![]()
![]()
Le praticien de la biologie halieutique recueille des données pour répondre aux deux questions principales: “combien de poissons se trouvent dans la zone où l'on a l'intention de pêcher” et “quelle est la quantité maximum qui puisse être pêchée annuellement sans affecter la capacité du stock de produire ce rendement”. Incidemment il peut satisfaire sa curiosité scientifique touchant la biologie des poissons qu'il étudie ou le milieu dans lequel ils vivent, mais il ne doit jamais perdre de vue son principal objectif: répondre aux deux questions.
Jusqu'à présent, les sections de ce manuel ont été consacrées aux données à recueillir et aux meilleurs moyens de les recueillir. Cette section décrira les utilisations auxquelles s'appliquent les données.
Il y a deux façons d'aborder ce problème. L'une est de déterminer la quantité totale de poissons présents en termes absolus, c'est-à-dire qu'il est répondu à la question en termes soit de poids total (3 millions de tonnes) soit de nombre total (60 millions de poissons). L'autre est d'y répondre en termes relatifs, c'est-à-dire qu'il est répondu à la question en termes de poissons capturés en une heure de pêche. Des revues des méthodes d'évaluation des ressources sont données par Alverson (1971) et Mackett (1973).
Cette méthode est basée sur le fait que si T individus sont marqués dans une population totale de N individus, la probabilité qu'un individu soit marqué dans un échantillon pris au hasard dans la population est T/N. Si on capture n spécimens et si m de ceux-ci se trouvent être marqués alors le nombre de poissons dans la population est N = nT/m. Pour fixer les idées, supposons que 3 000 poissons ont été marqués pendant une expérience (T = 3 000) et que dans les 12 mois suivant leur mise en liberté 3 millions de poissons de cette espèce ont été mis à terre provenant du stock dans lequel on a effectué le marquage (n = 3 × 106). Ce nombre serait calculé d'après le programme d'échantillonnage du marché (Sections 2 et 3); de ces quantités débarquées les pêcheurs ont renvoyé 750 poissons marqués (= m). Alors une estimation du nombre de poissons de taille marchande dans le stock (N) serait nT/m = 3 000 000 × 3 000/750 = 12 millions de poissons.
Dans ce calcul on fait plusieurs suppositions, à savoir:
(a) les poissons marqués et ceux non-marqués sont mélangés également (ce qui serait improbable après une expérience normale de marquage);
(b) les taux de mortalité ou les taux d'émigration ou tous deux des poissons marqués ou non sont les mêmes;
(c) les poissons marqués et les poissons non-marqués sont également susceptibles d'être capturés par l'engin;
(d) qu'il n'y a pas de mortalité causée par le marquage, pas de marques perdues, et que tous les poissons marqués sont retournés.
Même si toutes les conditions sont remplies l'estimation de N obtenue comporte une erreur systématique. Une meilleure méthode implique les captures et recaptures multiples, donnant la formule suivante (d'après Cormack, 1968):
 i = Zi = (Si/ri) + Mi
i = Zi = (Si/ri) + Mi
dans laquelle:
i = nombre estimé des individus marqués au temps ti
Zi = nombre de poissons marqués avant ti, non capturés à ti mais recapturés ultérieurement
Si = nombre de poissons marqués au temps ti
ri = nombre de poissons Si, recapturés ultérieurement
Mi = nombre de poissons marqués au début du temps ti
ensuite 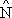 = (ni/mi). i
= (ni/mi). i
dans laquelle i = nombre de poissons dans la population au temps ti
ni = nombre de poissons capturés au temps ti.
La méthode dépend encore de la validité des assomptions a-b. Les méthodes de marquage et de recapture ont leur application potentiellement la plus utile dans les petites masses d'eau fermées comme les étangs et les petits lacs, mais même dans de petites mares il peut être très difficile d'obtenir des estimations sûres de la dimension de la population (voir Holden, 1963).
Des engins comme les chaluts et les sennes balayent un certain volume d'eau. Ce volume peut être calculé approximativement d'après les dimensions de l'engin. Si on suppose que l'engin capture tous les poissons sur sa route on peut déterminer une estimation minimum du nombre de poissons si le volume de la mer habité par le stock est connu. Cette méthode est une estimation minimum parce qu'elle suppose que tous les poissons se trouvant sur la route de l'engin sont capturés, ce qui est invraisemblable. Pour les chaluts pélagiques l'action d'éviter l'engin peut être si grande ou éliminée si efficacement par la pêche dirigée que la méthode trouve sa principale application aux engins trainés le long du fond de la mer. Les calculs sont faits en termes de surface. Dison que 1 000 traits de chalut ont été effectués au hasard sur 100 000 milles carrés (258 999 km²) de fond de la mer habité par un stock. Supposons que chaque trait de chalut balaye 0,5 mille carré (1,194 km²) du fond de la mer et que la pêche moyanne soit de 60 poissons. En balayant 500 milles carrés (129 499 km²) (1 000 traits × 0,5 mille carré) 60 000 poissons ont été capturés (60 poissons × 1 000 traits). Ainsi dans 100 000 milles carrés le nombre estimé de poissons est 60 000 × 100 000/500 = 12 × 106 poissons. Cette méthode peut être très utile pour donner une première estimation de la quantité de poissons présents.
L'emploi des études des oeufs et larves pour estimer les nombres du stock ont déjà été décrits dans la section 7.3.3. La méthode peut être très utile pour les poissons dont les oeufs sont pélagiques, si ces oeufs peuvent être identifiés rapidement comme appartenant à des espèces ou des groupes d'espèces.
De simples sondeurs à écho donneront quelque idée de l'abondance de poissons, mais les sondeurs à écho employés avec des intégrateurs donneront de très bonnes estimations de l'abondance de poissons, en particulier des espèces pélagiques. Les méthodes sont décrites en détail par Forbes et Nakken (1972).
Les sondeurs à écho fonctionnent simplement en envoyant une onde sonore dans l'eau. Si cette onde sonore frappe un objet qui va la réfléchir, une partie de cette onde sonore est réfléchie, la grandeur de cette partie dépendant du type de l'objet et de sa profondeur. Quand elle est reçue au retour, par le récepteur du sondeur à écho elle est transformée en un courant électrique qui est alors utilisé pour actionner un écran. Les intégrateurs fonctionnent en additionnant les voltages des courants électriques de retour et en représentant les réponses sur un tableau étalonné. Ils tiennent compte aussi de la profondeur de l'objet, ainsi une onde sonore réfléchie par un poisson à 50 m donnera la même valeur d'enregistrement qu'un poisson à 10 m. Les poissons d'espèces et de tailles différentes donnent des réponses différentes mais une fois que l'équipement a été étalonné pour l'espèce étudiée la quantité absolue de poissons peut être déterminée d'après l'enregistrement intégré. La pêche courante doit être pratiquée pendant l'étude pour déterminer à quelles espèces les enregistrements sont dûs, à moins que l'on sache qu'une seule espèce habite l'aire ou à moins que l'estimation de plusieurs espèces soit acceptable.
C'est une technique qui peut être utilisée si la majeure partie du stock est capturée et que la structure des âges de la capture soit connue avec une bonne précision. Dans cette méthode on utilise les résultats obtenus de l'échantillonnage du marché et des programmes de détermination des âges. En faisant deux suppositions, l'une sur le taux de mortalité naturelle et l'autre sur le taux de mortalité due à la pêche sur le groupe d'âge le plus ancien dans chaque classe d'âge il est possible de calculer par un processus réitératif les nombres de poissons dans chaque groupe d'âge dans le stock. La méthode fonctionne en considérant chaque classe d'âge séparément et en calculant la dimension de cette classe d'âge au début de chaque année et la mortalité due à la pêche agissant sur elle pendant toute cette année. Les estimations de ces deux paramètres deviennent de plus en plus précises quand les calculs procèdent des plus vieux aux plus jeunes poissons dans une classe d'âge. La précision des deux ou trois premières estimations dépend de l'exactitude de la valeur initiale du taux de la mortalité due à la pêche. Des détails complets de cette méthode sont données par Fry (1957), Gulland (1965) et Pope (1972).
Les méthodes relatives sont basées sur quelque forme de capture par indice d'unité d'effort, calculé d'après les informations fournies par un navire industriel ou de recherche. Elles sont relatives parce qu'elles indiquent seulement ce qu'il advient à l'abondance du stock et non sa dimension totale réelle; l'abondance est habituellement mesurée en capture par unité d'effort.
Les calculs des indices commerciaux de capture par unité d'effort ont été donnés dans la section 2, dans laquelle les méthodes de rassemblement des données sur les captures et unités d'effort ont été décrites et on a souligné l'importance de relever des données d'effort qui fournissent un indice significatif de l'abondance de poissons. Le dernier point est plus important, particulièrement quand le rendement de navires change ou que l'on introduit des types de pêche absolument nouveaux.
Les données sur la capture par unité d'effort sont utilisées de plusieurs façons différentes. Premièrement, elles peuvent être utilisées pour décrire les changements d'abondance du stock d'une ennée à l'autre. Utilisées avec les données de capture totale elles peuvent être utilisées pour décrire un modèle simple à partir duquel on peut calculer le rendement soutenu à long terme (Gulland, 1969). Elles sont aussi utilisées pour calculer les taux de mortalité totale d'après les nombres dans les groupes d'âge successifs d'une classe d'âge mis à terre pendant deux années consécutives. Par exemple, 1 000 poissons âgés de 5 ans peuvent être mis à terre une année et 500 poissons âgés de 6 ans l'année suivante. Comme ce sont des chiffres de captures totales ils ne donnent aucune indication de l'abondance de cette classe d'âge pendant les deux ans. Cependant, si l'effort de pêche pendant la seconde année était le double de celui de la première année, l'abondance relative était 1 000 : 250 (1 000/1 : 500/2). La survivance d'une année à la suivante de cette classe d'âge particulière était 0,25.
Une des difficultés des indices d'abondance industriels a déjà été mentionnée, c'est que le rendement des navires change d'une année à l'autre. Une autre est qu'il n'existe aucun contrôle sur les lieux ou la flotte pêche. Dans une pêcherie pour deux ou plusieurs espèces, la flotte va se concentrer sur ces espèces qui lui assurent les meilleurs profits financiers et il est probable que cela change avec la saison et annuellement. Le problème peut être surmonté jusqu'à un certain point en utilisant des indices calculés pour les périodes pendant lesquelles à la fois le stock et la pêcherie sont concentrés; par exemple, pendant la saison de reproduction.
Les expéditions de navires de recherche surmontent ces problèmes en grande partie et peuvent aussi être planifiées de façon que l'écart-type des réponses puisse être calculée. Un indice industriel peut montrer que la capture par unité d'effort est tombée de 5 à 4 tonnes par heure de pêche mais il n'est pas possible de dire d'après cela si l'abondance a baissé de 20 %. Cependant, la prospection par un navire de recherche peut montrer qu'elle a baissé de 5 ± 0,25 tonnes à 4 ± 0,25 tonnes par heure de pêche. Les limites de confiance de 95 % sont approximativement le double de l'écart-type (Section 2.3.3), il est donc certain à 95 % que la capture par unité d'effort a baissé d'entre 5,4 et 4,6 tonnes à un chiffre situé entre 4,5 tonnes et 3,5 tonnes par heure de pêche. Il est certain à 95 % qu'il y a eu une chute réelle de l'abondance.
Pour obtenir des résultats avec un écart-type connu il est nécessaire de baser les études sur l'emploi des techniques statistiques. Jones et Pope (1973) décrivent une étude du bano des Féroé dans laquelle ils avaient divisé la zone comme le montre la Fig. 9.1. C'est un exemple d'échantillonnage stratifié (Section 2.4). Dans chaque strate ils établirent une grille de stations potentielles et ensuite en choisirent deux au hasard (voir Section 2 … pour l'emploi de nombres pris au hasard). Ils ont aussi stratifié la pêche par intervalles de temps afin de pouvoir considérer les effets des variations diurnes de l'abondance.
Il est peu probable que les premiers poissons capturés soient un échantillon réel de la population et les mises à terre d'un navire de pêche industriel ont encore moins de chances de l'être parce qu'en mer on aura effectué quelque triage avec rejet à la mer du poisson écarté. Cela peut être vérifié en questionnant pour obtenir des informations sur les captures, l'effort de pêche et les lieux de pêche (Section 2.10). La capture dépendra d'une manière plus importante de la sélectivité de l'engin et de son grément.
La plupart des engins ne capturent pas un échantillon au hasard de tous les poissons présents; ils sélectionnent soit les gros poissons, soit les petits.
Un des cas le mieux étudié de sélectivité est celui des culs-de-chalut. Pour une dimension de maille donnée tous les petits poissons passeront à travers et tous les grands poissons seront retenus, mais quelque part au milieu de cette gamme il y aura un groupe de longueurs dans lequel des poissons d'une certaine taille sont capturés parce qu'ils sont trop gros pour passer à travers les mailles mais d'autres de la même taille passent à travers parce qu'ils sont suffisamment minces. Si on porte sur un graphique la proportion retenue à chaque longueur et la longueur on obtient une courbe sigmoïde (Fig. 9.2a). La caractéristique importante de cette courbe est le point des longueurs où la moitié des poissons sont retenus et la moitié s'échappent, c'est la longueur de sélection à 50 %. Divisée par la dimension de maille moyenne du cul-de-chalut elle donne le facteur de sélection. (La dimension de maille est mesurée avec una jauge spéciale à ressort tendu). D'autres détails de la sélectivité du chalut et des méthodes de son étude sont donnés par Pope (1966). Holden (1973) donne une bibliographie complète des références.
Une fois la courbe de sélectivité connue on peut effectuer la correction pour les poissons dans les longueurs retenues par le cul-de-chalut; c'est-à-dire, si 50 poissons ont été retenus dans la gamme de longueurs pour lesquelles la sélectivité était 0,25, 200 poissons ont pénétré réellement dans le cul-de-chalut (50/0,25 = 200).
Pour les filets maillants la sélectivité augmente jusqu'à un maximum, comme pour les culs-de-chalut, mais ensuite décline jusqu'à zéro quand les poissons deviennent trop gros pour être retenus par l'engin (Fig. 9.2b). Les méthodes pour étudier la sélectivité des filets maillants et pour corriger les données de capture obtenues avec des filets maillants sont données par Holt (1963).
Il y a aussi une sélectivité des hameçons; en général les hameçons prennent proportionnellement plus de grands poissons que de petits. Cela n'a pas été étudié en détail, jusqu'à présent il n'y a pas de méthode standard de correction des données de capture par l'hameçon.
Autre la sélection par les mailles, la sélection par les engins à la fois pour l'espèce et la taille est rencontrée fréquemment. On peut s'attendre à ce que différents engins ne soient pas également efficaces pour capturer différentes espèces. Quand les chalutiers ont pêché sur les mêmes fonds que les senneurs danois, la composition des espèces des captures a été très différente. D'après des résultats expérimentaux Margetts (1949) a estimé que pendant une période standard de 24 heures un senneur danois capturait 2,8 fois plus de plies qu'un chalutier. Les chalutiers capturaient relativement plus de soles, de baudroies, de turbots et de barbues que le senneur. On a aussi estimé que la surface du fond de la mer pêchée par le senneur, comprise comme la surface balayée par les funes et le filet, était environ 3 fois celle couverte par le chalut. Les rendements des deux engins, jugés d'après leurs capacités à capturer tant de poissons par unité de fond, étaient donc à peu près les mêmes pour la plie mais le chalut était plus efficace pour capturer des espèces telles que la sole.
Des différences mineures entre les engins peuvent aussi causer des différences d'efficacité. La majorité des expériences de sélection des mailles de cul-de-chalut ont montré que les grandes mailles capturent plus de poissons dont la longueur dépasse le point de rétention de 100 %. Le tableau 9.1 donne le taux des captures totales de plies obtenues pour des durées égales de pêche par deux chaluts dont les culs-de-chalut ont des mailles de taille différente. En théorie les taux ne doivent pas dépasser 1,0. Le désaccord peut provenir de différents écoulements de l'eau à travers les culs-de-chalut.
Les données de capture obtenues lors d'une comparaison entre les deux navires de recherche britanniques SIR LANCELOT et EXPLORER ont montré que le SIR LANCELOT avait capturé moins d'églefins, mais davantage de grands églefins que l'EXPLORER (l'EXPLORER avait capturé 1 262 églefins de 31–44 cm de long, 61 de 45–56 cm comparés aux 406 et 88 églefins des mêmes groupes dans les captures du SIR LANCELOT). Les chaluts étaient munis de culs-de-chalut ayant à peu près la même dimension de maille mais ils avaient des nombres différents de flotteurs et des bras différents. Plus tard, en utilisant la photographie sousmarine des chaluts on a déterminé leur forme en pêche. Le filet de l'EXPLORER était largement étalé mais avait una faible ouverture verticale, et le SIR LANCELOT avait une ouverture horizontale relativement étroite mais une beaucoup plus grande ouverture verticale. Cela suggère que le chalut du SIR LANCELOT capturait plus de gros églefins paroe que ceuxci nageaient plutôt plus au-dessus du fond que les plus petits.
| Longueur cm | Taux de capture en nombre par heure |
|---|---|
| 19,5 | 0,01 |
| 20,5 | 0,03 |
| 21,5 | 0,03 |
| 22,5 | 0,04 |
| 23,5 | 0,06 |
| 24,5 | 0,05 |
| 25,5 | 0,11 |
| 26,5 | 0,12 |
| 27,5 | 0,15 |
| 28,5 | 0,32 |
| 29,5 | 0,42 |
| 30,5 | 0,96 |
| 31,5 | 0,78 |
| 32,5 | 0,86 |
| 33,5 | 0,82 |
| 34,5 | 1,13 |
| 35,5 | 1,09 |
| 36,5 | 1,13 |
| 37,5 | 1,20 |
| 38,5 | 0,85 |
| 39,5 | 1,17 |
| 40,5 | 2,00 |
| 41,5 | 0,50 |
| 42,5 | 2,33 |
| 43,5 | 1,13 |
| 44,5 | 0,50 |
| 45,5 | 0,80 |
| 46,5 | 4,00 |
| 47,5 | 1,00 |
| 48,5 | 1,50 |
| (d'après Beverton et Holt, 1957) | |
Toutes les parties d'un filet maillant ne sout pas également efficaces pour capturer les poissons, comme le montrent les captures de saumon en relation avec leur distance de la ligne de plombs du filet (Tableau 9.2). Ce point était aussi mentionné dans l'Appendice 2, Echantillonnage d'une pêcherie de lac africain.
| Distance cm | >Nombre de saumons |
|---|---|
| 5–10 | 158 |
| 15–20 | 136 |
| 30 | 68 |
| 40 | 50 |
| (d'après Christensen 1970) | |
La possibilité pour une espèce d'être capturée peut changer quand le comportement s'altère avec l'âge. Les sprats (Sprattus sprattus (L.)) dans le Wash, sud de la mer du Nord, forment deux couches distinctes de distribution verticale, une couche supérieure de poissons de deux ans et une couche inférieure de poissons de trois ans. Industriellement, les sprats sont pêchés par des chaluts pélagiques mais comme les pêcheurs n'aiment pas les remorquer trop profondément dans cette région de bancs de sable, leurs captures se composent principalement de poissons de deux ans. Dans ce cas la capture par heure de chalutage est une mesure sujette à caution de l'abondance des poissons de trois ans (Cushing, 1968).
La possibilité d'être capturé peut aussi changer d'une saison à l'autre. Les Sardinella aurita adultes sont capturées pélagiquement dans les eaux côtières uniquement pendant la saison de la remontée d'eau de fond. Le reste de l'année elles sont probablement au-dessous du thermocline en des eaux plus profondes. Le meilleur indice d'abondance dans ce cas serait la capture par unité d'effort au plein de la saison de pêche.
Pour Sardinella il est évident qu'un indice d'abondance de la saison principale dans une année ne peut pas être comparé à un indice de la morte-saisson d'une autre année. C'est une règle générale qu'il ne faut pas faire cela; les indices de la même saison ou les moyennes de toute l'année sont préférables.
L'effet des changement diurnes a déjà été relevé dans la section 9.2.2.2.
Cette sous-section a traité de la question “combien de poissons sont présents”. Non seulement des méthodes pour déterminer cela ont été décrites mais aussi les erreurs systématiques inhérents aux méthodes d'échantillonnage utilisées. Les résultats de ces méthodes donnent aussi les réponses à “quelles espèces sont présentes” et à “quand et où elles sont disponibles pour être capturées”. Dans une pêcherie en développement la détermination de la composition en espèces disponibles décidera non seulement des meilleures méthodes de capture mais aussi des débouchés pour le produit. Savoir quand le poisson est disponible est aussi d'une importance vitale; une seule exploration effectuée pendant une période où les poissons étaient disponibles au maximum conduirait à une conclusion totalement fausse sur la viabilité d'une pêcherie potentielle. Savoir où les poissons sont disponibles détermine aussi le choix des engins et des unités de pêche. C'est la période de “Prospection”; maintenant nous nous tournons vers “L'évaluation des stocks” (Fig. 1.1).
La dynamique des populations est l'étude de l'effet de la pêche sur le poisson. La grande majorité des pêcheries s'effectue sur des stocks sauvages sur la reproduction, la croissance et la mortalité naturelle desquels l'homme n'a aucun contrôle. La seule façon selon laquelle il peut, d'une manière prévisible, affecter ces stocks est par le type de pêche qu'il adopte, bien qu'il puisse aussi affecter les trois autres facteurs d'une manière imprévisible.
Quand un homme veut prédire les effects de son interférence avec un système, il doit en construire un modèle mathématique. Le nombre de facteurs influençant un tel système est très élevé, et au début il est nécessaire de simplifier le modèle en laissant de côté plusieurs facteurs et en se concentrant sur ceux présentant le plus d'intérêt. Même si le modèle est très simple et même si on trouve très tôt qu'il ne décrit pas très bien le système, il peut être asses utile au chercheur en l'aidant à mieux comprendre la dynamique du système. C'est seulement en construisant et utilisant un modèle qu'il sera capable de poser des questions pleines de sens et de recueillir les données de la plus grande utilité à ce moment. (voir aussi section 2)
Russell (1931) a été un des premiers à exprimer les facteurs qui affectent la dimension d'un stock de poissons par la formule:
S2 = S1 = (A + C) - (C + M)
dans laquelle S1 = la dimension du stock au début de l'année
S2 = la dimension du stock à la fin de l'année
A = recrutement
G = croissance
C = capture
M = morts dues à des causes naturelles
Si le stock est en équilibre (S2 = S1), puis A + C = C + M ou C = A + G - M. C'est une façon très simple de considérer les facteurs qui gouvernent la dimension du stock mais la formule ne peut pas être utilisée telle quelle et dans les quarante dernières années les biologistes halieutiques ont décrit divers modêles pour exprimer les termes dans l'équation au moyen de paramètres qui peuvent être déterminés à partir d'observations. Comment obtenir ces informations? tel est le sujet du présent manuel. En plus il a indiqué comment déterminer l'effet des changements du paramètre que l'homme a potentiellement le pouvoir de contrôler, à savoir la quantité d'activité de pêche. Dans le cas simple on suppose que nombre de recrues, le taux de croissance d'un poisson individuel et la chance de survie d'un individu ne sont pas affectés par la densité du stock; en d'autres termes qu'ils sont les mêmes que la pêche soit nulle ou très grande.
On peut trouver les longueurs et poids moyens d'après les déterminations de l'âge, la longueur et le poids; ce sont les produits finaux des systèmes d'échantillonnage décrits dans les sections 2, 3 et 4. A partir de ceux-ci on peut contruire un modèle pour exprimer la longueur ou le poids comme une fonction de l'âge. La fonction du poids est la courbe A dans la Fig. 9.3. De la même façon, à partir des déterminations de l'âge et d'après les captures exprimées en nombres par unité d'effort on peut trouver la densité relative pour une classe d'âge à différentes occasions pendant sa vie et construire un modèle pour exprimer le nombre d'espèces comme une fonction du temps. Cette fonction apparait comme la courbe B dans la Fig. 9.3. Le produit du nombre présent à un certain âge et du poids moyen à cet âge donne la biomasse à ce moment (Courbe C, Fig. 9.3). La courbe C a un maximum et pour obtenir la capture maximum la classe d'âge doit être pêchée à cet âge. En pratique c'est impossible car cela nécessite un effort infiniment grand pendant une courte durée et pas de pêche pendant le reste de l'année. Aussi, les classes d'âge sont mélangées et il est impossible de les pêcher isolé ent. Ce modèle simple est insuffisant. Pour des raisons économiques l'effort doit être distribué sur toute l'année et débute donc en prenant sa part avant que le maximum de la biomasse soit atteint et continue après que le déclin a commencé. La pêche s'effectue aussi sur plus d'une classe d'âge à la fois et le modèle doit être amplifié pour tenir compte de cela.
Graham (1939) a décrit la courbe B (Fig. 9.3) d'une façon très simple en prenant un nombre initial de poissons, soit 1 000 et en calculant les survivants après un an pour un taux donné de mortalité totale, soit 50 % par an. A la fin de la première année il y aurait 500 survivants, à la fin de la seconde 250, à la fin de la troisième 125, et ainsi de suite. Le déclin est appelé “exponentiel” et peut être calculé plus facilement en utilisant les fonctions exponentielles, celles basés sur la puissance de la constante e. (La détermination et l'emploi de cette constante sont décrits dans la plupart des ouvrages sur le calcul). Cependant, c'est une méthode fastidieuse de calculer l'effet de la pêche et éventuellement les équations de rendement sont dérivées par Ricker (1948) et par Beverton et Holt (1957) qui ont permis de combiner en une seule équation les taux de mortalité et la croissance en poids (voir aussi Gulland, 1969).
Ces équations de rendement sont simplement des moyens pour intégrer les taux de croissance, de pêche et de mortalité naturelle pour donner des évaluations du rendement pour chacun des facteurs les deux autres étant constants. Comme les modèles sur lesquels elles sont basées supposent un recrutement constant les résultats qu'elles donnent sont en termes de rendement par recrue (Y/R). Un aperçu simple de ce modèle est donné dans la Fig. 9.4a. Ces modèles sont appelés déterministes, chaque facteur est fixé pour tout le cycle de la pêcherie et aucun d'eux n'est altéré par des changements des autres facteurs. Ces modèles ont été employés dans les pêcheries pendant les 20 dernières années et ont été très utiles dans l'évaluation des stocks. Ils ont permis aux chercheurs halieutiques de déterminer à quel point la pêche est en relation avec le rendement soutenable à long terme et l'effort de pêche (Fig. 1.1, points C, D. et E ou tout autre point), et de décrire les effects du changement de ce niveau d'effort de pêche (ou plus précisement du taux de mortalité due à la pêche). Ils ont aussi permis de calculer les effets de l'âge auquel les poissons sont capturés en premier (âge de recrutement) (Fig. 9.4a, b). L'inconvénient de ces modèles est qu'ils ne prédisent que qualitativement parce que toutes les réponses sont en termes de production par recrue. Des prévisions quantitatives dépendent de la connaissance soit du nombre de recrues, soit de l'utilisation de modèles qui incorporent la structure des âges du stock en nombres.
Dans les cinq dernières années les analyses de population virtuelle (section 9.2.1.5) ont été largement utilisées pour nous donner cette information, et utilisées avec des estimations du recrutement obtenues d'après des études des larves et des pré-recrues de poissons elles ont permis aux biologistes halieutiques de prédire quantitativement. Dans la CIPAN et jusqu'à un certain point dans la Commission des pêcheries de l'Atlantique du Nord-Est (CPANE) la pêche est contrôlée par des quotas qui ont été calculés de cette facon. Le traitement des données a aussi été rendu plus facile par l'emploi de calculateurs électroniques.
Les modèles qui permettent des variations des paramètres sont appelés “aléatoires” par comparaison à ceux qui sont “déterministes”. Ces modèles sont complexes et leur étude est difficile sans l'emploi de calculateurs électroniques mais ils permettent d'étudier les effects possibles de la dimension du stock sur les taux de croissance, les taux de mortalité et le recrutement (Fig. 9.4d). On ne connait rien des variations des taux de mortalité naturelle avec la dimension du stock mais on sait que les taux de croissance de plusieurs espèces dépendent de la densité du stock. Cependant, on a porté beaucoup d'attention à la question de la dépendance du recrutement sur la dimension du stock.
A présent c'est un sujet de controverse si la pêche peut ou ne peut pas affecter le nombre de recrues du stock pêchable d'une espèce de poissons téléostéens. Pendant longtemps on a pensé que dans la gamme d'abondance dans laquelle une pêcherie serait économiquement viable, le nombre de reproducteurs serait toujours suffisant pour produire suffisamment d'oeufs pour remplacer les pertes dues à la pêche et à la mortalité naturelle. Récemment cette supposition a été mise en question. Il semble hautement probable que les pêcheries de certains stocks comme le hareng de Hokkaido-Sakkalin, le hareng de Norvège, la sardine japonaise et la sardine de Californie se sont effondrées parce que le nombre de femelles adultes n'était plus suffisant pour maintenir le stock à un niveau où la pêcherie était profisable (Cushing, 1971). Pour incorporer les effects de la pêche sur le recrutement dans les modèles il pourrait être nécessaire de connaître à la fois la fécondité (section 5) et l'histoire biologique du poisson larvaire (section 7). D'autres utilisations qui peuvent être faites des données de base sont les études sur l'interaction entre les pêcheries et sur l'interaction entre les espèces dans la même pêcherie. Cette dernière a été mentionnée dans la section 6.
Le but final du chercheur halieutique est d'aider à maintenir le stock au niveau qui a été choisi (B, C, D ou E ou tout autre niveau, Fig. 1.1). Généralement, le niveau choisi est D ou à quelque point situé entre D et E. Pour ce faire il doit le surveiller (régulièrement relever toutes les données deorites dans les sections 2, 3 et 4 et conduire des études de pré-recrues) de façon à pouvoir prédire quelles captures peuvent être prélevées sur le stock sans affecter ses capacités de fournir des captures à ce niveau, dépendant du recrutement. Il doit aussi être capable de conseiller sur n'importe quels problèmes qui peuvent surgir soit avec l'introduction de nouvelles méthodes de pêche, soit de changements naturels dans la disponibilité ou l'abondance du stock.
Les richesses de la mer en dehors des limites sont ouvertes à la pêche de n'importe qui et l'exploitation des ressources vivantes de la mer devient de plus en plus importante pour un nombre croissant de pays. Les problèmes n'atteignent pas la même intensité dans les pêcheries en eau douce mais ils y existent aussi.
La coordination internationale est nécessaire pour protéger ces ressources de la surexploitation. Il ne rime à rien qu'une nation réglemente ses captures dans une pêcherie internationale si auoun autre nation n'en fait autant parce que la nation qui réglemente va perdre alors que les autres tient profit. Seulement, si chaque nation doit gagner également, parce que le modèle de ses chercheurs leur montre que cela augmentera le total généralement après quelques pertes initiales qui doivent aussi être partagés également les nations auront à coopérer.
Comme un stock de poissons est souvent exploité par plusieurs flottes de différents pays il est nécessaire que tous les pays relèvent des statistiques de pêche et de préférence de la même façon. Cela a été souligné dans les sections 2, 3 et 4. Pour d'autres études il n'est pas essentiel de relever des données d'une manière standard, bien qu'une tells pratique facilite les échanges d'informations. La coordination du travail est essentielle si plus d'une nation pêche sur le stock, si ce n'est que pour déterminer la capture totale qui provient du stock. Dans beaucoup de cas les nations pêchent différentes parties du stock de sorte que les captures par unité d'effort d'une flotte peuvent ne pas être représentatives de l'abondance du stock entier. Par exemple, la flotte chalutière britannique pêche dans la mer de Barentz sur la partie immature du stock de morues Arcto-norvégien, ces poissons sont âgés principalement de 3 à 7 ans. Les norvégiens pêchent ce stock aux îles Lofoten où il se reproduit et leurs captures se composent de poissons âgés de plus de 7 ans. Il y a une série d'indices élevés de capture par unité d'effort quand de grandes classes d'âge traversent la pêcherie. Initiallement c'est élevé dans la pêcherie britannique et ensuite élevé dans la pêcherie norvégienne. Pour des analyses de population virtuelle il est essential d'avoir la composition complète des âges pour les captures, ce qui signifie que ces nations qui pêchent le plus de poissons doivent fournir des données avant que les analyses puissent être tentées. La coordination du travail peut aussi rendre des études faisables en diminuant le coût pour chaque état individuel; les études du groupe O des téléostéens marins sont largement effectuées sur une base internationale.
La tâche de combiner les résultats de nombreux programmes de recherches et de les faire parvenir aux gouvernements qui dirigent la politique des pêches dans les différents pays est en grande partie entreprise par un certain nombre d'organisations internationales, surtout à travers les Commissions régionales se composant des pays exploitant la région. Un compte-rendu historique du développement de la collaboration dans les recherches halieutiques avec référence particulière à la région nord-atlantique est donné par Lucas (1963) et une revue des organisations internationales a été publiée par le U.S. National Oceanographic Data Center, 1969 sous le titre “Annotated Acronyms and Abbreviations of Marine Science Related International Organizations”.
Alverson, D.L., 1971 Manual of methods for fisheries resource survey and a praisal. Part 1. Survey and charting of fisheries resources. FAO Fish. Tech. Pap., (102):80 p.
Beverton, R.J.H. et S.J. Holt, 1957 On the dynamics of exploited fish populations. Fish. Invest. Lond. (2), (19):533 p.
Christensen, O., 1970 Undersøgelser i forbindelse med spørgsmålet om sommerfredning af laks i Østersøen. Skr. Dan. Fisk. Havunders., 3):26–33
Cormack, R.M., 1968 The statistics of capture-recapture methods. Oceanogr. Mar. Biol., 6:455– 506
Cushing, D.H., 1968 Fisheries biology. London, University of Wisconsin Press, 200 p.
Cushing, D.H., 1971 The dependence of recruitment on parent stock in different groups of fishes. J. Const. Int. Explor. Mer., 33:40–62
Forbes, S. et O. Nakken, (Eds), 1972 Manual of methods for fisheries resource survey and appraisal. Part. 2. The use of acoustic instruments for fish detection and abundance estimation. FAO Man. Fish. Sci., (5):138 p.
Fry, F.E.J., 1957 Assessment of mortalities by the use of the virtual population. ICNAF/ICES/ FAO Special Scientific Meeting, Lisbon, 1957. Paper P.15 (mimeo)
Graham, M., 1939 The sigmoid curve and the overfishing problem. Rapp.P.-V. Réun. Cons. Perm. Int. Explor. Mer, 110:15–29
Gulland, J.A., 1965 Estimation of mortality rates. Annex to Arctic Fisheries Working Group Report. Meeting in Hamburg, January 1956. ICES CM 1965, Doc.3 (mimeo)
Gulland, J.A., 1969 Manual of methods for fish stock assessment. Part 1. Fish population analysis. FAO Man. Fish. Sci., (4):154 p.
Holden, M.J., 1963 The population of fish in dry season pools of the River Sokoto. Fishery Pub. Colon. Off. Lond., (19):58 p.
Holden, M.J., 1973 Report of the ICES/ICNAF Working Groups on Selectivity Analysis. Coop. Res. Rep. ICES(A), (25):144 p.
Holt, S.J., 1963 A method of determinin gear selectivity and its application. Spec. Publs. ICNAF., (5):106–15
Jones, B.W. et J.G. Pope, 1973 A ground fish survey of Faore Bank. Res. Bull. ICNAF, (10):53– 61
Lucas, C.E., 1963 International collaboration in fisheries research. A historical review, with particular references to the North Atlantic Region. FAO Fish. Biol. Tech. Pap., (33):6 p.
Mackett, D.J., 1973 Manual of methods for fisheries resource survey and appraisal. Part 3. Standard methods and techniques for semersal fisheries resource surveys. FAO. Fish. Tech. Pap., (124):39 p.
Margetts, A.R., 1949 Experimental comparison of fishing capacities of Danish-seiners and trawlers. Rapp. P.-V. Réun. Cons. Perm. Int. Explor. Mer, 125:82–90
Pope, J.A., 1966 Manual of methods for fish stock assessment. Part. 3. Selectivity of fishing gear. FAO Fish. Tech. Pap., (41):41 p.
Pope, J.A., 1972 An investigation of the accuracy of virtual population analysis using cohort analysis. Res. Bull. ICANF (9)65:74
Ricker, W.E., 1948 Methods of estimating vital statistics of fish populations. Indiana Univ. Publ. Sci. Ser., (15)
Russell, E.S., 1931 Some theoretical considerations on the “overfishing” problem. J. Cons. Perm. Int. Explor. Mer, 6:3–20
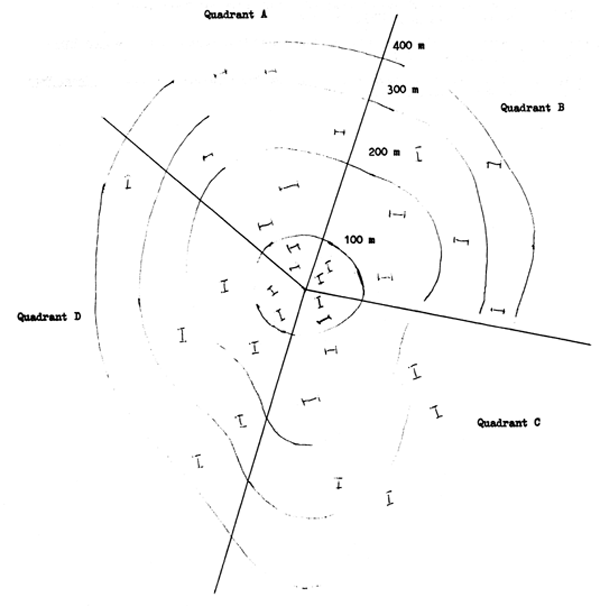
Fig. 9.1 Stratification de banc des Feroe par des quadrants A-D, chacun ayant les quatre mêmes zones de profondeur, moins de 100 m, 101–200 m, 201–300 m et 301–400 m. Traits de chalut choisis sur un quadrillage de stations potentielles par des nombres pris au hasard. Modifié par Jones et Pope (1974)
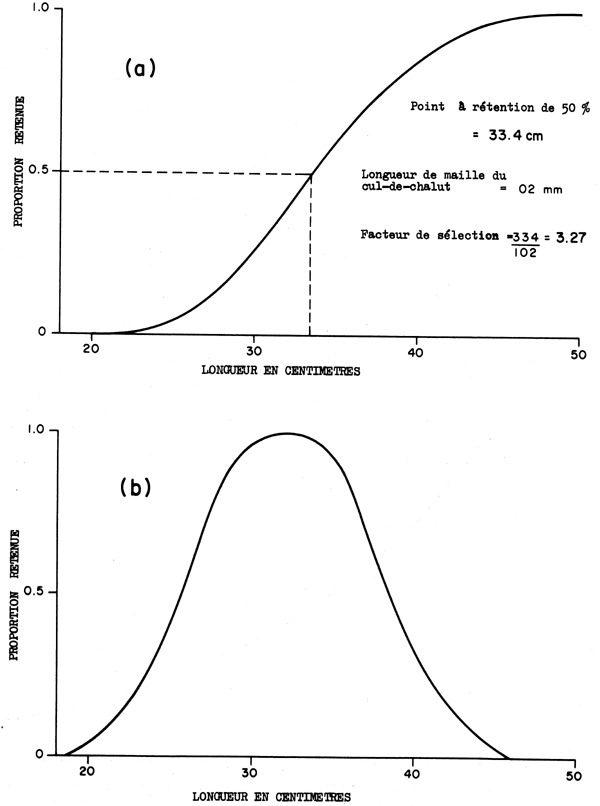
Fig. 9.2. Courbe de sélection de cul-de-chalut (a) et de filet maillant (b)
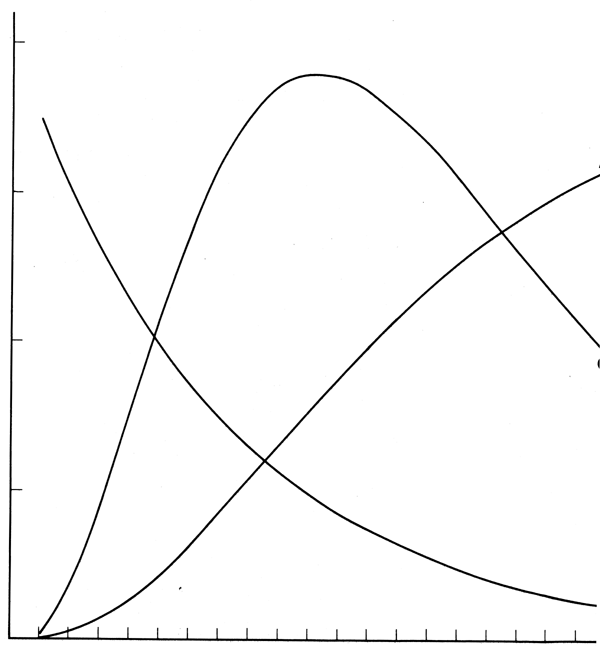
Fig. 9.3. Courbes montrant (A) augmentation de poids d'un poisson individuel, (B) déclin en nombre d'une classe d'âge, et (c) biomasses tatale d'une classe d'âge, le produit de (A) x (B), avec le temps (t)
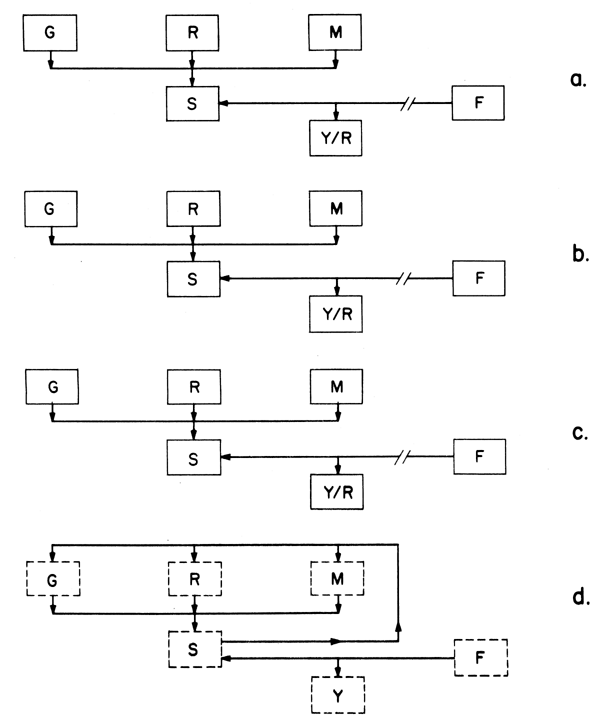
Fig. 9.4. Diagrammes sohématiques de modèles d'évaluation: G = croissance, R = recruitement, M _ taux de mortalite naturelle, F = taux de martalité due à la pêche, S = stock, Y = production, Y/R = production par recrue; a,b,c, sont basé s sur un modèle déterministique dans lequel F na pas d'effet sur G, R ou M. a) éltata initial, (b) longueur de maille augmentée (indiqué e par // ) et augmentation de Y/R, c) le taux de mortalité due à la pêche augmente et Y/R diminue, (d) représente le modè “stochastique” dans legal l'â ge de S affecte G,R et M ainsi que Y.
DOCUMENTS TECHNIQUES FAO SUR LES PÊCHES
Les documents de cette série constituent des éditions provisoires de volumes ou de chapitres des manuels FAO de science halieutique, publiés normalement dans la langue du manuscript original. Initialement, ils sont diffusés seulement aux collaborateurs participant au projet correspondant en vue d'obtenir leurs avis et commentaires, ainsi qu'au personnel de la FAO - du siège et sur le terrain - susceptible de les utiliser lors de stages, cours et séminaires de formation. Une fois mis au point, ils sont publiés par la FAO dans les langues officielles de I'Organisation.
Documents publiés depuis janvier 1973
| FIRM/T118 (En) | Manual of fisheries science. Part 1 - An introduction to fisheries science | February 1973 |
| (Es) | Manual de ciencia pesquera. Parte 1 -Una introducción a la ciencia pesquera | mayo 1973 |
| (Fr) | Manuel de science halieutique. Première partie - Introduction à la science halieutique | juillet 1974 |
| FIRD/T124 | Manual of methods for fisheries resource survey and appraisal. Part. 3-Standard methods and techniques for demersal fisheries resource surveys | September 1973 |
| FIRS/T115 (En) (Rev. 1) | Manual of fisheries science. Part 2 - Methods of resource investigation and their application | June 1974 |
| (Fr) | Manuel de science halieutique. Deuxième partie - Méthodes de recherches sur les ressources et jeur application | juillet 1974 |
![]()
![]()