![]()
![]()
![]()
por
ALFREDO DE LA CRUZ S.
Centro de Investigaciones Pesqueras
Instituto Nacional de la Pesca
Playa Habana, Bauta, Cuba
Extracto
Con el fin de conocer la morfología de las larvas del camarón comercial Penaeus schmitti Burkenroad y sus diferencias con las de Penaeus duorarum, se realizó un muestreo de plancton en Tunas de Zaza, un área camaronera al sur de Cuba, donde existe un predominio de P. schmitti, encontrándose hembras próximas a desovar. Se aplica el plan de trabajo y métodos empleados y se describen las nuevas larvas encontradas. Se discute la posibilidad de que las larvas descritas pertenenzan a P. schmitti o al género Sicyonia. Se hacen sugerencias para futuros trabajos.
PRESENCE OF CERTAIN PENAEID LARVAL STAGES IN A SHRIMPING AREA SOUTH OF CUBA
Abstract
With a view to studying the morphology of the larvae of the commercial shrimp species Penaeus schmitti Burkenroad and differentiating it from the pink shrimp, P. duorarum, a plankton sampling was made at Tunas de Zaza, a shrimping area south of Cuba, where P. schmitti predominates and females ready to cast their eggs were found. The plan of work and methods used are described as are also the young shrimp caught. There follows a discussion of whether the larvae described belong to the P. schmitti or the Sicyonia genus. Suggestions for future work are profferred.
PRESENCE DE QUELQUES STADES LARVAIRES DE PENEIDES DANS UNE ZONE A CREVETTES AU SUD DE CUBA
Résumé
Pour étudier la morphologie des larves de la crevette Penaeus schmitti Burkenroad, présentant un intérêt commercial, et la comparer avec Penaeus duorarum, on a procédé à des prélèvements d'échantillons de plancton à Tunas de Zaza, zone à crevettes située au sud de Cuba où prédomine P. schmitti et où l'on a rencontré des femelles prêtes à pondre. L'auteur décrit le plan de travail et les méthodes employées, ainsi que les nouvelles larves rencontrées. Il examine la question de savoir si les larves décrites appartiennent au genre P. schmitti ou au genre Sicyonia. Quelques propositions sont avancées pour les activités futures.
Los camarones son una parte muy importante en las pesquerías cubanas, pues desde hace muchos años se realiza una captura considerable de los mismos en las zonas de plataforma que rodean la Isla de Cuba.
Es sabido que los camarones poseen características biológicas complejas y que su estudio plantea problemas de muy diversa índole. Entre esos problemas, las migraciones de los adultos y la determinación de los lugares de desove y de cría, están relacionados con el estudio de las larvas de camarones.
En Cuba se ha registrado la presencia de más de diez especies diferentes de camarones, aunque de ellas solamente tienen importancia comercial el camarón acaramelado, Penaeus duorarum Burkenroad, 1939 y el camarón blanco, Penaeus schmitti, Burkenroad 1936. La captura de dichas especies se realiza casi exclusivamente en la plataforma sudoriental de Cuba, que se extiende desde Cienfuegos hasta Cabo Cruz. Según un muestreo, la composición por especies da un 66% de P. duorarum y un 34% de P. schmitti.
Los estadios larvales de P. duorarum a partir del huevo, los nauplios, protozoeas, mysis y post-larvas, son bien conocidos después del excelente trabajo realizado por Dobkin (1961) en aguas de la Florida. Hasta donde alcanzan nuestros conocimientos, los estadios larvales de P. schmitti no están descritos todavía. Por lo tanto, en Cuba, la investigación de la biología pesquera de los camarones debe basarse, principalmente en un estudio de los estadios larvales de P. schmitti para conocer su morfología, y establecer la diferenciación de las especies durante su período larval. Por esa razón, el Centro de Investigaciones Pesqueras, del Instituto Nacional de la Pesca, ha dedicado especial atonción a dicho estudio.
Los objectivos de la investigación sobre larvas de camarones han sido los siguientes:
describir los estadios planctónicos de P. schmitti;
señalar las principales diferencias entre los distintos estadios larvales de las dos especies comerciales cubanas.
El presente informe se elabora con los resultados obtenidos hasta ahora, ya que los trabajos continúan realizándose.
Deseamos agradecer a Miroslav Nikolió, biólogo pesquero de la FAO en Cuba, su constante apoyo y sugerencias en el desarrollo de este trabajo. También a Nancy Aguiar por la ayuda prestada en el laboratorio.
El modo más seguro para conocer los estadios larvales de una especie consiste en la cría de la misma desde el huevo en condiciones controladas de laboratorio, pero dicho método no nos ha sido posible seguirlo hasta el presente. No obstante, y teniendo en cuenta que las poblaciones de las dos especies comerciales cubanas de camarones se encuentran mezcladas, hemos tomado muestras de plancton de las áreas de pesca, basándonos en el siguiente plan de trabajo:
si existen especies cuyos adultos son predominantes en una región, y se encuentran en período de desove, sus larvas deben encontrarse en abundancia;
si al tomar una muestra de plancton en un área con las dos especies predominantes, las larvas de camarones encontradas coinciden morfológicamente con la descripción de Dobkin, se trata de larvas de P. duorarum;
si encontramos larvas que no coinciden con la descripción de Dobkin, y que a su vez son abundantes, podemos suponer con mucha probabilidad que se trata de larvas de P. schmitti, las cuales intentamos describir.
En el muestreo de la zona estudiada hemos encontrado dos tipo de larvas de camarones abundantes:
un tipo que concuerda con la descripción de Dobkin en casi todos sus detalles, aunque de tamaño algo menor. Se trata de larvas de P. duorarum;
un tipo que no concuerda con la descripción de Dobkin, que es tan abundante como el tipo anterior y que forma una serie lógica en la sucesión de los estadios.
El material para este trabajo consiste en 28 muestras de plancton recogidas a bordo del barco “Camarón” que se dedica a la pesca de arrastre en el plan de investigación de los camarones realizado por el Centro de Investigaciones Pesqueras. El viaje se efectuó del 14 al 23 de septiembre de 1966, en áreas cercanas a la costa de Tunas de Zaza, al sur de Cuba, una región de pesca tradicional de camarón blanco, cuyos sedimentos superficiales del fondo están formados por fango gris.
Se tomaron muestras horizontales de plancton en dos secciones que se extienden desde la costa hasta cuatro millas de distancia de la misma. La primera sección se hizo partiendo de la desembocadura del río Zaza, y la segunda en la Ensenada de Guayacanes. En las doce estaciones realizadas se recogió plancton en la superficie y a una profundidad que oscila entre 2 y 10 m. La red de plancton utilizada tenía 30 cm de diámetro en la boca y una tela filtrante de seda de 0,31 mm de abertura de malla. Al no utilizarse una red de cierre, la muestra horizontal subsuperficial tiene una pequeña mezcla con las aguas superficiales en el momento de su recogida. Todos los muestreos fueron de 1 minuto de duración y la velocidad del barco era de 1,8 m/seg.
A la par que los muestreos de plancton, se realizaron capturas por arrastres en las mismas estaciones. Utilizando una red tipo americano semibalón de 375 y otra tipo japonés de 350 mallas, se pescó durante 75 horas de trabajo en 10 arrastres diurnos y 24 nocturnos, capturando un total de 1.400 kg de camarones. De esa captura se hicieron 8 estudios biométricos tomando muestras al azar, analizándose en total 803 camarones, de los cuales el 95% pertenecía a la especie de P. schmitti y el 5% restante a P. duorarum. De las 379 hembras de P. schmitti, el 5,8% (22 ejemplares) estaban impregnadas.
En el laboratorio, las larvas conservadas en formol al 2%, se separaron de las muestras y se estudiaron. Al hacer las descripciones se han tomado en consideración solamente los caracteres más notables. Los dibujos han sido realizados en cámara clara y esquematizados. Al lado de cada esquema aparece una recta dividida en 0,1 mm a escala con el dibujo. Algunas setas se suprimen y en las setas plumosas los cilios no se dibujan en su totalidad para no oscurecer el esquema.
4.1 Primera protozoea
En las larvas que corresponden a este estadio no se han encontrado ejemplares que difieran de la descripción dada por Dobkin para P. duorarum.
4.2 Segunda protozoea
Se examinaron 12 larvas de segunda protozoea (Fig. 1). En el estadio protozoea las larvas fueron medidas desde el borde anterior del carapacho hasta el borde posterior de las ramas furcales para hacer comparable la longitud de este estadio con la del mysis. El largo de la segunda protozoea oscila entre 1,1 y 1,3 mm.
En este estadio los ojos ya formados aparecen fuera del carapacho, el abdomen se segmenta y aparece el rostro.
El carapacho es pequeño, bastante ajustado al cuerpo del animal, está algo levantado, y no cubre ningún segmento torácico. El carapacho tiene una longitud equivalente a la tercera parte de la longitud del animal. El rostro que aparece por primera vez en este estadio, es corto y está algo curvado ventralmente. La longitud del rostro es de un décimo de la longitud del carapacho aproximadamente. Las porciones laterales del borde posterior del carapacho están algo extendidas hacia atrás.
Los segmentos torácicos son seis, al igual que los segmentos del abdomen. El sexto segmento abdominal es más largo que los primeros cinco. Estos cinco primeros segmentos abdominales son ligeramente mayores que los torácicos y no presentan espinas en su parte dorsal.
No existe división entre el sexto segmento abdominal y la porción posterior del animal. Esta última porción está dividida en dos ramas furcales, cada una de las cuales tiene siete espinas. Seis de ésas son setosas y terminales y la séptima es lisa y dorsal.
Los ojos formados son pedunculados en este estadio y se proyectan hacia afuera, libres del carapacho.
La primera antena está formada por tres porciones, de las cuales la basal está segmentada a su vez en 5 partes. La segunda antena consta de un protopodio, un exopodio y un endopodio.
Los pereiópodos aparecen en este estadio como botones muy rudimentarios. No existen pleópodos ni urópodos.
Las principales diferencias entre la especie P. duorarum y las larvas encontradas en este estadio son las siguientes:
en P. duorarum el carapacho es ancho y llega hasta el tercer segmento torácico, mientras que en las larvas encontradas el carapacho es estrecho y no cubre ningún segmento torácico;
el rostro es largo en P. duorarum y corto en las larvas encontradas;
en las larvas encontradas no existen espinas supraorbitales, mientras que en P. duorarum son evidentes y bifurcadas.
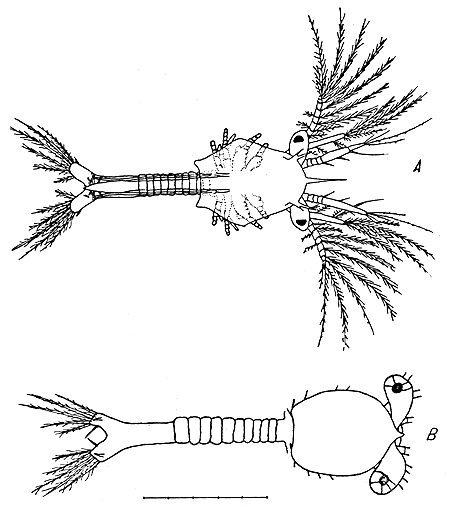 | 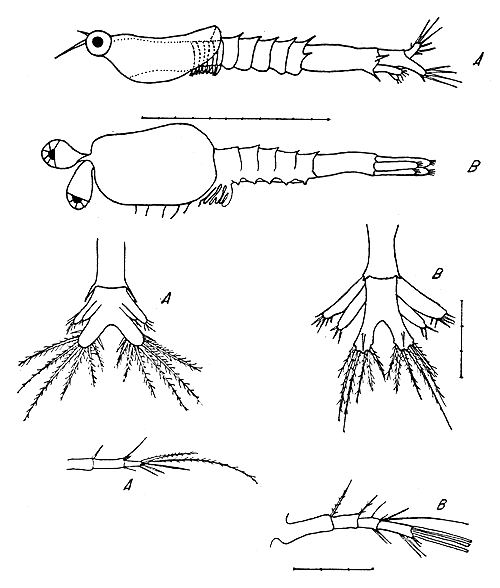 |
Fig. 1 Segunda protozoea, vista dorsal: | Fig. 2 Tercera protozoea, vista lateral, rama furcal, primera antena: |
4.3 Tercera protozoea
Se examinaron 32 larvas en el estadio de tercera protozoea (Fig. 2). La longitud total de las mismas oscila entre 1,4 y 1,9 mm.
Este estadio se diferencia del anterior principalmente por la aparición de los urópodos y las espinas del borde superior de los segmentos abdominales.
Visto desde arriba el carapacho alcanza el primer segmento torácico, y tiene dos expansiones laterales que se extienden hacia atrás, hasta el tercer segmento torácico. El rostro está presente, pero no se observan espinas supraorbitales.
Los segmentos torácicos son estrechos, mientras que los abdominales están mucho más desarrollados. Los cinco primero segmentos abdominales son de tamaño similar, pero el sexto segmento está muy desarrollado, siendo su longitud igual a la de los cuatro segmentos precedentes unidos. Existen espinas dorsomedianas en el borde posterior de los cinco primeros segmentos abdominales, pero no son muy pronunciadas. El segmento abdominal quinto posee un par de espinas posterolaterales. Además, el borde inferior de las pleuras de los cinco primeros segmentos abdominales se prolonga en una expansión terminada en punta, carácter que en este estadio no está muy desarrollado. El sexto segmento abdominal está separado del telson, es alargado, y lleva en su borde posterior un par de espinas posterolaterales situadas encima de la inserción de los urópodos y un par de espinas posteroventrales situadas debajo de la inserción de los urópodos. Todas las espinas del abdomen están dirigidas hacia atrás.
Cada rama furcal posee siete espinas, de las cuales una es dorsal y pequeña; otra es lateral, mediana y se cruza en la línea media con la del lado opuesto; y las otras cinco son grandes y terminales (Fig. 2).
Las primeras antenas (Fig. 2) son unirramosas, están dirigidas hacia delante y están formadas por cuatro segmentos. El primer segmento tiene una seta larga en su extremo distal, y el segundo lleva una seta mediana en su extremo distal. El tercer segmento tiene una seta mediana en su borde interno, dos setas en su extremo externo y tres setas en su extremo interno, de las cuales una es de gran dimensión. El segmento distal tiene dos setas en su borde externo y cinco setas en su extremo. Hay un total de quince setas en la primera antena. Las segundas antenas son birramosas y coinciden en sus detalles con la descripción de Dobkin.
Los pereiópodos son birramosos y rudimentarios. Los pleópodos no están desarrollados. Los urópodos se forman a nivel del extremo posterior del sexto segmento abdominal. Están formados por dos ramas, de las cuales la externa es mayor y tiene seis setas terminales, y la interna tiene tres setas terminales.
En el estadio de tercera protozoea las larvas encontradas se diferencian de la especie P. duorarum por los siguientes caracteres:
P. duorarum no posee expansiones en bordo inferior de la pleura de los cinco primeros segmentos abdominales, mientras que las larvas encontradas sí las presentan;
las larvas encontradas tienen siete espinas en las ramas furcales y P. duorarum tiene ocho;
P. duorarum tiene tres segmentos en la primera antena, mientras que las larvas encontradas tienen cuatro.
4.4 Primera mysis
Se examinaron 22 larvas en el estadio de primera mysis (Fig. 3). La longitud total oscila entre 1,9 y 2,2 mm. En el estadio mysis la longitud de las larvas se midió desde el borde anterior del carapacho hasta el borde posterior del telson.
El estadio de primera mysis se diferencia mucho del de tercera protozoea anterior. se caracteriza por su aspecto, más parecido a un camarón, por el desarrollo de los apéndices torácicos, la morfología de las primeras y, segundas antenas, el desarrollo de los urópodos y la formación del telson.
El carapacho no cubre el tórax. Tiene expansiones laterales en el borde posterior y se encuentra algo levantado. El rostro está proyectado en línea recta hacia delante y presenta una espina hacia la base. Se observan espinas supraorbitales cortas y un par de espinas en la esquina anterolateral del carapacho.
Los cinco primeros segmentos abdominales son de tamaño menor que el sexto. Estos segmentos tienen espinas dorsomedianas muy reducidas o desaparecidas, mientras que hacia hacia la cara ventral se observa el margen puntiagudo de las pleuras abdominales que están expandidas. El sexto segmento abdominal presenta una espina dorsomediana en su borde posterior, pero no presenta espinas ni por encima ni por debajo de la inserción de los urópodos. El quinto segmento abdominal no presenta espinas posterolaterales.
En este estadio se ha producido el acercamiento y fusión de las ramas furcales y el telson se presenta como un apéndice único en el extremo posterior de la larva (Fig. 3). La hendidura que se encuentra en la línea media es profunda. El punto más profundo de la hendidura está situado a nivel de la región entre la espina lateral y la espina terminal mas externa. El telson presenta seis pares de espinas terminales y un par de espinas laterales.
La primera antena (Fig. 3) está transformada. Consta de tres segmentos, siendo el primero tan largo como los otros dos unidos. El extremo distal presenta dos ramas, de las cuales la externa tiene doble longitud que la interna. El segmento basal presenta dos espinas, una corta en su base y la otra larga en la unión del tercio medio con el tercio distal. Los bordes y extremos de los segmentos de la primera antena tienen varias setas plumosas.
Los pereiópodos son birramosos y en este estadio alcanzan mayor tamaño. Los pleópodos no están desarrollados. Los urópodos están a ambos lados del telson. Consta de un protopodio, del cual salen el exopodio ancho y el endopodio estrecho, ambos de igual longitud.
En el estadio de primera mysis las larvas encontradas differen de las de la especie P. duorarum en los siguientes caracteres:
las larvas encontradas presentan una espina a nivel del rostro, mientras que en P. duorarum el rostro es liso;
las larvas encontradas tienen bien desarrolladas las expansiones del borde ventral de las pleuras de los cinco primeros segmentos abdominales, mientras que P. duorarum no tiene esas expansiones;
en el telson, las larvas encontradas tienen seis espinas terminales y una lateral, mientras que P. duorarum tiene siete espinas terminales y una lateral;
en el segmento basal de la primera antena las larvas encontradas presentan dos espinas, mientras que P. duorarum solamente presenta una.
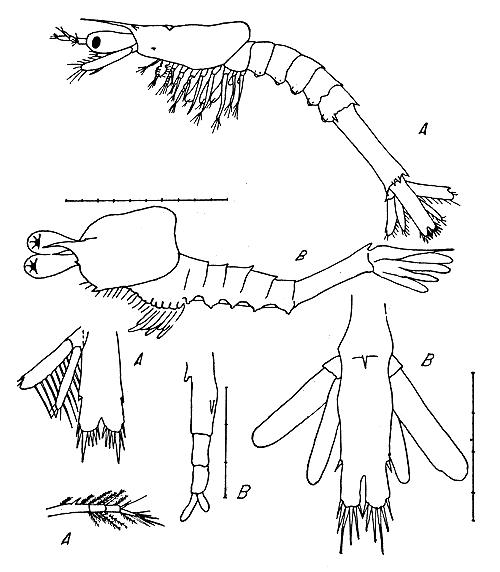
Fig. 3 Primera mysis, vista lateral, telson, primera antena:
A) P. duorarum (según Dobkin)
B) larvas encontradas.
4.5 Segunda mysis
Se analizaron 16 larvas en estadio de segunda mysis (Fig. 4). La longitud total oscila entre 2,1 y 2,4 mm.
Este estadio se diferencia del anterior por la menor profundidad de la hendidura del telson, la aparición de los pleópodos, el aumento de una espina detrás del rostro, y los estatocistos situados en la base de la primera antena.
El carapacho cubre escasamente los primeros segmentos del tórax, aunque sus expansiones laterales alcanzan hasta el cuarto o quinto segmento torácico. El rostro se extiende hacia delante en línea recta y en este estadio se encuentran dos espinas pequenas en la carina detrás del rostro. Las espinas supraorbitales son cortas y las anterolaterales pronunciadas. Entre ambas existe una expansión terminada en punta que corresponde a las espinas suborbitales.
En el abdomen es notable el desarrollo de los márgenes ventrales de las pleuras de los cinco segmentos, las cuales terminan en punta. Las espinas dorsomedianas están reducidas o desaparecidas, menos la del sexto segmento que es bien visible.
El telson (Fig. 4) tiene forma rectangular y en su borde posterior la hendidura es pequeña. Existen 6 espinas terminales y l espina lateral a cada lado. El punto más profundo de la hondidura se encuentra a nivel de la espina más externa.
La primera antena (Fig. 4) tiene tres segmentos; el segmento basal presenta el estatocisto y dos espinas. La rama externa del último segmento es de doble longitud que la rama interna. En esta antena hay varias setas plumosas.
La segunda antena consta de un protopodio provisto de una espina, un exopodio con una espina subterminal y un endopodio parecido a una varilla puntiaguda. El endopodio tiene una longitud de las tres cuartas partes del exopodio.
Los pereiópodos son birramosos y máyores en este estadio. Los pleópodos aparecen entre las expansiones de las pleuras abdominales como botones redondeados.
Las larvas encontradas y las de P. duorarum se diferencian en el estadio de segunda mysis por los siguientes caracteres:
las larvas encontradas presentan dos espinas a nivel del rostro, mientras que P. duorarum tiene un rostro liso;
los bordes laterales de las cinco primeras pleuras abdominales son muy pronunciados en las larvas encontradas, mientras que en P. duorarum no se observan esas expansiones;
el telson presenta 6 espinas terminales y l espina lateral en las larvas encontradas, mientras que en P. duorarum presenta 7 espinas terminales y l espina lateral;
la primera antena de las larvas encontradas tiene dos espinas en el segmento basal, mientras que en P. duorarum solamente presentan una. Además, las dos ramas del segmento distal son casi iguales en P. duorarum, mientras que en las larvas encontradas son de diferente longitud.
Los estadios larvales de camarones descritos previamente constituyen una serie lógica de acuerdo con las transformaciones que aparecen en su morfología. Queda por dilucidar si dicha serie corresponde a la especie P. schmitti o se trata de otra especie.
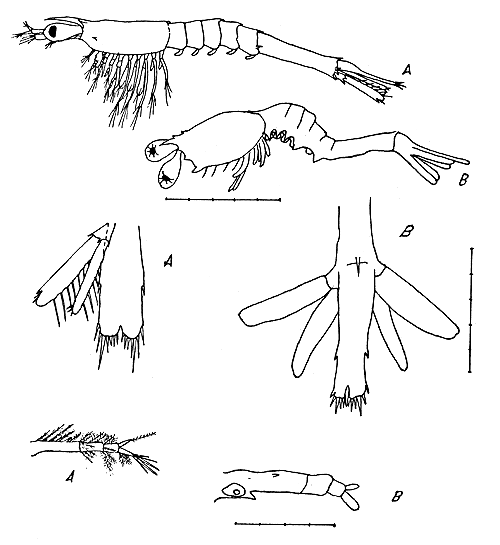
Fig. 4 Segunda mysis, vista lateral, telson, primera antena.
A) P. duorarum (según Dobkin)
B) larvas encontradas.
Pearson (1939) captura huevos en el plancton de las aguas de Florida y los cría en acuarios, obtenidendo una serie de nauplios, protozoeas y mysis, cuya morfología tiene un gran parecido con las larvas descritas en este trabajo. Sobre todo es significativo el parecido de las márgenes ventrales de las pleuras de los cinco primeros segmentos abdominales. Este autor atribuye dichos huevos a la especie Eusicyonia stimpson (Bouvier). En la subfamilia Sicyoninae se han registrado en Cuba las especies Sicyonia typica (Boeck) y Sicyonia brevirostris Stimpson (Pérez Farfante, 1954). Ambas especies tienen el exoesqueleto duro y son de menor tamaño que los camarones comerciales. Estas especies de camarones son conocidos vulgarmente como camarón reyecito y se encuentra en muy escasa cantidad en las zonas de camarón comercial. En la captura realizada durante el viaje de estudio a Tunas de Zaza no se observó en ninguna ocasión.
¿Pueden tratarse de larvas del género Sicyonia las descritas en este trabajo? Si bien nos inclinamos a pensar que las larvas encontradas pertenecen a la especie P. schmitti teniendo en cuenta la abundancia de las larvas, la de los adultos de camarón blanco y la presencia de hembras de P. schmitti próximas al momento del desove, no podemos descartar la posibilidad de que se trate de otra especie de Peneido. Debemos pensar en la probabilidad de que la captura con una red de arrastre puede no ser representativa de la fauna de una zona, ni siquiera de su fauna camaronera, que la abundancia de las larvas puede no ser proporcional a la abundancia de los adultos, y además, que cada especie puede tener su época y lugar de desove.
Es necesario tene también en cuenta otro factor. Los adultos de P. schmitti y de P. setiferus son muy cercanos morfológicamente. Sin embargo, las larvas encontradas difieren mucho de las descritas para P. setiferus (Pearson 1939; Heegaard, 1953), lo cual hace pensar que no se trate de una especie como P. schmitti.
Creemos, dados los razonamientos anteriores, que se puede dudar racionalmente de que las larvas descritas en este trabajo correspondan a P. schmitti. Esta incógnita solamente podrá ser resuelta mediante el cultivo de las larvas en condiciones de laboratorio para estar seguros de su identidad y como confirmación, obtener dichas larvas en los lugares y épocas de desove.
Para futuros trabajos el paso inmediato más importante es establecer con seguridad la identidad específica de las larvas encontradas lo cual puede ser logrado mediante el cultivo de las mismas, obtenidas en condiciones de laboratorio de hembras de P. schmitti.
El cultivo de las larvas puede servir además para la descripción de los primeros estadios larvales, es decir, los nauplios y la primera protozoea.
Un estudio completo de las muestras de plancton obtenidas en Tunas de Zaza puede servir para conocer la distribución vertical y horizontal de las larvas de camarones y señalar algún conocimiento sobre las zonas de desove y de cría de la especies comerciales.
Dobkin, S., 1961 Early developmental stages of pink shrimp, Penaeus duorarum from Florida waters. Fishery Bull.Fish.Wildl.Serv.U.S., 61(190): 321–49
Heegaard, P.E., 1953 Observations on spawning and larval history of the shrimp, Peaneus setiferus (L). Publs Inst.mar.Sci.Univ.Tex., 3(1):73–105
Pearson, J.C., 1939 The early life histories of some American Penaeidae, chiefly the commercial shrimp, Penaeus setiferus (L). Bull.U.S.Bur.Fish., 49(30):1–73
Pérez Farfante, I., 1953 Los camarones comerciales de Cuba. Mems Soc.cub.Hist.nat. “Felipe Poey”, 22(2):1–16, also issued as Contres Cent.Invest.pesq.Habana, (1):16 p.
![]()
![]()
![]()