![]()
![]()
par
INTRODUCTION
Deux espèces de seiches sont capturées par les pêcheries séné-gambiennes de céphalopodes: Sepia officinalis hierredda et Sepia bertheloti. Cette dernière espèce constituant moins de 1% des prises, c'est Sepia officinalis hierredda qui retiendra ici notre attention d'autant plus qu'elle représente plus de 95% des mises à terre de céphalopodes.
L'exploitation de cette seiche a commencé au Sénégal en 1973 par une flottille de chalutiers sénégalais. Celle-ci a été renforcée en 1975 par une flottille piroguière.
Dans ce travail, nous analyserons l'évolution de ces pêcheries et ferons le point des connaissances sur la biologie et l'identité des populations exploitées de ce mollusque sur les côtes sénégalaise et gambienne (figure 1).
1. EVOLUTION DE LA PECHERIE DES SEICHES DES COTES SENEGALAISES
1.1 EVOLUTION DES FLOTTILLES
La flottille chalutière est composée principalement de chalutiers glaciers opérant en boeuf et de quelques chalutiers-congélateurs classiques (tableau 1), tous spécialisés dans la pêche de la seiche.
Quant à la flottille artisanale piroguière, elle ne recherche pas systématiquement cette espèce. Elle développe son effort sur elle esentiellement au cours du premier semestre quand le mollusque gagne les petits fonds pour la reproduction. La pêche s'effectue alors à la turlutte et au casier. Nous ne connaissons pas avec exactitude l'effectif des pirogues qui s'adonnent à cette activité pendant la période favorable.
Nous pensons toutefois qu'elle avoisine 300 unités.
1.2 LES STATISTIQUES DE PECHE
1.2.1 Les prises
Les prises de seiche augmentent depuis 1973, aussi bien pour la pêche industrielle chalutière que pour la pêche artisanale piroguière (tableau 2). On note toutefois que les statistiques des années 1973–1975 sont peu précises.
Par ailleurs, la part des captures piroguières dans les prises globales ne cesse de croître passant de 22% en 1975 à 36% en 1977 et 46% en 1980.
1.2.2 Les rendements
Les chalutiers-boeufs étant plus spécialisés dans la pêche des seiches et effectuant plus de 40% des captures, leurs rendements ont été retenus comme indice d'abonance apparente.
Les figures 2 et 3 donnent les évolutions mensuelles et annuelles de ces rendements. On note qu'en 1980 ceux-ci ont été surtout élevés d'avril à juillet et se sont maintenus au même niveau annuel qu'en 1979.
1 Chercheur au Centre de recherches océanographiques de Dakar-Thiaroye (ISRA) B.P. 2241
Dakar, Sénégal
2 Biologist, Fisheries Department, 6 Marina Parade, Banjul, La Gambie
2. REPARTITION, REPRODUCTION ET CROISSANCE DE LA SEICHE SUR LES COTES SENEGALAISES
2.1 REPARTITION ET MIGRATIONS
2.1.1 Répartition
Les études réalisées au Centre de recherches océanographiques de Dakar-Thiaroye (Bakhayokho, 1980) montrent que les seiches se rencontrent sur toute la côte séné-gambienne, mais qu'elles sont plus abondantes sur la côte sud du Cap-Vert à la Casamance, par des fonds inférieurs à 50 m.
C'est d'ailleurs dans cette zone qu'opèrent les chalutiers et les pirogues, ces dernières travaillant plus vers la côte que les premiers.
Il faut noter aussi que les seiches de grande taille (supérieure à 30 cm de longueur du manteau dorsal) sont capturées uniquement sur la Petite Côte et principalement sur les faibles profondeurs, de janvier à avril. Elles ne sont jamais prises en Casamance.
2.1.2 Migrations
A l'intérieur de la zone d'abondance maximale définie ci-dessus, on observe des migrations côte-large et des migrations nord-sud.
Les seiches rejoignent en effet les petits fonds de janvier à juin et y effectuent des déplacements nord-sud. Elles regagnent ensuite le large au second semestre en migrant vers le nord (figure 4).
2.2 REPRODUCTION
2.2.1 Les périodes de reproduction
Les seiches se reproduisent de janvier à septembre sur les petits fonds, les grandes de janvier à avril, les petites et les moyennes de mai à septembre. La période de ponte intense se situe cependant de février à mai.
Elles fixent leurs oeufs sur les algues, herbes marines et débris solides allongés variés que l'on trouve principalement de Joal à Sangomar et en face des fleuves.
2.2.2 Echelle et taille à la première maturité
Une échelle de maturité a été étudiée (tableau 3) et utilisée pour déterminer la taille à la première maturité des femelles. Celle-ci se situe à 13,5 cm.
2.2.3 Fécondité
La fécondité a été estimée par le nombre d'oeufs en voie de maturation dans les ovaires. Sa relation avec la taille est donnée (figure 6) par la formule suivante:
Fe = 2,59 L1,77
où Fe = le nombre d'oeufs pondus par an
et L = la longueur du manteau dorsal en cm.
La fécondité varie donc de 250 oeufs pour les femelles de 13 cm à 1 400 oeufs pour celles de 35 cm (taille maximale des femelles). La fécondité relative résultante est de 0,52 oeufs par gramme de poids.
2.2.4 Capacité de reproduction
La capcité relative de production d'oeufs calculée suivant la taille révèle que ce sont les individus de 14 à 23 cm qui participent le plus à la reproduction (figure 7) dans la mesure où les captures constituent un échantillon représentatif de la population.
2.2.5 Sex ratio
Le premier trimestre correspond à la période où l'on rencontre toutes les tailles de seiches.
Un échantillon de 5 356 individus mesurés à bord d'un chalutier commercial en mars 1979 dans cette zone comprenait les pourcentages de femelles suivant la taille indiqués sur la figure 5.
Les faibles valeurs situées entre 20 et 23 cm et au-delà de 32 cm sont dues au fait qu'il y aurait deux classes d'âges (de 0 à 23 cm et de 24 à 40 cm) et que pour chacune d'elles les individus de plus grande taille sont constitués de mâles qui auraient une croissance plus rapide que les femelles.
On note que le sex ratio observé dans les captures varie au cours de l'année.
2.3 PREMIERE TENTATIVE D'ESTIMATION DE LA CROISSANCE
Le premier mode qui apparaît dans les histogrammes obtenus à partir des captures des chalutiers se situe à 9 cm de longueur de manteau dorsal.
La croissance a été étudiée chez les mâles, de 9 à 25 cm par la méthode de Petersen.
Connaissant les périodes d'éclosion, la croissance a été également estimée à la naissance, à 9 cm.
Les équations suivantes ont été obtenues (figure 8) :
L = 2,16 t + 10,10 (1) = intervalle d'observation de 9 à 25 cm
L = 2,06 t + 0,7 (2) = intervalle d'observation de l'éclosion à 9 cm
L en cm et t en mois.
La croissance en poids résultant de ces équations et de la relation taille-poids s'écrit (figure 9) :
W = 11,85 t2,15
avec W en gramme et t en mois.
En supposant que la croissance est identique pour les deux sexes jusqu'à la taille à la première maturité (13,5 cm), les femelles commenceraient leur première maturation sexuelle à l'âge de 5 à 6 mois.
2.4 RELATION TAILLE-POIDS
La relation entre la taille et le poids a été calculée à partir de 612 mâles et 471 femelles échantillonnés au cours de toute l'année 1979 sur la Petite Côte. Les résultats figurent dans le tableau ci-après:
| Nombre de seiches | Type d'ajustement | Coefficient corrélation | Equation des courbes d'ajustement F en g et L en cm | |
|---|---|---|---|---|
| Mâles | 612 (5 à 44 cm) | axe majeur réduit | 0,99 | P = 0,26 L2,74 ± 0,03 |
| Femelles | 471 (7 à 37 cm) | axe majeur réduit | 0,99 | P = 0,26 L2,75 ± 0,03 |
3. IDENTITE DES POPULATIONS DE SEICHES EXPLOITEES SUR LES COTES SENE-GAMBIENNES
L'étude de la répartition et de l'abondance des seiches sur la Petite Côte et en Casamance, à partir de statistiques de pêche chalutière (Bakhayokho, 1980) a conduit à émettre l'hypothèse qu'il pourrait exister deux stocks de part et d'autre de la Gambie.
En effet, les schémas de migration (figure 4) résultant des variations spatio-temporelles de l'abondance et des tailles (présence de grandes, petites et moyennes seiches sur la Petite Côte mais présence uniquement de petites et surtout de moyennes seiches en Casamance) sont différentes dans les deux zones.
Cependant, en absence d'études biologique, biométrique et éthologique des individus pêchés sur la Petite Côte, en Gambie et en Casamance, il est difficile de confirmer ou d'infirmer cette hypothèse.
Une campagne conjointe de chalutages de prospection effectuée en juillet et en août 1981 sur le plateau continental séné-gambien par le Centre de recherches océanographiques de Dakar-Thiaroye et le “Fisheries Department” de la Gambie, à partir d'un chalutier spécialisé de l'armement sénégalais SENEPESCA, a conduit aux résultats suivants sur la répartition et la biométrie de Sepia officinalis hierredda.
3.1 REPARTITION
Les seiches sont présentes sur toute la côte sud de Séné-Gambie. Cependant, les concentrations et les tailles varient suivant la latitude (figures 10, 11 et 12), la profondeur (figures 13, 14 et 15) et le type de fond (figures 16 et 17).
Toutefois l'histogramme des tailles regroupées (figure 18) montre un seul mode.
Il est connu que les seiches préfèrent les fonds sableaux (et sablo-vaseux ou vaseux pour les jeunes) où elles s'enfoncent quand elles sont au repos et cherchent les algues, herbiers et débris allongés pour fixer leur ponte (Bakhayokho, 1980). Il est également connu (Domain, 1976) que la Petite Côte est très sableuse et couverte de prairies d'algues et d'herbiers alors que le littoral de la Gambie et de la Casamance est plutôt vaseux et sablo-vaseux. La sédimentologie du plateau séné-gambien serait donc à l'origine de la répartition des seiches.
3.2 LE SEX RATIO
Le sex ratio global calculé en juillet 1981 à partir de 7 038 individus de toute la côte sud séné-gambienne est de 45%. Il varie cependant suivant la zone. Sur la Petite Côte, en Gambie et en Casamance, il est respectivement de 48, 44 et 44%, les calculs étant effectués à partir de 5 489, 1 020 et 390 individus.
Cette “différence” est liée au fait que les seiches de 12 à 18 cm, les plus abondantes pendant cette période sur toute la côte (figure 18), ont un sex ratio supérieur à 50% et se trouvent plus abondamment sur la Petite Côte.
Lorsqu'on examine le sex ratio suivant la taille en fonction de la latitude (figure 19) les courbes sont comparables. Elles restent toujours comparables dans le cas du sex ratio selon la taille en fonction de la profondeur sur les fonds de 10 à 25, voire 30 m (figure 20). Quelques différences apparaissent cependant au-delà de 26 m liées probablement à la faible taille des échantillons et par suite leur non-représentativité.
3.3 RELATION TAILLE/POIDS
La relation entre la taille et le poids a été calculée pour 611 mâles de 7 à 31 cm pêchés sur la Petite Côte et pour 223 mâles de 7 à 25 cm capturés en Casamance. Les équations suivantes ont été obtenues après ajustement par la méthode des moindres carrés:
Petite Côte: P = 0,27 L2,64 ± 0,02
Casamance : P = 0,28 L2,65 ± 0,01
P est mesuré en grammes et L en centimères. On constate que les courbes sont comparables.
4. CONCLUSIONS
Depuis 1976, les captures sénégalaises de seiches tendent à augmenter.
Dans le même temps, les rendements obtenus par les chalutiers-boeufs sénégalais (seuls bateaux ayant la seiche comme cible pendant toute l'année) sont restés pratiquement constants à part une chute en 1978 provoquée peut être par une augmentation de l'effort de pêche des bateaux coréens opérant à partir de la Gambie.
Par ailleurs, la taille moyenne des prises est restée constante. Sachant que les pêches piroguières ne portent que sur les adultes, il est donc vraisemblable que la ressource ne soit pas menacée.
Cette première conclusion doit cependant être considérée avec prudence, du moins jusqu'à l'obtention de données fiables sur la production maximale équilibrée du stock séné-gambien dont nous avons tout lieu de penser maintenant qu'il est unique, compte tenu:
de la distribution continue des seiches sur le littoral;
de la structure en taille unimodale des captures juste avant le recrutement en juillet;
de l'allure du sex ratio et de la relation taille/poids qui sont comparables chez les seiches pêchées sur la Petite Côte et en Casamance.
REFERENCES
Bakhayokho, M., 1980 Historique des pêcheries de céphalopodes (seiches, poulpes, calmars) des côtes sénégalaises. Pêche marit., (1244):634–40
Domain, F., 1976 Les fonds de pêche du plateau continental Ouest-Africain entre 17°N et 12°N D.S.P.Centre de Rech.Océanogr.Dakar, (61)
| Années | 1973 | 1974 | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 |
|---|---|---|---|---|---|---|---|---|---|
| Nombre de chalutiers-congélateurs | 2 | 2 | 3 | 3 | 5 | 3 | 3 | 2 | 0 |
| Nombre de paires de chalutiers-boeufs | 0 | 1 | 1 | 2 | 3 | 3 | 3 | 3 | 3 |
| Années | 1973 | 1974 | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 |
|---|---|---|---|---|---|---|---|---|
| Production totale | 30 | 794 | 1 316 | 2 295 | 2 099 | 2 379 | 2 695 | 2 771 |
| Prises industrielles | 30 | 794 | 1 023 | 1 742 | 1 332 | 1 465 | 1 791 | 1 496 |
| Prises artisanales | 0 | 0 | 293 | 553 | 767 | 914 | 904 | 1 275 |
| Stade | Aspect de la gonade | Aspect des oeufs | Taille des oeufs |
|---|---|---|---|
| 1. immature | Gonade petite, blanc-jaunâtre, plaquée sur la face dorsale de la poche d'encre | Petits oeufs jaunâtre | Moins de 2 ans |
| 2. maturation | Gonade moyenne, jaunâtre, oeufs moyens bien individualisés sur toute ou une partie de l'extérieur de la gonade | Petits oeufs Oeufs moyens, bien visibles, réticulés ou uniformément blanchâtres, ovoides | Moins de 2 mm 2 à 4 mm |
| 3. Préponte | Gonade grosse, occupe toute la partie postérieure concave du sépion, jaune, oeufs en grappes de taille croissante de l'intérieur vers l'extérieur. Masse gélatineuse verdâtre occupe la partie postérieure de la gonade. | Petis oeufs Oeufs moyens Grands oeufs jaunes, ronds et réticulés | Moins de 2 mm 2 à 4 mm 4 à 6 mm |
| 4. Ponte | Même aspect qu'au stade 3 mais avec des oeufs lisses, libres dans l'enveloppe gonadique, ou engagés dans l'oviducte; souvent plages jaunes et blanches sur la partie postérieure de la gonade | Petits oeufs Oeufs moyens Grands oeufs Très grands oeufs, lisses et translucides | Moins de 2 mm 2 à 4 mm 4 à 6 mm 5 à 9 mm |
| 5. Post-ponte | Gonade réduite blanc-jaunâtre dans une enveloppe presque vide, flasque | Petits oeufs Oeufs moyens Grands oeufs | Moins de 2 mm 2 à 6 mm |
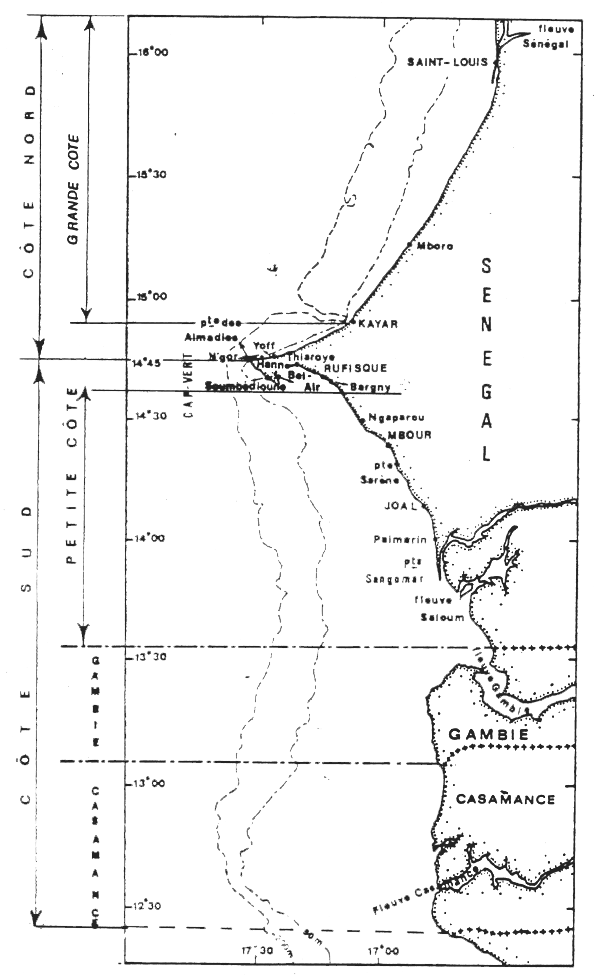
Figure 1.- La Côte séné-gambienne et les villages côtiers
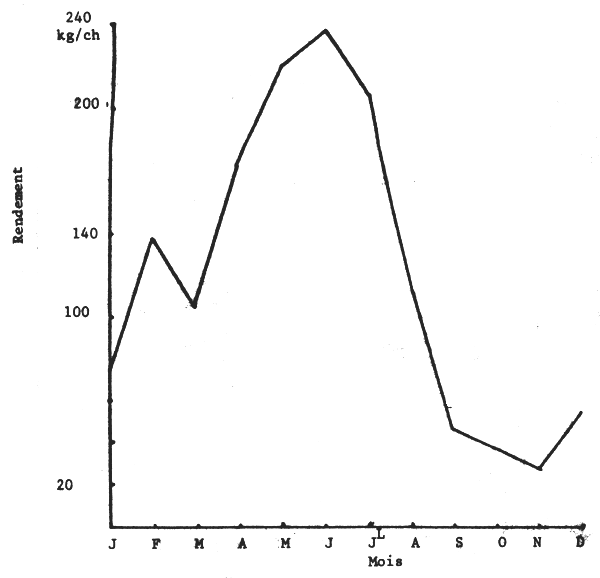
Fig. 2 - Evolution mensuelle des rendements (prises par heure de chalutage) en seiche des chalutiers-boeufs sur les côtes sénégalaises en 1980
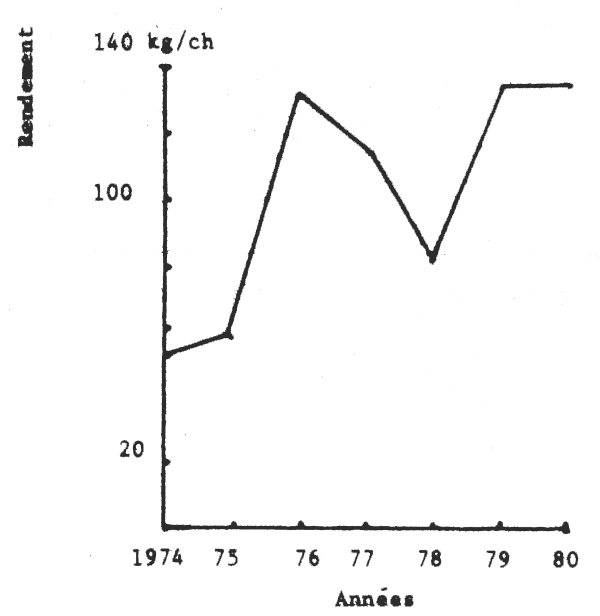
Fig. 3 - Evolution annelle des rendements (prises par heure de chalutage) en seiches des chalutiers-boeufs sur les côtes du Sénégal (1974–1980)
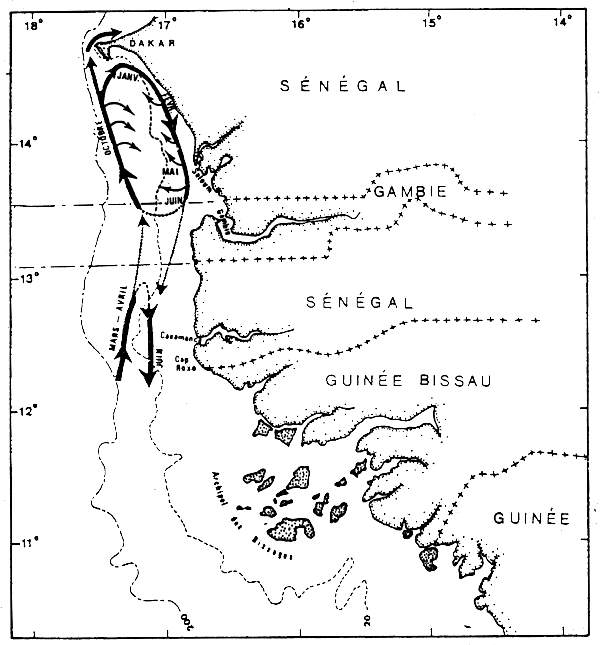
Figure 4.- Schéma de migration probable des seiches sur la Petite Côte du Sénégal
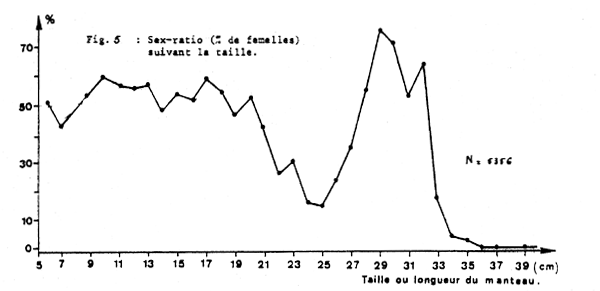
Fig. 5 : Sex-ratio (% de femalles) suivant la tasille.
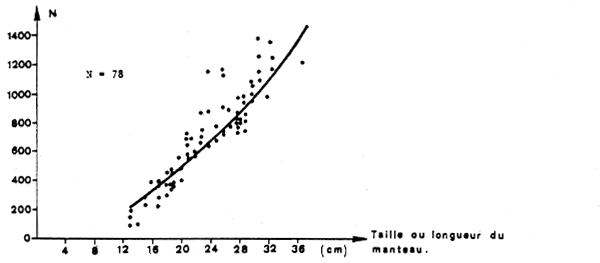
Fig. 6 : Evolution de la fécondité partielle individuelle (ou TOTAK) suivant la taille.
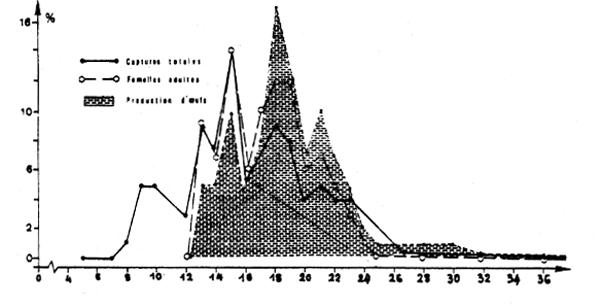
Fig. 7 : Importance relative suivant la taille : - des captures totales (femelles + mâles) - des captures totales de femelles adultes - de la production d'ceufs
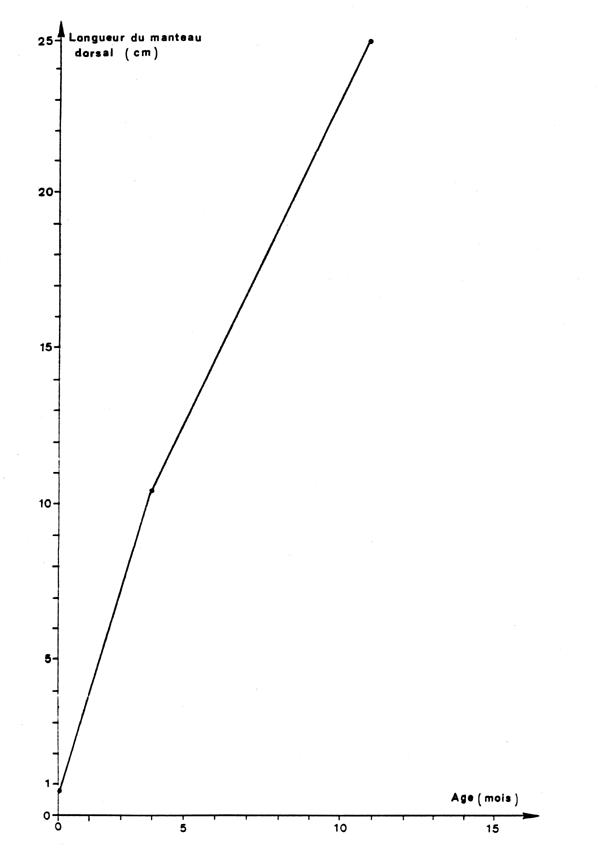
Fig. 8 : Courbe de croissance de la seiche mâle jusqu'à 25 cm de longueur du manteau dorsal
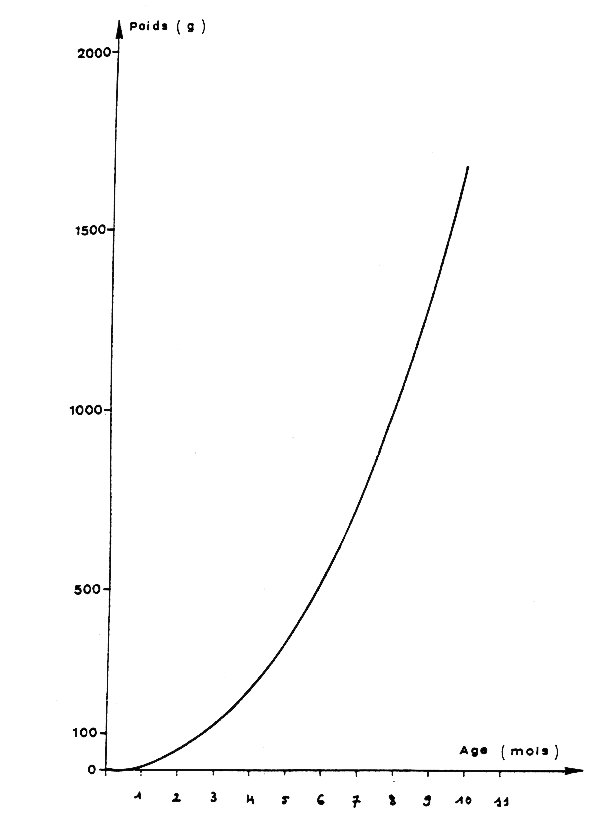
Fig. 9 .- Croissance en poids de la seiche mâle
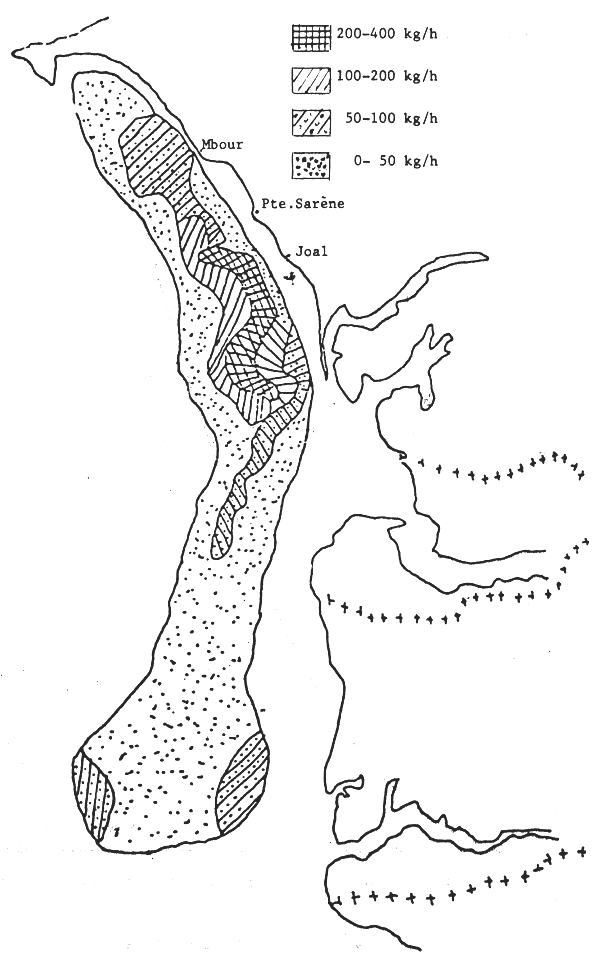
Figure 10.- Abondance apparente des seiches sur la côte sud de Séné-Gambie
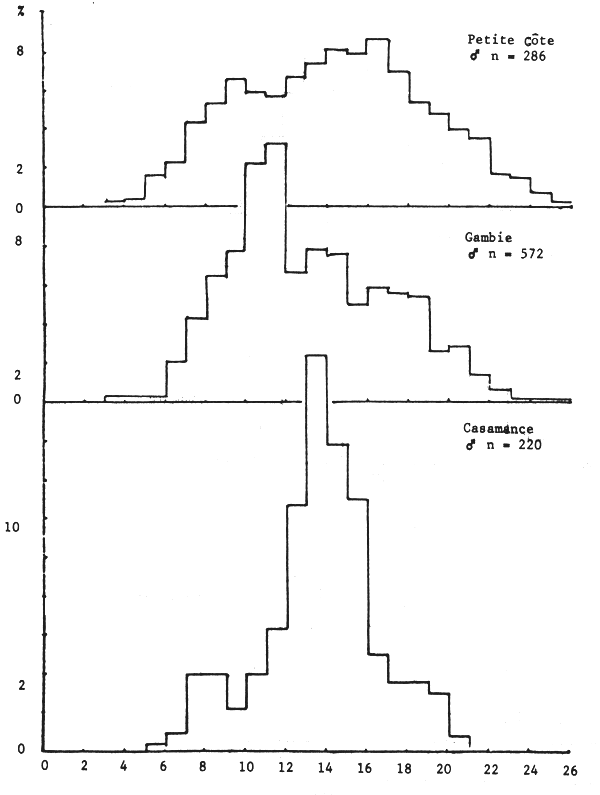
Figure 11.- Importance relative des tailles par secteur chez la seiche mâle
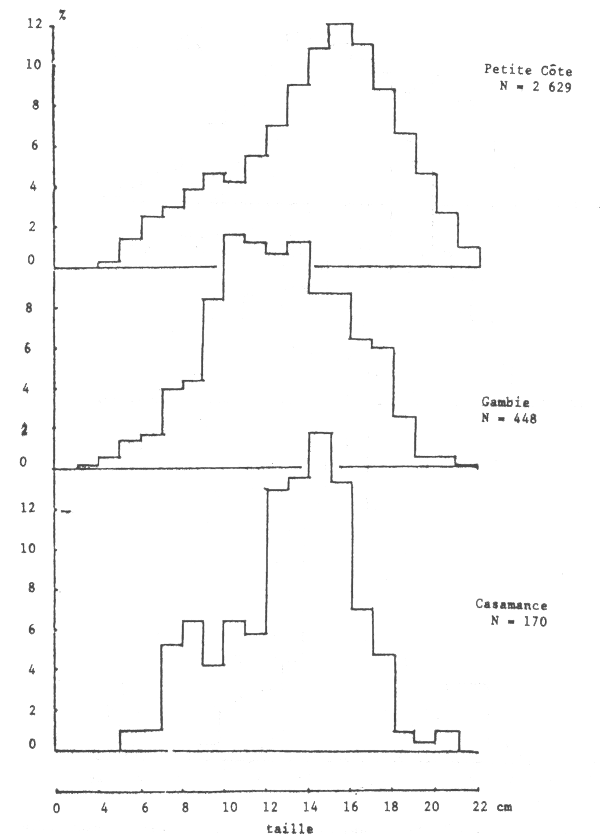
Figure 12.- Abondance relative des tailles des femelles par secteur
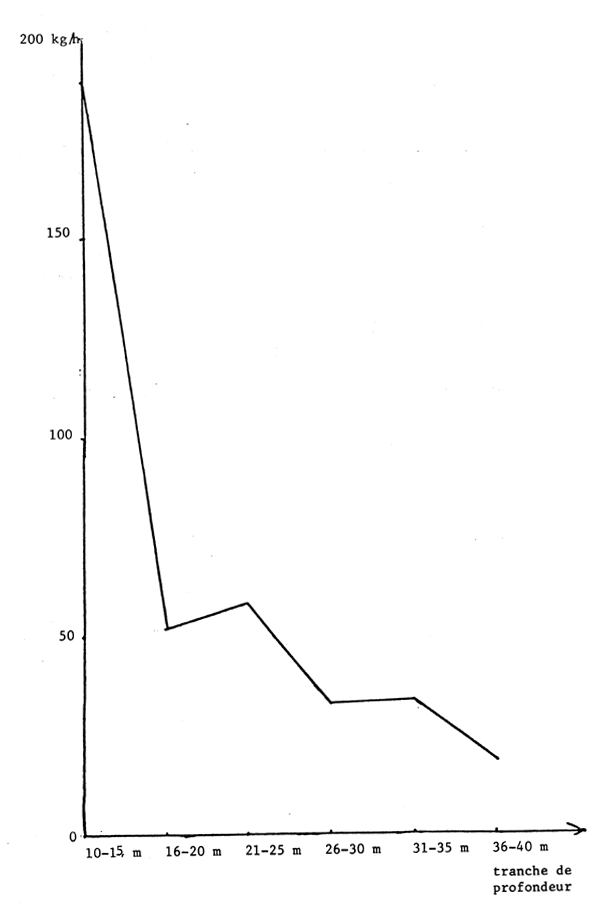
Figure 13.- Abondance apparente des seiches suivant la profondeur sur toute la côte sud
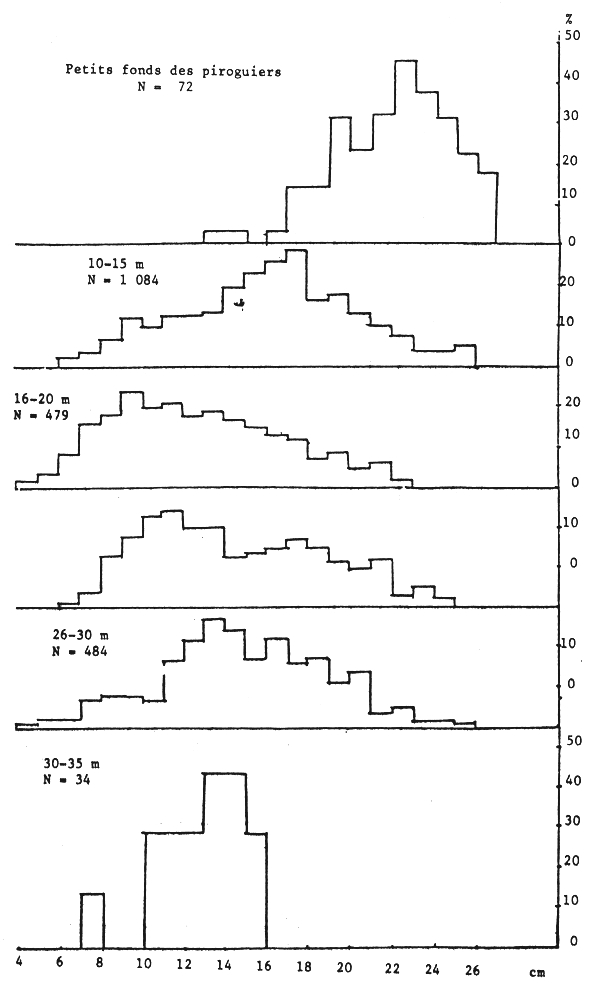
Figure 14.- Importance relative des différentes tailles de seiches mâles suivant la profondeur sur toute la côte sud
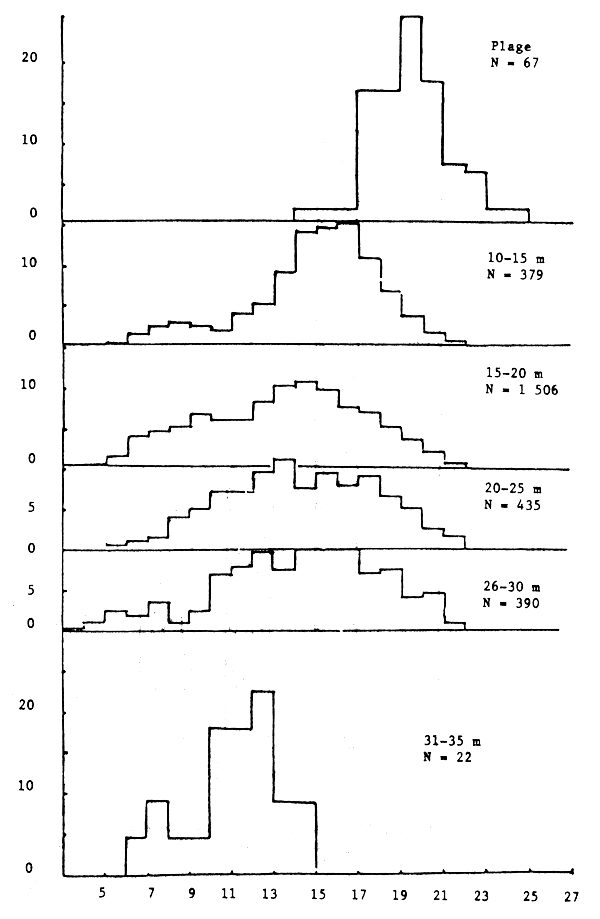
Figure 15.- Importance relative des différentes tailles de seiches femelles suivant la profondeur
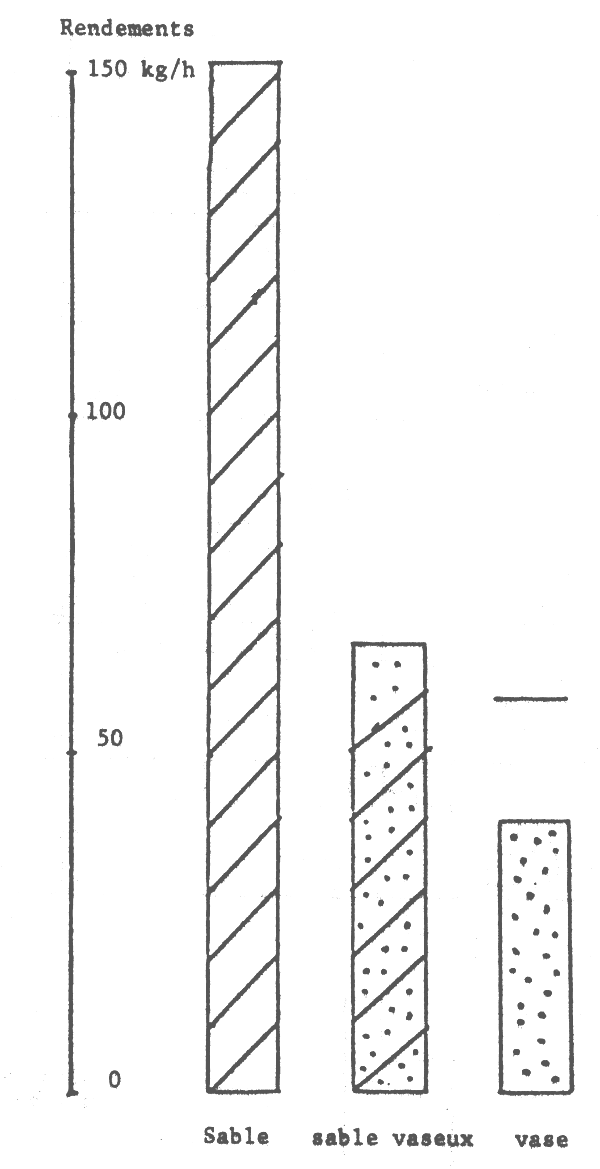
Figure 16.- Abondance apparente des seiches suivant le type de fond
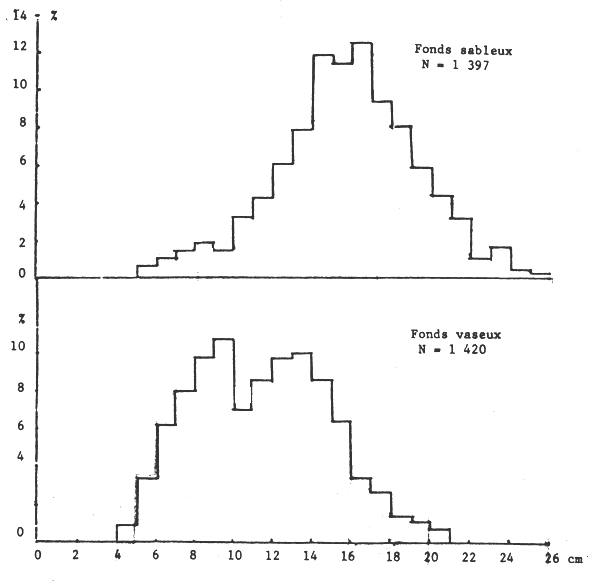
Figure 17.- Importance relative des différentes tailles suivant le type de fond (entre 10 et 40 m)
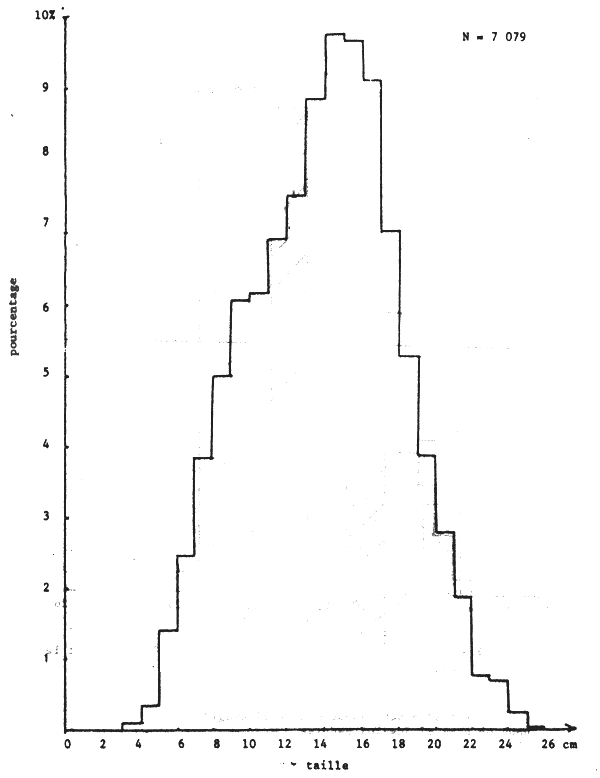
Figure 18.- Importance relative des tailles sur la côte sud
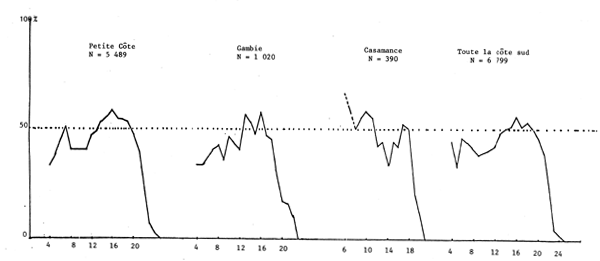
Figure 19.- Importance relative des femelles par classe de taille suivant la latitude (secteur)
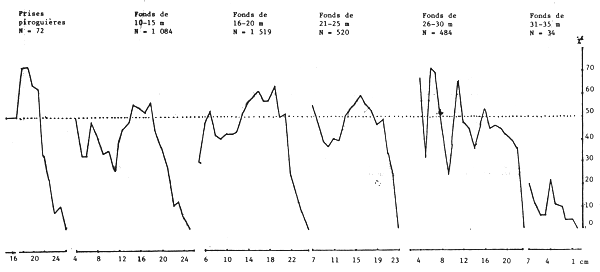
Figure 20.- Pourcentage de femelles suivant la taille par tranche de profondeur
Depuis la réunion du Groupe de travail spécial sur les stocks de céphalopodes de la région nord du COPACE (Santa-Cruz de Ténériffe, 27–30 juin 1981), de nouvelles données coréennes ont été communiquées par Joo Yeoul Lim à la sixième session du Groupe de travail du COPACE sur l'évaluation des ressources (Dakar, 2–6 février 1982). Ces données permettent notamment d'estimer comment les captures coréennes de céphalopodes et l'effort de pêche correspondants ont été répartis par stock à partir de 1977 (voir tableaux 1, 2 et 3).
Au tableau 1, les totaux pour 1978, 1979 et 1980 sont ceux estimés par Yamamoto et Ansa-Emmim (1979) et Shimura (1980 et 1981) à la base de l'échantillonnage des chalutiers à Las Palmas. Pour 1977, les nouvelles données coréennes de Lim ont été retenues puisqu'elles sont supérieures aux chiffres jusqu'à présent utilisés et résumés dans le Bulletin statistique N° 3 du COPACE (FAO, 1981). Il est également à noter que pour 1980, la répartition des captures par zone de Lim correspond assez bien à celle trouvée par Shimura. Une comparaison des données obtenues antérieures à 1980 n'est pas possible, le COPACE ne disposant pas de données sur la répartition par zone de pêche. La concordance en 1980 est néanmoins encourageante.
Le tableau 2 fournit une estimation de la répartition spatiale de l'effort coréen, basée également pour 1977 sur les données de Lim et pour 1978–1980 sur les données d'effort total obtenues par l'échantillonnage à Las Palmas.
Le tableau 3 résume les cpue coréennes en divisant les captures du tableau 1 par l'effort du tableau 2. Ces cpue nous permettent d'améliorer nos estimations de la cpue dans la zone du Cap Blanc (voir tableau 14). La série corrigée des captures, de l'effort et des cpue pour les poulpes du Cap Blanc est donnée au tableau 4.
Pour les stocks de Dakhla, une correction des captures totales obtenues par soustraction au tableau 14, s'avère également nécessaire, dans la mesure où les captures coréennes non ventilées par zone de pêche dans le Bulletin statistique N° 3 du COPACE, mais sûrement capturées dans la division statistique 34.1.3 ne sont pas incorporées dans le total de cette zone. Le tableau 5 résume les nouvelles données obtenues.
Appliquées aux modèles de production de Fox (figures 8 et 9 du rapport du Groupe de travail) pour les stocks de Dakhla et du Cap Blanc, il est apparent que ces nouvelles données ne changent pas beaucoup les résultats obtenus (figures 1 et 2): pour le Cap Blanc, le MSY passerait de 43 600 à 42 700 t pour un effort de 205 000 au lieu de 190 000 h. Pour la zone de Dakhla, le MSY augmente d'environ 5 000 t; 79 600 t contre 74 700 pour un effort correspondant de 685 000 au lieu de 635 000 h.
Ces changements ne modifient alors en rien les conclusions du Groupe de travail concernant l'état d'exploitation de ces stocks.
REFERENCES
FAO, 1981 Comité des pêches pour l'Atlantique Centre-Est, Bulletin statistique N° 3, COPACE. Captures nominales 1969–79. Bull.Stat.COPACE, (3):211 p.
Lim Joo Yeoul, 1982 Review on cephalopod catches by Korean trawlers in the northern zone of CECAF, 1977–1980. Annexe 15 au Rapport de la sixième session du Groupe de travail du COPACE sur l'évaluation des ressources, Dakar, Sénégal, février 1982
Shimura, T., 1981 Etude statistique sur les chalutiers internationaux basés à Las Palmas. Dakar, Sénégal, FAO, Comité des pêches pour l'Atlantique Centre-Est, COPACE/TECH/80/26:41 p.
Yamamoto, T. et M. Ansa-Emmim, 1979 CECAF Project Statistical Survey of Las Palmas based international trawlers, February 1978-February 1979. Dakar, Sénégal, FAO/UNDP Project, Development of Fisheries in the Eastern Central Atlantic, CECAF/TECH/79/15:15 p.
| OCTOPUS | Zone | Division COPACE | 1977 | 1978 | 1979 | 19801 | 19802 |
|---|---|---|---|---|---|---|---|
| 22–26°N | Dakhla | 34.1.3 | 4 234 | 5 646 | 3 954 | 724 | 1 419 |
| 19–22°N | Cap Blanc | 34.1.3 | 4 529 | 3 199 | 2 948 | 11 806 | 11 154 |
| 15–19°N | Nouakchott | 34.3.1 | 1 268 | 2 846 | |||
| }983 | }223 | }182 | |||||
| 13–15°N | Cap-Vert | 34.3.1 | 87 | 404 | |||
| Total | 10 118 | 12 093 | 7 885 | 12 755 | 12 755 | ||
| SEICHE | |||||||
| 22–26°N | Dakhla | 34.1.3 | 5 412 | 14 371 | 12 460 | 4 337 | 3 890 |
| 19–22°N | Cap Blanc | 34.1.3 | 1 409 | 2 589 | 5 082 | 9 542 | 10 835 |
| 15–19°N | Nouakchott | 34.3.1 | 1 309 | 4 498 | 2 972 | 1 763 | |
| }1 020 | |||||||
| 13–15°N | Cap-Vert | 34.3.1 | 208 | 483 | 0 | 79 | |
| Total | 8 339 | 21 940 | 20 493 | 15 745 | 15 746 | ||
| ENCORNET | |||||||
| 22–26°N | Dakhla | 34.1.3 | 6 089 | 6 108 | 3 683 | 654 | 517 |
| 19–22°N | Cap Blanc | 34.1.3 | 1 361 | 897 | 1 106 | 1 946 | 2 313 |
| 15–19°N | Nouakchott | 34.3.1 | 2 271 | 800 | 939 | 305 | |
| }74 | |||||||
| 13–15°N | Cap-Vert | 34.3.1 | 59 | 275 | 0 | 0 | |
| Total | 9 790 | 8 080 | 5 728 | 2 905 | 2 905 |
| Répartition de l'effort | |||||
|---|---|---|---|---|---|
| Zone | 1977 % | 1978 % | 1979 % | 1980 % | |
| 22–26°N | Dakhla | 62,9 | 60,1 | 62,8 | 27,0 |
| 19–22°N | Cap Blanc | 16,5 | 14,9 | 21,0 | 63,1 |
| 15–19°N | Nouakchott | 16,7 | 16,7 | 16,2 | 8,2 |
| 13–15°N | Cap-Vert | 3,9 | 8,3 | 0 | 1,7 |
| Total | 100,0 | 100,0 | 100,0 | 100,0 | |
| Effort en nombre de jours de pêche (= 12 heures d'après Joo Yeoul Lim) | |||||
| 22–26°N | Dakhla | 15 335 | 21 724 | 13 936 | 4 660 |
| 19–22°N | Cap Blanc | 4 035 | 3 154 | 4 660 | 12 276 |
| 15–19°N | Nouakchott | 4 081 | 3 536 | 3 595 | 930 |
| 13–15°N | Cap-Vert | 945 | 1 757 | 0 | - |
| Total | 24 3961 | 21 1712 | 22 1912 | 17 8662 | |
1 Estimation de Lim (1982), éventuellement sous-estimée. Ses estimations de l'effort(transformées en jours de pêche) sont pour 1978, 1979 et 1980 respectivement:20 963, 20 105 et 7 264 ou 1%, 9% et 60% inférieurs aux résultats de l'échantillonnageà Las Palmas
2 Totaux basés sur l'échantillonnage à Las Palmas
| Année | Effort
(103 h) | Cpue | Captures totales (103 t) |
|---|---|---|---|
| 1966 | 17 | 691 | 11,6 |
| 1967 | 46 | 645 | 28,7 |
| 1968 | 71 | 646 | 45,7 |
| 1969 | 60 | 573 | 34,2 |
| 1970 | 89 | 389 | 34,6 |
| 1971 | 120 | 318 | 38,2 |
| 1972 | 114 | 320 | 36,6 |
| 1973 | 100 | 292 | 29,2 |
| 1974 | 159 | 230 | 36,6 |
| 1975 | 214 | 186 | 39,9 |
| 1976 | 268 | 197 | 52,9 |
| 1977 | 125 | 292 | 36,6 |
| 1978 | 82 | 285 | 23,3 |
| 1979 | 78 | 171 | 13,4 |
| 1980 | 51 | 245 | 12,5 |
| Année | Effort (103 h) | Cpue | Captures totales (103 t) |
|---|---|---|---|
| 1966 | 55,6 | ||
| 1967 | 190 | 347 | 65,9 |
| 1968 | 168 | 416 | 69,8 |
| 1969 | 162 | 313 | 50,6 |
| 1970 | 198 | 185 | 36,7 |
| 1971 | 400 | 170 | 68,0 |
| 1972 | 665 | 126 | 83,8 |
| 1973 | 401 | 103 | 41,3 |
| 1974 | 952 | 63 | 60,0 |
| 1975 | 1 138 | 50 | 56,9 |
| 1976 | 1 239 | 59 | 73,1 |
| 1977 | 1 503 | 39 | 58,6 |
| 1978 | 1 442 | 43 | 62,0 |
| 1979 | 1 785 | 27 | 48,2 |
| 1980 | 1 491 | 34 | 50,7 |
| Zone | 1977 | 1978 | 1979 | 19801 | 19802 | ||
|---|---|---|---|---|---|---|---|
| OCTOPUS | 22–26°N | Dakhla | 276 | 444 | 284 | 155 | 305 |
| 19–22°N | Cap Blanc | 1 122 | 1 014 | 633 | 962 | 909 | |
| 15–19°N | Nouakchott | 311 | 805 | ||||
| }240 | }240 | }196 | |||||
| 13–15°N | Cap-Vert | 92 | 230 | ||||
| Total | 415 | 571 | 355 | 714 | 714 | ||
| SEICHE | 22–26°N | Dakhla | 353 | 1 129 | 894 | 939 | 835 |
| 19–22°N | Cap Blanc | 349 | 821 | 1 091 | 777 | 883 | |
| 13–19°N | Nouakchott | 321 | 1 272 | ||||
| }827 | }1 896 | }1 098 | |||||
| 13–15°N | Cap-Vert | 220 | 275 | ||||
| Total | 342 | 1 036 | 923 | 881 | 881 | ||
| ENCORNET | 22–26°N | Dakhla | 397 | 480 | 264 | 140 | 111 |
| 19–22°N | Cap Blanc | 337 | 284 | 237 | 159 | 188 | |
| 13–19°N | Nouakchott | 556 | 226 | ||||
| }261 | }328 | }80 | |||||
| 13–15°N | Cap-Vert | 62 | 157 | ||||
| Total | 401 | 382 | 258 | 163 | 163 |
1 Cpue par zone calculée d'après Lim et basée sur le total de Shimura (1981)
2 Estimations de Shimura (1981)
| Cpue Japon-Hatanaka (1979) Données Groupe de travail N° 2 (movennes mobiles) | Cpue Corée Joo Yeoul Lim (1982) et Cap Blanc | Rapport Japon/Corée | |
|---|---|---|---|
| 1977 | 292 | 1 122 | 0,26 |
| 1978 | 285 | 1 014 | 0,28 |
| 1979 | 171E | 633 | 0,27E |
| 1980 | 245E | 909 | 0,27E |
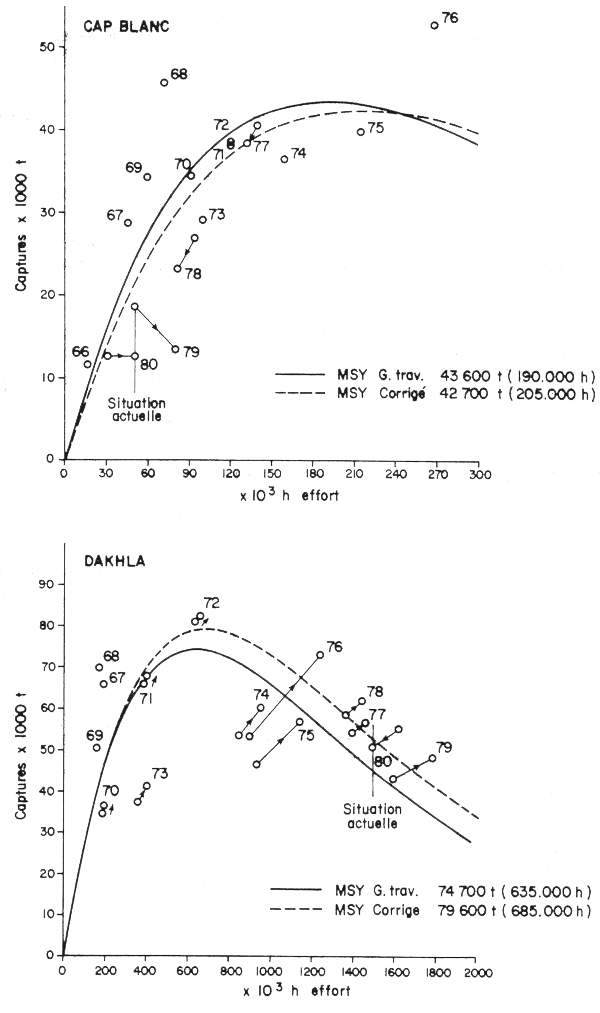
Figs. 1 et 2 - Corrections des modèles de Fox pour les poulpes du Cap Blanc et de Dakhla; les flèches indiquent les valeurs corrigées

![]()
![]()