![]()
![]()
![]()
Solar radiation is the source of heat for all water bodies. The amount and angle of incidence of sunlight would decide the energy entering the water body. Distribution of heat within the water by conduction is negligible because of very low heat conductivity of water. Much of heat mixing takes place by convection aided by wind action. Heat is lost due to evaporation and also by direct exchange to air and substratum.
Largely temperature variation in water bodies depend on their geographical location, latitude and altitude. In the tropics surface temperature of shallow water bodies, reach such as ponds, tanks and lagoons and backwaters can occasionaly near 40°C or above in peak summer and can cause mortality of fishes especially if the water body is less than a metre in depth; more often the summer daily maximums are well below this temperature limit; near 30 – 35°C. As indicated elsewhere most warm-water fishes tolerate and fare well in this range. In the temperate regions in which we are less concerned, the daily maxium temperatures rarely reach over 25°C, but in the high altitude tropics (e.g. in Kenya, Tanzania, Malawi, Zimbabwe in Africa) where we have introduced exotic fishes such as the trouts, the thermal conditions are similar to those of temperate region. Here summer daily maximum temperatures are more critical and introduction of coldwater fishes such as salmonids should not be made in sites where the summer daily maximum temperature would go over 20°C.
In artificial ponds where the depth is usually of 1 – 2 metres there will be only a minor difference between the surface and bottom water. This difference would increase with increase in depth of the water bodies. Even in the tropical shallow ponds and tanks the surface water temperature can be much higher in hot summer afternoons and it is likely that fishes congregating in the cooler and deeper portions of the pond survive such a critical condition. Also it must be noted that even the daily amplitude of temperature change can be high in certain tropical areas, the upper limit reaching near 40°C and the lower near 20°C in the same day. In such a condition it is questioned if fish mortality is due to even low temperature (a rare phenomenon in the tropical plains), for the fish acclimates itself to the highest temperature of the day and a sudden lowering of temperature could take the fish to its lower lethal point (Fry, 1971). We have suspected this to happen in Vaigai Resevoir (S. India). It is, however, to be stressed that the commonest cause of thermal depth in the tropics is the warming of water. A point made elsewhere should again be noted here, that quite often fish mortality observed is not due to a single factor such as temperature, but a combination of factors e.g. temperature x low oxygen x metabolite load x salinity.
Since most of the radiant energy of light is absorbed as heat by the surface and upper layers of water in the fish ponds, (because of the high concentration of dissolved organic and particulate matter on the top layer of water) the lower water layer becomes cooler. This at times is quite beneficial, for during the hottest part of the day the fish can move down safely to the lower layers. Mixing of the layers would however take place due to wind action. Boyd (1979) (Fig. 8.2) clearly shows a well developed thermocline (thermal stratification) in a fish pond - as we have discussed elsewhere herein this thermocline is not strictly according to the classical concept of thermocline (see below), as the change in temperature with depth in a small pond of say 2.5m is much higher than 1°C/one metre - e.g. given change from 28 to 21°C in a depth change of 1.8 to 2.2 metres (e.g. 7°C per 0.4m or 17.5°C/one metre). In shallow ponds (1m) the water would destratify on cooling at night, whereas in deeper ponds (1.5 – 2m) at Auburn, Alabama remained stratified throughout the warm months (Boyd, 1979).
There is usually a close correlation between air temperature and pond water temperature, though usually pond temperatures do not fluctuate as greatly as air temperature and epilimnion temperature. Temperature of turbid waters are greater than those of clear waters. At the beginning of a plankton bloom the surface temperature of a pond was 31°C, which increased to 35°C at the peak of the algal bloom (Idoso and Foster, 1974), but at a depth of 60cm the temperature remained unchanged.
This aspect of study is relevant to aquaculture in as much as it is important in stocking and management of man-made reservoirs, lakes and large coastal water bodies. A phenomenon of much significance in temperate lakes and also in the deeper tropical water bodies is the thermal stratication of water in summer (cf. fish pond). The warmer surface water remains in a separate zone on the top, referred to as epilimnion, and cooler deep water, being heavier, remaining in the bottom, referred to as hypolimnion (Fig. 8.3). The two zones are separated by the thermocline where the temperature changes sharply with increase in depth - according to Birge the thermocline is characterized by change in temperature of 1°C/metre - perhaps this definition need not be so rigid. According to this characteristic very few tropical lakes have thermocline, eventhough Ruttner in Indonesia and Sreenivasan in India recorded thermoclines in tropical lakes and reservoirs. Worthington and Beadle did not find thermocline in African lakes, Rudolyph, Victoria Nyanza and others, but did find a definite thermocline in Lake Edward. (See “Thermocline in fish pond” discussed earlier).
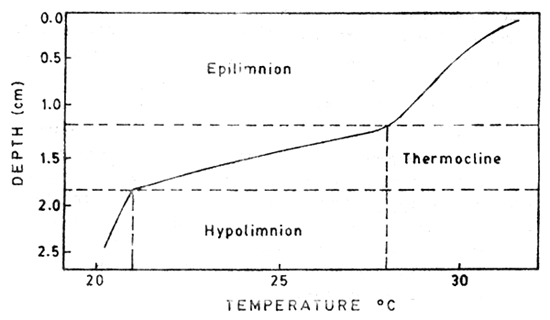
Fig. 8.2. Thermal stratification in a fish pond (after Boyd, 1979)
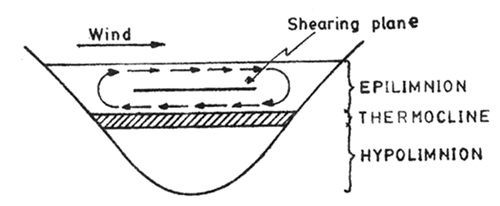
Fig. 8.3. Stratification of water in a natural water body (lake). Formation of horizontal water current and shearing plane in upper portion (epilimnion) is shown
Since water heated by sun remains in the epilimnion itself (mixing due to wind action restricted to the upper zone itself) there is a sharp difference in the physico-chemical and biological characteristics of epilimnion and hypolimnion. These aspects will be discussed separately. (See also water density).
Temperature is often referred to as the master factor among environmental factors affecting aquatic life. All animals have a temperature range, the ‘biokinetic range’, within which they can live indefinitely. This range is limited by the upper and lower tolerance limit, and beyond these critical temperatures the animals may live for a brief period resistance time but would eventually die (lethal zone). Both the tolerance range and resistance time in lethal temperatures are subject to the acclimation temperature i.e. the temperature to which the organisms have been previously exposed. Therefore it is a matter of great concern for aquaculturists not only to know the seasonal and daily changes in temperature of the water body selected for aquaculture but also the thermal tolerance range and thermal history of the organisms to be introduced in the water body.
The thermal tolerance of finfishes and shellfishes vary greatly, those with a wide range of tolerance being referred to as ‘eurythermal’ and those with a narrow range as ‘stenothermal’. The temperature range of a water body as obvious must depend on the latitude or geographical location and also the altitude at which it is located as already referred to; both these aspects have to be considered while transporting and stocking fishes.
In a continent as wide as Africa several temperature regimes would characterise specific locations. Generally the cichlids which are abundant in Africa have a high upper thermal tolerance, Oreochromis mossambicus having as high as 38–39°C, but their lower tolerance limit is also high i.e. they are unable to tolerate cold temperatures even by gradual acclimation; several tilapias cannot be taken below 10°C. They also need warmer temperature to reproduce or even to be active. O. mossambicus ceases swimming activity, in a tunnel apparatus at temperatures below 15 – 20°C, in contrast to several culturable fishes such as the salmonids (stenothermal) - the salmonids cannot tolerate temperatures above 20 – 25°C; the cyprinids are relatively eurythermal, the common carp Cyprinus carpio and goldfish are good examples of eurythermal fishes. Quantified information on thermal tolerance of several cyprinids (Fry et al, 1942; Kassim, 1978), some mullets (Kutty, 1981; Kutty et al, 1978), O. mossambicus (Allanson and Noble, 1968; Aranthakrishnan and Kutty, 1976) and several salmonids - (Brett, 1962) are available in literature. Thermal tolerance range of tilapias are given in Balarin and Hatton (1979) and Pullin and Lowe Mcconnell (1982). The temperature history of the water body and of the selected organisms to be cultured have to be precisely known since growth of fishes is a major interest of the aquaculturist. Information on optimum thermal range for growth has to be known. Here temperature acts as a controlling factor regulating metabolism and thereby growth. Temperature acts also as a directive factor when it causes preference and avoidance of warmer or cooler areas in a water body, causing concentration or complete absence. The importance of these reactions in aquaculture and fisheries is obvious. More details are discussed separately under “Species selection for aquaculture”.
In brackish water ponds the tropical shrimps, Penaeus aztecus, P. merguiensiss, P. mondon, P. indicus, P. japonius, and P. duorarun - P. notialis (= duorarum) in Nigeria are cultured. While the optimal temperature for these shrimps would differ, generally a range of 26 to 30°C is considered optimal for growth i.e. in terms of maximum yield.
The rate of growth of post-lerval P. aztecus increased with temperature upto 32.2°C. Survival for one month was greatly reduced at 32.5°C and no shrimp survived at 35°C (Zein-Eldin and Aldrich, 1965). In case of P. meruguiensis (8.5cm), Piyanka rchana et al (1975) observed that while above 34°C the shrimp became immobile mortality began at temperatures above 40°C. In the case of P. indicus they survived temperatures up to 39°C at salinities of 5–35 p.p.t. and their metabolic rates maintained steady increse up to 39°C (Kutty et al, 1972).
In the case of P. monodon survival was affected at temperatures above 37.5°C.
The temperature of water in a pond can be ususlly decreased during the warm season if water level is increased and vice versa. Temperature death due to high temperature takes place during peak summer and this can thus be avoided often. Provision of shade (cover) over a part of the pond can also be of help in hot months. It is significant that temperatures at which mortality occurs is so precise that change of even a fraction of degree of temperature can make difference of life or death of the aquatic organisms.
In cold countries, providing wind breaks during winter causes increase in temperature of ponds e.g. carp - spawning pond in Europe; Milkfish pond in Taiwan. Evacuation of bottom water by any device (properly designed monk or otherwise) can also help in warming of water. In certain temperate regions tilapias are reared in artificially warmed ponds.
It is also noteworthy that heated effluents from power generation units can be used for warm water facilities in cold months especially for rearing fish as indeed is done in certain cases (salmon and trout culture) in temperate regions.
Water depth in brackishwater ponds is often increased to avoid heat death of shrimps under culture. A minimum of 0.5cm depth is to be maintained and to avoid heat death the ponds can be further deepened up to 1.5 – 2m if the water is clear or deeper channels can be dug. However, it must be ensured that water does not stagnate and stratify - the layout of the pond must be such that wind action does cause water mixing.
![]()
![]()
![]()