![]()
![]()
![]()
BENT SØEGAARD
BENT SØEGAARD is Forest Geneticist at the Arboretum of the Royal Veterinary and Agricultural College, Horsholm, Denmark. He was assisted in the preparation of the final version of this chapter by H. D. Gerhold (United States). Other members of the drafting team were E. J. Schreiner (United States), E. Björkman (Sweden), R. T. Bingham (United States) and O. Fugalli (FAO).
Resistance of trees to insect attack is considered important by all thoughtful scientists, and there is an urgent need for increased work. Close co-operation between entomologists, physiologists and geneticists is required.
The best known example of resistance to insect attack is that of the white pine weevil, Pissodes strobi, and its relation to the eastern white pine, Pinus strobus; both the weevil and the pine have been much studied Other examples are the pine reproduction weevil, Cylindrocopturus eatoni, and its attack on Jeffrey and Coulter pines and the backcross hybrid, Pinus jeffreyi x (jeffreyi x coulteri), which has shown remarkable resistance. The Resin midge, Retinodiplosis spp., and the Dendroctonus bark beetles have also been studied on pines in California. In Douglas fir, Pseudotsuga taxifolia, studies on resistance to Chermes cooleyi have yielded interesting results.
Among broadleaved species mention is made of the resistance of Robinia pseudoacacia to the locust barer, Cyllene robiniae, and in poplars there are records of resistance to the Japanese bark beetle.
The term "resistance" refers to trees that are less damaged or less infested than others under comparable environmental conditions. The resistance may be due to the following reasons:
1. A tree may not be preferred for ovipositing, shelter or food.
2. A tree may possess a component of resistance called antibiosiss and may affect the biology of the insect.
3. A resistant tree may be tolerant and survive under levels of infestation that would kill or severely injure susceptible trees.
Variation in insect resistance is the basis for selection and breeding. Examples demonstrate the variation in resistance in white pine, larch, and Douglas fir. The basis of resistance has been investigated for the pine reproduction weevil and is attributed either to a cork layer surrounding the egg puncture in the cortex or some property of resin. Further examples deal with resin as a factor in resistance. Methods for investigating and improving resistance are based on selection. The comparison of direct or indirect selection suggests the use of an integrated selection system which combines the advantages of both methods. In testing Douglas fir for resistance to Chermes cooleyi, a glasshouse provided "laboratory" conditions. The mass production of the resistant material can be done by grafting, budding or the rooting of cuttings or by means of seed orchards.
Studies on the genetics and biology of Ips confusus and Dendroctonus pseudotsugae included work on the flight behavior.
Finally, attention is drawn to the importance of establishing international co-operation.
Increased interest in the resistance of plants to insect attack has become evident during the past 20 years, not only in agricultural but also in forest research. The use of insecticides and the subsequent development of resistance to many of them by insects has caused thoughtful entomologists and geneticists to realize that all possible means must be employed in insect control. Painter (1958) suggests that an important part of any well-rounded insect control project should be the search for sources of insect resistance, and the use of varieties and cultivars having such resistance in reducing the populations of insects and the damage done by them. Schreiner (1960) also calls attention to the importance of improving insect resistance in forest trees.
Very little has been done on breeding for insect resistance in forest trees either by entomologists or geneticists. A close co-operation with workers on problems of disease resistance is essential as the problems are essentially the same for both. Co-operation between forest entomologists, physiologists, and forest tree breeders is very necessary to understand more fully the causes of resistance.
Insects affect tree growth primarily by chewing plant tissues, sucking fluids from tissues, ovipositing and causing gall formation. They also influence growth by acting as vectors for some of the most important fungal, bacterial, and virus diseases of trees. Graham (1952) emphasized that no stage in the life cycle of a tree is free from insect attack. Seeds may be attacked before they are collected, and also later during storage. Seedlings and saplings are attacked and often killed by a wide variety of insects. In general, trees appear to acquire some resistance to insect attack when they have passed the sapling stage. However, as they approach maturity and their physiological activity declines, trees again become subject to widespread insect attack.
The white pine weevil Pissodes strobi, has long been recognized as the most important insect pest of eastern white pine, Pinus strobus, throughout its range. Injury to the tree results from killing of the leader during larval feeding. The first serious attack by weevil usually occurs when the trees are 5 years old and up to 1 meter (3 feet) in height; thereafter the attacks increase in severity until the trees are about 8 to 10 meters (25 to 30 feet) tall. Sullivan (1959) investigated the relationship between weevil activities and the climatic characters of the weevil habitat, and believes that 50 percent of incident light marks the dividing line between severe and moderate weevil damage. Wright and Gabriel (1959) state that there is enough proved resistance to form the basis of weevil-resistant cultivars with desirable growth characters, and that the production of Pinus strobus genetically resistant to the white pine weevil is worth attempting. Gerhold (1962) discusses what should be accomplished in the selection process and states that it is necessary to measure precisely the degree of weevil resistance and classify accurately the types of weevil resistance inherent in white pine progenies.
In an area with a high weevil population, scions of selected Pinus strobus, were top grafted onto young planted white pines, together with scions from heavily weeviled trees as controls. Grafts made with scions from a plantation of northern origin in southern Ontario were markedly less severely weeviled than grafts made with scions from weeviled local stock.
Trees with thinner bark and smaller leaders were found to be less susceptible by Barnes (1928), Prebble (1951), Kriebel (1954) and Sullivan (1957); while Fowler (1957) suggested that heavy resin flow in Pinus peuce may be responsible for the recovery of this species from attack. Experiments by Anderson and Fisher (1956, 1960) and observations by Barnes (1928) and MacAloney (1930) suggest that odors act both as attractants and repellents and are correlated with host specificity. The white pine weevil has also been reported on Pinus resinosa and P. banksiana (Heimburger, 1963). Even Picea abies is attacked (Hoist, 1955).
Investigations of the resistance of various pine species and their hybrids to attack and injury by the pine reproduction weevil (Cylindrocopturus eatoni) has been made at Berkeley and Placerville in California, U.S.A. It was demonstrated by Miller (1950) that Pinus coulteri and the backcross hybrid Pinus jeffreyi x (jeffreyi x coulteri) showed remarkable resistance to attack and injury by the reproduction weevil in caged tests under nursery conditions; by contrast, P. jeffreyi was severely attacked. Hall (1959) reported that the two experimental plantings where weevil damage occurred have demonstrated quite conclusively that the backcross hybrid Pinus jeffreyi x (jeffreyi x coulteri) has a high degree of resistance to attack and injury by the reproduction weevil under field planting conditions.
In Pinus ponderosa Austin et al. (1945) described the damage caused by the resin midge (Retinodiplosis sp.) and found that trees having glaucous shoots (covered with a waxy bloom) and glabrous shoots (dry and smooth) suffered little or no injury, while those having shoots that were viscid (sticky and covered with resin) suffered greater damage. It is suggested that trees susceptible to resin midge attack might be quickly identified for removal from the stand by observing the surface of the shoots.
In addition to breeding trees resistant to the pine reproduction weevil, the question of obtaining resistance to Dendroctonus bark beetles has received much study in California. Some promising leads have been obtained for identifying resistance and its cause.
Investigations in Denmark on the resistance of Pseudotsuga taxifolia to attacks of Chermes cooleyi involved the growth of clones under uniform conditions in a special glasshouse with great possibilities for infection by Chermes cooleyi. Nymphs were found on 32 clones which varied greatly in susceptibility judged by the wax deposits. One single clone remained unattached throughout the three years of the experiment (Petersen and SØEGAARD 1958).
Broadleaved tree species have also been examined for resistance to insect attacks. Thus, in his study of the locust borer (Cyllene robiniae), Hall (1942) states that areas were observed where certain races of Robinia pseudoacacia with excellent growth habit appeared to be highly resistant to injury by the borer. Two outstanding examples of this apparent resistance occur in "Shipmast" locust on Long Island and "Higbee" locust in southern Indiana. In the latter case, this apparent resistance has been tested over 4 years by the artificial application of over 2,000 locust borer larvae on a number of trees. Less than 20 of the larvae survived to the adult stage.
In Populus there is good evidence of clonal variation in resistance. Schreiner (1949) stated that the extremely wide variation in susceptibility among individual hybrids derived from the same parent trees is of great significance to forest tree breeding. Such differences were hardly expected for an insect like the Japanese beetle (Popillia japonica) which feeds on many species of plants. If the results obtained in 1947 are confirmed during the next few years they will justify intensive breeding for resistance to other forest insects.
These are only a few examples of different forest tree species which show some degree of resistance. However, it is now appropriate to define what is meant by the term "resistance." Resistant trees are those that are less damaged or less infested than others under comparable environmental conditions. According to Painter (1958), the resistance may be due to three basic components:
1. A tree may not be preferred for oviposition, shelter or food.2. A tree may possess a component of resistance called antibiosis, that is, the tree may affect the biology of the insect.
3. A resistant tree may be tolerant and survive levels of infestation that would kill or severely injure a susceptible tree.
Variation in resistance to insect attack has been observed in many of the examples given above. This variation is the basis for selection and breeding to obtain trees possessing a high yield capacity. Heimburger (1963) reports that the broad-crowned types of Pinus strobus, have greater survival value than the more slowly growing narrow-crowned ones, even when heavily attacked by Pissodes strobi. This provides an instance of Painter's group (3), "tolerant" trees. Heimburger op. cit., has also observed differences between provenances in resistance to weevil attack. When moving trees from south to north he found that trees planted to the north of their place of origin were more heavily and frequently weeviled than trees of local origin. Heimburger concludes that Pinus strobus, acceptable for planting in eastern North America, should be resistant both to blister rust, Cronartium ribicola, and to moderate weevil attack. Selection of narrow-crowned individuals with slender leaders that are less attractive to the weevil shows promise under conditions of moderate weevil attack. Screening for resistance to weevil attack is more difficult and time-consuming than screening for resistance to white pine blister rust. Therefore, it is easier first to screen seedlings in seed beds for their resistance to blister rust and then screen the survivors for resistance to the weevil.
Wright and Gabriel (1959) found variation in resistance to weeviling among geographic races of Pinus strobus, from a small provenance test in the Tug Hill area of New York. Pauley, Spurr, and Whitmore (1955) found that Pinus strobus, of three Ontario provenances suffered less weevil damage than did trees of Massachusetts, New Hampshire, or New York origin. They attributed the differences in weeviling to differences in growth rate.
Syrach Larsen (1956) demonstrated that Larix decidua is susceptible to a species of Chermes which is not found on Larix leptolepis. Callaham (1960) concluded from tests of species and hybrids that there was considerable heritable racial variation in resistance to weevils within Pinus ponderosa. Holst (1963) surveyed the susceptibility of species of Pinus in the series Lariciones1 and Australes to damage by the European pine shoot moth, Rhyacionia buoliana, and the Nantucket tip moth, Rhyacionia frustrana, and the species in each series have been rated for resistance to these moths. In the series Lariciones, Pinus nigra is most likely to provide highly resistant material. Pinus resinosa has significantly lower resistance than the other pines in the series. In the series Australes, longleaf pine (Pinus palustris) and slash pine (Pinus elliottii) are highly resistant. Species that have high resin yields are highly resistant to the two moths and within a species or series, high-resin yielders have shown more resistance than low-resin yielders.
1 The series Lariciones = series Sylvestres. The nomenclature followed here is that used by Wright (1962) in Chapter 3.
Syrach Larsen (1953) and Heitmüller (1954) have both reported the probable occurrence of single cases of complete resistance in Pseudotsuga taxifolia to Chermes attack. Petersen and Søegaard (1958) conclude that there are significant differences in the resistance of individual tees of Pseudotsuga taxifolia to attack by Chermes cooleyi. One single clone remained completely immune to attacks of the Chermes and in 1963 it was still not attacked even after being grafted under several different environmental conditions.
An important aspect of the resistance problem is that a satisfactory way of growing trees in one country or locality may be unsatisfactory elsewhere because of a change in the host/insect relationship. Holst (1963) gives examples of this in his study of the possibilities for breeding resistance of Pinus species to the Rhyacionia moths. The European pine shoot moth, Rhyacionia buoliana, was first discovered in Canada in 1915. It is now stabilized along the minimum winter isotherm of - 6.7°C (+ 20°F), although in Europe it extends into areas with very severe winters. The influence of the summer climate both on resin production and population increase is important both in Europe and in North America. Voute's (1946) study of European pine shoot moth on Pinus sylvestris in Holland over a 40-year period revealed severe damage after years with a hot July, usually associated with dry weather. In drought years the trees produce little resin and are thus unable to repel the shoot moth larvae. Neugebauer (1952) gave a somewhat similar explanation of his map which showed consistently high shoot moth populations in areas where the period May to July was dry. Areas with consistently high moth populations were those with low summer rainfall which causes low turgidity with consequent low assimilation, resin production and sap pressure.
Harris (1960) has shown for Pinus sylvestris in eastern England that in a warm summer the adults emerge, pair and lay eggs, the eggs hatch, and the larvae develop to the third instar in 5 weeks, whereas in a cool summer the same development takes about 14 weeks. Thus in a warm summer the third instar larvae attack the buds before they are well protected, whereas in a cool summer the larvae attack after the buds are protected by well-developed resin canals.
The problems of the cause or basis of the insect/host relationship always arise early in any study of resistance. Often the first question to be answered is whether trees surviving severe attacks really are resistant. To answer that question, it is necessary to test such "resistant" trees under controlled conditions in comparison with control trees. A further study of the basis of resistance requires a close examination of the various factors involved in the insect/host relationship. A few examples will serve to illustrate the problem.
Miller (1950) worked on the resistance of species of Pinus to the pine reproduction weevil, Cylindrocoptorus eatoni. He found that necrotic tissue develops around the egg puncture in the cortex, walling off this tissue. In trees which were killed, the small larvae penetrated the necrotic tissue and a cork layer and reached the cambium. In trees which survived, the larvae were unable to penetrate the corky tissue. Miller attributed resistance either to this layer which develops around the necrotic tissue, or to some property of the resin.
Salman and Bongberg (1942) and Keen and Salman (1942) developed a risk-rating system for classifying mature and overmature Pinus ponderosa and P. jeffreyi trees according to their relative susceptibility to Dendroctonus bark beetle attack. The trees so classified were used for one of the first intensive studies of the role of resin in resistance. Resin is considered a secondary plant product, and Fraenkel (1959) has stated, with reasonable evidence, that secondary plant products are very important in insect/host selection, susceptibility and resistance. Mergen et al. (1955) have found that certain characters of resin are under a high degree of genetic control.
Resin was first theoretically implicated in the resistance of Pinus ponderosa to Dendroctonus monticolae by Hopkins (1902), but for the next 25 years little was done to explore this possibility. Person (1931), working with Pinus ponderosa and Dendroctonus brevicomis, stated that the success of the first attacks on a tree depended on the ability of the beetle to overcome the flow of resin. Gordon (1932) found that certain constituents of resin were toxic to bark beetles, while Callaham (1955) showed that flow from a standard wound was prolonged in low-risk trees (those least likely to be killed by bark beetles). Callaham attributed failure of beetle attacks on low-risk trees to a continued flow of resin, but he was unable to associate total flow with susceptibility according to the risk classification. Smith (1961a) devised a technique for determining the toxic effect of saturated vapors of fresh pine resins to adult bark beetles. Using a limited number of Pinus species and three species of bark beetles, he suggested that Dendroctonus beetles could tolerate saturated resin vapors (that is, the volatile terpenes) of host pines but not of nonhost pines (Smith, 1961b).
Having noted the variation in resistance to insect attacks in a given species, the next problem is how to select in the forest trees with the desired qualities. A good method is described by Gerhold and Stroh (1963) of selecting Pinus strobus trees for resistance to Pissodes strobi. In their theoretical considerations concerning the employment of direct or indirect selection, Gerhold and Stroh state that indirect selection could be more effective than direct selection under certain conditions. If improvement in a character A is desired, it is usually better to select directly for A than for a character B which is correlated with A. But indirect selection could be more effective than direct selection if one or several of the following factors compensated for an incomplete correlation of B with A:
1. Less effort or cost is necessary in selecting for character B than for A.2. Less time per generation is needed for improving B than for A, as, for example, if selection is possible at an earlier age.
3. There is better precision in recognizing phenotype B than phenotype A as might be the case in the absence of insect infestations of sufficient intensity.
4. There is better correlation of progeny means with parental means for character B than for A.
Gerhold and Stroh emphasize the need to assess these relationships with due weight given to the importance of each in a particular breeding program and they continue: "The situation involving a decision between direct and indirect selection for insect resistance has, of course, been oversimplified. Insect resistance is often a complex trait, involving several mechanisms (Painter, 1958). In reality, then, it is likely that indirect selection would involve several traits rather than trait B only. Neglect of other traits would increase the risk of an insect adapting to the improvement in only a single trait. However, selection for several traits would undoubtedly multiply the work, and it might be questioned whether the genetic gain resulting from selecting n traits would be only 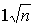 times the gain for single trait selection. Actually, the latter objection is not valid because selection for complex trait A indirectly involves all of its components, and selection acts upon a combination of these that is expressed as higher resistance."
times the gain for single trait selection. Actually, the latter objection is not valid because selection for complex trait A indirectly involves all of its components, and selection acts upon a combination of these that is expressed as higher resistance."
A technique for handling a large number of characters (traits) has been developed by Grafius (1956, 1959), Grafius and Kiesling (1958), and Grafius and Adams (1960), although not specifically for insect resistance. A model based on quantitative gene action was developed to predict the best combinations of parents that produce progenies which have means approaching an ideal for a large number of characters. Complex characters are broken up into their component parts to minimize epistatic interactions. In support of this novel approach, Grafius writes (personal communication of February 1963) "We now have data on... about 50 crosses in barley, and the data are in excellent agreement with expectations - this for a dozen different traits."
Gerhold and Stroh (1963) then discuss direct selection and state that two attributes of weevil attack make direct selection of Pinus strobus, for weevil resistance difficult. First, the percentage of trees damaged each year is normally quite low and only in exceptionally heavy infestations exceeds 50 percent. Secondly, the pattern of attack is at least close to being random; these characters make the precision of selection low. They suggest a procedure for preliminary selection of Pinus strobus, which is a variation of a technique suggested by Wright and Gabriel (1959). Stands in which selection is to be practiced should be fairly uniform, from 5 to 12 meters in height, and heavily weeviled for at least 6 years. First, tally the number of weevilings per tree by year in each of about 100 trees. Then determine the annual rate of weeviling and calculate the expected frequency distribution according to the number of weevilings per tree by expanding the binomial
(p1 + q1) (p2 + q2)......(pn + qn)
in which p = probability of attack and q = probability of escaping attack in each year. The actual and expected frequency distributions are compared and tested by chi square. Selection should be practiced only in stands in which there is a significantly larger than expected frequency of unweeviled trees and in which the ratio of actual to expected number of unweeviled trees is high. Direct selection of other white pines is discussed and found impossible in the northeast, with the possible exception of Pinus monticola.
In the discussion of indirect selection, Gerhold and Stroh (1963) state that several characters are correlated with weevil resistance in Pinus strobus, examples being trees as mentioned earlier with thinner bark and smaller leaders which were found to be less susceptible, and a decrease in susceptibility above an upper limit of leader diameter. The most promising characters emerging from their own investigations are bark thickness and depth to cortical resin ducts in the leader. From 20 to 49 percent of the variation in feeding in several plantations has been accounted for by measuring three characters in bark samples. It requires from 4 to 9 minutes per tree to obtain this data, depending on tree height. If chemical tests prove worthwhile, these would require about 20 minutes per tree in addition, plus the cost of analytical equipment and supplies. These figures are cited as evidence that indirect selection is several times as expensive as direct selection, which requires from 1 to 3 minutes per tree under a range of conditions.
Based on present limited knowledge, it seems that indirect selection would be too costly to be substituted for direct selection; but Gerhold and Stroh propose the employment of an integrated selection system in which the potential advantages of both selection methods complement each other.
In studying resistance to insect attacks empiricism has preceded laboratory tests. However, the investigation carried out by Petersen and SØEGAARD (1958) is very near to laboratory conditions. They brought together grafts of selected trees in a special glasshouse where it was possible to give the grafts uniform and the pest excellent conditions. Such "laboratory" tests have a great advantage where vegetative propagation can be employed, for it is then possible to combine laboratory investigations with field tests.
The assessment of resistance for trees propagated through seed had been discussed by Wright and Gabriel (1959) and they propose the following program to the end of the second generation for resistance to white pine weevil, Pissodes strobi. Phenotypic selection of 500 apparently resistant individuals; determination of the general combining ability of all 500 parents by 1-parent progeny tests; determination of specific combining ability of the best 32 parents by diallel, 2-parent progeny tests; phenotypic selection in the F1 test plantations; determination of the general combining ability of the 500 best F1 seedlings by 1-parent progeny tests; and determination of the specific combining ability of the 32 best parents by diallel 2-parent progeny tests.
The search for sources of resistance has preceded the study of inheritance of resistance and the closely related breeding work. In the breeding program on resistance of pines to the pine reproduction weevil, Smith (1960) has reported that eleven resistant Pinus species, varieties or hybrids had been discovered out of 30 which had been observed during the preceding 15-year period that the host relationship of the weevil had been studied. He found evidence suggesting that the female parent is more influential than the male in the transmission of resistance to the progeny. Research findings to date show that the possibilities for hybridizing pines to produce trees resistant to weevils are considerable.
Another example from Pinus concerns the breeding work done by Smith and Eaton (1963) which has produced 2 hybrids of particular interest, namely P. jeffreyi x ponderosa and P. (jeffreyi x coulteri). Each is a cross of a nonhost and host species with respect to both Dendroctonus brevicomis and D. monticolae. Resin vapors of these hybrids usually were toxic to D. brevicomis but not to D. monticolae.
When the breeder has produced trees resistant to attack by insects, he then becomes interested in the possibilities of reproducing the material on a large scale for use in the forest. If the species concerned is propagated vegetatively (as is the case with Populus and Cryptomeria japonica), the material obtained can be used through grafting, budding or the rooting of cuttings. If the species is propagated by seed, one method for mass production of the resistant cultivar is the establishment of seed orchards as described in chapters 7 and 10.
Smith and Eaton (1963) report that a large-scale program to produce weevil-resistant stock for planting in northern California was begun by the United States Forest Service 5 years ago (see Libby, 1958). The breeding program uses P. jeffreyi x coulteri F1 hybrids, and the natural hybrid. Current work is designed to screen a large number of trees of Pinus jeffreyi for their compatibility with Pinus coulteri. Hybrid-producing trees of Pinus jeffreyi are being grafted into an orchard for subsequent controlled pollinations. The annual production of weevil-resistant hybrids is now tens of thousands but the goal is several million.
It is recognized in resistance studies that the insect host relationship is of great importance as is also the study of the two separate factors involved - the tree and the insect. Observations and studies of the trees have been recorded in several of the examples mentioned above and Smith and Eaton (1963) stress the importance of the insect and have themselves begun work on the variable capacity of bark beetles to attack pines "...which could be equally as important as the variable capacity of the tree to resist the beetles."
Gara (1963) studied Ips confusus and found that the flight behavior, observed from take-off to landing, was influenced by wind velocity and direction, intensity of attractants, and the ground cover. The response to attractants occurred immediately, indicating that flight exercise is not a prerequisite to response in this species. The ability to respond to attractants was found to vary from beetle to beetle, with sex, within a brood, and from population to population. He concluded that the differences in response among individual insects and broods are due to their physiological condition.
Vité et al. (1963) report from their studies that a volatile substance responsible for the mass attraction of Ips confusus to trapping devices was obtained from the hindgut of mature male beetles, from their frass, and by condensation from air drawn from a box containing beetle-infested sections of logs. The attractant was found to be released only after emerging male beetles had fed for several hours on new host material and the gut, empty during flight, had been filled. In some of the laboratory experiments, the attractive substance seemed to act as an arrestant rather than an attractant.
Rudinsky (1963) has studied the response of Dendroctonus pseudotsugae to volatile materials emanating from infested logs of Pseudotsuga taxifolia. Immediately after the beetle's emergence, dispersal flight occurred at random and then became a concentrated flight in response to an attraction factor caused by the invasion of the first bark beetles. The attractive substance has been concentrated in the laboratory, and when it was released in the field it caused the flight population to respond. This suggests its possible use in the control and prevention of outbreaks of Dendroctonus pseudotsugae.
International co-operation has been established in the study of diseases. In Denmark, Finland, Norway and Sweden, co-operation between forest entomologists has begun, although on a rather limited scale. The small number of eight investigators engaged in forest entomology in the Nordic countries makes such co-operation all the more valuable for each individual country and sometimes permits the mounting of a research project on a much larger scale than would have been possible in any one of the countries by itself. Moreover, this co-operation permits a greater degree of specialization, the investigation of much larger geographical and climatic areas and often results in a more representative sampling of the variable to be investigated.
If co-operation such as this can be enlarged - first, to make it possible for the group mentioned to cooperate with similar groups established in other geographic regions of reasonable size and improvement; and second to permit the single group within the respective region to make arrangements for close co-operation with the forest geneticist and tree breeders of the region - then highly profitable international co-operation will he the result.
ANDERSON, J. M. & FISHER, K. C. 1956. Repellency and host specificity in the white pine weevil. Physiol. Zool., 29: 314-324.
ANDERSON, J. M. & FISHER, K. C. 1960. The response of the white pine weevil to naturally occurring repellents. Canada J. Zool., 38: 547-564.
AUSTIN, L., YUILL, J. S. & BRECHEEN K. G. 1945. Use of shoot characters in selecting Ponderosa pines resistant to resin midge. Ecol., 26 (3): 288-296.
BARNES, T. C. 1928. A biological study of the white pine weevil with special reference, to anatomy, flight, phenology, parasitism, behaviour, and injuries to young trees. Unpub. doctoral dies., Harvard Univ., Cambridge, Mass.
CALLAHAM, R. Z. 1955. Oleoresin production in the resistance of Ponderosa pine to bark beetles. Ph. D. thesis, Univ. of California, Berkeley, Calif. 120 p.
CALLAHAM, R. Z. 1960. Observations on pine susceptibility to weevils. U.S. Forest Serv. Pac. Southwest Forest and Range Expt. Sta. Tech. Paper 51. 12 p.
FOWLER, D. P. 1967. Discussion of tree breeding at Maple, Ontario. Proc. 5th Northeast. For. Tree Impr. Conf., p. 26.
FRAENKEL, G. S. 1959. The raison d'être of secondary plant substances. Science, 129 (3361): 1466-1470.
GARA, R. I. 1963. Studies on the fight behaviour of Ips confusus (Lec.) (Coleoptera: Scolytidae) in response to attractive material. Contrib. Boyce Thompson Inst., Jan-March.
GERHOLD, H. D. 1962. Testing white pines for weevil resistance. Proc. 9th Northeast. For. Tree Impr. Conf., 1961: 44-50.
GERHOLD, H. D. & STROH, R. C. 1963. Integrated selection for white pine weevil resistance and its components. FAO/FORGEN 63-6b/1.
GORDON, A. 1932. Tree injection experiments in white fir and ecological studies of insects attacking yellow pine. U.S. Dept. Agric., Bur. Ent., Forest Insect Lab., Berkeley, Calif. Unpub. Prog. 10 p.
GRAFIUS, J. E. 1956. Components of yield in oats: a geometrical interpretation. Agron. Jour., 48: 419-423.
GRAFIUS, J. E. 1959. Heterosis in barley. Agron. J., 51: 551-554.
GRAFIUS, J. E. & KIESLING, R. C. 1958. Vector representation of biologic fields of force. Agron. J., 50: 757-760.
GRAFIUS J. E. & ADAMS, M. W. 1960. Eugenics in crops. Agron. J., 52: 519-523.
GRAHAM, S. A. 1952. Forest entomology. New York, McGraw-Hill.
HALL, R. C. 1942. Control of the locust borer. U.S. Dept. Agric., Circ. 626. 19 p.
HALL, R. C. 1959. Field tests of the resistance of hybrid pinees to the pine reproduction weevil. U.S. Forest Serv., Pac. Southwest For. and Range Expt. Sta., Tech. Paper 33. 21 p.
HARRIS, P. 1960. Natural mortality of the pine shoot moth, Rhyacionia buoliana (Schiff.) (Lepidoptera: Olethreutidae), in England. Can. J. Zool., 38: 755-768.
HEIMBURGER, C. 1963. The breeding of white pine for resistance to weevil. FAO/FORGEN, 63-6b/5.
HEITMÜLLER, H. H. 1954. Beobachtungen über individuelle. Resistenz gegen Gilletteella cooleyi Gill. an Douglasie. Z. Forstgenet., 3: 99-100.
HOLST, M. J. 1955. An observation of weeril damage in Norway spruce. For. Res. Div., Tech. Note No. 4, Ottawa.
HOLT M. J. 1963. Breeding resistance in pines to Rhyacionia moths. Canada; Dept. of Forestry, Forest. Res. Branch Contr. (In Press)
HOPKINS, A. D. 1902. Insect enemies of the pine in the Black Hills forest reserve. U.S. Div. Ent. Bull. 32. 24 p.
KEEN, F. P. & SALMAN, K. A. 1942. Progress in pine beetle control through tree selection. J. For., 40 (11): 854-858.
KRIEBEL, H. B. 1954. Bark thickness as a factor in resistance to white pine weevil injury. J. For., 54 (2): 133-34.
LIBBY, W. J. 1958. The backcross hybrid Jeffrey x (Jeffrey x Coulter) pine. J. For., 56: 840-842.
MACALONEY H. J. 1930. The white pine weevil (Pissodes strobi Peck). Its biology and control. N. Y. State Coll. of Forestry, Tech. Publ. 28. 87 p.
MERGEN, F., HOEKSTRA P. E. & ECHOLS, R. M. 1955. Genetic control of oleoresin yield and viscosity in slash pine. For. Sci., 1 (1): 19-30.
MILLER, J. M. 1950. Resistance of pine hybrids to the pine reproduction weevil. U.S. Forest Serv., Calif. Forest and Range Expt. Sta. Res. Note 68. 17 p.
NEUGEBAUER, W. 1952. Die Bekämpfung des Kieferntriebwicklers. Forstarchiv, 23: 159-165.
PAINTER, R. H. 1958. Resistance of plants to insects. Ann. Rev. Entomology, 3: 267-290.
PAULEY, S. S., SPURR, S. H. & WHITMORE, F.W. 1955. Seed source trials of eastern white pine. For. Sci., 1: 244-256.
PERSON, H. L. 1931. Theory in explanation of the selection of certain trees by the western pine beetle. J. For., 29 (5): 696-699.
PETERSEN, B. B. & SØEGAARD, B. 1958. Studies on resistance to attacks of Chermes cooleyi (Gill.) on Pseudotsuga taxifolia (Poir.) Britt. Forstl. Foresøgsv. Danm., 25: 37-45.
PREBBLE, M. L. 1951. Summary of investigations of attack by the white pine weevil in plots at the Petawawa Forest Exp. Sta. 1936-1948. For. Insect Lab. Sault St. Marie, Ontario.
RUDINSKY, J. A. 1963. Response of Dendroctonus pseudotsugae, Hopkins to volatile attractants. Contrib. Boyce Thompson Inst., Jan-March.
SALMAN, K. A. & BONGBERG, J. W. 1942. Logging high-risk trees to control insects in pine stands in northeastern California. J. For., 40 (7): 533-539.
SCHREINER, E. J. 1949. Poplars can be bred to order. U.S. Dept. Agric. Yearbook, p. 153-157.
SCHREINER, E. J. 1960. Objectives of pest-resistance improvement in forest trees and their possible attainment. Proc. 5th World For. Congr., Seattle, U.S.A., 2: 721-727.
SMITH R. H. 1960. Resistance of pines to the pine reproduction weevil Cylindrocepturus eatoni. J. Econ. Ent., 53: 144-148.
SMITH, R. H. 1961. Techniques for determining the toxicity of pine resin vapors to Dendroctonus brevicomis and D. jeffreyi. J. Econ. Ent., 54 (2): 359-365.
SMITH, R. H. 1961b. The fumigant toxicity of three pine resins to Dendroctonus brevicomis and D. jeffreyi J. Econ. Ent., 54 (2): 365-369.
SMITH, R. H. & EATON, C. B. 1963. Studies on resistance of pines to insects. FAO/FORGEN, 63-66/2.
SULLIVAN, C. R. 1957. A biological study of the white pine weevil, Pissodes strobi Peck, with special reference to the EFFECT of physical factors on its activity and behavior. Ph. D. thesis, McGill Univ.
SULLIVAN, C. R. 1961. The effect of weather and the physical attributes of white pine leaders on the behaviour and survival of the white pine weevil, Pissodes strobi Peck. in mixed stands. Canad. Ent., 53: 721-724.
SYRACH LARSEN, C. 1953. Studies of diseases in clones of forest trees. Hereditas, 39: 179-192.
SYRACH LARSEN, C. 1956. Genetics in silviculture. Edinburgh, Oliver and Boyd.
VITÉ, J. P., GARA, R. I. & KLIEFORTH, R. A. 1963. Collection and bioassay of a volatile fraction attractive to Ips confusus (Lec.) (Coleoptera: Scolytidea). Contrib. Boyce Thompson Inst., Jan-March.
VOUTE, A. D. 1946. De oorzaak van het optreden van gradaties van de dennenlotrups (Evetria buoliana [Schiff.]). en de mogelijkheid deze te voorkomen [Cause of the occurrence of the pine-shoot moth Evetria buoliana and the possibility of controlling it]. Ned. Boschb. Tijdschr., 18:
104-111, 128-135.
WRIGHT, J. W. & GABRIEL, W. J. 1959. Possibilities of breeding weevil-resistance white pine strains. Northeast. For. Expt. Sta. Paper 115. 35 p.
![]()
![]()
![]()