![]()
![]()
![]()
Las actividades humanas pueden producir la erosión de la variación genética y la extinción de los peces a través de varios mecanismos como los siguientes:
la contaminación y otros cambios del medio que ponen en tensión a una población y causan mortalidad diferencial, extinción o ambos;
la presión de pesca que puede favorecer algunos genotipos sobre otros;
la selección artificial y la domesticación que pueden resultar en consanguinidad y empobrecimiento genético, consciente o inconscientemente;
la introducción de especies exóticas y de enfermedades.
En esta sección los factores anteriores son recapitulados a la luz de experiencias recientes en las pesquerías y en la acuicultura.
Diferentes tipos de contaminación (por ejemplo, la eutroficación, toxinas tales como el mercurio, el DDT, PCB) han tenido efectos desastrosos en muchas comunidades ícticas. En la mayor parte de los países occidentales, la detección de toxinas en ciertas especies de peces, las ha hecho no aptas para el consumo humano. En algunos casos la legislación ha tenido éxito para revertir la situación. Por ejemplo, una gran parte del lago Vänern, Suecia (el tercer lago en tamaño en Europa) fue puesta en la lista negra, pero la situación ha mejorado por la modificación de los procedimientos de purificación del agua de los molinos de papel.
El problema más importante en Escandinavia y otros lugares, ha sido la lluvia ácida (SO2) proveniente de industrias que utilizan combustibles fósiles como fuente de energía. Por ejemplo, grandes porciones del sur de Noruega y del sudoeste de Suecia han perdido casi todos sus peces debido a lluvias ácidas originadas en Gran Bretaña y en la República Federal de Alemania. Las especies de peces difieren en su respuesta a la acidificación. El bermejuelo (Leuciscus rutilus) y la mayor parte de los salmónidos son extremadamente sensibles a la disminución del pH. Se ha intentado añadir cal como un agente de “respiración artificial” (SKr 1 millón al año, en Suecia), pero es obvio que este problema no puede ser resuelto sin algún tipo de negociación internacional. Varias poblaciones de trucha alpina se han extinguido recientemente debido a su sensibilidad. El efecto primario de la acidificación se da aparentemente en las etapas reproductivas tempranas.
La contaminación por nutrientes (especialmente PO4) provenientes de las comunidades humanas ha sido más desastrosa en las regiones densamente pobladas en las zonas templada, subtropical y tropical. A pesar de ello en las regiones árticas o alpinas con medios ambientes extremadamente oligotróficos, la eutroficación por adición de nutrientes, ha sido promovida como un método de incrementar la productividad. Esto está siendo estudiado experimentalmente en Escandinavia, así como en Canadá (Milbrink y Holmgren, 1981; Schindler y Fee, 1974).
En algunos casos, la “contaminación nutricional”, ha sido tratada exitosamente. En todos estos casos la construcción de plantas de purificación ha producido un mejoramiento significativo de la calidad del agua. En el lago Washington (EE.UU.) las vías a las zonas de desove del salmón del Pacífico han sido restauradas actualmente, y nuevamente se produce el desove natural. De la misma manera, existe una recolonización natural de las especies nativas del bajo Támesis en Inglaterra. La construcción de plantas de purificación en todas las comunidades alrededor del lago Malaren en Suecia ha hecho posible reintroducir el salmón del Báltico y la trucha parda en un curso de agua que pasa a través de Estocolmo.
La construcción de plantas de energía hidroeléctrica también ha afectado a las poblaciones de peces de lagos represados y de las porciones de ríos entre las plantas de energía. Una observación importante en las aguas escandinavas es que existen subpoblaciones de salmónidos que son afectadas de diferente modo. Por ejemplo, la trucha parda está a menudo representada por poblaciones diferentes en sitios distintos dentro de un mismo río, aparentemente con poca cruza interpoblacional. Durante la construcción de las presas, las poblaciones que desovan en la zona de la desembocadura se extinguen. La trucha alpina y el pescado blanco corégono están a menudo representados por dos o más poblaciones distintas, que son afectadas de manera diferentes. Globalmente, las poblaciones pelágicas son menos afectadas que las bentónicas.
Quedan sólo unos pocos ríos en Suecia donde se lleva a cabo el desove natural del salmón. La mayoría del salmón del Báltico proviene de las piscifactorías suecas. Las poblaciones liberadas han sido cuidadosamente seleccionadas para que representen stocks nativos originales. Sin embargo, hay evidencias de un cambio genético, tendiente a menor tamaño y maduración temprana, lo que es probablemente un efecto de la sobrepesca.
Las poblaciones continentales del salmón del lago Vänern están representadas por dos subpoblaciones que no parece se intercruzan. Estas son el salmón “Klarälven” que pasa tres años en el río Klarälven y regresa a desovar después de tres o cuatro años, y el salmón “Gullspangsälven” que pasa dos años en este río y regresa al lago después de cuatro o cinco años. Las tasas de crecimiento de las dos poblaciones son notoriamente distintas. Experimentos de marcaje de las crías procedentes de las piscifactorías han mostrado que después de 40 meses en el lago, el salmón de Gullspangsälven alcanza un peso de 5 kg, mientras que el salmón de Klarälven alcanza sólo 2.5 kg. También están separados especialmente durante su vida en el lago, sugiriendo una capacidad innata de crecimiento y de selección de habitat.
Ambas poblaciones están en peligro de extinción (figura 1), principalmente debido al desarrollo hidroeléctrico. La población de crecimiento rápido del río Gullspangsälven está muy próxima a la extinción, pero se están realizando esfuerzos para salvarla a través de la cría en piscifactorías y de su introducción en otros lagos.
La sobrepesca del salmón del Báltico ha resultado en un crecimiento más lento, tamaño menor en la madurez y regreso temprano a los sitios de reproducción. Resultados muy parecidos fueron obtenidos por Gwahaba (1973) en el caso de Tilapia nilotica en el lago George, Uganda (figura 2), quien afirmó que el tamaño restringido de las zonas de cría, las tormentas que mataban los peces y la depredación, han sido los factores de la disminución de la propagación, pero que el aumento de la pesquería comercial fue la razón principal de los cambios.
Turner (1976) menciona otro caso de modificación del ecosistema debido a la pesca intensiva de los cíclidos del lago Malawi. Mientras que en un principio la pesca estaba constituida principalmente por especies mayores, en la actualidad predominan las especies pequeñas. Este ejemplo muestra dramáticamente los efectos de la sobrepesca sobre una comunidad íctica integral. Se puede predecir que si se continúa pescando con redes no adecuadas, las especies mayores pueden estar amenzadas de extinción, en parte porque los estadios más pequeños de las especies mayores compiten en desventaja con las etapas adultas de peces pequeños de especies más abundantes.
Observaciones semejantes fueron hechas comparando poblaciones diferentes de la trucha alpina ártica, que se encuentran sujetas a diferentes tasas de explotación. La figura 3 ilustra que la población de crecimiento estimado más rápido madura antes que la de crecimiento lento, no explotada (Nilsson y Filipsson, 1971). En este caso la diferencia está sin duda determinada genéticamente (Nyman, 1972).
Desde el principio de la década de los setenta, mucha información se ha acumulado sobre las serias consecuencias de las introducciones de peces exóticos de otros lugares sobre la fauna nativa (por ejemplo, Walford y Wicklund, 1973). Por ejemplo, muchos lagos originados por desglaciación, previamente sin peces, fueron dotados de peces por el pueblo lapón en Escandinavia, y por los indios y por los colonizadores europeos en Norteamérica. A medida que la tecnología de las piscifactorías ha mejorado, la práctica se ha extendido.
La frecuencia de tales introducciones se ha incrementado en años recientes. De acuerdo a Everett (1973), la trucha arco iris (Salmo gardnieri) fue introducida en la cuenca del lago Titicaca en Perú y Bolivia en 1942. En la década de los cincuenta, las siguientes especies de trucha también fueron introducidas: S. trutta, Salvelinus fontinalis, S. namaycush y el aterínido Basilichthys bonariensis (Villwock, 1972). En 1972 dos de las especies más valiosas (y mayores) del género endémico Orestias del lago Titicaca prácticamente se extinguieron por la depredación o la competencia por alimento con las especies de truchas. Los parásitos esporozoarios que fueron introducidos pasivamente junto con las especies exóticas explican la drástica disminución de la mayoría de las especies endémicas de Orestias (Frey, 1975).
Otro ejemplo bien conocido es el del lago Lanao en la isla de Mindanao en las Filipinas. De acuerdo a Villaluz (1966: citado por Frey, 1969) personal de la Universidad Estatal de Marawi City en Mindanao, introdujo Clarias batrachus (Siluridae), Ophicephalus striatus (Ophiocephalidae) y Tilapia mossambica (Cichlidae). Aun más seria fue la simultánea introducción accidental de Glossogobius giurus (Gobiidae). De acuerdo a Frey, por lo menos algunas especies bentónicas y pelágicas del género endémico Barbodes (Puntius), se han vuelto muy escasas como lo muestra su virtual desaparición de las pescaderías locales.
Estos dos grupos de especies, el Orestias endémico del lago Titicaca y el Barbodes del lago Lanao eran los principales recursos económicos y nutricionales de los residentes. Estos no pueden ser reemplazados por las especies introducidas (Villwock, 1972) porque no son fácilmente aceptadas por la población local. Los efectos sobre los recursos genéticos de las especies endémicas de Orestias o Barbodes son irreversibles. Algunas especies están evidentemente extintas y aún aquellas que hayan podido sobrevivir probablemente se han visto disminuidas a una o pocas poblaciones pequeñas y muy dispersas.
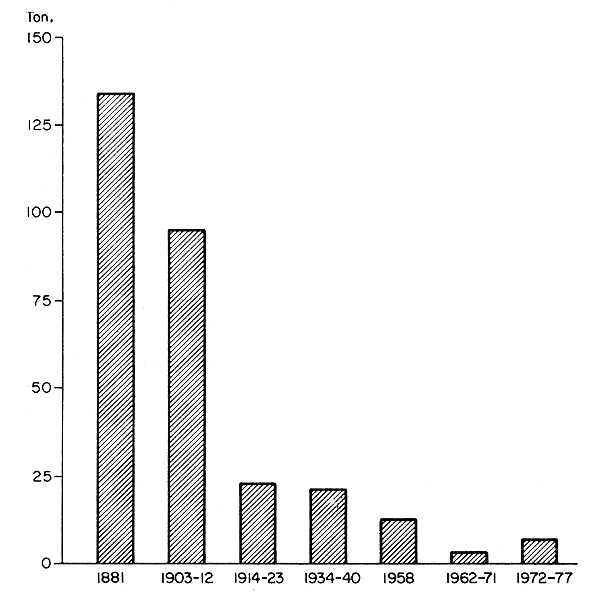
Figura 1 Capturas de salmón en las aguas continentales del Lago Vänern, Suecia, de 1881 a 1977 (Nilsson, 1979)
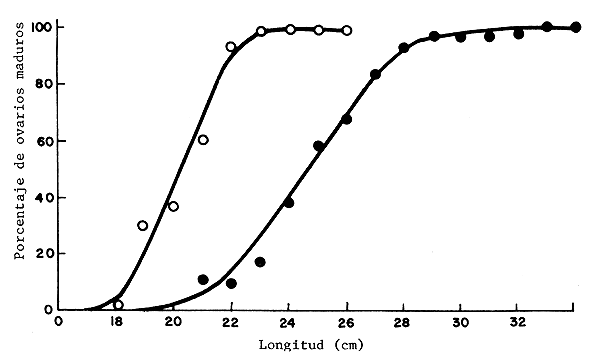
Figura 2 Porcentaje de hembras de Tilapia nilotica con ovarios maduros a diferentes longitudes. La curva de madurez para el estudio de 1971 (o) es comparado con el elaborado anteriormente por Fry y Kimsey (1960) (•)
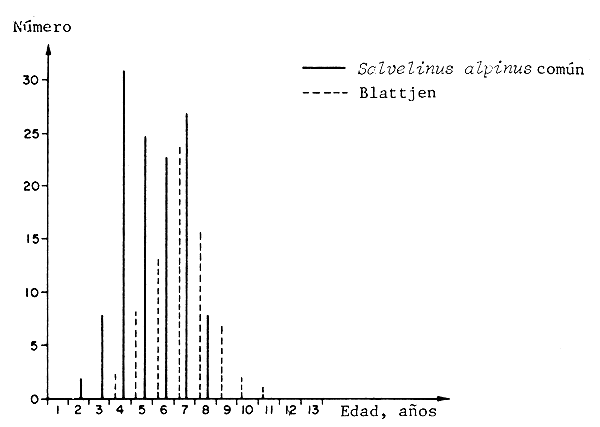
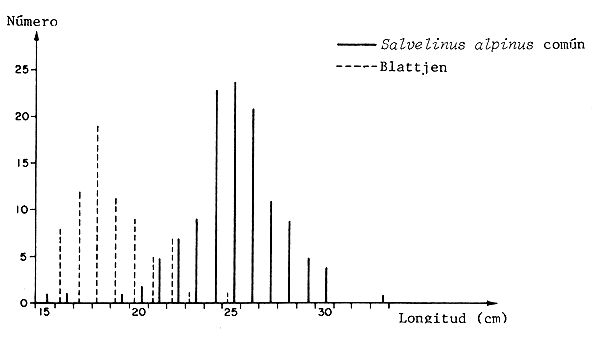
Figura 3 Diferencias en edad y longitud entre dos poblaciones de Salvelinus en madurez, en el lago Ovre Bjorkvattnet, Suecia. El Salvelinus alpinus común crece más rápidamente y es más explotado (Nilsson y Filipsson, 1971).
Los pescadores deportivos y los acuaristas han sido responsables de numerosas introducciones que han originado cambios detrimentales en la fauna nativa. Se ha demostrado en estudios llevados a cabo en Escocia y Escandinavia que las especies nativas pueden extinguirse por competencia ecológica con especies exóticas. Por ejemplo, Svärdson (1979) describió el efecto de la introducción del pescado blanco corégono y la subsecuente extinción de Salvelinus alpinus en Escandinavia. Como resultado de la competencia entre las dos formas, mostrado en la figura 4, la talla en la edad madura, edad, peso y volumen de pesca con red agallera de la trucha alpina disminuyó continuamente resultando en su virtual extinción alrededor de 1965 (Nilsson, 1967).
La hibridación entre especies nativas y especies emparentadas introducidas (Svärdson, 1979; Moyle, 1976) también ha presentado serias consecuencias en los recursos genéticos. En la segunda mitad del siglo XIX, diferentes especies de truchas fueron introducidas en California, haciendo más confusa la compleja situación de la fauna nativa de truchas (Hoopaugh, 1974: citado por Moyle, 1976). Eventos similares han sido mencionados por Moyle (1976) para especies de Gila y de Gasterosteus. El resultado más frecuente es la invasión genética de las especies nativas. El gran número de hibridaciones de que se tiene noticia en Florida fueron producidas por acuaristas o por la industria de peces de acuario (Courtenay et al., 1974). Veinte especies exóticas y cinco poblaciones híbridas se han establecido como poblaciones reproductivas.
Moyle (1974) informó acerca de las introducciones de peces en California para combatir las hierbas e insectos. Además de varias gambusias (Gasterosteus) y de dos especies de Tilapia (T. Mossambica y T. zillii), el pejerrey (Menidia aureus, Atherinidae) fue introducido ilegalmente en 1967. Desde entonces se ha convertido en la especie más abundante y extendida. Las Tilapia y las Menidia han alterado la abundancia relativa de los diferentes peces y el número total de organismos zooplanctónicos a los cuales los peces nativos estaban muy adaptados. Sin embargo, es difícil predecir las consecuencias finales para las especies nativas.
La competencia por alimento y la depredación de las larvas de especies nativas por parte de los peces exóticos han llevado a menudo a la extinción. En menos de una década las gambusias y Mollinesia, introducidas en el sur de Europa y norte de Africa, fueron responsables de la disminución de la población nativa de Aphanius sp. (Cyprinodontidae) (Villwock, 1977 y datos no publicados). De forma similar el género Cyprinodon del sistema del río Colorado está amenazado por la competencia de los peces vivíparos introducidos.
Las introducciones accidentales se han dado de varias formas. Entre estas están las redes sucias (“contaminación por huevos”) y el escape de carnada (Johannes y Larkin, 1961). Otro peligro es representado por el escape de especies exóticas de tanques de acuicultura, por ejemplo, a través de inundaciones durante la temporada de lluvias. Introducciones accidentales se han producido también a través de canales o túneles utilizados para la navegación y para plantas de energía. El caso más famoso es el de la lamprea marina (Petromyzon marinus) presente en aguas continentales interiores, que penetró en los Grandes Lagos Superiores a través del Canal Welland. Movimientos similares se han dado en el mar Mediterráneo desde la apertura del Canal de Suez.
Muchas introducciones han sido económica y estéticamente exitosas. Por ejemplo, la mayor parte del salmón norteamericano, así como la lobina negra han sido introducidos en aguas dulces europeas y varias especies de Oncorhyncus en las aguas marinas del norte de Rusia y en el Báltico. El “kokanee” (Oncorhyncus nerka) ha sido introducido en aguas dulces de Escandinavia. A pesar de que la mayor parte de estas introducciones han fallado, la lobina negra en Alemania, y la trucha arco iris en Gran Bretaña se han establecido exitosamente. La trucha parda y la carpa europea se han introducido en América del Norte, Nueva Zelandia y la mayor parte de las áreas alpinas donde los europeos han colonizado. La introducción de la trucha de lago (Salvelinus namayacush) que se inició en Fennoscandia en 1960, parece haber sido exitosa en algunas aguas suecas y finlandesas.
Se conoce poco sobre el impacto de estas introducciones en los genomas de las especies, pero donde se practican el cultivo intensivo y la domesticación, por ejemplo, de la trucha arco iris, el cambio genético es inevitable. Lo mismo debe ocurrir para la trucha parda en Europa y otros lugares.
Un estudio muy intenso de los efectos del transplante de las especies de Coregonus y Salvelinus, ha sido llevado a cabo en Suecia e Italia (Svärdson, 1979; Nilsson, 1978; Berg y Grimaldi, 1966; Nyman, 1972). Se ha demostrado que las introducciones de especies exóticas conducen a los siguientes cuatro resultados posibles:
extinción de homólogos ecológicos; el ejemplo más dramático ha sido la extinción de la trucha ártica (complejo Salvelinus alpinus) debido a la introducción de ciertos stocks de pescado blanco corégono;
hibridación con los efectos concomitantes en la genética de las poblaciones de peces originales;
fracaso de la introducción, originado en parte por la competencia con las especies establecidas;
coexistencia, lo que implica que la especie introducida ha encontrado un nicho vacante en la comunidad, con una segregación interactiva de niche, como resultado obvio.
La figura 5 ilustra el resultado normal de la introducción de especies en el norte de Escandinavia y su impacto en la comunidad zooplanctónica. Aparentemente la utilización del zooplancton como alimento tiene un papel muy importante, ya que los consumidores más eficientes de zooplancton se convierten en las especies dominantes (Nilsson y Pejler, 1973; Svärdon, 1976).
Introducir especies exóticas como organismos de presa, es en la actualidad una práctica corriente, especialmente en Estados Unidos y en Canadá, así como en Escandinavia. En Canadá y Suecia, por ejemplo, introducciones de relictos glaciales (especialmente el crustáceo Mysis relicta) han sido llevadas a cabo durante varias décadas (Furst, Bostrom y Hammar, 1978). Tales introducciones parecen haber sido exitosas en conjunto, sin embargo, en muchos casos, se ha observado una fuerte repercusión sobre la comunidad zooplanctónica nativa.
La selección artificial se ha asociado normalmente con cambios genéticos deseables. Sin embargo, en muchos casos el resultado de la selección artificial puede ser no deseable ni predecible. Esto es particularmente cierto cuando la selección y manejo de reproductores llevan a la pérdida de variación genética. Por su misma naturaleza, ejemplos específicos del empobrecimiento genético debido a tales prácticas, no están fácilmente disponibles. Sin embargo, existen suficientes antecedentes para poder aconsejar tener precaución. Por ejemplo, el rendimiento determinado genéticamente expresado en un conjunto de circunstancias (por ejemplo, en piscifactorías) puede no estar correlacionado con el rendimiento en otras circunstancias (por ejemplo, en ríos, lagos, zonas naturales). Las prácticas de selección de reproductores deben proyectarse de modo que se evite la selección y la consanguinidad inadvertidas, si la meta es liberar stocks en un medio distinto del en que fueron criados.
A este respecto, la accesibilidad de stocks reproductores a los administradores de pesquerías no asegura su superioridad genética. Por ejemplo, se ha demostrado que una carpa grande escogida de una población de individuos de idéntica edad, no necesariamente representa un genotipo superior (Wohlfarth y Moav, 1978). Tal individuo puede haber sido expuesto a un conjunto favorable de condiciones medioambientales que amplifiquen una pequeña diferencia de tamaño de origen no genético, en una talla superior. El uso de stocks reproductores compuestos por tales individuos conduciría a un progreso genético muy reducido o nulo, y bien podría llevar, si el número de individuos seleccionados es pequeño, a la consanguinidad y a la deriva génica (sección 3). Además, individuos de gran talla podrían presentar fenotipos conductuales agresivos bajo control genético, y la selección de tales individuos podría llevar a la población a un nivel de agresividad no deseable.
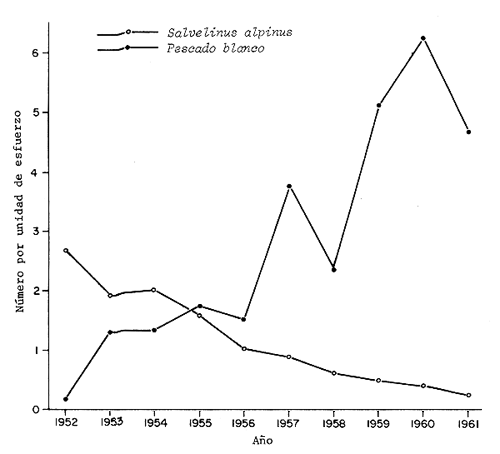 | 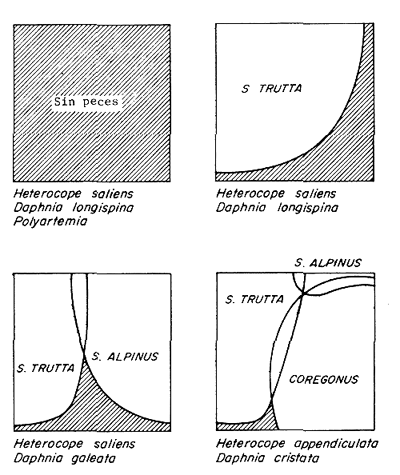 | ||
| Figura 4 | Disminución de la captura con red de enmalle de Salvelinus alpinus a medida que el pescado blanco (Coregonus sp.) introducido, aumentaba. Lago Vastansjo, norte de Suecia. (Svärdson, 1976). | Figura 5 | Modelo de las dimensiones de los nichos de la trucha parda (Salmo trutta), de Salvelinus alpinus y pescado blanco (Coregonus sp.) en alopatría y simpatría, y las especies de zooplancton dominantes. (De acuerdo a Nilsson y Pejler, 1973). |
En algunos casos el stock de reproductores seleccionado de unidades de cultivo en las cuales las clases anuales estén mezcladas (como es probable en tanques tropicales no drenados), puede presentar una mezcla de genotipos superiores e inferiores. Consecuentemente poco progreso genético, o aún erosión, pueden ser esperados por la selección fortuita de reproductores. Por ejemplo, cuando se utilicen como reproductores hembras grávidas de tanques no drenados pescadas con red, ellas pueden ser tanto ejemplares jóvenes de talla (de genotipo superior) como ejemplares viejos de talla pequeña (de genotipo inferior). Además, podría suceder que los machos de genotipo inferior por ejemplo crecimiento lento pero que “no caen en la red”) se reproduzcan más a menudo con estas hembras.
Esta es una de las razones por las que los cultivos de Tilapia en tanques ha fallado en algunas zonas trópicales: al cosechar los tanques de Tilapia es frecuente que se seleccionen los peces grandes y se regresen los pequeños. Este procedimiento favorece la reproducción de peces pequeños y a crecimiento lento, y según se ha demostrado experimentalmente, tal selección puede producir un cambio genético en la población hacia peces de talla menor (y menos deseables). El problema de la Tilapia se complica por el cuidado bucal durante la crianza y por la defensa de los juveniles hasta que tengan un tamaño que los libere del canibalismo. Los resultados son tanques sobrepoblados con individuos enanos. En canibalismo puede ser ventajoso genética y económicamente en algunos sistemas de acuicultura.
La domesticación se define como un proceso de selección genética mediado por aislamiento geográfico y reproductivo, por consanguinidad y tamaño de población pequeño para producir cambios evolutivos profundos y deseables en un stock genético. La domesticación involucra, al menos, la capacidad para vivir la mayor parte del ciclo de vida en condiciones artificiales. Se espera que incluya además adaptaciones genéticas al hacinamiento, manipulación y dietas artificales. Normalmente también se sobreentiende un mejoramiento genético hacia un crecimiento mayor, modificación de la forma del cuerpo y disminución de la conducta de huída producida por el temor. Los aspectos antropológicos de la domesticación están bien documentados (Zeuner, 1963), pero la atención de los científicos no se ha centrado en el proceso mismo a causa de las limitadas oportunidades, para estudiar, documentar y controlar los cambios fenotípicos y genotípicos que se han dado en la domesticación del ganado, las aves o animales de compañía. Lo contrario sucede en la acuicultura.
Existen probablemente muchas razones por las que el hombre no ha domesticado tantos peces como mamíferos, aves y plantas. A causa de las dificultades del transporte terrestre de peces vivos, la domesticación pudo sólo haberse dado en cultivos sedentarios. Existe, de hecho, evidencia de cultivos de peces en Sumeria, contemporáneos con las etapas más primitivas del desarrollo agrícola, que han precedido los inicios documentados de la acuicultura en China en 600 A.C.
También es probable que los intentos para aplicar a los peces o vertebrados poiquilotermos, los criterios de selección aprendidos en la cría de animales de ganado y en el cultivo de plantas, hayan sido generalmente contraproducentes. Por la razón que sea, y con la notoria excepción de la carpa dorada ornamental, y tal vez de la carpa común, pocos peces pueden considerarse domesticados aun cuando algunas cepas de truchas, por ejemplo, claramente están mucho mejor adaptadas a las condiciones de cultivo en piscifactorías que sus contrapartes silvestres (Moyle, 1969; Hines, 1976). Otras especies como las carpas chinas, carpas indias, Tilapia sp. y los bagres americanos están en proceso de domesticación.
Como se hizo notar en la sección 5.4, las mismas condiciones de cultivo, junto con la selección consciente hecha por el hombre, facilitarán la domesticación. En gran manera los resultados de este proceso no sólo son deseables, sino también predecibles. Los investigadores deben recordar, sin embargo, que la historia raramente informa de fracasos en tales intentos. A este respecto los investigadores, acuicultores y criaderos deberían situarse en la perspectiva que estos esfuerzos representan, en muchos casos, los pasos iniciales de la domesticación incipiente de un nuevo grupo animal. Debe diseñarse un estudio científico para monitorear y documentar este proceso.
Se ha reconocido que debido a la actuación inevitable de la selección natural en los medios ambientes de cultivo (sección 5.4) la propagación sin domesticación es, tal vez, imposible. Sin embargo, en algunas circunstancias la cría sin cambio genético es la meta, como en el caso de los programas de refuerzo de los stocks de pesquerías naturales, en los cuales es deseable preservar los genotipos originales no domesticados. Por ello se debe tener precaución para evitar el desarrollo de los rasgos que históricamente han caracterizado la domesticación (por ejemplo, pérdida del temor por los depredadores, de la agresividad, de la irritabilidad). Tales pérdidas pueden resultar muy útiles en la administración de stocks de reproductores porque estos rasgos resultan en una mayor accesibilidad. Sin embargo, el desarrollo de dichos rasgos irá en contra de las metas del refuerzo de stocks en pesquerías naturales.
En algunos casos de refuerzo de stocks, los cambios conductuales genéticos producidos durante la domesticación son útiles y deseables. Por ejemplo, el anegamiento de ciertas zonas en proyectos de recuperación de costas o en reservorios continentales pueden crear grandes zonas de hábitats seminaturales acuáticos útiles para la acuicultura. En tales situaciones, puede ser deseable la pérdida de hábitos migratorios y otras conductas sociales no apropiadas.
Puede esperarse que, en base al conocimiento científico ahora existente, se logre una domesticación relativamente rápida de las especies acuáticas, tal como ha sido sugerido por Moav, Brody y Hulata, (1978). Técnicas tales como la ginogénesis probablemente faciliten notoriamente el proceso de domesticación. La fecundidad normalmente alta de las especies ícticas ha sido mencionado como un factor que contribuye a la domesticación rápida. Ya sea así o no, es ciertamente fundamental, como fue hecho notar en las recomendaciones de la Conferencia Técnica de la FAO sobre la Acuicultura en 1976, llevada a cabo en Kyoto, que los criadores de peces proporcionen una documentación muy cuidadosa del historial de la crianza de las líneas que se están domesticando. Esta documentación, como es reconocido por los criadores de mamíferos y por los agricultores, será de gran importancia cuando después sea deseable volver a obtener genes, por ejemplo, de resistencia a enfermedades, que hayan sido perdidos en el proceso de domesticación. También se ha propuesto (Malecha, Sarver y Onizuka, 1980) que existe la oportunidad de estudiar las etapas tempranas del proceso mismo de domesticación, a través de la domesticación de especies acuáticas, de una manera que ya no resulta posible en el ganado.
![]()
![]()
![]()