![]()
![]()
![]()
by
V.A. Shevchenko
Institute of General Genetics
Academy of Sciences
Moscow, U.S.S.R.
Mutational variability results in hereditary changes, serving as a starting point for evolution. Useful mutations are picked up by selection and, after a certain number of generations, affect the entire population. Neutral mutations occur in nature as a function of forward and reverse mutation frequencies. Detrimental mutations forced into the population by mutation pressure are gradually eliminated from the population. The majority of mutations appearing in natural populations can be regarded as detrimental changes. Natural selection clears the populations of detrimental mutations and takes care of the adaptive features of a species. Genetic death, affecting in each generation a part of the population, saves the species from a dangerous accumulation of mutations.
Selection is not a fortuitous (accidental) reproduction of genotypes. The life-flow of a population could be regarded as consisting of particles, continuously changing, dividing, merging and disappearing, flowing from the past into the future through a number of extremely complicated sieves. Selection can be said to have occurred if the difference in the composition of the flow in question in two successive sections is too great to be attributed to selection error (drift) or a mutation.
In trying to understand selection effect, one must be sure that the section of the flow under investigation does not ramify (no emigration) and does not take tributaries (no immigration).
If a population is genetically heterogeneous, the additions will be in favour of successful reproduction of some genotypes rather than others. Thus, certain types of genetic information will occur in the gene-pool of a population more and more often, and other types more and more seldom. The gene frequency p and q will be a function of time instead of remaining constant as might be expected under the Hardy-Weinberg equilibrium conditions. When a non-accidental selected group of genotypes produces more offspring than other types, selection is said to have occurred. In a broad sense selection reduces the variety of living beings; those containing certain types and combinations of genetic information are not viable or do not produce offspring. It is this process that is responsible for what we perceive as a certain order in nature. Only in a limited sense, in the sense of counteracting the tendency for enthropy increase, is selection a creative factor.
There are two types of selection: natural selection, whereby the environment determines which genotypes are most adapted, and artificial selection, whereby this is determined by man. It is noteworthy that while natural selection must act through the phenotype, artificial selection may sometimes have an immediate effect on the genotype.
The term “struggle for existence” is relevant only in Darwin's broad metaphorical interpretation: "involving the dependence of one being upon another and involving, above all not only the survival of one particular species, but prospects of reproduction as well (Darwin, 1867). Struggle for existence is always associated with extermination and failure to reproduce. This side of the struggle for existence in modern literature is termed “elimination”. Another important manifestation of the struggle for existence is competition between organisms struggling for living and reproductive conditions.
In the former aspect of the struggle for existence notion the stress is laid on the organism, its occurrence and taxonomic relations. In this case the structure and functions of the organism are regarded as function of the whole set of living conditions comprising the environment. In the latter aspect the stress is laid on eliminating factors, at the same time the analysis of the environment factors is performed, determining the survival and reproduction factors of the organism in question.
In the different types of competition the positive characteristic of the struggle for existence is emphasized. According to Schmalhausen (1946), one should differentiate between intragroup (individual and interfamily) competition and intergroup competition. Intragroup competition is characterized by differences between the individuals of one population, in their struggle for living and reproduction conditions and for their own survival, as well as that of the offspring. Intergroup competition occurs between individuals that are members of different populations, bound by common origin and reproduction. The negative side of the struggle for existence is manifested in their direct or indirect extermination and failure to reproduce. The struggle for existence is always associated with the elimination of the least adapted; and only through it does it lead to natural selection. Elimination is an integral part of all forms of struggle for existence. There is a great number of various forms of elimination, characterized by the eliminating factors (physical, physiological and biological) as well as the objects of elimination (individuals, families, populations) their age (age elimination) and the nature of elimination (general, spontaneous, random, permanent, recurrent, seasonal).
There are three main types of natural selection in populations - directional, stabilizing and disruptive. Directional selection means a selection whereby there is a shift in the average value of the character under investigation. Stabilizing selection manifests itself in a reduced adapatability of extreme variants accompanied by a reduction in the dispersion of a given character, which results in the population being more homogeneous. Disruptive selection occurs when the conditions are favourable for two or more different types and unfavourable for the intermediate types. This type of selection usually increases dispersion and under certain conditions may lead to population splitting. Directional selection at its simplest may be represented by a chart (Fig.1). The first phase (A) represents a typical variation curve characterizing a known trait of all individuals in a population. The second curve characterizes the variability of the same trait in individuals surviving and producing offspring (the reproducing part of the population). The area between the two curves represents the zone of variations, which are eliminated in the struggle for existence.
When the environment changes in a certain direction, shifting the adaptability of the organism along the given lines, some individuals formerly within adaptative limits will find themselves less adapted, and some others, formerly eliminated, may gain certain advantages. As a result, at one side of the variation curve (Fig.1 B) there is a reduction in the number of the surviving quota and a respective widening of the eliminated variations zone; on the other side is an expansion of the quota, accounted for by the variations zone, formerly eliminated. The survival curve becomes asymetric and the mean shifts in the favourable direction. The accumulation of variations and, consequently, mutations, formerly eliminated, results in the expansion of the variation curve outside the limits observed hitherto (Fig.1 C). This process is slower than the first phase of the evolution (Fig.1 B). However, in favourable conditions the evolution becomes self-accelerating. The majority of data proving directly or indirectly the existence of natural selection conform to this simple form.
Stabilizing selection leads to increased stability of the quota through the elimination of all accidental fluctuations. During the change of environment factors or during mutation the stability of form generating systems falters. During evolution the form generating systems are to a certain extent restored due to the continuous effect of stabilizing selection.
The mechanism of stabilizing selection can be analyzed by means of a usual variation curve (Fig.2). The elimination of all ontogenetic fluctuations (shown under the abscissa) leading to the development of extreme variants is connected with the selection and accumulation through a number of generations of minor mutations, whose phenotypic expression under normal development conditions does not exceed an established conventional norm.
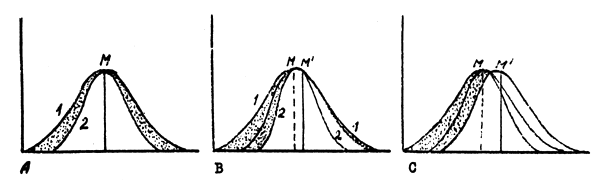
Fig.1 Chart of the trend selection mechanism
A. Selection under stabilized conditions
B.C. Stages in the process of selection with conditions changing in a certain direction
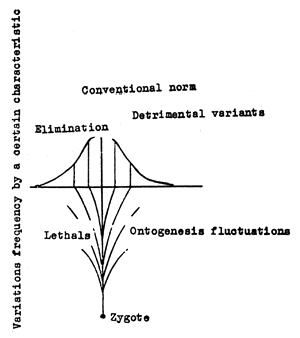
Fig.2 Chart of the stabilizing selection mechanism.
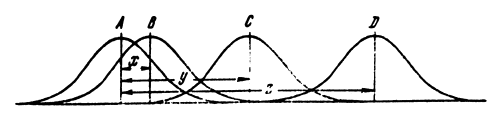
Fig.3 Chart of the different degrees of population divergency. Variability in populations may partly overlap (A,B and C) or be completely separate (A and D).
Under specific evolutionary conditions, the directional and stabilizing effects of natural selection manifest themselves simultaneously. On the one hand, there is a stabilization of characteristics that are becoming positive under the given fluctuations of environmental conditions, as well as other factors, influencing the quota. On the other hand, gradual changes of the same factors in certain directions introduce new requirements and cause the respective changes of the norm to take place.
Disruptive selection in its simplest form is shown in Fig.3. Changes in the genetic structure of populations may disrupt the population completely (there are no common genotypes) or just shift the genotype distribution to various degrees (there are a number of common genotypes). Fig.3 shows four populations at different stages of genetic divergency. Populations with an average A and B are only slightly shifted as regards one another. The differences of the average of these populations are designated by the X segment. The curves, characterizing genotype distribution in these populations overlap in large segments. The overlap of the C and A curves is not so great. The differences between the averages in these populations are designated by the y segment. The D curve is completely disconnected from the A curve, and their differences are designated by the Z segment.
The effectiveness of selection is associated with the degree of characteristic inheritance. In certain cases this primarily concerns the crop capacity and productiveness, quantitatively dependent on hereditary peculiarities as well as the positive effect of environment.
Part of the variability due to modifications is not inherited by the next generation. For example, if a given character is only 20 percent inheritable, then obviously selection for this characteristic will be much weaker as compared to a character that is 100 percent inheritable. The differences in the selection effectiveness with 100 percent heredity of a character and with the obscuring effect of the adequate modificational variability are shown in Fig.4, which shows the data of the offspring selection from two initial phenotypically identical populations. The left half represents the selection process with a 100 percent heritability of the characteristic, while the right half shows 20 percent heritability. We can see a striking difference between the two selections within only four generations.
For each given environment the product of the relative fitness of a given genotype by its relative reproduction potential is adaptability or fitness of the genotype. Fitness comprises of relative productivity, length of the reproductive period, capacity to find a mate, general endurance of the specimens of a given genotype, etc.
Fitness is conventionally expressed by values from O (adaptive value of the zygote, homozygous by the lethal gene) to 1 (adaptive value of the genotype producing maximum gametes taking part in the reproduction of the next generation). Thus fitness should be understood as genotype effectiveness in passing genetic information on to the next generation. To compare the genotype effectiveness a scale is used where O represents absence of information transmission and 1 represents the ability to transmit all information of the genotype. The creative aspect of selection is almost completely connected with the environment, which by means of selection affects the genetic structure in such a way as to result in the origin of adapted phenotypes.
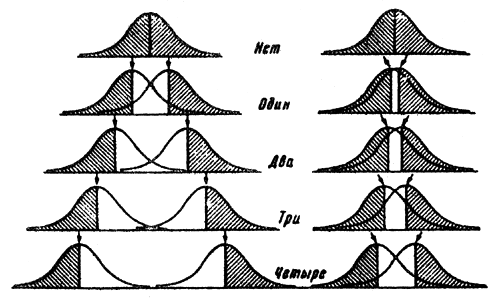 | Fig.4 Selection effectiveness in populations with complete heritability of a characteristic (left) and 20% fluctuation in heritability. |
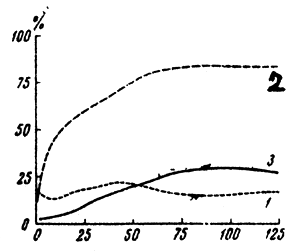 | Fig.5 The accumulation of recessive lethals on the second chromosomes in experimental populations of Drosophila melanogaster. |
1. Populations radiated by a large dosage at the beginning of the experiment. 2. Populations radiated in each generation for 126 generations. 3. Control population, not radiated. | |
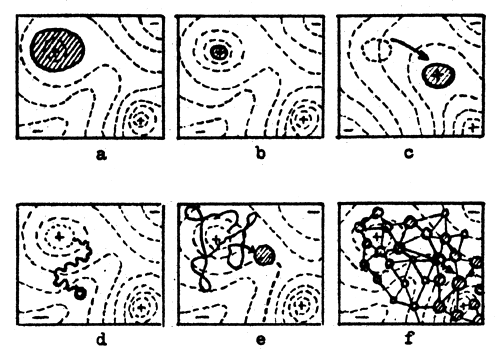 | Fig.6 Fields of gene combinations occupied by different types of populations under different conditions. |
A lot of work has been done to describe in terms of mathematics the various selection types. If we have a number of genotypes (AA, Aa and aa) we can, by assigning to each a certain fitness W, carry out a quantitative estimation of their ability to pass their genes on to the gene pool of the next generation. The genotype with maximum adaptability will have W=1, whereas less adaptive combinations will be designated by smaller values of W. Thus, the following situation is possible:
| genotype fitness | AA 1 | Aa 1 | aa 1 - S |
In this case, homozygous recessives are inferior in their adaptability to heterozygotes and to dominant homozygotes. The degree of disadvantage of this genotype is determined by the selection factor S and its fitness is expressed as 1 - S. The selection factor may vary from 0 (no negative selection) to 1 (recessive homozygotes fail to survive). The following is an analysis of an example where homozygous recessives completely fail to survive (S = 1).
The proportion of gene frequency in any two generations may be determined from
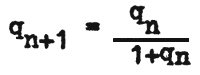
where the indices represent ordinal numbers of generations.
Thus
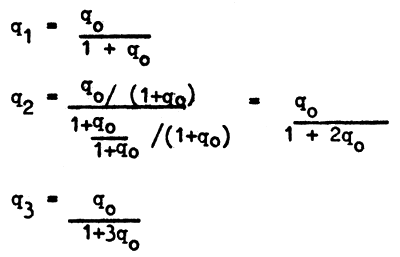
these successive values q (gene frequencies) form a harmonic series, i.e., a series whose elements are inverse to elements of arithmetic progression. With the initial gene frequency known, the frequency for any subsequent generation can be found from
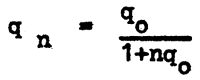
The gene frequency balance in the first generation is designated as ∆ q and can be found from
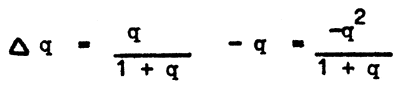
In this case the rate of variation is a function of gene frequency. When the gene frequency is high, the gene is soon eliminated from the population. The reason for such a rate of change is that with an increase in the frequency of a given gene the share of recessive genes in heterozygous condition rapidly increases. In heterozygotes, recessive genes are protected from selection effect, as they are lethal only in homozygous condition. The lower the gene frequency is the smaller the share of recessive genes subject to elimination in homozygotes, and accordingly, the slower the process of elimination of the gene from the population. This can be clearly seen from the table below:
| Gene frequency | Frequency drop within one generation |
| 0.9 | 0.426 |
| 0.5 | 0.167 |
| 0.1 | 0.009 |
| 0.05 | 0.0024 |
| 0.01 | 0.000099 |
Selection against dominant genes is relatively effective. For example, lethal dominant genes are eliminated from the population within one generation.
A situation where dominant specimens have advantages over recessive ones, but whereby homozygous recessives, nevertheless, contribute to the gene pool of a subsequent generation, occurs more often than complete lethality of homozygous recessives.
If we assume the adaptation value of both dominant forms (AA and aa) to be 1, and that of homozygous recessives to be 1 - S, then with a selection effective of 1, we shall have p2 for AA specimens, 2pq for Aa specimens and q2 - Sq2 of aa specimens from the total of p2+2pq+q2-Sq2 is equal to 1 1-Sq2 specimens. Using the same procedure as in the previous instance we shall have the gene frequency balance for one generation equal to
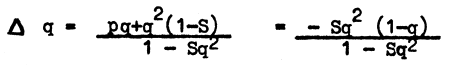
Thus, under these conditions, the gene frequency change will be small when q is very high or very low, and it will be comparatively high with average q values. When q is high, the process will be slow due to a comparatively high contribution of recessive homozygotes to gene reproduction. When q becomes small, the protective action of the heterozygote dominant phenotype slows down the process - as in the case of lethality of homozygous recessives.
In a situation where both homozygous genotypes are inferior to heterozygotes in their adaptability value (fitness), a given locus is said to be over dominant. This situation may be described thus:
| Genotype | AA | Aa | aa |
| Adaptability value | 1 - SA | 1 | 1 - Sa |
q change within one generation will finally become
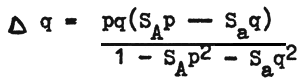
Consequently SAp > Saq , q will be positive as the gene frequency a increase. With SAp < Saq, then ∆ q=0 and the gene frequency a is in equilibrium. In this case the equilibrium value of q is determined only by the selection factors SA Sa.
If the adaptability values of all the three genotypes remain constant, the equivalent gene frequency value will also be constant. If due to any cause the gene frequency is shifted from the equilibrium point, selection will restore the equilibrium. That is why this type of equilibrium is called stable.
Polymorphism is the occurrence in the same population of several distinct forms. Usually the term is applied only to cases where the rarer form is maintained by some mechanism other than mutation.
In a kind of salmon spawning in Northern rivers, there are two types of males. Some of the males, after migrating to the sea, stay there and grow to lengths of 100 cm. They arrive in the spawning area together with females at the age of 5–7 years. Other males are dwarfs, not longer than 15 cm, and reach puberty at the age of 2–3 years. These males live only in the river. Together with their sea brethren they take part in the fertilization of spawn and reproduce both normal offspring that descend to the sea to grow and return for spawning after intensive growth. It is believed that the presence of dwarf males is characteristic of populations where the sex ratio is out of balance when the fish return for spawning. Thus their existence provides the necessary sex ratio for normal reproduction.
Polymorphism occurs, among other things, when heterozygotes are superior to both homozygous forms. In such cases the gene frequency equilibrium results in balanced polymorphism. This type of polymorphism is very important for evolution as it allows the variability in a population to be kept at a definite level. Due to balanced polymorphism a population may be able to respond promptly to environmental changes.
Another mechanism, slowing down loss of variability in any locus is called dominance, whereby it becomes possible to mask undesirable mutations in heterozygotes. However, judging from a different angle, both these phenomena could be said to contribute to keeping unwanted genes in a population. To understand this point of view the question of genetic load should be considered.
The situation whereby various recessive mutations, disguised as normal phenotypes, can accumulate in populations was first demonstrated by Tschetverikoff (1926) with heterozygous species. His study revealed the existence in natural populations of a great variety of detrimental mutations existing in the heterozygous state. Since these mutations are segregated from populations in the homozygous state in accordance with the Hardy-Weinberg law, populations not only carry detrimental genes in homozygotes, but also homozygous individuals are sacrificed. This served as the foundation for the genetic load theory. The principal notion in the genetic load theory is the notion of genetic death. Genetic death should be understood as failure to survive, caused by negative mutations, which, by being removed from the homozygote, arrest development and result in the death of the zygote or of the developing organism.
The genes arresting development to a considerable degree were termed semi-lethal mutations, the ones whose detrimental influence is insignificant were termed sub-vital. While studying the natural populations of the Drosophila, it was demonstrated that each chromosome actually contains one, two or even a few detrimental mutations.
If at a certain time one genotype in a population has an advantage over the others, it can be assumed that a population comprising of only that genotype would have the highest fitness possible. If we assume the fitness of the fittest genotype to be W = 1.0 then the adaptability of the whole population W will also be 1. as W=∑ Wiqi (where Wi = the fitness of the i-th genotype, qi = frequency of the first genotype, i = any numeral from 1 to n and n = the number of different genotypes. According to this model, any composite genotype population will be less adapted than an ideal monomorphic population. The difference between the factual and ideal adaptability is the genetic load L.
L = log n W or W = e-L, that is L is the average of potential deaths for one individual and W is the genetic survival potential, the possibility for an individual to survive despite the properties of its genome. Assuming this model, two extreme cases can be visualized. In one, the ideal genotype is homozygous in all its loci. In the other all loci may be overdominant as regards fitness, and the multiple heterozygote will be the ideal genotype. There is evidence to the effect that overdominance is of primary importance for adaptability in animals. At the same time quite a number of plants have genetic systems providing near homozygosis. Thus both possibilities to form ideal genotypes do occur in nature.
In a population existing under stable conditions, the genetic contents in general and the genetic load in particular are stabilized at a certain level. In the stable phase it is impossible to study regularities in the motion of mutations with different viability in populations.
The principal regularities of the genetic load dynamics in a population are discovered by artificially changing the genetic load with the help of mutagenic actions. An especially detailed study of the genetic load dynamics was carried out after ionizing radiation treatment of populations. The first study of this kind was accomplished by Wallace (1956). Wallace demonstrated that after radiating, populations of Drosophila in laboratory conditions showed a great number of mutations, reducing the specimens' viability to various extents. These mutations gradually being eliminated from the populations, exceeded the control level in 50–70 generations. The findings were corraborated during the study of radiation effect on Drosophila populations in natural conditions. Among other things, during the study of the effects of the Bikini island thermonuclear explosions on the genetic properties of Drosophila populations, radiation effects were registered only a few years after the explosions. Thus, the induced genetic load cannot be eliminated by selection within one or several generations, but remains in the population for a very long time (scores of generations) affecting its viability.
With permanent radiation, the genetic load of populations increases. Fig.5 shows the amounts of second chromosomes with lethal mutations in Drosophila accumulating in populations no.2 for as many as 126 generations after the experiment commencement. Control population no.3, originally devoid of lethal and semilethal mutations, was influenced by the natural mutation process and thus acquired more and more lethal mutations. However, the process stabilized at 25–30 percent as there began the segregation of homozygotes and consequent suppression of lethals. This state of equilibrium of selection and mutation, reproduced under experimental conditions, is a fair model of the populations structure in the majority of animals and plants.
In population no.2, the level of the total lethal concentrations is considerably higher than in the control; it reaches equilibrium only in 80 percent of lethal chromosomes. (Fig.5, generations 75–126). In this case equilibrium is noticeably shifted towards high concentrations of lethals owing to each Drosophila generation being radiated by a dose of 2 000 R.
After radiating only one initial generation (population 1 Fig.5) there is a drop in the number of lethals which appeared as a result of radiation, and then a few generations later the genetic load of this population stabilized at a lower level (generations 60–126, Fig.5).
According to Wallace, species which are heterozygous in recessive lethal mutations are quite viable. He demonstrated that viability of individuals in continuously irradiated population (no.2) is only slightly lower than that of the control individuals. The average viability of specimens in population no.2 was 98 if that of the specimens from population no.3 is assumed to be 100.
Thus, even considerable dosages of radiation do not result in the death of the Drosophila populations. The populations continuously radiated at 2 000 R maintain a high adaptation level, thanks to selection. However, the survival and steady reproduction of such a population is associated with high mortality caused by mutational pressure. With a considerable part of a population continuously dying out, it is only the high reproduction potential that allows selection to maintain the high viability of the population. Naturally, such a situation would not avail in animals with a limited number of individuals or with considerable longevity.
The genetic load constantly remaining in populations under variable environment conditions may be transformed into hereditary components, which are picked up by natural selection processes, and is the object of progressive evolution. In a number of cases this provides high rates of evolution processes. A classic example is the melanism mutation in butterflies. Due to industrialization of towns and provinces, soot and smoke caused the landscape to darken; as a result, the black variety, due to their physiological peculiarities maintained in populations by balanced polymorphism, has become dominant in certain butterfly species within the last generation due to extensive selection.
All this goes to show that a change in environment results in genes, formerly maintained at the mutation or balanced genetic load level, acquiring the properties of useful mutations and beginning to be subjected to trend selection. Selection increases the level of mutations concentration to such an extent as to enable the population's survival in the new conditions. The normal allele formerly dominating in the population is suppressed and moves to the genetic load category; whereas the mutation, formerly an element of genetic load, becomes one of the adaptative properties of the population's genotype.
In organisms with sexual reproduction a great number of gene combinations is found. In a species having as few as 1 000 loci, each represented by a series of 10 alleles, various genotypes might be obtained by recombination (101000). This figure is far greater than the number of electrons in the Universe. It is obvious that such a species would have a great potential of possible genotypic variation. Among this vast number of feasible combinations there should be a great number of highly adaptive combinations, alongside a considerable number of unfavourable and even lethal combinations. Wright (1931) represented this multitude as a two dimensional trace chart with adaptive peaks and non-adaptive hollows and said that the problem of evolution is the problem of a mechanism whereby a species could find a way from lower peaks to higher ones.
Fig.6 shows several instances of populations of certain types in the adaptability field under various conditions. This field is represented as a topographic chart where isolines correspond to various adaptation levels. The bold trace line shows the initial positions of populations and the arrows show the directions of a subsequent change.
Fig.6-a shows the result of a mutation frequency increase, or a decrease of the average adaptability of a population and a general adequate widening of the variability range. Part of a population may occupy lower slopes of another adaptation type, higher than the original. If this takes place, the whole population will begin to shift, to move to another peak.
With the selection effect weakened, the significance of mutation processes in the change of a population's genetic structure considerably increases. The pressure of negative mutation may function as a purgative in historical degeneration of traits no longer important. In this case, uncontrolled accumulation of negative mutations leads to the degeneration (disappearance) of the organ.
An example of such types of evolution caused by mutational pressure is the reduction of eyes and pigmentation in cave animals, the reduction of many organs in vermin and the degeneration of wings in birds having no need to fly.
A more complicated situation may be observed when a whole population with quite a number of adaptative traits finds itself under conditions where the effect of natural selection is reduced. In this case the process of mutation begins to ruin the adaptation basis of the population as a whole, due to the accumulation of negative mutations.
Among the most illustrative examples of such a trend in mutation processes is the example of populations of man. The consequences of selection increase and the mutation pressure drop may be found in Fig.6-b. Here the average adaptation level increases due to the drop in evolutional flexibility. The chances for conquering the next peak have dropped.
Fig.6-c shows the consequences of profound environmental changes. Here adaptive peaks become hollows and vice versa. The fate of a species depends on selection extensiveness and the rate of environmental change. A species occupying a small field, when pressed by selection, may prove not to be variable enough to pass on to the newly formed peaks; thus it may remain in the hollow and degenerate. Under less extensive selection conditions, populations will just float as the environment changes.
Fig.6-d shows the effect of a considerable reduction in the population number and close inbreeding. As a result of accidental fixation and a loss of genes the population descends in a curvilinear manner from its adaptive peak and inbreeding reduces the adaptive field occupied by the population. This process eventually leads to degeneration.
Fig.6-e shows the consequences of intermediate relation between the number of individuals in a population and mutation rate. The population descends from the peak and begins to float, keeping, however, close to it. This is a situation of experiment and error, which may lead to conquering new and higher peaks; but under the above conditions the process would be painfully slow.
In Fig.6-f a species with a considerable area is sub-divided into numerous semi-isolated populations. The area, occupied by each of the sub-populations shifts at a high rate, mainly in a non-adaptive manner and much faster than in the former case (as the shifting is determined not by mutation frequency, but by the rate of intrapopulational migration). Under these conditions a sub-population may find itself in the vicinity of a higher peak and climb it. In that case, this sub-population will increase its number and gradually the whole species will move to the new peak's zone of influence. This inter-group selection allows one to experiment with new gene combinations with a hazard for the species lower than in the case of intra-group selection. A divided population like this presents by far the best conditions for evolutional changes, associated with a considerably smaller risk.
So far we have been discussing processes causing the appearance, inheritance and maintenance of differences in a population. Now we shall analyse the processes resulting in differences and the appearance in a species of new forms, races, sub-species; i.e., we shall discuss the initial steps of speciation. Species are the basic actual individualized categories into which falls the life flow on earth. The evolution of populations leads to speciation. Completed with the appearance of a new species, the evolution creates the species genotype as an integral coadapted system of genes, meeting all the adaptation requirements.
The process of speciation, associated with the distance separation of populations, is termed allopatric speciation, whereas the appearance of new forms without distance separation is termed sympatric speciation. Some investigators suppose that allopatric speciation is the principal way of speciation. For example, out of 1 200 species of fish to the east and west of the Panama Isthmus, which by geological standards was formed recently, only 6 percent are found in both the Atlantic and the Pacific oceans, whereas a lot of species are geographically separated forms which diverged a long time ago. A similar picture is observed here as regards other animals such as mollusca, sea urchins, worms, crustacea, etc.
According to Darwin the area's width may form an immediate contribution to speciation. Geographic races or subspecies may be considered to be potential new species. However, to realize these potentialities and for sub-species to become species will require further divergence, resulting eventually in reproductive isolation and the appearance of various adaptative mechanisms. Geographical races are always found under different conditions, which determines their greater or smaller ecological specialization. For this reason they can be regarded as ecological races at the same time. Ecological specialization inside a species may take place in adjacent areas without a significant geographical separation of the differentiating forms, the possibility of genetic information exchange between specimens of neighbouring populations being preserved. In this respect, the multitude of endemic forms in large isolated water basins is very illustrative. In Lake Baikal, for example, 300 species of crustacea are found, most of which occur nowhere else in the world. In the Njasa Lake 174 endemic species of a fish of the family Cichlidae were found. The origin of these endemic peculiarities is most likely the result of ecological separation and differences in selection effects in locally independent populations.
Of importance is seasonal separation, manifesting itself in different reproduction seasons. Berg (1953) described five forms of trout in the Sevan Lake (Salmo ischchan) having considerable differences in spawning time and a number of other biological characteristics. He also described spring and winter races in the salmon, sturgeon and carp, spawning independently in spring and autumn. The basis for the differences in morpho-physiological characteristics of geographic and ecological races is the divergence in the genetic structure of the component populations. If such a divergence progresses to a considerable degree, it results in the development of reproductive separation mechanisms, capable, up to a point, of restricting the crossing of specimens representing different races.
The basic forms of reproductive separation manifest themselves in:
Change in mating selectivity often precedes the appearance of other forms of reproductive separation in animals. This may be explained by the fact that sexual behaviour of individuals of opposite sexes is a complicated, specialized, highly specific combination of reactions, which are controlled by heredity. Mating behaviour in many species of fish, birds and mammals can serve as an example of these types of selection.
The existence of fertilization selectivity has been ascertained in experiments whereby a comparison is made of the fertility of sperm from males of different lines and breeds by crossing them with females of a different genotype. Of the other two mechanisms of reproductive separation - a decrease in fertility and the hybrids' viability - their origin is the consequence of considerable genetic differences, when the diverged forms can be described as independent species.
Sympatric speciation inevitably involves distant separation of populations. Animals and plants have a number of mechanisms which provide for intraspecies divergence.
The basis for sympatric speciation is inherent in the hereditary phenotypic polymorphism of populations which are caused by different environmental conditions. In plants, essential genetic factors for sympatric speciation may be polyploidy or mutations which determine the incompatibility of genes and cytoplasm. In animals, such factors may be sex heterogametism, sterility genes and cytoplasmic separation.
Whatever the actual relation between sympatric and allopatric speciation, it should be borne in mind that the primary importance in intraspecies divergence and the formation of separating mechanisms is attached to selection and genetic mechanisms, initiating and controlling the processes of microevolution.
Berg, L.S., 1953 Spring and winter races in migratory fishes. “Essays on general problems in ichthyology” (Ocherki po obschem voprosam ikhtyologi)
Darwin, C., 1859 Of the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. J.M. Dent and Sons, Ltd., London
Schmalhausen, I.I., 1946 Faktory evolyutsii (Factors of evolution). Akad.Nauk SSSR, Moscow
Tschetverikoff, S.S., 1926 On certain features of the evolutionary process from the point of view of modern genetics. Zh.exp.Biol., 2:3–54
Wallace, B., 1956 Studies on irradiated populations of Drosophila melanogaster. J.Genet. 54 (2):280–93
Wright, S., 1931 Evolution in Mendelian populations. Genetics, 16:97–159
![]()
![]()
![]()