![]()
![]()
![]()
Les recherches sur les contenus stomacaux ont été traditionnellement un domaine d'activité important dans la biologie halieutique, mais c'est un domaine dans lequel il existe de grandes difficultés pour mettre en corrélation les résultats avec les recherches effectuées dans les autres domaines. Les recherches sur la nourriture des poissons ne peuvent pas être considérées isolément mais doivent être discutées en relation avec l'ensemble du milieu marin dont les poissons constituent de simples éléments. Il faut dono faire une brève revue générale des processus les plus importants de l'écologie aquatique en portant une attention particulière à l'alimentation.
Les organismes vivants réagissent les uns sur les autres et avec leur environnement non-vivant (abiotique) de diverses façons; il n'existe pas d'organisme indépendamment de son environnement. C'est l'étude de ces interrelations qui est appelée écologie. Il est possible d'étudier l'écologie d'une espece en relation à son environnement ou à un groupe complet d'espèces et leurs interactions à la fois entre elles et avec leurs environnements. L'écologie touche done non seulement aux disciplines biologiques, mais aussi aux sciences physiques et chimiques.
Toute aire de la nature dans laquelle des matériaux sont échangés entre des organismes vivants et leur environnement abiotique forme un système écologique ou écosystème. Ce concept est utile car il souligne l'interdépendance des composants mis en jeu. Bien qu'il soit à peine possible de délimiter une aire dans la nature qui ne soit pas influencée par ses voisines, on peut néanmoins considérer un étang, un lac ou même une partie d'une forêt comme un écosystème.
Pour comprendre la dynamique d'un tel écosystème indépendant comme une unité fonctionnelle, ses composants doivent être d'abord considérés en détail.
D'un point de vue fonctionnel on peut reconnaître les étapes suivantes dans le fonctionnement de l'écosystème (Fig. 6.1 et 6.2):
Les producteurs principaux sont les plantes vertes à chlorophylle; les bactéries synthétisantes ne jouent généralement qu'un rôle très mineur.
Les consommateurs comprennent tous les autres composants vivants présents. Ils comprennent les herbivores (végétariens), qui s'alimentement directement aux dépens des producteurs, et les carnivores (prédateurs) qui s'alimentent aux dépens des herbivores ou d'autres carnivores. Ils comprennent aussi les parasites, les nécrophages (mangeurs de charogne) et les saprophytes. Bien qu'il convienne de porter sur la liste les “décomposeurs” comprenant des bactéries et des champignons, comme une entité séparée à cause de leur rôle spécifique et de leur caractère indispensable ils sont naturellement aussi consommateurs.
Les éléments nutritifs inorganiques comprennent un grand nombre d'éléments présents sous forme de sels dissous. Les plus importants sont l'azote et le phosphore suivis du potassium, du calcium, du soufre et du magnésium. Des éléments sont nécessaires en quantités extrêmement petites et sont donc connus comme micro-éléments nutritifs.
La production de substances organiques (nourriture) par photosynthèse est un processus impliquant la transformation de l'énergie lumineuse en énergie chimique potentielle. Le transfert de cette énergie alimentaire des producteurs à une série de consommateurs est appelé chaîne alimentaire, chaque organisme par lequel elle passe étant un maillon de la chaîne.
Pour simplifier on peut reconnaître trois chaînes alimentaires différentes:
En réalité la nourriture peut passer par des parties des trois chaînes avant d'être décomposée finalement en éléments nutritifs inorganiques par les bactéries et champignons trouvés à la fin de chaque chaîne alimentaire.
En d'autres termes, les populations d'espèces dans une communauté ou un écosystème forment plusieurs chaînes alimentaires qui s'interconnectent, s'anastomosent ou se croisent entre elles d'une manière complexe, que l'on cite habituellement sous le nom de réseau alimentaire.
Les organismes qui appartiennent au même maillon de la chaîne alimentaire comptés au niveau des producteurs sont déclarés appartenir au même niveau trophique. Les plants constituent donc le premier niveau trophique, les herbivores le second, et les carnivores se nourrissant des herbivores le troisième niveau trophique. Les carnivores secondaires se nourrissant sur les carnivores du troisième niveau appartiennent au quatrième niveau trophique, et ainsi de suite. Cependant, il existe une limite bien définie au nombre de maillons possibles dans una chaîne alimentaire, et, par conséquent aussi au nombre de niveaux trophiques dans tout écosystème. La raison en est que 10 pour cent seulement de l'énergie disponible est assimilée en passant d'un niveau trophique au suivant. Au bout de la chaîne alimentaire il y a habituellement un ou deux majeurs prédateurs. Le nombre d'espèces dans chaque couche trophique augmente quand on se rapproche de la première couche, laquelle engendre ce qui est appelé une pyramide de nombres. Pour les majeurs prédateurs l'introduction de petites quantités de polluants dans la première couche trophique peut avoir des conséquences fatales parce qu'ils s'y concentrent éventuellement.
Les lois de la thermodynamique établissent que l'énergie ne peut être créé ni détruite, mais aussi qu'elle ne peut être transformée d'un type en un autre sans dissipation partielle en énergie thermique. Cela signifie que la transformation de l'énergie lumineuse en énergie chimique potentielle sous forme de composés organiques dans les plantes ne peut pas être efficace à 100 pour cent. Une très petite partie seulement de l'énergie lumineuse absorbée par les plantes vertes est transformée en énergie alimentaire (production brute) parce que la plus grande partie est dissipée sous forme de chaleur. En outre, une partie de la production brute synthétisée est utilisée par les plantes dans leurs propres processus respiratoires, laissant une quantité encore plus petite d'énergie potentielle (la production nette) disponible pour être transférée au niveau trophique suivant.
La perte d'énergie est généralement appelé la perte respiratoire parce que les organismes utilisent l'énergie des aliments en les oxydant. A cause de la perte respiratoire les chaînes alimentaires ne peuvent pas être très longues et le nombre de niveaux trophiques dans les communautés naturelles dépasse rarement quatre ou cinq et souvent se tient à ces chiffres. Cela signifie aussi que la quantité totale de nourriture disponible diminue quand le niveau trophique s'élève. Pour cette raison, on trouve les plus grands animaux se nourrissant soit de plantes, soit d'autres animaux qui sont dans un niveau trophique inférieur, comme par exemple, les baleines mangent le krill et les éléphants les plantes.
Chez les animaux la production brute correspond à la nourriture assimilée, ce qui signifie la nourriture ingérée et absorbée par l'intestin. La production nette est égale ici à la nourriture assimilée moins la respiration.
Alors que la plus grande perte d'énergie dans un écosystème est due aux processus respiratoires, il existe d'autres pertes qui affectent les organismes individuels. Des aliments potentiels ne sont pas ingérés, mais sont soit décomposés directement, soit emmagasinés ou sortent du système ou de la communauté. Une autre source de perte est que toute la nourriture ingérée n'est pas réellement assimilée, une partie traverse le tractus alimentaire et est perdue sous forme de fèces.
Comme il l'a été dit plus haut, quand les organismes meurent ils sont attaqués par les décomposeurs qui en tirent leur énergie en réduissant leur contenu organique en éléments nutritifs inorganiques. Comme il l'a déjà été indiqué, ces éléments nutritifs peuvent être utilisés de nouveau par les producteurs avec pour résultat que les matériaux en jeu circulent continuellement dans le système. Cependant, le courant d'énergie passe strictement par une “rue à sens unique”. Pour maintenir le fonctionnement d'un écosystème il faut fournir continuellement de l'énergie lumineuse.
Comme nous l'avons vu. la vrais production de matière organique prend place seulement chez les plantes à chlorophylle et certaines bactéries synthétisantes, et cela a été appelé production primaire. Cependant, les copépodes et les euphausidés, par exemple, sont souvent cités comme producteurs de chair ou animaux de “l'industrie-clé” parce qu'ils convertissent le matériel végétal en protéine qui peut être assimilée par les animaux qui les mangent mais qui eux-mêmes ne peuvent pas subsister sur le matériel végétal. Naturellement, en réalité, ils assimilent et emmagasinent seulement l'energie dérivée des producteurs primaires. Pour éviter la confusion il serait mieux de les appeler producteurs secondaires, un terme qui, naturellement s'applique aussi bien à des animaux à des niveaux trophiques plus élevés parce qu'eux aussi - bien qu'indirectement - utilisent la production primaire des plantes.
D'un point de vue pratique il est souvent désirable de rechercher quelle est la grandeur de la production secondaire de certains animaux dans une aire donnée, soit un banc de pêche, ou même plus important, si une production connue peut être augmentée. Les estimations de production doivent être basées sur des facteurs tels que la “standing crop” (biomasse), le taux d'enlèvement de matériel et le taux de croissance, comprenant la croissance des jeunes nés ou éclos pendant la période du recensement. Le taux de renouvellement présente aussi de l'intérêt quand des espèces à vie courte sont impliquées comme c'est pratiquement toujours le cas dans un écosystème dans la mer. La production biologique doit être exprimée par unité de temps. Une grande biomasse n'est en aucune façon synonyme d'un taux de production élevé. Pour prendre un exemple imagé, un pâturage où paissent des vaches peut avoir une très petite biomasse d'herbe parce que la production est mangée aussitôt produite, mais il peut néanmoins avoir un taux de production plus élevé qu'un pâturage voisin non pâturé avec une très grande biomasse.
Les relations quantitatives entre les divers niveaux trophiques peuvent être calculées pourvu que l'on connaisse les taux de production pour chaque niveau concerné. Les relations de cette nature dans les niveaux trophiques présentent aussi un intérêt considérable. Les résultats de oes calcule exprimés en pourcentage sont souvent cités comme rendements écologiques parce qu'ils sont en rapport avec le rendement du transfert d'énergie à differents points le long de la chaîne alimentaire. Ils sont donc importants pour notre compréhension de la dynamique dans les écosystèmes. En outre, la plupart des taux de rendement sont significatifs en ce qui concerne les populations d'espèces seules, aussi bien que tous les niveaux trophiques.
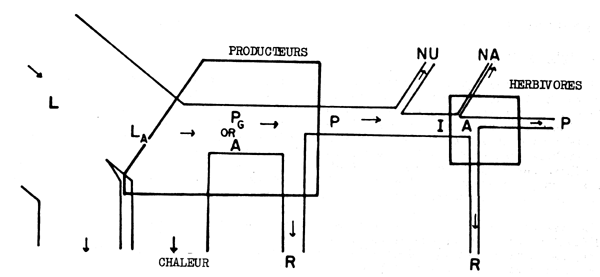
Malheureusement il existe une grande confusion dans les termes utilisés par divers auteurs, et il n'est pas toujours clair de savoir à quel taux de rendement un auteur a réellement désiré se référer. Odum (1959) a fait une tentative pour définir les divers taux, en se basant sur le diagramme de l'écoulement de l'énergie et c'est certainement une bonne méthode pour illustrer les complexités míses en jeu (Fig. 6.3).
En biologie halieutique la facon la plus courante pour décrire le rendement est l'emploi du facteur de conversion, c'est-à-dire, le rapport du poids de nourriture consommée par le poisson à la croissance en poids. Certains auteurs expriment ce facteur de conversion comme coefficient de nutrition. D'après la Fig. 6.3 on voit que le facteur de conversion est la valeur réciproque de ce qu'Odum appelle rendement de la croissance écologique. La valeur du facteur de conversion est traditionnellement définie comme étant 10, mais en fait il peut être beaucoup plus élevé ou beaucoup plus faible, p. ex., dans les élevages de truite, du nord de l'Europe, le facteur de conversion est de 5 environ.
Cependant, il est important de souligner qu'aucun des rendement écologiques n'est constant pour la population de n'importe quelle espèce ou pour un niveau trophique entier. Ils dépendent d'un certain nombre de facteurs abiotiques tels que la température et la salinité, de même que facteurs biotiques comme le type, l'abondance et la distribution de la nourriture disponible, et l'âge des consommateurs. Par exemple, les larves et les jeunes de toutes les espèces étudiées ont des facteurs de conversion beaucoup plus faibles que les animaux plus âgés.
Presque tout le travail qui a été fait sur la nourriture ingérée par les poissons a été qualitatif plutôt que quantitatif. C'est-à-dire que les chercheurs ont décrit la présence de nourriture trouvée dans les tractus digestif, généralement dans l'estomac seulement. Cela indique ce que le poisson a mangé et approximativement en quelles proportions mais ne décrit pas combien de chaque espèce de nourriture est mangé. La raison du manque de travaux quantitatifs est qu'ils nécessitent beaucoup de temps.
Le plus tôt possible après la capture du poisson tout le tractus digestif est disséqué, et si les recherches ne peuvent pas être terminées immédiatement, possiblement à cause du manque d'installations de laboratoire, le matériel doit être conservé dans du formol à 4 pour cent tamponné.
Plus tard, au laboratoire on ouvre les différentes parties du tractus digestif et les groupes individuels d'organismes son triés pour identification. Ceci se fait très facilement avec un microscope binoculaire à dissection. Pour illustrer les méthodes. employées et pour décrire leurs avantages et inconvénients une série de résultats seront étudiés, en se basant sur l'examen du contenu de l'estomac de deux Lates niloticus (Tableau 6.1).
La présence de chaque aliment est relevée et exprimée en pourcentage de la présence de tous les organismes de la nourriture. La méthode est très rapide et facile mais elle sousestime l'importance des grandes espèces. Dans l'exemple 40 pour cent de toutes les présences de nourriture étaient des nymphes d'Odonata. Le contenu de l'estomac de ces deux L. niloticus aurait pu consister presque entièrement de nymphes, chacun avec un très petit Tilapia galilaea mais les résultats ne donnent pas d'indications si ce peut être une interprétation vraie des données.
Les nombres de chaque pièce d'aliment sont relvés et les résultats exprimés en pourcentage du nombre total de pièces d'aliments présentes. Cette méthode surestime l'importance des petits organismes de la nourriture, dans ce cas les nymphes d'Odonata. Elle n'est pas satisfaisante avec la nourriture végétale, sauf avec l'examen microscopique des algues. Dans l'exemple donné le matériel végétal pouvait être un petit mourceau d'herbe ou un grand morceau.
La méthode est similaire à celle des fréquences de présence sauf que les résultats sont exprimés en termes du nombre de poissons dans lesquels se trouvait l'aliment exprimé en pourcentage du nombre total de poissons. La méthode donne une indication de ce que sont les principales espèces alimentaires mais n'indique pas leur importance relative comme sources d'énergie.
On mesure le volume total de l'estomac. Le contenu est ensuite trié en différents types de nourriture et on détermine le volume de chaque type. Les résultats sont en général rapportés en pourcentage du volume total.
Cette méthode demande plus de temps que n'importe laquelle des méthodes précédentes mais elle décrit avec précision l'importance relative de chaque aliment et le volume total (∑) indique la quantité de chaque aliment mangé.
Cette méthode est identique à la précédente sauf que le volume de chaque aliment est d'abord exprimé en pourcentage de volume de tout le contenu de l'estomac dont il provient et ensuite en moyenne relevée pour tous les poissons échantillonnés. L'avantage de cette méthode sur la précédente est que les résultats finaux seront plus représentatifs des habitudes alimentaires d'un groupe de poissons si un ou deux individus de ce groupe ont beauooup consommé un aliment particulier. L'inconvénient est que l'information sur le volume réel du contenu de l'estomac est perdue.
On détermine les poids totaux de tous les aliments. Cette méthode est très similaire à la méthode volumétrique, sur laquelle elle a un léger avantage à moins que l'échantillon soit sec, auquel oas on détermine seulement le poids sec. Les résultats sont habituellement présentés en pourcentage du total. Si on détermine les poids humides les aliments doivent être placés sur un buvard pendant un moment peur enlever l'exoès d'humidite. Ce problème d'enlèvement de l'excès d'humidité rend la méthode quelque peu subjective et par conséquent le poids sec est meilleur, bien que demandant beaucoup plus de temps.
Chaque aliment recoit des points. Le nombre de points dépend à la fois de deux choses: si l'organisme est très commun dans le contenu de l'estomac (nombre le plus élevé de points) ou rare (le plus faibles nombres) et de sa taille (plus de points pour les grandes que pour les petites tailles). La méthode peut aussi être modifiée pour tenir compte de la plénitude de l'estomac. Elle est rapide, facile et ne nécessite aucun appareil spécial; avec de l'expérience, la méthode peut être très précise. Dans l'exemple donné (Tableau 6.1) les points alloués donnent un résultat similaire à ceux des méthodes volumétrique et gravimétrique.
Rounsefell et Everhart (1953) donnent d'autres détails sur ces méthodes.
Toutes ces méthodes décrivent, avec un degré plus ou moins grand de précision, la quantité de nourriture que les deux L. nilotious, qui ont été pris comme exemples, avaient dans leur estomac, les meilleures étant les méthodes volumétrique et gravimétrique. La somme des volumes réels et des poids indique aussi le degré de plénitude de l'estomac, qui permet de décrire les cycles de prise de nourriture. Le système des points est bon aussi s'il est utilisé objectivement et a les avantages supplémentaires de la facilité et de la rapidité. Il peut aussi être adapté pour tenir compte la plénitude de l'estomac.
Cependant, aucune des méthodes ne décrit la quantité ingérée de chaque aliment. Dans l'exemple utilisé la digestion de T. galilaea peut être bngue et ce poisson peut être identifiable longtemps après que les nynphes d'Odonata aient cessé de l'être. Pendant le temps nécessaire pour digérer un T. galilaea 20 à 30 nymphes d'Odonata peuvent avoir été ingérées et digérées mais à n'importe quel moment 4 à 6 seulement peuvent être reconnaissables dans l'estomac. D'autre part, des aliments peuvent avoir été régurgités à la capture. Alors que les méthodes qualitatives permettent la construction de chaînes alimentaires, elles ne décrivent pas le courant d'énergie à travers ces chaînes alimentaires ni ne permettent pas de déterminer l'importance de, par exemple, la prédation d'une espèce commerciale de valeur sur une autre espèce également de valeur. Si on connaissait les réponses à ces problèmes on pourrait dans quelques cas trouver recommandable de réduire l'abondance d'une espèce prédatrice pour permettre une augmentation de l'abondance de sa proie, spécialement si la proie était à un niveau trophique inférieur(meilleur rendement). On ne peut répondre à ces questions que par des études qualitatives.
Des études quantitatives d'alimentation prennent beaucoup de temps et sont aussi le travail des spécialistes. Il ne sera donné qu'un bref aper¸u des méthodes utilisées. Premièrement, on choisit l'unité dans laquelle le courant d'énergie doit être mesuré, les calories et l'azote étant les deux plus communes. En second lieu, les expériences d'alimentation sont effectuées dans des aquariums avec les différents aliments que l'espèce étudiée mange afin de déterminer quel est son rendement pour convertir l'aliment en chair de poisson. Pour les calories, cela signifie déterminer la valeur calorifique de l'aliment choisi, en nourrissant le poisson avec un poids connu de cet aliment, en recueillant ses fèces et en déterminant la valeur calorifique. Afin de traduire les observations sur les poids et les valeurs calorifiques de la nourriture trouvée dans l'estomac les taux de digestion doivent être établis en nourrissant des poissons et ensuite en les tuant à intervalles connus. Ceci peut devoir être fait pour plus d'une température. D'après ces informations on construit une image de l'ingestion quotidienne de nourriture de poissons de la même espèce de différentes tailles, et combien de l'énergie provenant de cet aliment ils utilisent pour l'entretien et la croissance. Une telle étude de Daan (sous presse) montre que les grandes morues du sud de la mer du Nord ont probablement une grande influence sur la taille des classes d'âge en mangeant les petites morues du groupe O, et qu'elles mangent aussi de grandes quantités de harengs et de soles. Boddeke (1971) a aussi montré que de récentes grandes classes d'âge de morues ont réduit l'abondance de la crevette grise (Crangon crangon) au large de la côte hollandaise. Daan a suggéré que la production totale d'espèces commerciales du sud de la mer du Nord puisse être augmentée en réduisant l'abondance de la morue en pêchant intensément sur le stock. La solution de telles relations prédateur-proie peut altérer la manière selon laquelle des estimations de poissons ont été faites.
Allee, W.C. et al., 1949 Principles of animal ecology. Philadelphia, W.B. Saunders
Boddeke, R., 1971 The influence of the strong 1969 and 1970 year classes of cod on the stock of brown shrimp along the Netherlands coast in 1970 and 1971. ICES C.M. Shellfish and Benthos Committee. Paper K32:3 p (mimeo)
Clarke, G.L., 1954 Elements of ecology. New York, Wiley and Sons, 534 p.
Daan, N., A quantitative analysis of the food intake of North Sea cod. Gadus morhua, L. Neth. J. Sea Res., (in press)
Hardy, A.C., 1959 The open sea; its natural history. Pt. 2. Fish and fisheries with chapters on whales, turtles and animals of the sea floor. London, Collins, 322 p.
Laevastu, T., 1965 (comp.), Manual of methods in fisheries biology. FAO Man. Fish. Sci., (1): pag. var.
Marshall, N.B., 1966 The life of fishes. Cleveland, Ohio, World Publishing Co., 402 p.
Nikolskii, G.V., 1963 The ecology of fishes. London, Academic Press, 352 p.
Odum, E.P., 1959 Fundamentals of ecology. Philadelphia, W.B. Saunders, 546 p.
Rounsefell, G.A. and W.H. Everhart, 1953 Fisheries science: its methods and applications. New York, Wiley and Sons, 444 p.
Steele, J.H. (ed.), 1970 Marine food chains. Edinburgh, Oliver and Boyd, 552 p.
Tableau 6.1 Exemple de résultats obtenus en utilisant différentes méthodes d'estimation de contenus stomacaux de deux Lates niloticus
Contenus stomacaux
| L. niloticus 1. | 1 | Tilapia galilaea 20 cm long, poids 80 g, volume 100 ml. |
| 6 | Nymphes d'Odonata (libellule) chacune de 1 cm de long, 0.5 g, 1 ml Matériel végétal, poids 5 g, volume 7 ml | |
| L. niloticus 2. | 1 | T. galilaea 10 cm, 10 g, 15 ml |
| 4 | Nymphes d'Odonata chacune de 1 cm, 0,5 g, 1 ml |
Résultats
| Aliment | Méthode | Poisson | Ln 1 | Ln 2 | ∑ | % | Total dont % est exprimé |
|---|---|---|---|---|---|---|---|
| Présence | |||||||
| T. galilaea | 1 | 1 | 2 | 40 | présence de tous les aliments | ||
| Odonata | 1 | 1 | 2 | 40 | |||
| Végétaux | 1 | 0 | 1 | 20 | |||
| Numérique | |||||||
| T. galilaea | 1 | 1 | 2 | 14,4 | tous les organismes alimentaires | ||
| Odonata | 6 | 4 | 10 | 76,9 | |||
| Végetaux | 1 | 0 | 1 | 7,7 | |||
| Dominance | |||||||
| T. galilaea | 1 | 1 | 2 | 100 | tous les poissons | ||
| Odonata | 1 | 1 | 2 | 100 | |||
| Végétaux | 1 | 0 | 1 | 50 | |||
| Volume total | |||||||
| T. galilaea | 100 | 15 | 115 | 87,1 | volume total des aliments | ||
| Odonata | 6 | 4 | 10 | 7,6 | |||
| Végétaux | 7 | 0 | 7 | 5,3 | |||
| % volume | |||||||
| T. galilaea | 88,5 | 79 | 167,5 | 83,8 | volume des aliments | ||
| Odonata | 5,3 | 21 | 26,3 | 13,1 | |||
| Végétaux | 6,2 | 0 | 6,2 | 3,1 | |||
| Gravimétrique | |||||||
| T. galilaea | 80 | 10 | 90 | 90 | poids total de nourriture | ||
| Odonata | 3 | 2 | 5 | 5 | |||
| Végétaux | 5 | 0 | 5 | 5 | |||
| Points | |||||||
| T. galilaea | 50 | 10 | 60 | 85,7 | aliments | ||
| Odonata | 5 | 4 | 9 | 12,9 | |||
| Végétaux | 1 | 0 | 1 | 1,4 |
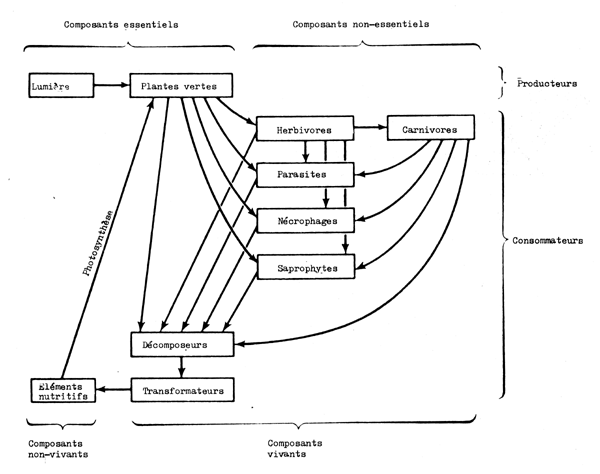
Fig. 6.1 Principales étapes et composants d'un écosystéme autonome (d'après Clarke, 1954)
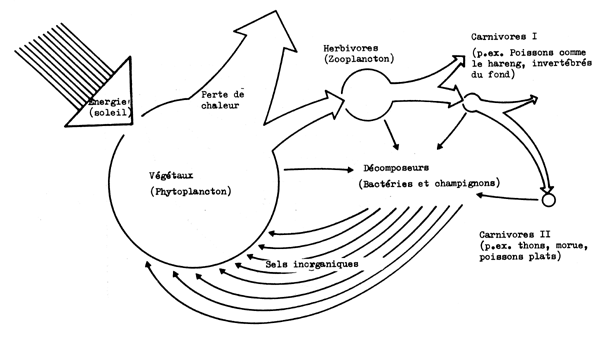
Fig. 6.2 Exemple simplifié d'un écosystème marin. L'écoulement de l'énergic est représenté par des flèches ouvertes
| Relaciones | Designación y explicación | |
|---|---|---|
| A. Rapports entre les niveaux trophiques | ||
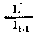 | Niveau trophique d'absorption d'énergie ou rendement (de Lindeman). Pour le niveau primaire c'est | |
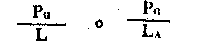 | ||
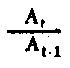 | Niveau trophique du rendement de l'assimilation | Pour le niveau primaire P et A peut être en terms de L ou La comme ci-dessus; At/At-1 = ItIt-1pour le niveau primaire, mais pas pour les niveaux secondaires. |
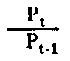 | Niveau trophique du rendement de la production | |
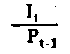 | 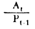 Rendements d'utilisation Rendements d'utilisation | |
| B. Rapports à l'inérieur des niveaux trophiques | ||
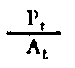 | Rendement de la croissance des tissus | |
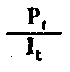 | Rendement de la croissance écologique | |
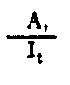 | Rendement de l'assimilation | |
Fig. 6.3 Divers types de rendements écologiques. Les symboles employés sont (voir diagramme): L = lumiére (totale), La = lumiére absorbé e, PG = photosynthése totale (production brute), P = production de biomasse, I = énergie absorbée, R = respiration, A = assimilation, NA = absorbée mais pas assimilée, NU = inutilisée par le niveau trophique montré, t = niveau trophique, t-1 = niveau trophique précédent (d'après odum, 1959)
![]()
![]()
![]()