![]()
![]()
![]()
L'étude de la croissance des poissons et des phénomènes qui y sont liés, tels que maturation, migration nourriture et habitudes alimentaires, est le pivot de la biologie des pêches et est essentielle pour établir une gestion adéquate et adapté aux conditions locales.
L'ichtyofaune de L'Etang Saumatre est constitué d'au moins 13 éspèces, provenant de 9 familles (voir tableau 3.).
Les premierès neuf espèces sont indigènes dont Strongylura notata et Gobiomorus dormitor sont des prédateurs marins qui ont été enfermés dans le lac dans le passé.
Tilapia mossambica et Cyprinus carpio ont été introduits dans les années cinquante par un projet FAO. Tilapia nilotica a été aperçu seulement dans un endroit près de la ferme commerciale d'Avelech.
C. haitiensis, T. mossambica et G. dormitor sont les principaux espèces exploités par les pêcheurs.
Tableau 3.: L'ichtyofaune de l'Etang Saumatre
| NOM SCIENTIFIQUE | FAMILLE | REMARQUES |
| 1. Limia tridens | Poecilidae | indigène |
| 2. Limia melanotata | Poecilidae | indigène |
| 3. Gambusia hispaniolae | Poecilidae | indigène |
| 4. Gambusia hispaniolae | Poecilidae | indigène |
| 5. Cyprinodon bondii | Cyprinidontidae | indigène |
| 6. Gobionelleus sp. | Gobeidae | indigène |
| 7. Dormitator maculatus | Eleotridae | indigène |
| 8. Gobiomorus dormitor | Eleotridae | indigène |
| 9. Strongylura notata (?) | Belonidae | indigène |
| 10. Cichlasoma haitiensis | Cichlidae | indigène |
| 11. Tilapia mossambica | Cichlidae | introduit |
| 12. Tilapia nilotica | Cichlidae | introduit |
| 13. Cyprinus carpio | Cyprinidae | introduit |
L'étude des peuplements ichtyologiques de l'Etang Saumatre est basée sur la réalisation des pêches avec des filets maillants expérimentaux. Le filet experimental est composé de 6 panneaux de 8.2 m de longeur et de 2.4 m de profondeur et sont associés à des mailles étirées de 25, 37.5, 50, 62.5, 75 et 87.5 mm.
Ces pêches ont été effectuées suivant une périodicité relativement courte, generalement chaque 1 ou 2 mois dans des different secteur du lac.
Lors de chaque pêche, la faune recoltée dans chaque panneaux a été triée à l'éspèce, puis chaque individu a été pésé (au gramme) et mésuré ( longueur totale en mm).
Le sexe et le stade sexuel ont été déterminé. L'échelle de maturation utilisée pour la détermination des stades sexuels par examen macroscopique des gonades est inspirée de celle donnée par Kesteven (voir Annexe 1.).
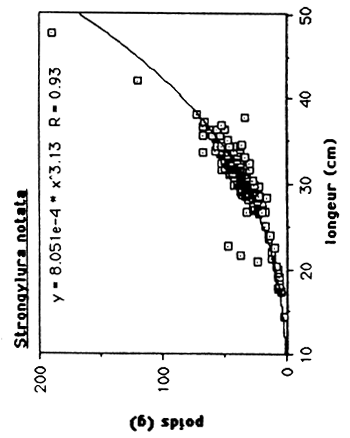
Les rélations poids/longueur ont été calculées pour les trois espéces économiquement les plus importants, Cichlasoma haitiensis, Tilapia mossambica, et Gobiomorus dormitor. On a inclus Strongylura notata. vu son composition élèvé dans les captures experimentals (environ 25% de la biomasse totale).
Pour établir ces rélations des analyses de regression simples basées sur des logarithmes ont été appliquées pour chaque espèce par la relation
W = a × Lb
| dans laquelle, | West le poids en gramme |
| L est la longueur en cm | |
| a est l'intercept ou l'ordonnée à l'origine | |
| b est la pente ou le coefficient de régression |
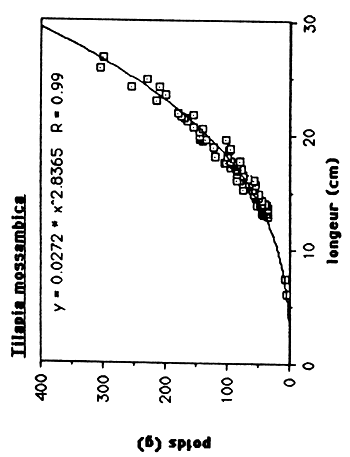
Les résultats sont exprimés dans le tableau 4. et dans le figure 1.. Quand on compare ces régressions de T. mossambica et C. haitiensis avec celles des populations de Miragoine et de Trou Caiman (voir tableau 5. et figure 2. et 3.), on observe que la croissance en masse de la population d'Etang Saumâtre est égal aux celles du Miragoine mais supérieur aux celles de Trou Caiman. Vu que la croissance des poissons est déterminer en premier lieu par des conditions physico - chimiques du milieu, il semble que la salinité (8–15 ppt) de l'Etang Saumuâtre et son dureté élèvé n'influence pas la croissance en masse de T. mossambica et C. haitiensis.
Tableau 4: Les rélations poids/longueur de C. haitiensis, T.mossambica G. dormitor et S. notata de l'Etang Saumatre.
| ESPECE | RELATION | R |
| C. haitiensis | W = 0.0498 L 2.5533 | 0.96 |
| T. mossambica | W = 0.0272 L 2.8365 | 0.99 |
| G. dormitor | W = 0.0034 L 3.3098 | 0.98 |
| S. notata | W = 0.0008051 L 3.31 | 0.93 |
La valeur R ou coefficient de corrélation indique le degré d'une corrélation positive et varie entre - 1.00 et + 1.00..
Figure 1.: Les relations poids/longueur des quatre espèces principales de Etang Saumâtre.
 |  |
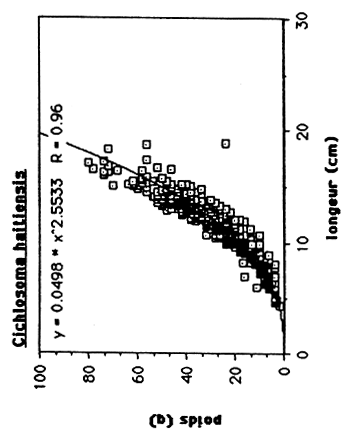 | 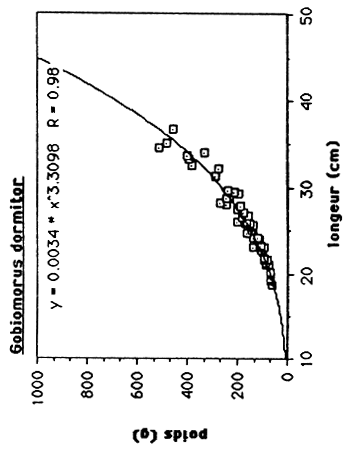 |
Figure 2.: Les relation poids/longueur de C. haitiensis selon les lacs.
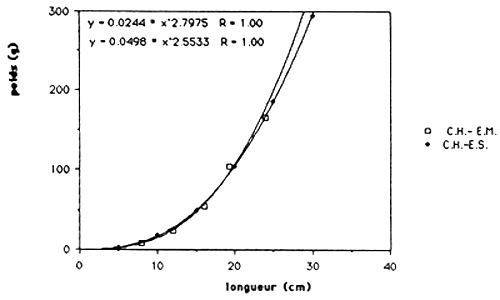
Figure 3.: Les relations poids/longueur de T. mossambica selon les lacs.
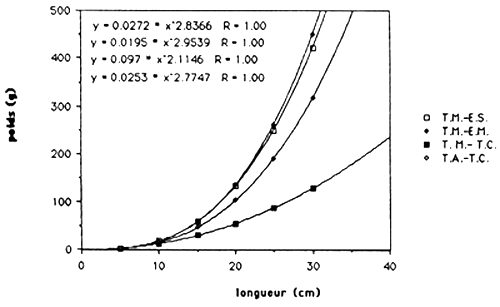
Tableau 5.: Les relations poids/longueur de C. haitiensis, T. mossambica et T. aurea de Trou Caiman et Etang Miragoine.
| ESPECE | LOCALITE | RELATION |
| C. haitiensis | Miragoine (ref 18) | W= 0.0244 L 2.7975 |
| T. mossambica | Miragoine (ref 18) | W= 0.0195 L 2.9539 |
| T. mossambica | Trou Caiman (ref 7) | W= 0.0972 L 2.1146 |
| T. aurea | Trou Caiman (ref 7) | W= 0.0253 L 2.7746 |
Le sex-ratio global (effectif des femelles/ effectif des mâles) des quatre principales espèces capturées en Etang Saumatre est donné dans le tableau 6.
Tableau 6.: Sex-ratio global des quatre principal espèces de Etang Saumatre.
| ESPECE | % MALES | % FEMELLES | SEX-RATIO | EFFECTIF |
| C. haitiensis | 58.9 | 41.1 | 0.69 | 679 |
| T. mossambica | 70.4 | 29.6 | 0.42 | 125 |
| G. dormitor | 51.6 | 48.4 | 0.94 | 273 |
| S. notata | 29.6 | 70.4 | 2.38 | 257 |
Les mâles sont plus nombreux chez les Cichlidae et G. dormitor, tandis que chez S. notata c'est l'inverse. Chez cette dernière espèce le sex-ratio montre une remarquable évolution en fonction de la taille (figure 4.). On observe dans les petits tailles une nette preponderance des males ( 75 à 80 %), puis le sex-ratio tend progressivement à s'équilibrer vers 30 cm pour ensuite s'inverser en faveur des femelles. Au dessus de 35 cm, seulement des femelles ont été capturés.
Une tendence totalement inverse a été observée chez G. dormitor, ou dans les petits tailles il y a un forte pourcentage des femelles et dans les grands tailles seulement des mâles sont capturés (figures 4.).
Chez les Cichlidae le sex-ratio est équilibré dans les petits tailles et dans les grandes tailles seulement des mâles ont été capturés.
Plusieurs hypothèses peuvent être invoquées pour rendre compte du sex-ratio global déséquilibré en faveur des mâles chez T. mossambica et en faveur des femelles chez S. notata et de la relation sex-ratio/taille observée.
Vu les différences comportementales chez T. mossambica entre les mâles et les femelles durant la reproduction où le mâle défend la niche et la femmelle se disperse après la ponte, les mâles ont une répartition plus bordière que les femelles. Alors ils sont plus accesible aux les engins de pêche.
Les schémas comportementaux associés à la reproduction et aux soins parantaux prodigés aux öeufs et aux alevins repartis nettement les quatre espèces.
T. mossambica est un incubatuer buccale. Lors de la reproduction, le mâle fait un nid, dans des eaux peu profondes, dans lequelle les oeufs sont déposés et fecondés avant d'être rapidement repris en bouche par la femelle. L'éclosion des oeufs a lieu dans la cavité buccale de cette dernière et ce n'est pas qu'après la résorption complète de la vésicule viteline que ce comportement de protection parantale s'achève et que les alevins devienent définitivement autonome.
G. dormitor est un pondeur sur substrat et se reproduit à des profondeurs de plus de 10 m. Au moment de la ponte les oeufs sont déposés en masse dans des étroites fissures des roches et font l'objet d'une garde parentale vigilante qui limite l'action des prédateurs éventuels. Après deux ou trois jours l'éclosion est terminé et les alevins se dispersent durant la nuit. Notons que G. dormitor est capable de produire plusieures pontes dans une période relativement courts (6 à 7 jours, ref 12, 13, 16).
Chez C. haitiensis il n'y a aucune référence quant à sa biologie. Neanmoins, des observations sur le terrain montrent que cette espèce utulise le littoral supérieur comme zone de reproduction.
Concernant la comportement reproductif de S. notata, une recherche bibliographique n'a donné aucune information. La concentration des aiguilettes autour des sources subaquatiques durant certaines périodes de l'année et la grande dimension des oeufs (flottant ou semi - flottant ?) nous suggèrent que c'est une espèce qui se reproduit en bande et que l'eau des sources subaquatiques tiennent les oeufs en suspension jusqu à l'éclosion. Une fois l'éclosion terminés, les alevins se dirigent vers le littoral.
La taille de première maturation sexuelle (L50) est ici définie comme la longueur totale à laquelle 50% des poissons se trouvent à un stade avancé (égal ou supérieur au stade 4 de l'échelle de maturation) du premier cycle sexuel.
De plus, afin de mieux cerner l'étalement de la gamme dans laquelle la première maturation est susceptible d'intervenir, il est utile de préciser également la longueur du plus petit individu mature capturé et aussi la taille à laquelle la quasi totalité (95 %) des poissons observés est en stade de maturité avancé.
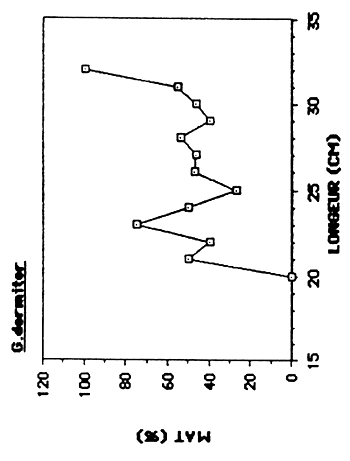
L'aquisition progressive de la maturité sexuelle en fonction de l'accroissement en taille des poissons est représentée sur la figure 5.. La taille du plus petit individus mature capturé ainsi que le L50 et L95 détermineé graphiquement sont présentés dans le tableau 7.
Tableau 7.: Caractéristiques de la taille de première maturation chez les quatre espèces principaux présentes en Etang Saumatre.
| ESPECE | EFFECTIF(F) OBSERVEE NBR | L50 (MM) | L95 (MM) | PLUS PETIT FEMELLE MATURE (MM) |
| C. haitiensis | 296 | 125 | 160 | 98 |
| T. mossambica | 35 | 170 | 190 | 129 |
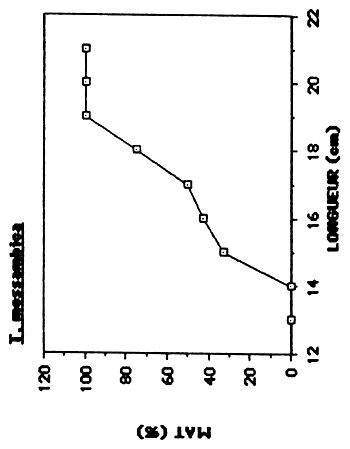
| G. dormitor | 144 | 210–310 | 320 | 195 |
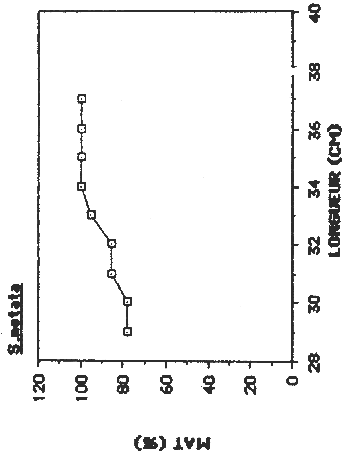
| S. notata | 181 | < 290 | 330 | 215 |
Figure 4.: Evolution du sex-ratio en fonction de la taille des quatre espèces principales de l'Etang Saumatre.
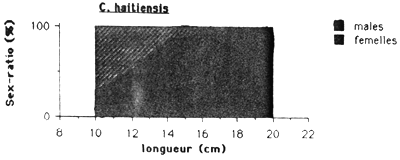
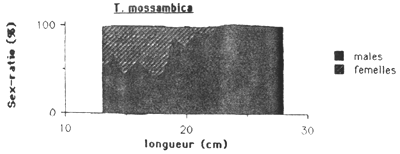
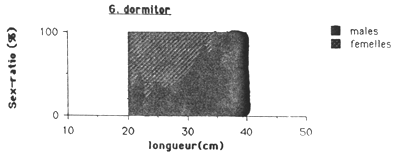
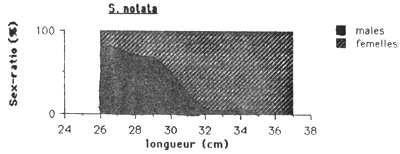
Figure 5. Détermination de la taille de première maturation sexuelle chez les quatre espèces principales de l'Etang Saumâtre.
 |  |
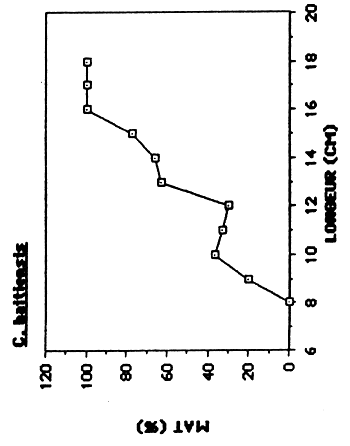 |  |
On constate que les femelles de C. haitiensis entrent en maturation a une taille très inférieur (125 mm) à celle observée pour les femelles de T. mossambica (170 mm).
Chez G. dormitor, le nombre restreint d'individus capturés par classe de taille rend une détermination précise du L50 difficile, qui est situé entre 210 et 300 mm.
Chez S. notata on observe dans les plus petits tailles capturés (290 mm) déja un forte pourcentage des femelles (77.7%) en stade de maturation avancée, donc L 50 est plus petit que 290 mm.
Il n'existe pas un saison de reproduction bien marqué chez C. haitiensis et il peut se reproduire tout au long de l'année. On observe en effet des femelles à tous des stades sexuels dans les échantillons mensuels. avec une proportin toujours d'important d'individus en maturation. Ce même tendence est observé chez T. mossambica.
Chez S. notata on observe une concentration plus élevée pendant le mois de juillet ou 68.36 % des femelles capturés étaient dans une stade de maturation avancée. C'est pendant cette période qu'on a remarqué le bande d'aiguilettes autour les sources subaquatiques.
Chez G. dormitor on observe une concentration plus élèvée pendant le mois d'avril. Ceci corrobore avec les observations de Mc Kay, qui rapporte que la reproduction de cet espèce parvient à la fin de la saison sèche, juste avant le début de la saison pluvieuse, au Nigaragua (ref 12.)
Des observations régulières sur le terrain apportent des précisions sur la régime alimentaire de certaines espèces. La diète alimentaire chez C. haitiensis est basé principalement sur des algues et des petits coquillages qui se trouvent en abondance sur les végétation aquatiques. T. mossambica a un regime alimentaire qui est constitué de phytoplancton des eaux de surface.
G. dormitor et S. notata se nourissent avec des alevins de T. mossambica., C. haitiensis et des poecillidae.
La connaissance du type de croissance des poissons est essentielle pour la plupart des estimations de stock, étant donné que c'est la croissance individuelle des poissons qui regit d'année en année les captures effectuées par une pêcherie.
Les données biologiques qui peuvent être utilisées pour estimer ces paramètres de croissance sont de trois types principaux;
Vu qu'une étude de marquages demande beaucoup de materiel et de temps et que l'examen des écailles n'étaient pas concluant, la methode graphique de Gulland et Holt, basée sur l'analyse des fréquences de tailles était choisi pour estimer les paramètres (ref.: 17)
Le tableau 8. recapitule les paramètres de croissance L8 et K pour les quatre espèces principales de L'Etang Saumatre. Les fréquences de longueurs des ces quatre espèces capturés dans ce lac et les calcules relative se trouvent en Annexe 2.
Tableau 8.: Les paramètres de croissance pour les principales espèces de Etang Saumâtre
| ESPECE | L8 (cm) | K |
| C. haitiensis | 22.10 | 0.139 |
| T. mossambica | 31.90 | 0.457 |
| G. dormitor | 50.91 | 0.304 |
| S. notata | 48.12 | 0.633 |
La mortalité totale Z a été estimée pour C. haitiensis, T. mossambica et G. dormitor à partir des structures démographiques. (voir Annexe 3). La mortalité naturelle M a été estimer à partir l'equation
Log M = -0.0066 -0.279 Log L8 + 0.6543 Log K + 0.4634 Log T
ou T est le temperature moyen.
Vue que Z = F+M, on peut calculer la mortalité par pêche F. Le tableau 9. recapitule ces estimations sur les différents mortalités (ref.: 17).
Lors qu'on dispose des valeurs de F et M, un taux d'exploitation E peut être calculer par
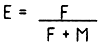
dans laquelle F est la mortalité par pêche
M est la mortalité naturelle d'un stock donné
Cette equation permet d'estimer si un stock est surexploité ou non, à partir de l'hypothese que la valeur optimal de E est proche de 0.5
Tableau 9.: Les taux d'exploitation pour C. haitiensis, T. mossambica et G. dormitor à l'Etang Saumatre.
| ESPECE | Z | F | M | E |
| C. haitiensis | 0.5872 | 0.04 | 0.54 | 0.07 |
| T. mossambica | 1.2066 | 0.14 | 1.06 | 0.12 |
| G. dormitor | 2.7435 | 1.35 | 1.39 | 0.49 |
On observe que le taux d'exploitation des deux Cichlidae est très inferieur à 0.5, donc ses deux espèces sont sous-exploités. G. dormitor, au contraire est exploité au maximum.
Pour rendre compte ces diffèrences entre le taux d'exploitation des Cichlidae et de G. dormitor, l'hypothèse suivant peut être invoqué. Les Cichlidae éffectuent des migrations inter-habitats au cours de leurs cycles biologiques, de façon à satisfaire aux besoins des divers stades. Au stade de jeunes immatures ils se dispersent dans l'ensemble du lac pendant la maturation des gonades. A maturité sexuelle ils retournent au littoral supérieur pout y frayer et élèvér les alevins. Une fois l'élèvage terminé, les femelles se dispersent encore pendant que les mâles gardent les nids. G. dormitor, par contraire, reste prèsque toujours dans le littoral durant sa vie. Il descend seulement dans les profondeur plus élèvés pendant la saison de reproduction. Vu que actuellement la pêche s'éffectue seulement dans le littoral, par manque de transport adéquate, le G. dormitor est plus accèsible aux pêcheurs.
Maintenant, pour augmenter la taux d'exploitation des Cichlidae, il est souhaitable d'améliorer et augmenter la capacité de transport des pêcheurs et en même temps d'essayer et puis introduire des nouveaux techniques de pêches, comme le chalut au boeufs ou des filets maillants de fond.
En géneral, il est indésirable que les jeunes poissons qui deviennent disponibles à la pêcherie sont capturés avant qu'ils sont matures parce-que ainsi la capacité de reproduction de la population est réduite. Alors des recommandations doivent être faites pour assurer que la majorité des poissons capturés ont eu l'opportunité de se reproduire au moins une fois.
Pour déterminer la dimension minimum des mailles pour des filets maillants, deux types d'information sont nécessaire;
Après la collecte de ces données, L 50 est comparé avec L min et seul les dimensions des mailles qui captures des poissons dont L min est égal ou plus petit que L 50 sont considerées comme désirables et la plus petite de ces dimensions est la dimension minimale.
La détermination de la dimension minimale des mailles pour la protection des poissons immatures et son application est une méthode effective pour la gestion d'une pêcherie pour obtenir des poissons de bonne qualité. Neanmoins ceci risque de diminuer la quantité des poissons pêché parce-que d'autres espèces plus petites et généralement plus abondantes sont exclus des captures.
Dans le cas de l'Etang Saumatre et en prenant en considération l'empoissonement massif prévu de Tilapia aurea, il serait souhaitable de déterminer la maille minimale à 75 mm (voir tableau 10 et figure 6.). Par cette action, on élimine la capture de C. haitiensis par des filets maillant qui est actuellement l'espèce la plus abondante. L'autre obstacle est, si une maille minimale est determiner, de le faire respecter. Vu qu'il n'y a pas actuellement un système de controle pour la pêcherie établit, ceci doit être un des responsabilités des pêcheurs pour qu'ils pulssent eux-mêmes veiller à l'application des réglements.
Figure 6.: Sélectivité des filets maillants pour C haitiensis, T. mossambica et G. dormitor.
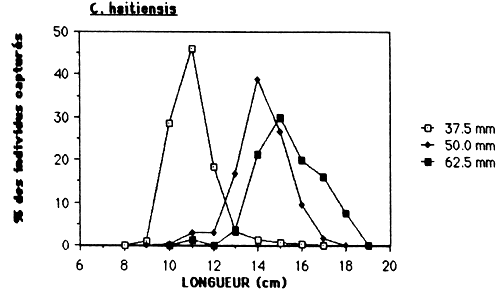
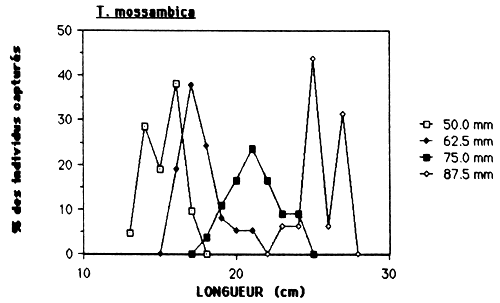
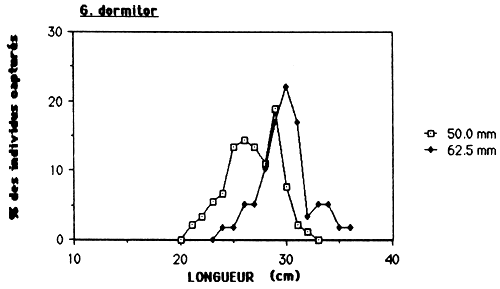
Tableau 10.: Caractéristiques de la sélectivité des filets maillants pour C. haitiensis, T. mossambica et G. dormitor à l'Etang Saumåtre.
| ESPECE | L 50 | M.E. 37.5MM | M.E. 50.0MM | M.E. 62.5MM | M.E. 75.0MM | ||||
| moy. | L min | moy. | Lmin | moy. | Lmin | moy. | Lmin | ||
| mm | mm | mm | mm | mm | mm | mm | mm | mm | |
| C. haitiensis | 125 | 110 | 98 | 140 | 126 | 150 | 136 | - | - |
| T. moss. | 170 | - | - | 150 | 139 | 170 | 160 | 210 | 191 |
| G. dormitor | 210–310 | 240 | 202 | 270 | 236 | 300 | 272 | 340 | 293 |
![]()
![]()
![]()