![]()
![]()
![]()
J.J. HAVEL
J.J. HAVEL is the Principal of the Forestry School, Bulolo, New Guinea. W. Finlayson assisted in reading and correcting the manuscript.
IT IS SAID, with some truth, that the tropical rainforest is the botanist's dream but the forester's nightmare. The excessive richness of species hinders the assessment of resources, the initial utilization of the forest and its subsequent regeneration and management. A mere plot assessment becomes a major taxonomic exercise, yet volumetric assessment without identification gives no indication of the likely harvest. The harvesting and processing of such a varied crop, and especially its marketing, present almost unsurmountable difficulties, with the result that usually only a minor proportion of the forest is logged. The regeneration of the logged-over forest is often a puzzle, seemingly without solution, which frequently drives the foresters to clear-fell and replant with exotics.
The often unsatisfactory state of forestry in tropical countries is at least partially attributable to the inability of foresters to identify reliably even a minor proportion of all the species. Most professional foresters are trained in temperate countries and their first encounter with tropical rainforests is after their graduation. What is more, the countries in which they work generally lack complete floras. The use of indigenous names and the employment of local inhabitants as tree spotters are not without difficulties. In many areas, particularly in New Guinea, the fragmentation of languages is such that the matching of tribal and scientific names is as difficult as the field identification itself. Furthermore, the scientific and the tribal concepts do not always coincide. In the case of trees of economic importance to the local population names exist for barely distinguishable varieties, whereas species and even genera without economic value are bundled together under one common name, such as the Malayan term medang.
However, the primary cause of difficulty is the sheer magnitude of the problem.
In contrast to temperate or subtropical forests, the tropical rainforest is extremely rich in both species and families. Except under special climatic and edaphic conditions, no species is of overwhelming importance, ecologically or economically. The number of tree species in Malaya has been estimated by Menon (1961) at 2,000 to 2,500; the estimates for New Guinea are even higher. They belong to a very wide range of families, including those which in temperate countries are composed largely of herbs; for example, datiscaceae.
As a result, no single feature, other than flower structure, is sufficiently varied to provide adequate means of identification. The situation has been examined in Malaya by Menon, who concluded that the woody tissues are too simple to carry the identification as far as the species, and that in many cases it is even impossible to separate related genera,. A key to the tree genera of Indonesia, based on vegetative organs only, has been developed by Endert (1956), but it necessitates such detailed morphological and anatomical examination of twigs that its use is restricted to professional botanists.
Although the use of bark in tree identification has been considered recently by a number of botanists (Wood 1952, Wyatt Smith 1954, Whitmore 1963) it cannot by itself provide exact and reliable identification for all species.
It is obvious that a forester, for whom taxonomic botany is only one of many spheres of work, has no hope of mastering all the species of the rainforest with which he is concerned; yet, paradoxically, foresters seem to prefer to deal with individual species. There are many reasons for this. Many features used by foresters in identification, such as the outer bark or the form of a tree, are frequently characteristic of individual species. It is often easier to recognize a particular species than to grasp the concept of a whole genus. Two species of the same genus frequently differ markedly in their economic value.
In contrast to this, the botanists favor the study of families for equally valid reasons. In many of the common families, such as the Lauraceae and the Meliaceae, the classification and nomenclature are at present far from complete, and valid identification of species is difficult, if not impossible.
Foresters and botanists also seem to be divided on the means of identification. Foresters generally avoid the use of fertile material, on the grounds that flowers and fruits of the large forest trees are virtually inaccessible, and in many cases, only available seasonally. They prefer to use the overall appearance of the trees and the structure of their bark and wood, which are readily accessible from the ground, supplemented by the characteristics of fallen leaves. Botanists, on the other hand, tend to depend on features which can be seen in fertile herbarium specimens, that is, leaves, flowers and fruits. Frequently they cannot recognize in the field a species which they have repeatedly handled in the herbarium. It is not for me to decide who is wrong - they both are. Fallen fruits and flowers are far more useful, frequent and reliable features than the foresters are prepared to admit. On the other hand, once a reliable correlation has been developed between fertile material and the bark and wood, the objections of the botanists to the use of vegetative features lose their force.
However, the fact remains that scientific classification and nomenclature are based on fertile specimens, and that positive initial identification is only possible on this basis. Putting it simply - the botanists pipe the tune to which the foresters dance. After the initial positive identification, subsequent identifications based on a correlation between reproductive and vegetative features are not only possible, but are far more efficient in routine practice.
One reason why greater use is not made of such correlations is that they are not always simple. It is possible for genera closely related on the basis of flower structure to be widely divergent in wood structure or, on the other hand, for the same anatomic feature to be present on otherwise quite unrelated genera (Menon 1961). This, after all, is not foreign to the taxonomists: identification and classification are not, and indeed cannot be, based on a single feature alone. Rather they are based on a combination of features of varying complexity.
The less common and the more striking the features are, the fewer of them are needed. The combination of features becomes increasingly more important but also more difficult to grasp, as one moves from species to genus and to family, the chief reason being that one moves toward wider, more general and therefore less clearly defined concepts.
We thus arrive at the crux of the problem: the individual species comprising the forests of tropical countries are too numerous to be dealt with as distinct, unrelated entities. There is, therefore, a need for generalization through the concepts of genus and family. Such a generalization facilitates the retention of acquired knowledge by providing a framework into which the data can be meaningfully filled. It also reduces the number of facts that need to be memorized, because many features are common to all members of a family. Finally, it provides a way to future expansion of knowledge, in that trees not previously encountered can be assigned to a family and then identified, by means of a key to the family, at generic or specific level. However, before this can be done, the difficult and often elusive concept of a family must be developed.
To aim solely for such an advanced stage may be too much of a gamble. If, however, the ecologically or economically important species are employed in the development of the concept, the ability to recognize these is gained even though the more advanced stage of concept formation may prove too difficult.
The formation of the genus and family concepts is a difficult task, which must be tackled thoroughly. Two principles are suggested. Firstly, the trees should be treated as a whole, not merely as pieces of wood covered by bark, or twigs glued to a herbarium sheet. An excellent example of the use of a wide range of features in tree identification is given by Wyatt Smith (1954). Secondly, teaching or learning the concepts should follow the discoveries of educational psychology in this field.
There are two ways of developing the concept of a family; of these, the formal, deductive approach is more commonly used, especially in textbooks. It consists of enumerating the features of the family, splitting it by means of a key, and describing its component genera and species. Seemingly, it is simple and orderly. Its main weakness is that few men can visualize the family from the definition, because the defining features are rarely clearly cut. It is not unusual for the genera of a family to have alternate, opposite and whorled leaves, or four, five or six petals. This is an incomplete reasoning process, perhaps suitable for the specialist but quite out of place at a lower level.
The alternative approach is inductive. It consists of studying a group of trees known to belong to a particular family and arriving by observation and reasoning at the concept of the family. It is a natural approach, possible even where precise description of features is difficult. It can be and is being done, by people lacking formal education; for example, few New Guinea students fail to place Viticipremnanovae pomeraniae next to Vitex cofassus, despite the fact that the two differ in having palmately compound and simple leaves respectively. Many recognize the introduced Tectona as a "brother" of the indigenous Gmelina; some will go as far as placing the four genera mentioned here, within a "clan," though they have never heard about the family Verbenaceae.
The inductive development of a concept can be broken into five component steps:
(a) observation of the various objects;(b) formation of hypothesis about their common elements;
(c) testing of the hypothesis in specific instances;
(d) selection of a general category which fits all the objects;
(e) giving the category a name.
The same sequence needs to be followed in the study of taxonomy. The concept of the family will be arrived at more rapidly if a teacher can assist the student but, given adequate basic training and a suitable reference book, the task can be undertaken by the student alone. Some comments will now be made on the implementation of the above reasoning.
OBSERVATION OF TREES KNOWN TO BELONG TO A FAMILY
Instructions on how to observe trees may seem superfluous but they certainly are not, A student must be taught to recognize relevant features; for instance, many Leguminosae have spectacular flowers, whose color immediately catches the eye, yet the colors are irrelevant; what matters is the arrangement of the floral parts, the type of fruit and the nature of the wood.
The numerous features observable on each tree may be divided into three main categories:
(a) main features useful in recognizing the tree as a member of a particular family or genus;
(b) striking features useful in the identification of the particular species;
(c) commonplace features useful only in the checking of the preliminary identification.
Such a fragmented observation must, however, be balanced by perception of the whole tree as an entity, because the whole is more than the mere sum of its component parts. It is possible to recognize members of the more clear-cut families, such as Myristicaceae, at a glance without mentally checking the characteristics of the family. The feature on which such recognition is based - the slant of the branches, the arrangement of the leaves, the nature of the bark - are difficult to describe. For this reason, recognition-by-glance is only developed by experience, through gradual reduction of the number of clues necessary for positive identification and through shift in emphasis from detailed, clear-cut feature to overall impression.
A good herbarium and wood collection, and ready access to the forest are, of course, indispensable for such study. Access to the forest presents less difficulty for lower level training institutions than for universities.
The herbarium material, wood samples and trees should be seen in close spatial and temporal proximity. If an afternoon is set aside for the study of a particular family, the members of other families should be avoided, no matter how conveniently nearby they may occur. The clearness of concept achieved this way is well worth the extra effort.
Because speedy identification is only achieved by repeated observations, the establishment of a botanical walk with trees bearing name tags, is also well worth while
FORMATION OF THE HYPOTHESIS
Here, even more than in the first step, the teacher can be of great help in drawing attention to the common features of the family. Before this can be done, the students will need to know basic taxonomic terminology. The terms employed should cover the whole tree and not merely its reproductive features, but excessive precision and detail are not warranted. Features not readily seen with the naked eye or ten-magnification lens are best omitted, as they are of little use in field identification. It is worth noting that many of the terms used in taxonomy and wood anatomy are in themselves already fairly complex concepts, describing the interrelation of the component parts, such as epigyny in flowers and confluent parenchyma in wood. As such, they are useful steps in the development of the family concept.
Where a textbook is being used, parallelism of descriptions and illustrations help to bring out the similarities. As the concept formation involves the combination of many specific experiences into one generalized abstraction, the use of such aids as floral formulas or diagrams speeds up the process.
During the study of the family, and in subsequent encounters with it, there should be a continual revision of the concept, in which the earlier perceptions and the new observations are fused. The resulting new concept will either be wider or more precise than the earlier one, depending on the selection of the earlier examples. In the light of his wider experience, the teacher is justified in avoiding examples which lead to an unnecessarily wide and general concept. In most families the study of timber trees of a particular country will lead to a narrower, more precise concept than the study of the herbs and trees of the whole world.
The family concept is easier to develop in some families than others. Small compact families, such as the Myristicaceae, are much easier to group than the larger variable families, such as the Euphorbiaceae.
TESTING THE HYPOTHESIS
This is an important step because it not only makes possible a check of the effectiveness of past study, but encourages the active participation of the students. Two possible approaches suggest themselves: either the omission of an important species from the initial concept formation, or the use of a minor, relatively unimportant species. In either case, the species should be casually introduced so that the learners may attempt to place it without assistance. The rejection of features which are only of specific value helps to clarify the family concept.
DEFINITION OF THE FAMILY AND ITS NAMING
Definition of the family may be attempted by the students, but usually the assistance of the teacher will be needed. At this stage, the name of the family can be attached to the concept. This may seem a back-to-front approach, yet without the prior development of the concept, the name is merely a hollow empty thing.
Earlier, a mention was made of the use of the family concept as an aid to memory, in that the facts are made more meaningful. It is worth considering whether the same principle applies to larger units of classification, such as the orders. In most countries, the herbaria use the Engler and Prantl system, which is useful in that it groups families together according to their similarities. For instance, the sequence of dicotyledonous families progresses from the absence of perianth, through perianth of one series, to perianth of two spirally arranged series, continuing on to cyclic arrangements of two series, with the segments of the inner series distinct and finally to segments of inner series fused. Such an arrangement provides a further and larger framework for the orderly arrangement of data. There are also parallel but less constant series from apocarpy to syncarpy, and from hypogyny through perigyny to epigyny. In view of the vast volume of data that needs to be remembered, any approach which facilitates retention is worth trying.
FINAL STAGES
During the development of the family concept, it is desirable to facilitate transfer of learning to the second stage, when the student will be working alone. Thus the terminology used initially should, as far as possible, follow the reference works on which the student will ultimately depend. In the case of the Malaysian region, these are unquestionably the Flora Malesiana and the Commonwealth Scientific and Industrial Research Organization (CSIRO) card key to the timbers of the southwest Pacific area. The chief problem in this respect is the wide range of technical terms used by professional botanists.
As a final step, students should be taught a suitable approach to the identification of a species previously not seen. One such approach, used by the writer, is as follows: If the unknown species can, on the basis of initial impression, be placed within a genus or a family, the "hunch" should be immediately checked by comparing the features of the unknown species with the known characteristics of the genus or family. If there is no immediate "hunch," the features of the unknown species are written down and compared with the known characteristics of the various families. Often the mere enumeration leads to a breakthrough. If an approximate identification is then possible, a thorough check should again be made. Where a herbarium is available, no better check can be made than a direct comparison with the actual specimens; elsewhere, reference books must suffice. An integral part of the second stage is the use of taxonomic keys. As this is a subject adequately covered in most textbooks, it will not be dealt with here.
In conclusion, it may well be questioned whether the suggested approach is feasible. The writer has found that some interested and intelligent trainees with lower high school education, after studying 150 of the more important New Guinea species over a period of one year, could place many previously unseen species in the correct family or genus. Other trainees could only recognize those species which they were actually taught, and could not grasp the family concept. Even where identification is given, it is not always correct and the wending of a specimen for confirmation by a herbarium is encouraged. The main point is that a foundation has been laid and a framework has been established, thus making further progress possible. The higher the education, the better the results which can be anticipated.
ENDERT, F.H. 1956. Key to the tree genera of Indonesia. English translation by R. D. Hoogland. Canberra, Commonwealth Scientific and Industrial Research Organization.
KRECH, D. & CRUTCHFIELD, R. S. 1959. Elements of psychology. New York, Knopf.
LAWRENCE, G.H.M. 1951. Taxonomy of vascular plants. New York, Macmillan.
MENON, P.K.B. 1961. Taxonomic value of wood anatomy seen through Malayan woods. Malayan Forester, 24: 290-301.
MERRILL, E.D. 1954. Plant life of the Pacific. New York, Macmillan.
WHITMORE, T.C. 1963. Bart, morphology as an aid to forest recognition and taxonomy of Dipterocarpaceae. Flora Malesiana Bulletin, 18: 1017-1019.
WOOD, G.H.S. 1952. Bark as means of tree identification. Journal of the Oxford University Forest Society. Ser. 3, 6: 15-27.
WYATT SMITH, J. 1954. Suggested definitions of field characteristics. Malayan Forester, 17: 170-183.
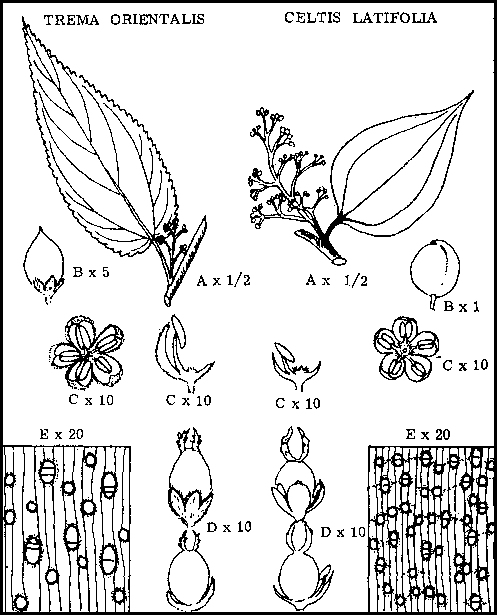
Celtis and Trema, together with two other unimportant genera in New Guinea resembling Trema, belong to the family Ulmaceae. The characteristics of the family, as seen in Celtis and Trema, are as follows:
Leaves are simple, alternate, with entire or toothed margin.
Trees are monoecious, flowers mostly unisexual, consisting of a 4- to 6-lobed peranth, with equal number of stamens in male flowers, and with a single one-celled ovary with two styles in female flowers.
Fruit is mostly a small drupe, with thin outer flesh and inner hard shell.
Timbers have narrow rays, small- to medium-sized pores in short multiples and parenchyma surrounding pores.
Note that, in the following more detailed descriptions of Celtis latifolia and Trema orientalis broken underlining refers to family features, continuous underlining to important specific features.
CELTIS LATIFOLIA PLANCH.
|
Habit |
A large tree up to 140 ft (42 in) tall and 3 ft (1 in) in diameter. Bole buttressed, long, slender, tapering, crown of medium width. |
|
Bark |
½ in. (1.27 cm) thick. Outer bark grey, smooth except for pustules and some raised horizontal, ridges. Middle bark green, inner bark cream to straw, with prominent red-brown "salt and pepper" pattern, hard non-fibrous. Cut darkens rapidly, |
|
Wood |
Cream to straw colored, moderately hard and heavy, rays narrow, pores of medium size, moderately numerous in short radial multiples, parenchyma surrounding pores. Winglike and confluent. |
|
Leaves |
Simple, alternate, ovate, often assymetrical, 1 ½ x 3 in (3.8 x 7.62 cm), tip acuminate, base round, margin entire, venation multicostate, ternate, of medium thickness, leathery, dark green glossy above, light green glabrous beneath. |
|
Flowers |
In axillary or terminal panicles, small, yellow, green, faint smell. The flowers either unisexual and monoecious, or seemingly bi-sexual. Male flowers consist of 5 perianth segments, with one stamen opposite each segment. Female flowers again with 5 perianth segments, with a large ovoid ovary with 2 styles and with weakly developed stamens. |
|
Fruit |
A globose drupe with pointed tip and thin flesh, shiny blue-black when mature. |
|
Occurrence |
In lowland rainforest throughout the Territory, associated with the various broad leaf species. |
|
Other species |
There are six species of Celtis in the Territory, differing mainly in size and shape of leaves. In two of them, the venation is not markedly ternate. The identification in the bush is based on the "salt and pepper" pattern in the bark, on wood structure and, in most cases, on ternate venation |
TREMA ORIENTALIS (Linn) Bl.
|
Habit |
Small- to medium-size tree, up to 70 ft (21 in) tall and 1-5 in (38 cm) diameter. Bole spurred, straight; crown of medium width to broad, foliage thin. |
|
Bark |
¼ - 1/3 in (0.63 - 0.84 cm) thick; outer bark grey, smooth, inner bark greenish cream, very fibrous, used as bush rope. |
|
Wood |
Soft and light, straw colored, coarse grained, pores of medium size in short chains, rays narrow, wavy, parenchyma surrounding pores. |
|
Leaves |
Simple, alternate, districhous, ovate, with acuminate tip, cordate base and serrate margin, venation unicostate, but two lowest veins strongly developed, of medium thickness, green, weakly hairy above, silvery, densely hairy beneath. |
|
Flowers |
In small axillary panicles, flowers minute yellow green, unisexual, trees monoecious. Male flowers consist of 5 lobed perianth with a stamen opposite each lobe. Female flowers consist of 5 lobed perianth with a large ovary which bears two styles. |
|
Fruit |
Small - 1/8 in (0.32 cm) diameter, ovoid, black drupe with thin outer flesh and inner hard shell, with persistent perianth lobes. |
|
Other species |
There are two other species of Trema in the species Territory, differing mainly in the hairiness of leaves. |
|
Occurrence |
The three species of Trema occur throughout New Guinea, in lowlands and mountains up to about 5,000 ft, (1,500 in), very common on old garden sites. |
Trema orientalis - Celtis latifolia
![]()
![]()
![]()