
Preliminary Studies on the Effects of Methyltestosterone on Macrobrachium rosenbergii Juveniles
| NACA/WP/86/46 | November 1986 |
|
Preliminary Studies on the Effects of Methyltestosterone on Macrobrachium rosenbergii Juveniles |
Research Conducted under Secondment of Young Scientists Programme
NETWORK OF AQUACULTURE CENTRES IN ASIA
BANGKOK, THAILAND
Hyperlinks to non-FAO Internet sites do not imply any official endorsement of or responsibility for the opinions, ideas, data or products presented at these locations, or guarantee the validity of the information provided. The sole purpose of links to non-FAO sites is to indicate further information available on related topics.
This electronic document has been scanned using optical character recognition (OCR) software. FAO declines all responsibility for any discrepancies that may exist between the present document and its original printed version.
(CRUSTACEANA : PALAEMONIDAE)1
Jocelyn L. Antiporda2
Macrobrachium rosenbergii juveniles were fed hormone-free and hormone-treated diets at 30% crude protein. Methyltestosterone (MT) was incorporated in the diets at 3 levels (25 mg MT/ kg feed, 75 mg MT/kg feed and 125 mg MT/kg feed and administered for 60 days. Growth in all treatments after 30 days post-hormone treatment showed no significant differences (P 0.05) while sex ratios were significantly different in 3 treatments : control v diet I (25 mg MT/ kg feed); diet I v diet IV (125 mg MT/kg feed) (P≥ 0.05). As artificial control of growth and monosex cultures through hormones are possible tools in any cultural management, this preliminary study may inspire further exploration along these lines particularly on the culture of the giant freshwater prawn.
The economic importance of the giant freshwater prawn Macrobrachium rosenbergii in many countries cannot be underscored. A major drawback in its culture, however, is the wide variation in size and the fact that when raised in mixed populations, growth of females is inhibited. This major difference in the growth strategy of the males and females strongly influences the marketable yield (Sagi et al., 1986; Smith et al., 1978; Brody, et al., 1980; Cohen et al., 1981). Therefore, where market demands cater for more uniform sizes, the possibility of monosex culture may come into focus. Production techniques based on monosex sub-populations have been applied in the culture of other domesticated animals and more recently was successfully applied to intensive production of Tilapia spp. (Clemens and Inslee, 1968; Guerrero, 1975; Jensen and Shelton, 1979; Macintosh et al., 1985; Mires, 1977; Nakamura, 1975; Tayamen and Shelton, 1978; Yamazaki, 1976). The production of a monosex population through sex reversal maybe achieved by administering androgens or estrogens. This would eliminate unwanted reproduction and permit farmers to achieve high yields of desirable size.
Orally-administered androgens such as ethynyltestosterone (ethisterone) and methyltestosterone were found more effective than naturally occuring androgens like testosterone, androsterone, adrenosterone, androtenedione, which are more potent when injected intraperitoneally (Clemens and Inslee, 1968; Guerrero, 1975; Nakamura, 1975, Tayamen and Shelton, 1975; Macintosh et al., 1985). Estrogenic steroids were also used to alter the sex of fishes but were found to be less successful than the androgens (Jensen and Shelton, 1979).
The administration of methyltestosterone through prepared feed has been proved successful not only in reversing sex but also as a growth promoter. Fagerlund et al., (1979) fed coho salmon (Oncorhynchus kisutch) fry with moist diet supplemented with 1 mg/kg of 17∞-methyltestosterone and found that hormone-treated fish grew faster and utilized food more efficiently than controls. Acceleration of growth was observed in goldfish (Carassius auratus), horai masu, a variant of rainbow trout (Salmo gairdneri) and kokanee (Oncorhynchus nerka) with the incorporation of methyltestosterone in the diet at a concentration of 1 ppm (Yamazaki, 1976).
This experiment is the first attempt to explore possibility for artificial control of growth and sex differentiation in the fresh-water giant prawn M. rosenbergii using mainly methylstestosterone.
Four treatment feeds were prepared using the same feed composition (Table I) at 30% crude protein. One treatment feed served as control and the three others were formulated to contain 25 mg MT/kg feed, 75 mg MT/kg feed and 125 mg MT/kg feed prepared by the alcohol preparation method (Guerrero, 1975). The various hormone levels were selected based on experiments with tilapias (Clemens and Inslee, 1968; Guerrero, 1975; Nakamura, 1975; Tayamen and Shelton, 1978).
Juveniles were obtained from Chachoengsao Fisheries Station, Chachoengsao, Thailand and acclimated in aquaria at the NIFI Hatchery Building. A total of 800 juveniles (average weight 0.07 g and 16.8 mm in length) were measured from the tip of the telson to the base of the eye orbit and were randomly stocked in 16 indoor glass aquaria measuring 94cm × 47cm × 47cm at the rate of 50 individuals per aquarium. The experiment was conducted with 4 replicates per treatment using completely randomized design. Sampling was done every two weeks with a minimum of 20 animals measured for weight and length in each aquarium.
After the treatment phase of 8 weeks, all prawns were fed hormone-free pelleted diets at 30% CP for one month. Individual prawns was then examined phenotypically under a dissecting microscope to note the presence or absence of appendix masculinus in the second pleopod (Fig. 1) and sex ratios were then determined. All measurements were subjected to analysis of variance using IBM PC Computer (Epistat, Microstat Programs) to determine differences among treatments and t-test to determine which treatments are significantly different from one another.
Table 2 gives a summary of the responses of M. rosenbergii juveniles fed hormone-free and hormone-treated diets. The initial weights were 0.06, 0.08, 0.06 and 0.06g for control, diet I, diet II and diet III respectively, with the corresponding percent weight gains of 1448, 1474, 1412 and 1607 (Table 3 and 5) after 95 days. Diet III has a relatively higher percent weight gain compared with the other treatment groups, although statistically there is no significant difference in all the treatments (P≥ 0.05) as shown in Table 6. Figure 2 shows the growth response of the juveniles for 95 days culture period. In all the treatments, percent survivals are moderate ranging from 53 to 71% with diet II showing the highest survival. Initial lengths (mm) of the test juveniles ranged from 15.8 – 18 with an average of 16.6, 17.4, 16.8 and 16.3 for control, diet I, diet II and diet III, respectively (Table 7). Correspondingly, percent gain in length are 123, 122, 120 and 131% with diet III showing the highest increase in length (Table 9). Analyses of variance showed no significant difference in all the treatments with respect to percent gain in length (Table 10). The growth response of the juveniles expressed in length is shown in Fig. 3.
As shown in Table 11, diet I comes closest to the 1:1 male/ female ratio with the sex ratio value of 0.92. Analysis of variance (Table 12) established a significant difference between the means of 3 treatments. The sex ratio of the control diet is significantly different from that of diet II and diet III is also significantly different from each other, also diet II is not significantly different from diet I (Table 12).
While other researches on fishes and Tilapia spp. in particular, have shown considerable success in inducement of sex reversal and as a growth promoter, studies on crustaceans along this line are practically absent. The findings of Ra'anan and Sagi (1986) that the growth of female M. rosenbergii is strongly affected by the presence of males resulting in growth inhibition of the female, may have given the first call for research efforts directed towards the establishment of prawn monosex populations, in addition for which prawn farmers have long ago suffered the wide variations in the sizes of the cultured prawn. As in Tilapia, the male prawn grows faster and bigger hence production yield will be strongly enhanced through monosex culture. A number of experiments demonstrated the effect of methyltestosterone as a growth promoter and being an effective agent for sex reversal (Macintosh et al., 1985; Fagerlund et al., 1979; Tayamen and Shelton, 1978; Yamazaki, 1976). Although growth in all treatments in the present study, did not show significant differences both in weight and lengths (Table 4 and 8), the treatment levels may prove to be still low as indicated by the result that diet III (125 mg MT/kg feed) has a higher percent weight and length gain than the other treatments or that the hormone used may not have the same growth-promoting effect when administered to other animals such as the freshwater prawn. Macintosh et al. (1985) demonstrated that the anabolic and sex reversal effect of MT on tilapia are proportional to the duration of treatment and the quantity of hormone administered, which may also be true for the prawn. Yamazaki (1976) has observed that 1 ppm MT in the diet, accelerated growth of goldfish (Carassius auratus), a variant of rainbow trout (Salmo gairdneri) and Oncorhynchus nerka. Macintosh et al. (1985) showed that Oreochromis mossambicus fed with 30 ug MT g-1 diet for 60 days grew better than those fed with hormone-free feed. Fagerlund et al. (1979) suggested that coho salmon (Oncorhynchus kisutch) grew faster when fed moist diet supplemented with 1 mg 17 ∞ -MT/kg feed.
The failure to induce sex reversal in M. rosenbergii juveniles in this study may have been caused by a number of factors. The administered doses (25 mg MT/kg feed, 75 mg MT/kg feed and 125 mg MT/kg feed) may be insufficient under crustacean standards but not insufficient as compared to those given to Tilapia spp. (Guerrero, 1975; Mires, 1977; Tayamen and Shelton, 1978; Macintosh et al., 1985). Another factor may be that the treatment was given outside the labile period which covers from the undifferentiated to the sexually differentiated gonadal phase, in which case, hormone must be administered at an earlier stage. In tilapia, for example, sex reversal was successful when administration of hormone was done during the early stages of their lives (Guerrero, 1975; Nakamura and Takahashi, 1973 Clemens and Inslee, 1968; Tayamen and Shelton, 1978). The amount of hormone given for 60 days, in this experiment, still have failed to inactivate the naturally occuring hormones in the gastrointestinal tract or liver, and probably a longer period of treatment maybe necessary, as the effect of the hormone is dose- and time-dependent (Macintosh et al., 1985).
In this experiment, sex ratio in the treatment recieving diet I (25 mg MT/ kg feed), showed the highest value approaching to a 1:1 male/ female ratio and was significantly different from control, and diet III (125 mg MT/ kg feed) but not significantly different from diet II (75 mg MY/ kg feed). The diet 1 amount falls within the effective concentration for tilapia, 10–40 ppm (Clemens and Inslee, 1968); medaka and goldfish, 25 ppm (Yamamoto, 1958 ; Yamamoto and Kajishima, 1968; zebra fish, 1–100 ppm (Yamazaki, 1976). A 30-day post-treatment culture period may not have been sufficient to really obtain a more conclusive findings since it was observed that some prawns were developing appendix masculinus. It is probable that the hormone effect may take longer time than 30 days. One other possibility for the uncertainties in the sex ratio values may be due to the mortalities which might have been too high to affect the interpretation of the results.
While the present study, did not achieve the expected sex reversal by methyltestosterone and has not shown the hormone as a growth promoter in M. rosenbergii juveniles, nevertheless a broad area for similar investigations poses a great challenge. These initial findings may certainly serve as baseline data for further exploratory research. Sex determination in the earlier stages and differentiation, gonadal alteration and artificial control of growth by hormonal treatment in crustacea in general and M. rosenbergii in particular, are still far from established.
I am greateful to all the people/institutions who had in one way or another contributed to the fulfillment of this study. My special thanks goes to Ms. Nanthiya for the things she shared, technical or otherwise.
Brody, T., Cohen, D., Barnes, A. and Spector, A., 1980. Yield characteristics of Macrobrachium rosenbergii in temperate zone aquaculture. Aquaculture, 21:375–385.
Clemens, H.P. and Inslee, T., 1968. The production of unisexual broods by Tilapia mossambica sex-reversed with methyltestosterone. Trans. Amer. Fish. Soc., 97(1): 18–21.
Cohen, D., Ra'anan, Z. and Brody, T., 1981. Population profile development and morphotypic differentiation in the giant freshwater prawn Macrobrachium rosenbergii (de Man). J. World Maricult. Soc., 12 (2): 231–243.
Fagerlund, U.H.M., McBride, J.R. and Stone, E.T., 1979. A test of 17 - Methyltestosterone as a growth promoter in a coho salmon hatchery. Trans. Amer. Fish. Soc., 108:467–472.
Guerrero, R.D., III. 1975. Use od androgens for the production of all-male Tilapia aurea (Steindachner). Trans Amer. Fish. Soc. 104(2): 342–348.
Jensen, G.L. and Shelton, W.L., 1979. Effects of estrogen on Tilapia aurea: Implications for production of monosex gametic male tilapia. Aquaculture, 16:233–242.
Macintosh. J., Vaghese, T.J. and Satyanarayana, G.P. Rao. 1985. Hormonal sex reversal of wild-spawned Tilapia in India. J. Fish. Biol. 26:87–94.
Mires, D. 1977. Theoretical and practical aspects of the production of all-male Tilapia hybrids. Bamidgeh, 29(3):94–101.
Nakamura, M., 1975. Dosage-dependent changes in the effect of oral administration of methyltestosterone on gonadal sex differentiation in Tilapia mossambica. Bull. Fac. Fish., Hokkaido Univ. 26(2): 99–108.
Tayamen, M.M. and Shelton, W.L., 1978. Inducement of sex reversal in Sarotherodon niloticus (Linnaeus). Aquaculture, 14:349–354.
Sagi, A., Ra'anan, Z., Cohen, D. and Wax, Y., 1986. Production of Macrobrachium rosenbergii in monosex populations: Yield characteristics under intensive monoculture conditions in cages. Aquaculture, 51: 265–275.
Schulte, H. 1975. The laboratory culture of the Palaemonid prawn Leander squilla. The European Symposium on Marine Biology, Ostend, Belgium, Sept. 17–23 vol. 1: 437–454.
Smith, T.I.J., Sandifer, P.A. and Smith, H.H., 1978. Population structure of Malaysian prawns, Macrobrachium rosenbergii (de Man), reared in earthen ponds in South Carolina, 1974–1976. Proc. World Maricult. Soc., 9:21–38.
Yamazaki, K. 1976. Application of hormones in fish culture. J. Fish. Board Can. 33:948–958.
Yamamoto, T. 1958. Artificial inductions of functional sex reversal in genotype females of the medaka (Oryzias latipes). J. Exp. Zool., 132(2):227–264.
Yamamoto, T. and Kajishima, T. 1969. Sex differentiation. In W.S. Hoar and D.J. Randall (eds.), Fish Physiology, vol 3 Academic Press, New York, N.Y. pp. 117–275.
Yamamoto, T. and Kajishima, T. 1968. Sex hormonic induction of reversal of sex differentiation in the gold fish and evidence of the male heterogamety. J. Exp. Zool. 162: 215–222.
Takahashi, H. 1975. Functional masculinization of female guppies, Poecilla reticulata, influenced by methyltestosterone before birth. Bull. Jpn. Soc. Fish. 41(5): 499–472.
Table 1. Feed composition
| Feed Ingredients | Test Diets* | |||
| Control | Diet I | Diet II | Diet III | |
| Fish meal (55.3%CP) | 21.1 | |||
| Shrimp head meal (28.8%CP) | 5.2 | |||
| Rice bran (10.5%CP) | 64.7 | |||
| Bassfin | 1.0 | |||
| Vitamin mix** | 0.5 | |||
| Mineral mix*** | 0.5 | |||
| Cellulose | 0.6 | |||
| Oil | 6.3 | |||
| Methyltestosterone**** | 0 | 25 | 75 | 125 |
* Feed composition is the same in all treatments at 30%CP except for the amount of methyltestosterone.
** Vitamin mix provided the following in mg or units per kg of feed: Vit. A 8,818; Vit. D3 2,205 ICU; riboflavin 20; d-Ca panthothenic acid 50; choline chloride 600; niacin 100; Vit. E 50; Vit. B12 20ug; Vit. C 100; folic acid 5; pyridoxine 20; thiamine 20; Vit. K 10; inositol 100; biotin 1.
*** Mineral mix provided the following: Fe 19.8; Mn 60; I 1.2; Zn 44.1; Cu 2; Co 0.2; Dicalcium phosphate 32% calcium, 18% phosphorus. (Adapted from: Balazs, G. and E. Ross 1976. Effect of protein source and level on growth and performance of the captive freshwater prawn, M. rosenbergii. Aquaculture 7: 299–313.
**** The amount is in mg/kg feed.
Table 2. Summary of the response of M. rosenbergii juveniles fed different levels of methyltestosterone. All values are means of 4 replicates.
| Test Diets | ||||
| Control | Diet I | Diet II | Diet III | |
| Initial weight (g) | .0629 | .0765 | .0650 | .0601 |
| Final weight | .9737 | 1.2043 | .9826 | 1.0259 |
| Weight gain | .9108 | 1.1278 | .9176 | .9658 |
| Percent weight gain | 1448 | 1474 | 1412 | 1607 |
| Initial length (mm) | 16.6 | 17.4 | 16.8 | 16.3 |
| Final length | 37.0 | 38.6 | 36.9 | 37.7 |
| Length gain | 20.4 | 21.2 | 20.1 | 21.4 |
| Percent gain length | 123 | 122 | 120 | 131 |
| No. of males | 9 | 14 | 12 | 6 |
| No. of females | 25 | 15 | 24 | 20 |
| Ratio of male/female | .35 | .92 | .48 | .31 |
| Percent Survival | 68 | 59 | 71 | 53 |
Table 3. Mean body weights of prawns (g) fed varying levels of Methyltestosterone.
| Days on Experiment | Test Diets | |||
| Control | Diet I | Diet II | Diet III | |
| 0 | .0629 | .0765 | .0650 | .0601 |
| 18 | .1062 | .1108 | .1065 | .1190 |
| 38 | .2367 | .2562 | .2052 | .2051 |
| 52 | .2785 | .3099 | .2899 | .2875 |
| 67 | .5978 | .6043 | .5561 | .5942 |
| 81 | .7224 | .8211 | .7763 | .7814 |
| 95 | .9737 | 1.2943 | .9826 | 1.0259 |
Table 4. Analysis of variance for mean body weight of the test juveniles.
| SOURCE | SUM OF SQUARES | D.F. | MEAN SQUARE | F RATIO | PROB. |
| Between | .016 | 3 | 5.2216E-03 | .038 | .9899 |
| Within | 3.302 | 24 | .138 | ||
| Total | 3.318 | 27 |
Table 5. Percent gain of M. rosenbergii juveniles based on weight measured every 2 weeks.
| Time interval bet. samplings | Test Diets | |||
| Control | Diet I | Diet II | Diet III | |
| (days) | ||||
| 18 | 68.8 | 44.8 | 63.8 | 98.0 |
| 20 | 122.9 | 131.2 | 92.7 | 72.4 |
| 14 | 17.6 | 20.9 | 41.3 | 40.2 |
| 15 | 114.6 | 94.9 | 91.8 | 106.7 |
| 14 | 20.8 | 35.9 | 39.6 | 31.5 |
| 14 | 34.8 | 46.7 | 26.6 | 31.3 |
| Total % gain | 1448.0 | 1474.2 | 1412.0 | 1607.0 |
Table 6. Analysis of variance for percent gain of test juveniles based on weight.
| SOURCE | SUM OF SQUARES | D.F. | MEAN SQUARE | F RATIO | PROB. |
| Between | 64.875 | 3 | 21.625 | .015 | .9975 |
| Within | 29404.250 | 20 | 1470.213 | ||
| Total | 29469.125 | 23 |
Table 7. Mean body lengths (mm) of prawns fed varying hormone levels.
| Days on Experiment | Test Diets | |||
| Control | Diet I | Diet II | Diet III | |
| 0 | 16.6 | 17.4 | 16.8 | 16.3 |
| 18 | 19.2 | 19.6 | 19.3 | 19.8 |
| 38 | 24.7 | 25.3 | 23.5 | 23.2 |
| 52 | 25.8 | 26.2 | 25.7 | 25.5 |
| 67 | 32.8 | 32.0 | 32.3 | 33.5 |
| 81 | 33.8 | 35.4 | 33.5 | 34.5 |
| 95 | 37.0 | 38.6 | 36.9 | 37.7 |
Table 8. Analysis of variance for mean body length of the test juveniles.
| SOURCE | SUM OF SQUARES | D.F. | MEAN SQUARE | F RATIO | PROB. |
| Between | 3.201 | 3 | 1.067 | .017 | .9968 |
| Within | 1472.909 | 24 | 61.371 | ||
| Total | 1476.110 | 27 |
Table 9. Percent gain of M. rosenbergii juveniles based on length measured every 2 weeks.
| Time interval (days) bet, samplings | Test Diets | |||
| Control | Diet I | Diet II | Diet III | |
| 18 | 15.7 | 12.6 | 14.9 | 21.5 |
| 20 | 28.6 | 29.1 | 21.8 | 17.2 |
| 14 | 4.4 | 3.6 | 9.4 | 9.9 |
| 15 | 27.1 | 22.1 | 25.7 | 31.4 |
| 14 | 3.0 | 10.6 | 3.7 | 3.0 |
| 14 | 9.5 | 9.0 | 10.2 | 9.3 |
| Total % gain | 122.9 | 121.8 | 119.6 | 131.3 |
Table 10. Analysis of variance for percent gain of juveniles based on length.
| SOURCE | SUM OF SQUARES | D.F. | MEAN SQUARE | F RATIO | PROB. |
| Between | 120.985 | 3 | 40.328 | .387 | .7634 |
| Within | 2082.685 | 20 | 104.134 | ||
| Total | 2203.670 | 23 |
Table 11. Sex ratios of M. rosenbergii determined phenotypically at 30 days post-treatment culture.
| Diets | Duration of treatment | Conc. of
M.T. (ppm) | No. of prawns sexed | Male: female | % survival |
| Control | - | 0 | 136 | .35 | 68 |
| Diet I | 60 days | 25 | 101 | .92 | 59 |
| Diet II | -do- | 75 | 142 | .48 | 71 |
| Diet III | -do- | 125 | 106 | .31 | 53 |
Table 12. Analysis of variance for sex ratio of test juveniles.
| SOURCE | SUM OF SQUARES | D.F. | MEAN SQUARE | F RATIO | PROB. |
| Between | 1.152 | 3 | .384 | 6.632 | 6.843E-03 |
| Within | .695 | 12 | .058 | ||
| Total | 1.847 | 15 |
t-Test
| t | ||
| control v diet I | 2.84 | P ≥.05 |
| control v diet II | 2.28 | not significant |
| control v diet III | 0.41 | not significant |
| diet I v diet II | 2.08 | not significant |
| diet I v diet III | 2.89 | p ≥ .05 |
| diet II v diet III | 2.20 | not significant |
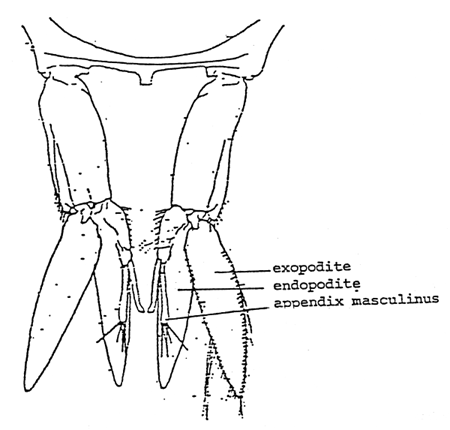
Fig. 1. Second pleopod, sex appendix of M. rosenbergii
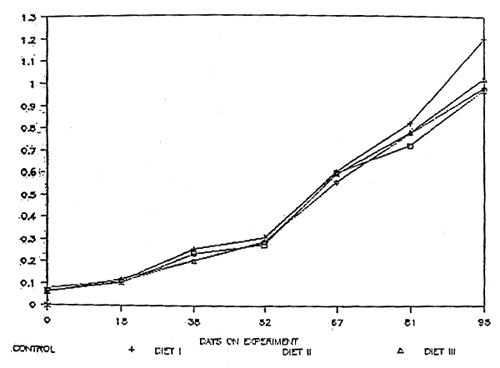
Fig. 2. Growth response of the test juveniles expressed in weight (g).
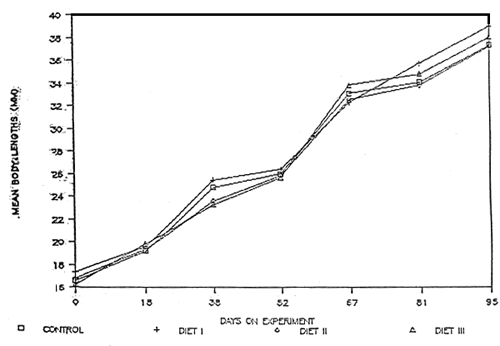
Fig. 3. Growth response of the test juveniles expressed in length (mm)

![]()