![]()
![]()
![]()
Objective: Attempt as far as possible to identify and differentiate between the various species found (bearing in mind that some Fiji species have not yet been described), and to at least preserve voucher specimens for detailed taxonomic work later on.
Samples of likely economic seaweeds were collected where found in significant quantities, and were preserved in 5% formalin in seawater for subsequent examination. Efforts were made to collect a range of plant sizes and not just the biggest plants. This was because some lifecycle stages tend to be smaller than others and might get overlooked. In the laboratory, plants were sorted according to lifecycle stage, if fertile material could be found, by observation under a dissecting microscope. Hand sections were made of cystocarpic and tetrasporangial material, and mounted on compound-microscope slides in Karo syrup and aniline-blue stain in accordance with standard methods. Observed features were compared with descriptions in available manuscripts (in particular; South 1993a, and South 1995). Opinions were also sought about specimens from G. Robin South and Antoine N'Yeurt (Marine Studies Programme, USP), and some specimens were sent to Prof. H. Yamamoto at Hokkaido University in Japan.
For many red seaweeds, it is necessary to collect all lifecycle stages before one can positively identify the species, unfortunately some lifecycle stages only appear at certain times of the year. Some species (for example, one of the Gracilaria species found at Serua) are still unnamed because all lifecycle stages have not yet been found and so the species has remained undescribed. The taxonomy of Gracilaria is enormously difficult since many taxonomic features form continuous rather than discrete characters, and one must compile tables listing various attributes before deciding whether, upon a balance of probability, it is one species or another.
Apart from formalin-preserved plants, material was also dried and pressed for preservation as herbarium sheets. These are now lodged in the Pacific Regional Herbarium at the University of the South Pacific, under collection numbers 0893–0901 (Tailevu specimens), 0902–0907 (Suva Harbour specimens), 0938–0944 and 0965 (Serua specimens). These herbarium sheets will enable subsequent investigators to independently identify (and, if necessary, update the nomenclature for) the particular plants reported on here.
Five species of Gracilaria were found during the survey. Most abundant was G. maramae, and all lifecycle stages were found among collections from Nasese and Kaba. Tetrasporophytes were dominant, followed by cystocarpic plants. Spermatangial plants were rare. The external appearance, thallus dimensions, and spermatangial and cystocarpic cellular arrangements were all consistent with the description in South (in press).
Also present alongside G. maramae at Nasese and Kaba, but smaller and less abundant, was a so-far unidentified species of Gracilaria (Plate 1(b)). This species was also quite abundant on Cakalevu Reef. Material from Corpus Cristi and Nasese appeared to be all either tetrasporophytic or sterile plants. All lifecycle stages were present among the material collected from Kaba Peninsula (though spermatangial plants were rare). This is the first time that male gametophytes have been found for this species in Fiji, and it enabled an identification to be made by H. Yamamoto who found it to be G. edulis (Gmelin) Silva. The edulis epithet has been widely applied to specimens throughout Asia, and its use may be controversial. We will leave to subsequent investigators the issue of whether or not it is correctly applied, meanwhile care should be taken in making any comparisons between Fiji G. edulis and published results from Asia.
Gracilaria edulis has a different growth habit from G. maramae, despite often living in the same environment (Plate 4 (b)). It has more branching, but short inter-node distances, making plants smaller and bushier than G. maramae. Its colour is also different, being more of an olive or khaki hue than the more translucent and golden-looking G. maramae. G. edulis often forms a turf of tightly inter-locking plants on hard substrates, while G. maramae will be individual plants growing in sand-bottomed pools.
A third species of Gracilaria was found as two small and isolated plants, one at Muaivuso near Suva and one at Qalotu Reef near Kaba Point. These plants have a very thick thallus for a Gracilaria (5 mm) and in outward appearance resemble a small Kappaphycus plant. The plant from Muaivuso is definitely a Gracilaria judging from the structure of its cystocarps. It has been tentatively identified as G. eucheumoides.
Of the four species found at Serua, one was readily identifiable as G. maramae South, and another was the same as plants found at Kaba which were subsequently identified by H. Yamamoto as G. edulis. In addition, two other species were found that had not previously been seen in this study. One was G. arcuata Zanardini v. snackeyi Weber van Bosse (H. Yamamoto, pers. comm.), while the other has still not been identified due to insufficient material.
The identification of G. arcuata v. snackeyi was made by H. Yamamoto from tetrasporic and cystocarpic plants found in the Serua sample. However the G. edulis material from the Serua site was all either tetrasporic or sterile, so identification was made using representative material previously collected from another site (Kaba Peninsula) which included all life history stages. The fourth species, distinctive in its bright emerald-green colour and arcuate branching pattern, could not be identified and is possibly a new species (H. Yamamoto pers. comm.). Voucher specimens of the Gracilaria species found at Serua have been deposited in the South Pacific Regional Herbarium based at the University of the South Pacific, with the following catalogue numbers: G. maramae 0939, 0940; G. arcuata v. snackeyi 0943; G. edulis 0965; G. sp. 0938. The representative material for G. edulis collected from Kaba and used to help identify Serua specimens is lodged under catalogue numbers 0896, 0897 and 0898.
A species of Hypnea (Plate 2 (a)) was found at many of the sites visited, and the collected material was either sterile or tetrasporic plants. Tetraspores were arranged in a “saddle-like” fashion on the undersides of stichidia (see South 1993). This arrangement is characteristic of Hypnea pannosa (other Hypnea species have tetraspores in different arrangements, such as in rings around the stichidium). The use of the epithet “pannosa” may be in doubt for this species, but no one has yet come up with a better name (G.R. South, pers. comm.).
A single species of Acanthophora was found (Plate 2(b)), and it was quite common throughout the region surveyed. No fertile plants were found, however their morphology was consistent with the description of A. spicifera in South (1993) being brittle and bearing proliferous outgrowths of spinous branches. There are no close lookalikes in Fiji for A. spicifera to be confused with (G.R. South, pers. comm.).
A conspicuous feature of the subtidal seabed off Kaba Peninsula is the criss-crossing network of Caulerpa racemosa stolons. These had erect branches up to 100 mm tall, and bore the grape-like stalked ramelli that are a familiar sight in Fiji's municipal markets (known as “nama”). Another species present was Caulerpa taxifolia, with erect branches flattened and fern-like in shape. There are several recognised varieties of C. racemosa, and the bulk of the plants present around Kaba most closely resembled the var. lamourouxii specimens found in the Pacific Regional Herbarium collection.
Two species of Laurencia were conspicuous on rocks and in tidal pools at Kaba Peninsula, particularly towards Kaleli Settlement. These plants are small (>50 mm) and grape-like in appearance. One was quite fleshy, and found on rocks in the intertidal (an example from Nasese is shown in Plate 3(a)). The other was finer, and found in tidal pools. Both have pits in the branch apices that are similar to those in Chondria, however Chondria has a characteristic arrangement of 5 central cells in cross section which Laurencia lacks. With the current state of knowledge, it was not possible to positively assign species names to these plants, however the larger, fleshy plants resemble herbarium specimens labelled as L. patentiramea. Laurencia contains phycocolloids that have been of academic interest, but is not being commercially used as a source of phycocolloid anywhere in the world.
Species listed as present in Fiji by South and Kasahara (1992) with potential commercial value overseas, but not located during this survey, include Gelidium pusillum, Pterocladia nana, Gracilaria coronopifolia, Gracilaria lichenoides, Hypnea cervicornis, Hypnea cornuta, and Hypnea musciformis. South and Kasahara (1992) also list some additional Gracilaria (G. eucheumoides, G. furcellata) and Hypnea (H. charoides, H. divaricata, H. esperi) species which may be worthy of investigation at a future date.
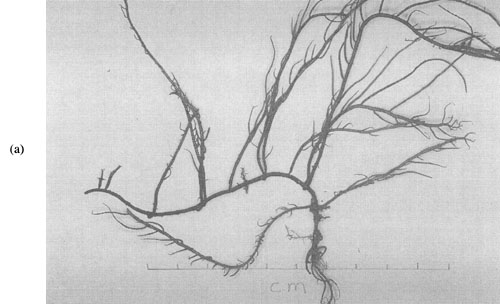

Plate 1: (a) Gracilaria maramae
(b) Gracilaria edulis
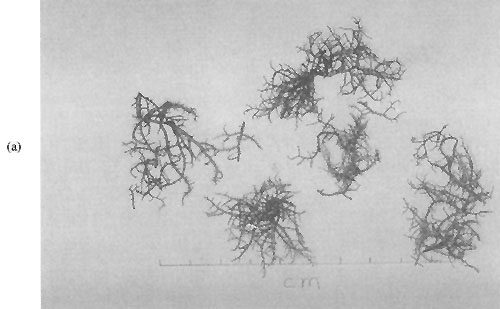

Plate 2: (a) Hypnea pannosa
(b) Acanthophora spicifera


Plate 3: (a) Laurencia sp.
(b) Gelidiella sp.
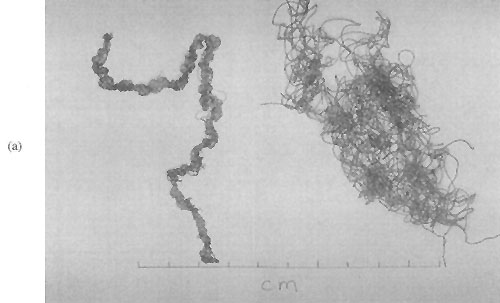

Plate 4: (a) Two species of Enteromorph
(b) Comparison of Gracilaria edulis and Gracilaria maramae growth habit
![]()
![]()
![]()