![]()
![]()
![]()
LISTE DES EXPERTS
| V. ANIKIN | Représentation – URSS |
| M. ANSA–EMMIM | Secrétaire du COPACE – BP 154, |
| Dakar, SENEGAL | |
| I. BA | CNROP, BP 22, Nouadhibou, MAURITANIE |
| M. O/CHEIKH | CNROP, BP 22, Nouadhibou, MAURITANIE |
| L. DANKE | Institut für Hochseefischerei und |
| Fischverarbeitung | |
| 2500 Rostock – Marienehe | |
| DDR | |
| F. DOMAIN | CNROP, BP 22, Nouadhibou, MAURITANIE |
| M. DUPRET MELO | Direction Générale des Pêches, |
| CAP VERT | |
| E. JOSSE | CNROP, BP 22, Nouadhibou, MAURITANIE |
| J. J. LEVENEZ | CRODT, BP 2241, Dakar, SENEGAL |
| J. LOPEZ | CRODT, B.P. 2241, Dakar, SENEGAL |
| C. MAXIM | * IRCM, ROUMAINE |
| S. OVERKO | AtlantNIRO, URSS |
| F.C.ROEST | Project COPACE, BP 154, Dakar, SENEGAL |
| B.SAMB | CRODT, BP 22, Dakar, SENEGAL |
| O. SARAFOLEANU | IPO - ROUMAINE |
| A. SOW | CNROP, BP 22, Nouadhibou, MAURITANIE |
| I. STAICU | * IRCM, ROUMANIE |
| H. SANTA RITA VIEIRA | Direction Généerale des Péches, |
| CAP VERT | |
| R. WEISS | Institut für Hochseefischerei und |
| Fischverarbeitung | |
| 2 500 Rostock – Marienehe | |
| DDR |
* Institut Roumain de Recherches Marines, Constanza, ROUMAINE
ORDRE DU JOUR ANNOTE
1. OUVERTURE
2. ADOPTION DE L'ORDRE DU JOUR
3. INVENTAIRE DES DONNEES BIOLOGIQUES 1
3.1 Bibliographie
Afin de créer une base de référence, chaque participant est prié d'apporter à réunion une liste de référence des articles pertinents sur les chinchards et les maquereaux et, si possible, des photocopies des documents les plus importants.
3.2 Données biologiques
Chaque participant compilera, en tenant compte de toutes les sources disponibles, les paramétres basaux des populations. Ceci permettra un choix rapide des paramétres à utiliser lors de la réunion pour l'évaluation des stocks. Si la distribution géographique des espéces de chinchards et maquereaux n'est pas limitée à la zone nord du COPACE, des paramétres pour les autres régions pourraient également #tre inclus ; de même, on pourrait inclure des données pour des espéces voisines à l'intérieur ou en dehors de la zone nor du COPACE. L'information requise concerne :
le rapport longueur–poids ;
la taille et/ou l'âge de (50%) maturité sexuelle et les périodes et zone de reproduction ;
les migrations, la répartition géographique et la séparation des stocks ;
la composition en longueur ;
les paranétres de croissance (K, Loo, to) ;
les taux de mortalité (F, M et Z) ;
les changements à long terme et les conditions climatologiques (en Afrique Occidentale et ailleurs) ;
parasitisme chez ces espéces.
3.3 Données sur les péches
Les péches doivent #tre décrites (pays, navires, engins, espéces cibles, captures annexes) y compris d'éventuels changements dans le temps (espèces cibles, péches, etc..).
Des tableaux sur les captures, les captures par unité d'effort et l'effort de péche seront également présentés. Information requise :
zones de péche, saisons de péche ;
nombre de navires et caractéristiques ;
engins de péche et maillages utilisés ;
captures totales par espéces : cibles et autres espéces (à spécifier) ;
tendances en effort, différentes mesures d'effort ;
captures par unité d'effort, indices d'abondance.
4. ETUDES DE LA SELECTIVITE
5. EVALUATION/ETAT DES STOCKS
5.1 Modéles de production : pertinence, résultats
5.2 Modéles analystiques
5.3 Autres études, prospections acoustiques, chalutages
6. CONCLUSIONS ET RECOMMANDATIONS
6.1 Recherche
6.2 Aménagement
7. ADOPTION DU RAPPORT
BIBLIOGRAPHIE
Cette bibliographie comportant 182 titres est basée sur les références fournies par les experts de la Mauritanie, de la RDA, de la Roumanie, du Sénégal, de l'URSS ainsi que du Project COPACE.
Elle n'est probablement ni exhaustive ni tout à fait sans erreurs, la plupart des références n'ayant pu #tre vérifiée à Dakar.
Aboussouan, A., 1967 Oeufs et larves de téléostéens de l'ouest africain, V. Caranx rhonchus Geoffr. St.Hil. (Carangidae). Affinités avec T. trecae Cadenat. Bull.IFAN (A), 29 (3) : 1039–1050.
Aleev, J.G., 1956 O Sistematiceskoj polozenii stavridy Cornogo Moria. Voprosy Ichiologii 7 : 174–184.
Aleev, J.G., 1957 Stavridy morej SSSR trudy Sevastopolskoj Biostanoji, AN SSSR 9 : 167–242.
Alekseev, F. E, 1969 Quelques données sur la biologie du maquereau des côtes nord–ouest africaines, Tr. AtlantNIRO 22 : 308–316 (en russe).
Alekseeva, E.I., 1973 Oogenése chez le maquereau Scomber colias Gmelin des côtes nord–ouest africaines. Tr. AtlantNIRO : 86–88 (en russse)
Alferova, N.M. and G. N. Nefedov, 1973 Electrophoretic investigation of esterases of some species in the Eastern Atlantic. Bioch.genetics of fishes 50 : 192–200 (en russe).
Aloncle, H., 1960 Remarques numériques préliminaires sur les branchicténies de Trachurus trachurus (Linné 1758). Bull.Inst.Péches.Maroc 4 : 3–8
Aloncle, H., 1964 Note sur la croissance et quelques caractéres numériques de Trachurus trachurus (Linne 1758) des côtes atlantiques du Maroc. Bull.Inst.Pêches Marit.Maroc 11 : 25–38.
Barkova, N. A., 1973 The peculiarities in distribution and biology of horse mackerel (Tr. trecae Cad.) from the Northwest African coast.
Barkova, N., P. Bukatin, J. Bourdin, M. Domanevskaya, L. Domanevsky, S. Krivospitchenko, A. Kozhemiakine, N. Mylnikov, S. Overko, Yu. Sazanov et K. Sedykh., 1981 Répartition et cycles vitaux de quelques espéces de poissons dans les secteurs côtiers (34.1.3 et 34.3.1) de l'Atlantique Centre–Est, pp 31–69 in: Contributions soviétiques à l'évaluation des ressources halieutiques dans la région du COPACE. COPACE/TECH/81/31 (Fr.)
Barraca, I.F., 1964 Quelques aspects de la biologie et de la pêche du chinchard Trachurus trachurus (L) de la côte portugaise. Notas e Estudos do Instituto do Biologia Maritima, 29.
Bellemans, M., 1982 Common names of West African marine animals/Noms usuels des animaux marins de l'Ouest africain. CECAF/TECH/82/44 : pag.var.
Benko, Y. K. et E. Pozo, 1975 Algunas caracteristicas de la biologia de jurel Trachurus trachurus (L., 1758) que habita junto a las costas de Africa occidental. ICSEAF Coll.Sc.Pap.2 : 27–33.
Berenbeim, D.Y., 1974 On regularities in the variability of the spawning terms of horse mackerel (Trachurus trachurus Linné) within the spawning area. ICES C.M.1974/J : 14 : 9p (mimeo).
Berenbeim, D.Y., S.M. Overko and V.A. Sedletskaya, 1973 On regularities in the variability of the optimum surface temperature in the course of development of the eggs and larvae of horse mackerel, Trachurus trachurus Linné. ICES C.M. 1973/J : 8 : 4p (mimeo).
Birjukov, N.R., 1960 Résultats de la troisiéme expédition de recherche et de pêche dans les eaux équatoriales des côtes ouest-africaines. Tr.BaltNIRO 5 : 60–90 (en russe).
Blanc, M., et M.L.Bauchot, 1961 Sur quatre genres de Carangidae (téléostéens perciformes de la côte occidentale d'Afrique : Decapterus, Caranx, Trachurus, Saureus). Affinités et rapports phylogéniques. Bul.Mus.Natl.Hist.Nat. (2° Ser.), Paris, 32(6) : 484–497.
Boely, T., 1979 Les poissons pélagiques au Sénégal. La pêche de Caranx rhonchus. Bull.Inst.Fondam.Afr.Noire (A) 4 : 859–886.
Boely, T., 1974 et al., Composition par espéce et par taille des captures du navire usine ASTRA. Mars/avril 1973–juin 1973–février-mars 1974.Doc.Sci.Cent.Rech.Océanogr.Dakar-Thiaroye ORSTOM (58) : 6p. (also issued as ICES C.M. 1974/J : 16 : 6p. (mimeo).
Boely, T., J. Chabanne, et P. Fréon, 1979 Schéma des cycles migratoires, lieux de concentrations et périodes de reproduction des principales espéces de poissons pélagiques côtiers dans la zone sénégalo-mauritanienne. COPACE/PACE/Series/78/10 : 63–70.
Boely, T. et P. Fréon, 1979 Les ressources pélagiques in Les ressources halieutiques de l'Atlantique Centre-Est. FAO.Doc.Tech.Pêches (186.1) : 13–78.
Boely, T., et O. J. φstvedt, 1976 Les poissons pélagiques côtiers au Sénégal. Observations faites à bird du navire-usine ASTRA de la Mauritanie aux Iles Bissagos. Bull.Inst.Fondam.Afr.Noire (A), 38 (3) : 677–702.
Boely, T., A. Wysokinski et J. Elwertowski, 1973 Les chinchards des côtes sénégalaises et mauritaniennes. Biologie-déplacement-ressources. Doc.Sci.Provis.Cent.Rech.Océanogr.Dakar-Thiaroye, ORSTOM, (46) : 47p.
Bonnet, M., 1970 Données biométriques sur les Scomber colias des côtes nordouest africaines.Rapp.P.-V. Réun. CIEM, 159 : 189–190.
Borodatov, V.A. et J.L. Karpechenko, 1958 Perspectives de dévelopment de la pêche dans la partie orientale du moyen atlantique. Rybn.Khoz., Mosk., 34 (6) : 10–19 (en russe).
Bukatin, P.A., A.I., Kozhemiakin, S.G. Krivospitchenko, 1981 Marquage de poissons et de céphalopodes dans l'Atlantique Centre-Est, Rybnoe khoziaistvo 1 : 27–28 (en russe).
Cadenat, J. 1949 Description de quatre téléostéens nouveaux de la côte occidentale d'Afrique, Cepola pauciradiata, Scorpenodes elongatus, Trachurus trecae, Chromis lineatus. Bull.Mus.Natl.Hist.Nat. (2° Ser.), Paris, 21 (6) : 663–671.
Carliste, J.G., 1971 Food of the Jack mackerel, Trachurus symmetricus, Calif. Fish Game 57(3) : 205–208.
Chabanne, J. et J. Elwertowski, 1973 Cartes des rendements de la pêche des poissons pélagiques sur le plateau continental nord-ouest africain de 11° à 26°N. Doc.Sci.Provis.Cent.Rech.Océanogr.Dakar-Thiaroye, ORSTOM (49).
Chlapowsky, K., B. Draganik, and M. Wyszynski, 1982 The Polish horse mackerel fishery in the Cunene and Cape Cross Divisions and its possible impact on hake. ICSEAF Coll.Sc.Pap. 9 (2) : 65–71.
Conand, F., et C. Franqueville, 1973 Identification et distribution saisonniére des larves des principales espéces de Carangidae des côtes du Sénégal et de la Gambie. Doc.Sci.Provis.Cent.Rech.Océanogr.Dakar– Thiaroye ORSTOM (47) : 36p. (also issued as Bull.IFAN(A) 35 (4) : 951–978 (1973).
Czajka, W., and W. burawa, 1970 Trawl nets used by Polish trawlers on fishing grounds of the West African shelf. Rapp. P.–V. Réun. CIEM, 159 : 264–271.
da Franca, P., et M. L. Paes da Franca, 1958 Contribuçao para o conhecimento da biologia dos Trachurus de Angola. Trab.Cent.Biol.Pisc., Lisb., (21–25) : 53–135
da Silva Marécos Godinho, M. L., 1974 Preliminary report on age and growth of the Cunene horse mackerel (Trachurus trachurus trecae, Cadenat.) in Angola. Collect.SciPap.ICSEAF, 1 : 117–123 (also issued as : Pub.Mimeogr.Missao Estud.Bioceanol.Pescas Angola, (12).
do Campos Rosado, J. M., 1972 Estudos sobre a pescaria de carapau com artes de cerco em Angola. 1. Estimativa do componente wt do modelo de Beverton–Holt em Trachurus trachurus. Publ.Mimeogr.Missao Estud. Bioceanol. Pescas Angola, (5) : 33p.
do Campos Rosado, J. M., 1972 Estudos sobre a pescaria de carapau com artes de cerco em Angola. 2. Variaçoes da abundancia de Trachurus trecae Beverton–Holt em Trachurus trachurus. Publ.Mimeogr.Missao. Estud.Bioceanol.Pescas Angola, (8) : 37 p.
Domanevsky, L. N., 1970 Biology and distribution of the main commercial fishes and peculiarities of their fishing by trawl on the shelf from Cape Spartel to Cape Verde.Rapp.P.–V. Réun.CIEM, 159 : 223–226.
Domanevsky, L. N., 1970 Peculiarities of the fish biology and fish resources of the East Central Atlantic pp. 61–75 in The present state of biological productivity and biological resources of the World Ocean and the prospects of their utilization. Kaliningrad. (en russe).
Domanevsky, L. N., 1979 The peculiarities of spawning of the fish in the East Central Atlantic, pp. 84–86 in Rep. to the 5th All–Union Conference “Problems of fishery oceanography of the WorldOcean, AtlantNIRO, Kaliningrad. (en russe).
Domanevsky, L. N., 1980 Ichthyogeographic complexes and their structure off the Atlantic Coast of Africa, pp. 35–36 in “Problems of Marine Biogeography” .Acad. Sci.USSR. DHTZ, IBM, Vladivostok. (en russe).
Domanevsky, L. N. 1980 A combined method for preliminary assessment of the fish biomass, pp. 49–50 in “Quantitative methods in the ecology of animals”. Zool.Inst.Acad.Sci. USSR. (en russe).
Domanevsky, L. N. 1982 Life cycles and distribution of the fish in the East Central Atlantic, pp. 1–2 in Rep. of the Sc.Tech.Meeting “Application of physical irritants aimed at the development of the marine fishery”. (en russe).
Domanevsky, L.N. and S.G. Krivospitchenko, 1979 Some peculiarities of the abundance fluctuations of mackerel in the East Central Atlantic, Sahara Division (34.1.3), pp. 46–47 in Reports to the All–Union Conf. Kaliningrad. (en russe).
Domanevsky, L.N., N.I. Mylnikov et M.V. Stiopkina, 1978 Caractéristiques de la nutrition de quelques espèces de poisson des côtes nord–ouest africaines. Tr. AtlantNIRO 74 : 44–48 (en russe).
Draganik, B., 1977 Horse mackerel (Trachurus trachurus, L.) fished in the Southeast Atlantic, Coll.Scient.pap.Int.Comm.Se.Atl.Fish.4 : 25–65.
Elwertowski, J., 1970 Notes sur la pêche des poissons pélagiques par les chalutiers congélateurs polonais sur les fonds mauritanosénégalais en février/mars/avril 1970. Rapp.Sc.Proj.Dev.Pêche pélag.Côt-FAO/PNUD Abidjan (2/70) : 56p.
Elwertowski, J., 1972 Statistiques de pêche des chalutiers congélateurs polonais dans les eaux internationales du Sénégal, de la Mauritanie et du Rio de Oro.Off.Central Pêches, Pologne, 2p., 44 tableaux.
Elwertowski, J., et T. Boely, 1971 Répartition saisonnière des poissons pélagiques côtiers dans les eaux mauritaniennes et sénégalaises. Doc.Sci.Cent.Rech.Océanogr.Dakar-Thiaroye ORSTOM (32) : 15p.
Elwertowski, J., P. Gonzalez Alberdi, J. Chabane et T. Boely, 1972 Première estimation des ressources pélagiques du plateau continental nord ouest africain (zone de transition nord de l'Atlantique Centre-Est). Doc.Sci.Provi.Cent.Rech.Océanogr.Dakar-Thiaroye ORSTOM, (42) : 33p.
Falk, U., 1965 Résultats biologiques du voyage en Afrique du nord-ouest du bateau de capture et de traitement de poissons “Walter Dehmel” en novembre-décembre 1964. Fisherei-Forsch.,Rostock, 4(1) : 59–67 (en allemand).
Falk, U., 1967 Sediments as food of chub mackerel (Scomber colias Gmelin) off northwest Africa, CIEM C.M. Comm.Poiss.Pélag.5.
Falk, U., 1967 Chub mackerel (Scomber colias) off the coast of Northwest Africa CIEM C.M. Comm.Poiss.Pélag. 5.
FAO, 1965 Fisheries Division, Fisheries Statistics Section, Economics Branch The Atlantic fishery for tuna-like and mackerel-like scombriforms by fishing craft from Japan, FAO, Fish.Cir. (48) : 7p.
FAO, 1965 Fisheries Division, Fisheries Statistics Section, Economics Branch, Tuna-like and mackerel-like scombriforms (sampled collection only) : Japanese catches in the Atlantic. FAO.Fish.Circ., (47) : 15p.
FAO, 1965 Fisheries Division, Fisheries Statistics Section, Economics Branch, Tuna-like and mackerel-like scombriforms : catches in the Atlantic. FAO.Fish.Circ. (45) : 29p.
Fréon, P., B. Stequert, et T. Boely, 1978 La Pêche des poissons pélagiques côtiers en Afrique de l'ouest des Iles Bisagos au nord de la Mauritanie : description des types d'exploitation. Cah. ORSTOM (Océanogr.) 16 (3–4).
Fréon, P., B. Stequert, et T. Boely, 1978 La pêche des poissons pélagiques côtiers des Iles Bissagos au nord de la Mauritanie : description et interaction des pêcheries. Document présenté au ICES. Symposium sur le courant des Canaries : upwelling et ressources vivantes. Las Palmas, Gran Canaria, 1978.Doc.(93) 11–14 avril.
Fréon, P., B. Stequert, et T. Boely, 1979 Les pêches sénégalaises : description et analyse des captures et des rendements des principales espèces pélagiques côtières. In Rapp. du Groupe de travail ad hoc sur les poissons pélagiques côtiers ouest africains de la Mauritanie au Libéria (26° à 5°N), Dakar, Sénégal, 19-24 juin 1978. COPACE/ PACE Sér. 78/10 : 27–62.
Gail, R., 1952 Trachurus. Etat sexuel et croissance du saurel des côtes atlantiques du Maroc (Trachurus trachurus L.) Ann.Biol.CIEM 8 : 90–91.
Gail, 1955 R. Nouvelles observations sur Trachurus picturatus Bowdich.Rapp. P.-V. Réun. CIEM (137) : 57–58.
Garcia, R.C., 1966 Investigaciones del Instituto Espanol de Oceanografia sobre los escombridos de las aguas ecuatoriales africanas. Publ. Tec. Junta Estud.Pesca, (5) : 85–139.
Gaudio, A. 1976 La pêche dans l'Atlantique Saharien, pactole inestimable. Pêche Marit. 1181 : 471–473.
Gréboval, D., 1982 Analyse bioéconomique des principales pêcheries démersales dans la région nord COPACE. COPACE/TECH/82/45 : 118p.
Habashi, H., and J. Wojciechowski, 1973 Observations on the biology of Scomber japonicus off northwest Africa, ICES C.M. 1973/J : 20 : 10p. (mimeo).
Hagen, E., and R. Weiss, 1979 Mesoscale coastal upwelling dynamics off the NW African coast and hypothetic relations to the chub mackerel concentrations. ICES. C.M. 1979/C Hydrographic Committee.
Hagen, E., and R. Weiss, 1980 On the dependency of the variability of chub mackerel concentrations on the dynamics of the mesoscale coastal upwelling off the NW African coast. Fischerei-Forsch. Rostock, 18 : 19–31. (en allemand).
Hartsuijker, L., 1972 Preliminary results of the sampling of the purse seiners landing at the fishing harbour of Abidjan. Supplement 2 : The mackerel Scomber japonicus. Rapp.Sci.proj.Dev.Pêche Pélag.Côt. FAO/PNUD Abidjan, (10/72) : 1–35.
Hartsuijker, L., 1972 L'échantillonnage des débarquements des sardiniers au port de pêche d'Abidjan. 2. Analyse de la couverture du débarquement par l'échantillonnage en 1970 et 1971 pour les espèces : Sardinella aurita, S. eba, Scomber japonicus. Rapp. Sci.Proj.Dev.Pêche Pélag.Côt. FAO/PNUD, Abidjan, (13/72) : 1–45.
Holzlöhner, S. and G. Kloxin, 1982 On the species composition and the length– age structure of the important pelagic fishes off North West Africa, 1981, ICES. C.M. 1982 H:55 Pelagic Fish Committee.
Holzlöhner, S., and R. Weiss, 1978 Fishery research by the GRD off NW Africa Seewirtschaft 10 : 508–509 (en allemand).
Kalinina, E.M., and V.A. Sedletskaya, 1979 Quantitative composition and distribution of ichthyoplankton along the Mauritanian coast. Voprosy ikhtiologii 19, 4 (117) : 737–742. (en russe).
Karnicki, Z., 1970 The characteristics of the fishing ground at cap Timiris on the coast of West Africa. Rapp. P.–V. Réun. CIEM 159 : 218–222
Kiliachenkova, V.A., 1970 Development and distribution of eggs and larvae of Trachurus trachurus L. Rapp.P.–V. Réun. CIEM : 194–198.
Klimaj, A., 1970 Fishing yield and species composition of trawl catches of M.T. ‘RAMADA’ on the fishing grounds on the shelf off Mauritania and Senegal in 1967. Rapp. P.–V. Réun.CIEM, 159 : 254–260.
Klimaj, A., 1976 Fishery atlas of the NW African shelf. Center for Scientific, Technical and Economic Information, Warsaw, Poland, 221p.
Kolesnikov, V.G., 1971 Habitat and distribution of horse mackerels in the area off NW Africa depending on the oceanographic environment.
Kolesnikov, V.G. and K.Y. Mratov, 1970 Vertical circulation and migrations of fish in the area off West Africa. In The Atlantic ocean, fish scouting research. Publ.AtlantNIRO Sér.3 (en russe).
Kolesnikov, V.G. and S.M. Overko, 1970 Features of dynamics processes in the Central eastern Atlantic Ocean and their effects on horse mackerel distribution. ICES. C.M. 1970/J : 6.
Kolesnikov, V.G. and S.M. Overko, 1971 Habitat and distribution of horse mackerel in the area off NW Africa depending on the oceanological environment ICES C.M. 15 : 11.
Komarov, J.A., 1972 Biological peculiarities of South African horse mackerel (T. trachurus). ICES C.M. Scomber 114 : 1–2.
Komarov, J.A., 1964 On the reproduction of the horse mackerel (Trachurus trachurus) off the South–Western coast of Africa, ICES C.M. Atl. Committee (94) : 5p.
Komarov, J.A., 1971 On the differentiation of unit stocks of South African horse mackerel (Trachurus trachurus capensis Castelnau). Tr.AtlantNIRO 41 : 33–42 (en russe).
Kompowski, A., 1975 Food and feeding of some Carangidae in the Cape Blanc area, ICES C.M. 1975/k : 42 (miméo).
Kompowski, A., 1975 The intraspecific geographical variability of the horse mackerel Trachurus trachurus (L.) in the West African shelf waters. Acta Ichtyol.Pisc. 5 (1) : 13–29.
Kompowski, A., 1976 A study on the food and feeding habits of Trachurus trachurus, Tr. trecae, Tr. picturatus and Caranx rhonchus in the region of Cape Blanc.Acta Ichthyol.Pisc. 6 (1) : 35–57.
Krivospitchenko, S.G., 1981 Peculiarities of applying the Fox model to the description of mackerel fishing in the Central Eastern Atlantic. pp. 34–39 in “State of stocks and principles of rational fishery in the Atlantic Ocean. AtlantNIRO, Kaliningrad (en russe).
Kuderskij, S.K., 1962 Influence of hydrological conditions on the distribution and behaviour on the South African horse mackerel (T. trachurus). ICES C.M. 1962 124 : 1.
Kwei, E.A., (n.d.), The biology, distribution and effect of fishing on the migration of Scomber japonicus (Spanish mackerel). Paper presented to the Sixth biennial Conference of the Ghana Science Association.
Letaconnoux, R., et A.E.J. Went, 1970 Symposium sur les ressources vivantes du plateau continental Atlantique Africain du détroit de Gibraltar au Cap Vert. Rapp.P.-V. Réun. CIEM 159 : 289p.
Lozano Cabo, F., 1958 Los escombridos de las aguas españolas y marroquies y su pesca. Primera parte. Trab.Inst.Esp.Oceanogr., (25) : 254p.
Marchal, E., 1961 Quelques observations complémentaires sur les Scombridae pêchés par le PYRRHUS. Bull.Inst.Fr.Afr.noire (A), 23(1) : 102–107.
Marchal, E., 1963 Description des states post-larvaires et juvéniles de quatre espèces de Scombridae de l'Atlantique tropico-oriental. Mém.Inst. Fondam.Afr.Noire. 68 : 201–40
Kompowski, A., and W. Slosarczyk, 1975 Biological characteristics of Polish catches of the horse mackerel Trachurus trachurus capensis Castelnau 1961 in the region of the South West African shelf. Acta Ichthyol.Pisc. 5(2) : 25–38.
Kompowski, A., and W. Slosarczyk, 1976 Some investigations on horse mackerel (Trachurus trachurus, L.) in the ICSEAF area, in 1973, ICSEAF Coll.Sc.Pap. 3 : 27–31.
Kompowski, A., and A. Wysokinski, 1975 Formation of growth zones in otoliths of scad (Trachurus trachurus) from the Cape Blanc Region. Prace Morskiego Inst. Ryback 18 (A) :
Krivospitchenko, S.G., 1975 Changes in the length composition in the catches of Atlantic mackerel (Scomber colias) in the Central East Atlantic from 1968 to 1975. Annales biologiques 32 : 176–177.
Krivospitchenko, S.G., 1978 Size structure of Atlantic mackerel (Scomber colias) in the Sahara sub-area of the Central east Atlantic, 1975–1978. Annales biologiques 35 : 215.
Krivospitchenko, S.G., 1979 Le maquereau Scomber japonicus devant le littoral saharien. COPACE/PACE Series/78/10 : 130–133.
Krivospitchenko, S.G., 1979 Some biological features of mackerel, Scomber japonicus Gmelin 1789, of the Azore and Canary aggregate Banks, pp. 62–63 in “State of Stocks and dynamics of pelagic fish abundance in the World ocean”, Rep.of All-Union Conference, Kaliningrad. (en russe).
Marchal, E., et T. Boely, 1977 Evaluation acoustique des ressources en poissons du plateau continental ouest-africain des Iles Bisagos (11°N) à la Pointe Stafford (28°N). Cah.ORSTOM (Sér.Océanogr.), 15(2) : 139–162.
Maxim, C., 1980 Assessment of the natural mortality of the horse mackerel (Trachurus trachurus trachurus L.) in the Atlantic Ocean off Southwest Africa Halieutica 4 : 79–89 (en roumain).
Maxim, C., 1982 Assessment of stock and calculation of recruitment and total admissible catch on the horse mackerel (Trachurus trachurus trachurus) in the East-Central Atlantic Ocean-Mauritanian waters (en roumain). Halieutica 2 : 75–88.
Maxim, C., et I. Staicu, 1976 Observations sur la biologie et dynamique du stock de Saurel (Caranx rhonchus Geof.St.Hist.) dans l'Atlantique Est-Central. Cercetari marine, I.R.C.M. 9 : 215–226.
Maxim, C. et I. Staicu, 1980 Assessment of the Atlantic horse mackerel stock in the Eastern-Central Atlantic ocean (Division 34.1.3 and 34.3.1) using provisional 1980 data, Cercetari marine, I.R.C.M., 13 : 163– 172.
Maxim, C. and I. Staicu, 1980 Evaluation of the horse mackerel stock in the Northwest African area of Atlantic ocean, (en roumain). Halieutica 2 : 103–116.
Morales, B., 1982 Considerations on scorpionfish, horse mackerel, and kinglip otoliths in the Southeast Atlantic and proposed guidelines for interpretation.Coll.Scient.pap.int.Comm SE.Atl.Fish, 9(2) : 199–219.
Mylnikov, N.I., 1976 Age and growth of horse mackerel, Trachurus trachurus trachurus L., of the Northwest African waters, p. 10 in “Problems of investigation of pelagic fishes and invertebrates of the Atlantic Ocean”. Reports of the Conference of Young Scientists AtlantNIRO, Kaliningrad (en russe).
Mylnikov, N.I., 1979 Some data on age and growth of bigeye scad. p.66 in “State of stocks and dynamics of pelagic fish abundance in the World Ocean”. Report of the All-Union Conference, Kaliningrad. (en russe).
Nefedov, G.N., N.M. Alferova and Y.A. Chuskin, 1978 Polymorphism of muscle esterases in horse mackerel of the Northeast Atlantic, In Biology of the sea 2 : 64–74. (en russe).
Nehring, D., 1978 Research into the influence of the environmental factors in the NW African shelf area. I The meteorological and oceanographic conditions between 22 and 26 °N, on 25–31 march 1976. Fischerei Forsch, Rostock, 16 : 39–45 (en allemand).
Nekrasov, V.V. 1970 Stavridy (sem.Carangidae) vostocnogo pobereza Afriki. Tr. AzcerNIRO 29 : 89–138.
φstvedt, O.J., 1973 Report on the Norwegian pelagic fishery off West Africa during 1970 and 1971. FAO Fish.Rep., (136) : 66–69.
φstvedt, O.J., and J. Blindheim, 1973 Studies of the abundance and distribution of fish off West Africa, November–December 1972, ICES. C.M. 1973/ J 24 : 16p. (miméo).
φstvedt, O. J. et al., 1973 Report on a cruise by the RV “G.O. SARS” to West Africa, 23 October–15 December 1972, ICES, C.M. 1973/J 13 : 14p (miméo).
φstvedt,J.O. and S. Myklevoll, 1975 Report on the Norwegian pelagic fishery off West Africa (1970–1973) FAO Fish.Rep., (158) 63 : 84
Overko, S.M., 1964 Sur la biologie et la pêche de chinchard près des côtes de l'Afrique Nord–Occidentale. Tr. AtlantNIRO 11 : 45–64 (en russe).
Overko, S.M., 1969 Données sur la fécondité des Carangidés des côtes nord-ouest africaines, Tr. AtlantNIRO 22 : 72–85. (en russe).
Overko, S.M., 1969 Sur la biologie de reproduction des chinchards des genres Trachurus et Decapterus près des côtes nord-ouest africaines. Tr. AtlantNIRO 22 : 86–102. (en russe).
Overko, S.M., 1971 Size composition of Trachurus trecae stock in the shelf waters of the NW African coast. ICES. C.M. Pelagic Fish Committee J : 3.
Overko, S.M., 1971 Studies in length structure of the Caranx rhonchus stock of the West African coast (Cape Verde-Cape Blanc). ICES C.M. Pelagic Fish (Southern) Committee G : 3.
Overko, S.M., 1971 Sur la position systématique des chinchards du genre Trachurus près des côtes ouest-africaines. Tr. AtlantNIRO 41 : 82–87 (en russe).
Overko, S.M., 1971 Morphologie, biologie, et pêche du chinchard vulgaire de l'Atlantique Centre-Est.Tr. AtlantNIRO 41 : 102–121 (en russe).
Overko, S.M., 1971 Dynamics of the length structure of the commercial stocks of horse mackerel (Trachurus trachurus) in the Central Atlantic waters.ICES. C.M. Pelagic Fish Committee.
Overko, S.M., 1973 Caractéristiques morphologiques de Decapterus rhonchus de l'Atlantique Centre-Est. Tr. AtlantNIRO 53 : 34–42 (en russe).
Overko, S.M., 1974 On peculiarities in the biology of horse mackerel (Trachurus trachurus) from the Central East Atlantic. ICES. C.M. J : 13.
Overko, S.M., 1979 Morpho-biological characteristics of Caranx rhonchus in the Eastern Central Atlantic. COPACE/PACE/Series 78/10 : 127–129.
Overko, S.M., and N.A. Barkova, 1973 The peculiarities in distribution and biology of horse mackerel Trachurus trecae Cad. from the Northwest African coast. ICES. C.M. Pelagic Fish (Southern) Committee J : 7 :9p.
Overko, S.M., and V.G. Kolesnikov, 1970 Features of dynamics processes in the Central-Eastern Atlantic Ocean and their effects on horse mackerel distribution. ICES CM J : 6 Pelagic Fish (Southern) Committee.
Overko, S.M., et N.I. Mylnikov, 1979 The biology and fishery of horse mackerel (Trachurus trecae) in the Eastern Central Atlantic. CECAF/ECAF Series/78/10 : 81–85.
Overko, S.M., and V.A. Sedletskaya, 1974 Studies of the size structure dynamics of the commercial stock of mackerel (Scomber colias) in the Central East Atlantic. Annales Biologiques ICES : 1–2.
Podsevalov, V.N., and L.A. Perova, 1975 Seasonal variations of the fat content in the meat of mackerel and horse mackerel. Rybn, Khoz., Mosk., (3) : 72 (en russe).
Poinsard, F. and L. Villegas, 1975 Analyse de la pêche côtière au chalut dans l'Atlantique marocain. Trav.Doc.Dév.Pêche.Marit.Maric., (12) : 37p.
Postel, E., 1955 Contribution á l'étude de la biologie de quelques Scombridae de l'Atlantique tropical oriental. Ann.Stn.Océanogr.Salambô, 10 167p.
Radu, Gh., 1982 Report on the results of the researches carried out in the East-Central Atlantic ocean, on the continental shelf of the I.R. of Mauritania, between October 1981 and January 1982 (en roumain). Halieutica 2: 7–74.
Razniewski, J., 1957 An observation on the biology of Scomber colias of the north-west African shelf. ICES, C.M. 1967/J : 4 Pelagic fish Committee.
Riazantseva, E. I., 1976 Electrophoretic studies of myogens and esterases of Scomber scombrus L., and Scomber colias, p. 24 in “Problems of investigation of pelagic fish and invertebrates of the Atlantic Ocean. Rep.Conf.Young.Scientists.Kaliningrad. (en russe).
Ritzhaupt, H., 1955 Les zones de pêche des côtes africaines (en allemand) Fischerei Forsch., Rostock, 4 (1–2) : 26–29.
Sedlestskaya, V.A., 1970 Development and distribution of eggs and larvae of Trachurus trachurus L., Rapp.Proc.Verd.Réun. 159 : 194–198.
Sedletskaya, V.A.,1971 Spawning dynamics of horse mackerel (Trachurus trachurus L.) off the NW African coast. Tr. AtlantNIRO 41 : (en russe).
Sedletskaya, V.A., 1972 Quelques aspects de la biologie de reproduction des poissons pélagiques de l'Atlantique Centre-East, pp. 34–35 in Résumés des rapports de la Conférence de l'Institut de l'industrie alimentaire, 1–10 mars 1972. (Section Aquaculture et Technologie de la Conservation) (en russe).
Sedletskaya, V.A., 1973 Sur la répartition des oeuvs et des larves du chinchard sud-africain (Trachurus trachurus L. ) Tr. AtlantNIRO 53 : 157–163 (en russe).
Sedletskaya, V. A., 1975 Dynamics of maturing of horse mackerel (Trachurus trachurus L.) in various areas of the East Central Atlantic and distribution of spawning grounds relative to the seasonal position of the hydrological frontal zone. Voprosy ikhtiologii vol. 15, 2 (91) : 267–273 (en russe)
Sedletskaya, V.A., 1976 Biomass and abundance of the larval fish in the East Central Atlantic in February and August 1973. Tr. AtlantNIRO 69 45–50 (en russe).
Sedletskaya, V.A., 1977 Reproduction des principaux poissons pélagiques des côtes nord–ouest africaines. Sebastopol, Inst. de la biologie des mers méridionales, 24p. (en russe).
Sedletskaya, V.A., 1979 On the ecology and reproduction of the most massive pelagic fishes of the East Central Atlantic, pp. 88–89 in “State of stocks and dynamics of pelagic fish abundance in the World Ocean”. Rep. of All-Union Conf., Kaliningrad (en russe).
Sedletskaya, V.A., 1973 and D.J. Berenbeim, On regularities in the variability of the optimum surface temperature in the course of development of the eggs and larvae of the horse mackerel Trachurus trachurus Linné. ICES CM Pelagic Fish Committee N : 8.
Shaboneev, I.E., 1973 Elements of the biology of the horse mackerel (Trachurus picturatus picturatus Bowd.) of the East Central Atlantic. Tr. VNIRO 93 M: 91–101 (en russe).
Shaboneev, I.E., 1980 Systematics, morphological characteristics and origin of horse mackerels of the genus Trachurus. Voprosy ikhtiologii 20, 6, (125) : 787–799 (en russe).
Shaboneev, I.E., 1977 and E.I. Riazantseva, On the population structure of the oceanic horse mackerel, Trachurus picturatus picturatus Bowd. Voprosy ikhtiologii 17, 6 (107) : 1127–1131 (en russe).
Sosa, M., 1981 Edad y crecimiento del jurel (Trachurus trachurus Linnoesas) de la Division 1.3 del Area de ICSEAF. Colln.Scient.pap.int. Comm SE/-tl.Fish.8(2) : 255–266.
Staicu, I.1980 Report of the results of the researches carried out in the east-Central area of the Atlantic Ocean, on the continental shelf of the I.R.Mauritania, between March and May, 1980 (en roumain) : Halieutica 4 : 9–24.
Staicu, I., 1974 et C. Maxim, Observations sur la biologie et la dynamique du maquereau espagnol (Scomber japonicus colias Gmelin) dans l'Atlantique Est-Central. Cercetari marine I.R.C.R. 7 : 113–128.
Staicu, I., 1975 C. Maxim, et I. Panait, Recherches ichthyologiques roumaines dans la zone de l'Atlantique Est-Central, en 1974. Cercetari marine I.R.C.M. 8 : 119–135.
Staicu, I., 1976 C. Maxim, et I. Panait, Recherches roumaines sur l'ichthyofaune de l'Atlantique en face du Sahar espagnol pendant la période 1973–1975. Cercetari marine I.R.C.M. 9 : 191–204.
Starks, E.C., 1910 On the osteology and mutual relationship of the fishes belonging to the family Scombridae. J. Morph. 21 (1).
Terre, J.J., 1976 Crecimiento y fecundidad del Jurel (Trachurus trachurus, Linnaeus) de las divisiones 1.3 y 1.4 del area de CIPASO, ISCSEAF Coll. Sci.Pap. 3 : 39–44.
Thorne, R.E., 1977 O.A. Mathisen, R.J. Trumble and M. Blackburn, Distribution and abundance of pelagic fish off Spanish Sahara during CUEA Expedition Joint 1. Deep Sea Res. 24 : 75–82.
Trewavas; E., 1946 Fishes of the genus Decapterus at St. Helena, Ann.Mag.Nat. Hist. Sér. 11, 13 : 623–625.
Vasiliev, O.P. et A.I. 1966 Cherstnev, Investigations hématologiques des chinchards de l'Atlantique Est. Rec.Informat. Scient. Tech. (VNIRO) 5: 21–26 (en russe).
Villegas, L., 1972 Abundance estimates of mackerel found east of Abidjan (Ivory Coast) during 1971 cold season. Rapp.Sci.Proj.Dev.Pêche.Pélag. Côt. FAO/PNUD, Abidjan (12/72) : 1–24.
Vyskrebentsev,B.V.,1970 Data on biology of mackerel Scomber japonicus colias Gmelin, of the West African coast. Tr. AzCherNIRO 29 : 114–167 (en russe).
Ware, D.M., 1977 Spawning time and size of atlantic mackerel Scomber scombrus in relation to the plankton. J.Fish.Res.Bd.Canada,34 : 2308–2315.
Weiss, R., 1971 On the occurrence of fish schools at the water surface on the fishing grounds off NW Africa. Fischerei-Forsch., Rostock, 9 : 11–14 (en allemand).
Weiss, R., 1974 Feeding behaviour and formation of fish concentrations in the chub mackerel (Scomber colias) in the NW African fishing grounds ICES.C.M. 1974 J : 15 Pelagic fish (Southern) Committee.
Weiss, R., 1975 Chub mackerel fishery on the fishing grounds of NW Africa and long-term temperature conditions ICES CM/J : 14
Weiss, R., 1980 Research into the population differentiation in the chub mackerel Scomber japonicus Houttuyn off NW Africa. Fischerei-Forsch., Rostock, 18 : 11–14 (en allemand).
Weiss, R., 1981 On the reproductive biology of the chub mackerel (Scomber japonicus Hottuyn, 1872) in the NW African waters. Fischerei-Forsch., Rostock 19 : 19–29 (en allemand).
Wengrzyn, J.,1975 Age and grouwth of T. trachurus L., from North West african waters. ICES CM. J : 19 : 17 p.
Wengrzyn, J., 1976 The catches and characteristics of the SW African horse mackerel stock in the exploitation phase. ICSEAF. Coll.Sci.Pap. 3 : 45–49.
Wengrzyn, J., 1976 Results of investigations on age and growth rate of the horse mackerel (Trachurus trachurus) in South–West African waters. ICSEAF Coll.Sc.Pap., Vol. 3 : 51–57.
Williams, F., 1975 Les ressources halieutiques de l'Atlantique tropical du Centre-Est : Prospection, utilisation et gestion depuis 1960 Tech.Sér.IOC.11 : 34–52.
Wysokinski, A. 1973 Polowy i zarys ostrobokow (Trachurus trachurus i Trachurus trecae) i chropika (Caranx rhonchus) w rejonie polnocnozachodniej Afryki Mor. Inst. Ryb. Studia i Materialy, Ser. B. (23), Gdynia
Wysokinski, A., 1970 et Porebskiij, Pêche et biologie des chinchards T. trachurus T. trecae et C. rhonchus des eaux nord-ouest africaines. Manuscrit MIR.
Zei, M., 1969 Sardines and related species of the eastern tropical Atlantic. Review papers and Contributions of the Proceedings of the Symposium of Oceanography and fisheries Resources of the Tropical Atlantic, 20–28 Oct. 1966, Abidjan, pp. 101–108.
Zei, M., and M. Ansa-Emmim, 1970 The catches of Ghanaian long-range trawlers on the shelf off Mauritania and Senegal in 1965–67 Rap. P.-V. Réun. CIEM 159 : 261–263.
Zenkin, V.S., 1979 Différenciation génétique des populations de chinchards noirs (Trachurus trachurus L. et Trachurus trecae) dans la région du plateau continental de l'Afrique du Nord-Ouest. COPACE/PACE Séries 78/10 : 76–80.
Zenkin, V.S. et Y.A. Komarov, 1981 Genetic differentiation in the Cape horse mackerel (Trachurus trachurus capensis Castelnau) population. Colln.Scient.Pap.int. Comm SE Atl. Fish. 8(2) : 291–298.
Zenkin, V.S., 1976 O.D. Losev, and E.I. Riazantseva, Biochemical and seriological investigations of population composition of horse mackerel Trachurus trachurus. L., of the West African shelf, pp. 23–24 in “Problems of investigation of pelagic fish and invertebrates of the Atlantic Ocean”. Rep. of Conference of young Scientists, Kaliningrad (en russe).
Zenkin, V.S., 1976 E.I. Riazantseva and O.D. Losev, Biochemical polymorphism of muscle esterases and population structure of horse mackerel Trachurus trachurus trachurus L., and bigeye scad of the Eastern Central Atlantic pp. 209–210 in “Ecological physiology and biochemistry of fishes”, Vol. 2, Rep. to the 4th All-Union Conf. Astrakhan (en russe).
Zenkin, V.S., E.I., Riazantseva and O.D. Losev, 1979 polymorphysm of muscle esterases and analysis of the population structure of horse mackerels, Trachurus trachurus trachurus L. and Trachurus trachurus capensis Cast. of the West African shelf. pp. 94–98 in “Biochemical and population genetics of fishes”, Coll.Sci.Papers L. (en russe).
Zharov, V.L. and A.M. Zhudova, 1964 Some data on the occurence of larvae of Scombriform species (order Perciformes, sub-order Scombridei) in the open parts of the tropical Atlantic. ICES CM 1964 Scombriform Fish Comm. (81) : 6p. (also issued as Tr.AtlantNIRO 18 : 201–214 (1967).
RESUEM DE LA SITUATION DE LA PECHE AUX CHINCHARDS ET MAQUEREAUX
EN REPUBLIQUE DU CAP VERT
par Helena Santa Rita Vieira et Mecildes Dupret de Melo
1. INTRODUCTION
Le Service des statistiques de pêches au Cap Vert n'a commencé à fonctionner qu'en Décembre 1980. Le manque de cadres ajouté aux limitations d'ordre budgétaire n'ont pas encore permis l'installation d'un systéme de collecte des statistiques dans toutes les régions. De même, il n'existe aucune structure de recherches.
Malgré le fait que les chinchards sont capturés en petites quantités, ils sont d'une trés grande importance parce qu'ils sont utilisés comme appât vivant pour la pêche du thon. Ainsi, les résultats de cette pêche dépendent largement du succès de la pêche de l'appât convenable á chaque espéce de thonidés.
2. DONNEES DISPONIBLES
Les quelques données actuellement disponibles sont :
l'estimation des captures de la pêche artisanale en 1981 (Tableaux 1 et 3). Dans cette estimation, les statistiques de Selar et Decapterus rhonchus ont été confondues car l'identification de ces deux espèces n'a pas toujours été possible, de même pour les Decapterus macarellus et D. punctatus;
les rapports de visites au marché de Praia pendant l'année 1982, où nous avons enregistré la fréquence d'occurence (Tableau 2) et la fréquence mensuelle des tailles des Selar, Decapterus macarellus et D. punctatus (Tableau 4) ;
les captures de la pêche industrielle qui n'ont pas été utilisées comme appât (Tableau 5);
les rapports des recherches menées par la RDA (N/O “Ernst Haeckel”) en avril et octobre 1976 et par le N/O “Dr. Fridtjof Nansen” en novembre 1982 (voir Figure 1).
Les chinchards sont capturés : (i) par 26 thoniers afins d'être utilisés comme appât vivant (1981), (ii) par 3 bateaux, qui ne font que la pêche d'appât par les thoniers, l'excédent étant vendu au marché soit frais, en farine ou en boite, (iii) par les pirogues artisanales.
Les engins utilisés sont les sennes de plage, les sennes tournantes et les lignes.
Les espèces concernées sont au Cap Vert :
Selar crumenophthalmus et Decapterus rhonchus, qui sont trouvées pendant presque toute l'année au marché de Praia et qui ont également été trouvées par les deux navires de recherches cités ci-dessus.
Trachurus trachurus, trouvée seulement au mois d'octobre 1982 au marché de Praia en petites quantités. Décrite sous le nom erroné de Trachurus picturatus, elle figurait parmi les trois espèces les plus fréquentes capturées par le N/O “Ernst Haeckel” et était plus fréquente en octobre qu'en avril. Un seul exemplaire de cette espèce a été trouvé par le N/O “Dr. Fridtjof Nansen” en novembre 1981.
Decapterus macarellus et Decapterus punctatus, présentes au marché de Praia pendant toute l'année 1981, sauf aux mois de juin et d'août, ont également été trouvées par les deux navires de recherche. Le rapport du “Dr. Fridtjof Nansen” indique que en général, ces stocks ne sont pas mélangés.
Scomber japonicus n'a jamais été trouvée au marché de Praia, ni par le N/O “Dr. Fridtjof Nansen”. Le N/O “Ernst Haeckel” l'a par contre, capturée en petites quantités.
| Espèces | ENGINS | ||
|---|---|---|---|
| Lignes | Filets | TOTAL | |
| Katsuwonus pelamis | 4 | 4 | |
| Thunnus albacares | 3 967 | 3 967 | |
| Thunnus obesus | 41 | 41 | |
| Auxis + Euthynnus | 1 | 1 | |
| Acanthocybium solandri | 2 272 | 2 272 | |
| Caranx spp. | 81 | 81 | |
| Selar + Decapterus rhonchus | 55 | 553 | 608 |
| Decapterus spp. | 28 | 52 | 80 |
| Caranx lugubris (?) | 9 | 9 | |
| Selene dorsalis | 8 | ||
| Trachurus spp. | 2 | 5 | 7 |
| Lichia spp. | 27 | 27 | |
| Elagatis bipinnulatus | 2 | 2 | |
| Mycteroperca rubra | 7 | 7 | |
| Epinephelus guaza | 4 | 4 | |
| Cephalopholis taeniops | 82 | 82 | |
| Sphyraena guachancho | 40 | 40 | |
| Muraena spp. | 36 | 36 | |
| Bodianus spp. | 42 | 42 | |
| Smaris melanurus | 23 | 23 | |
| Callyodon hoefleri | 51 | 51 | |
| Belone spp. | 2 | 2 | |
| Hemirhamphus brasiliensis | 1 | 1 | |
| Exocoetidae | 17 | 17 | |
| Sardinella maderensis | 10 | 100 | 110 |
| Coryphaena hippurus | 61 | 61 | |
| Lutjanus spp. | 3 | 3 | |
| Megalops atlanticus | 1 | 1 | |
| Parapristipome humile | 1 | 1 | |
| Diplodus spp. | 1 | 1 | |
| AUTRES | 551 | 2 | 553 |
| TOTAL | 7 429 | 713 | 8 172 |
| NOMBRE DE SORTIES | 181.575 | 4.599 | 186.174 |
| Mois | J | F | M | A | M | J | Jl | A | S | O | N | D |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nombre de visites | 13 | 15 | 20 | 19 | 18 | 16 | 14 | 18 | 19 | 21 | 25 | 26 |
| Selar + D. rhonchus | 12 | 11 | 13 | 18 | 16 | 16 | 14 | 17 | 16 | 21 | 25 | 21 |
| Decapterus sp. | 3 | 2 | 1 | 1 | 1 | 0 | 4 | 9 | 6 | 10 | 26 | 29 |
| Trachurus trachurus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Pirogues | ||||||
|---|---|---|---|---|---|---|
| Motorisées | à rames | à voile | TOTAL | Nombre de pêcheurs | Nombre de plages de débarquement | Engins |
| 171 | 758 | 61 | 990 | 2 669 | 70 | Ligne |
| Ligne + harpon | ||||||
| Sennes de plage : 101 | ||||||
| Selar | Decapterus sp. | Trachurus | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1(cm) | J | F | M | A | M | O | N | D | J | F | M | O | N | D | 1(cm) | OCT.82 |
| 8 | 1 | |||||||||||||||
| 9 | 9 | 1 | ||||||||||||||
| 10 | 13 | 3 | ||||||||||||||
| 11 | 25 | 10 | ||||||||||||||
| 12 | 2 | 54 | 27 | 4 | ||||||||||||
| 13 | 7 | 55 | 44 | 6 | 5 | 28 | 1 | |||||||||
| 14 | 2 | 36 | 81 | 17 | 4 | 4 | 29 | |||||||||
| 15 | 11 | 62 | 25 | 9 | 15 | |||||||||||
| 16 | 3 | 27 | 37 | 18 | 5 | 30 | 4 | |||||||||
| 17 | 2 | 1 | 22 | 14 | 7 | 31 | ||||||||||
| 18 | 9 | 6 | 6 | 6 | 7 | 5 | 16 | 6 | 32 | |||||||
| 19 | 10 | 3 | 18 | 22 | 1 | 2 | 3 | 1 | 44 | 34 | 33 | 3 | ||||
| 20 | 21 | 10 | 38 | 11 | 15 | 4 | 2 | 5 | 43 | 47 | 34 | 2 | ||||
| 21 | 2 | 17 | 22 | 5 | 44 | 32 | 23 | 6 | 38 | 61 | ||||||
| 22 | 7 | 5 | 15 | 3 | 61 | 100 | 67 | 2 | 31 | 73 | 35 | |||||
| 23 | 21 | 11 | 22 | 26 | 77 | 88 | 7 | 5 | 1 | 19 | 58 | |||||
| 24 | 21 | 12 | 22 | 42 | 35 | 37 | 5 | 5 | 13 | 24 | ||||||
| 25 | 4 | 5 | 7 | 15 | 34 | 5 | 11 | 6 | ||||||||
| 26 | 4 | 10 | 4 | 4 | 19 | 16 | ||||||||||
| 27 | 1 | 1 | 2 | |||||||||||||
| 28 | 1 | 2 | ||||||||||||||
| TOTAL | 109 | 11 | 80 | 152 | 20 | 406 | 531 | 369 | 26 | 10 | 10 | 59 | 266 | 327 | 10 | |
| MOIS | BATEAU | POSITION | ENGIN | ESPECES | |||
|---|---|---|---|---|---|---|---|
| Decapterus(MT) | Selar + Caranx (MT) | ||||||
| 06 | Cruzinha | CANAL DE | S. VICENTE | lignes | 0.4 | - | |
| 7 | " | " | " | +NN St. | |||
| ANTAO | 0.4 | - | |||||
| 8 | " | " | " | " | 12.0 | - | |
| 9 | " | " | " | " | 11.0 | 0.2 | |
| 10 | " | " | " | " | 10.0 | 0.0 | |
| 11 | " | " | " | " | 3.0 | - | |
| 12 | " | " | " | " | 6. | - | |
| 06 | Frigos(2) | TARRAFAL St. ANTAO | Senne | 12.0 | - | ||
| 07 | " | " | " | Tourn | 4.0 | - | |
| 08 | " | " | " | " | 2.0 | - | |
| 10 | Belfish | TARRAFAL - St. NICOLAU | " | 2.0 | - | ||
| 11 | " | " | " | " | 5.0 | - | |
| 12 | " | " | " | " | 8.0 | - | |
| 10 | Persistencia | S.FRANCISCO (STIAGO) | " | 0.1 | |||
| 10 | Rabil | S.TOME | " | 10.0 | 2.3 | ||
| 12 | " | S.TOME E PORTO DA PRAIA | 1.9 | 1.4 | |||
LA PECHE ROUMAINE ET L'EVALUATION DES STOCKS DE CHINCHARDS
DANS LA REGION NORD DU COPACE
par
I. Staicu et C. Maxim
Institut Roumain de Recherches Marines, Constantza, Roumanie
1. LA PECHE ROUMAINE
Les navires de pêche roumains ont commencéleurs activités dans l'Atlantique Centre-Est en 1965 et dès lors cette zone a été fréquentée chaque année et chaque saison.
Pendant la période 1965–1971, l'activité de pêche s'est étendue sur un grand aréal (26–14°N), mais, par la limitation de l'accès aux ressources halieutiques suite à l'élargissement des zones d'intérêt économique, les activités de la flotte roumaine se sont limitées les dernières années à la seule zone économique de la République Islamique de Mauritanie (20°35–16°28 N), en condition de licence.
Le Tableau 1 résume tous les changements survenus dans l'historique de cette pêcherie. L'effort de pêche, les captures totales et les captures de chinchards ont constamment augmenté, surtout à partir de l'année 1978, après l'introduction des chalutiers de type Atlantique 2 et des superchalutiers. En général (Tableau 2), les captures des chinchards augmentent en relation avec l'augmentation de l'effort.
Trachurus trachurus L. domine dans les captures roumaines de chinchards ; le chinchard Cunène (Trachurus trachurus Cadenat) est séparé seulement pour les périodes 1973–1976 et 1981–1982. Dans les autres années, T, trecae est inclus dans la catégorie Trachurus spp. Les captures de ces deux espèces sont présentées par division statistique au Tableau 3.
2. DONNEES BIOLOGIQUES
2.1 Composition en taille et âge
La composition en tailles et classes d'âge, des captures des deux espèces varie d'une période à l'autre et même d'année en année, étant donné qu'à la base des captures sont des individus âgés d'un à 5 ans (Figs. 1, 2, 3, 4).
2.2 Croissance
Pour l'espèce Trachurus trachurus, les différences en croissance en longueur et en poids sont très petites entre mâles et femelles (Tableau 4). Le taux de croissance n'est pas constant durant toute la vie ; il est plus grand au cours des premières années (Tableau 5, 6, 7 et 8).
2.3 Rapport longueur-poids
Les rapports longueur-poids ont été calculés par année :
| Trachurus trachurus | Trachurus trecae | |
|---|---|---|
| 1973 | Wt = 0,0197 L2,85 | Wt = 0,0247 L2,80 |
| 1974 | Wt = 0,0106 L3,05 | Wt = 0,0314 L2,72 |
| 1975 | Wt = 0,2077 L2,85 | Wt = 0,0145 L2,96 |
| 1976 | Wt = 0,0089 L3,09 | Wt = 0,0101 L3,07 |
| 1977 | Wt = 0,0292 L2,73 | |
| 1978 | Wt = 0,0208 L2,83 | |
| 1979 | Wt = 0,0446 L2,60 | |
| 1980 | Wt = 0,0125 L2,98 | |
| 1981 | Wt = 0,0198 L2,95 | Wt = 0,0083 L3,11 |
| 1982 | Wt = 0,0128 L2,99 | Wt = 0,0248 L2,80 |
2.4 Rapport taille-âge et poids-âge
2.4.1 Trachurus trachurus (Figs. 8 et 10)
Lt = 38,98 (1-e-0,2775724 ( t + 1,1613564))
Wt = 760,36 (1-e-0,2775724 ( t + 1,1613564))3
2.4.2 Trachurus trecae (Figs. 9 et 11)
Lt = 41,75 (1-e-0,2166723 (t + 1,4034547)) Wt = 1005 (1-e-0,2166723 (t + 1,4034547))3
| Effort de pêche | Capture (tonnes) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Année | Secteur ° N | No. navires | Jours | Heures | Coups Chalut | Total Zone | T.trachurus | T.trecae | Total Chinchards |
| 1965 | 26–14 | 2 | - | 1 527 | - | 2 732 | 2 000 | - | 2 000 |
| 1966 | 26–14 | 2 | - | 4 990 | - | 5 096 | 3 800 | - | 3 800 |
| 1967 | 26–14 | 2 | - | 3 085 | - | 8 601 | 4 800 | - | 4 800 |
| 1968 | 26–14 | 2 | - | 2 193 | - | 5 098 | 2 700 | - | 2 700 |
| 1969 | 26–14 | 2 | - | 3 850 | - | 7 709 | 4 600 | - | 4 600 |
| 1970 | 28–14 | 5 | 343 | 3 135 | 1 624 | 4 838 | 2 300 | - | 2 300 |
| 1971 | 29–12 | 9 | 999 | 8 361 | 4 715 | 15 945 | 3 800 | - | 3 800 |
| 1972 | 26–17 | 17 | 1 487 | 12 225 | 6 503 | 33 465 | 13 800 | - | 13 800 |
| 1973 | 26–20 | 18 | 1 909 | 17 040 | 8 559 | 43 863 | 6 613 | 22 491 | 29 104 |
| 1974 | 26–20 | 23 | 2 914 | 27 655 | 14 728 | 72 004 | 21 862 | 12 678 | 34 540 |
| 1975 | 26–20 | 21 | 2 932 | 24 556 | 13 797 | 78 084 | 20 254 | 6 158 | 26 412 |
| 1976 | 25–20 | 22 | 1 457 | 13 867 | 6 798 | 35 804 | 13 016 | 460 | 13 476 |
| 1977 | 29–17 | 21 | 2 964 | 31 127 | 13 573 | 78 453 | 38 643 | - | 38 643 |
| 1978 | 24–05 | 27 | 1 837 | 18 419 | 8 053 | 49 202 | 33 740 | - | 33 740 |
| 1979 | 23–09 | 6 | 1 030 | 9 669 | 4 369 | 45 222 | 41 464 | - | 41 464 |
| 1980 | 20–16 | 17 | 1 842 | 20 647 | 8 503 | 77 509 | 64 034 | - | 64 034 |
| 1981 | 20–16 | 17 | 2 375 | 29 099 | 11 536 | 85 567 | 18 878 | 39 203 | 58 081 |
| 1982 | 20–16 | 15 | 2 768 | 35 325 | 11 457 | 84 142 | 16 114 | 18 559 | 34 673 |
| EFFORT | C.P.U.E. | ||||
|---|---|---|---|---|---|
| Année | Jours effectués | Heures chalutage | Tonnes jour | Tonnes heure | Captures (tonnes) |
| 1965 | - | 1 527 | - | 1,300 | 2 000 |
| 1966 | - | 4 990 | - | 0,760 | 3 800 |
| 1967 | - | 3 085 | - | 1,550 | 4 800 |
| 1968 | - | 2 193 | - | 1,230 | 2 700 |
| 1969 | - | 3 850 | - | 1,190 | 4 600 |
| 1970 | 343 | 3 155 | 6,700 | 0,730 | 2 300 |
| 1971 | 999 | 8 361 | 3,800 | 0,450 | 3 800 |
| 1972 | 1 486 | 12 225 | 9,280 | 1,129 | 13 800 |
| 1973 | 1 909 | 17 040 | 15,224 | 1,708 | 29 104 |
| 1974 | 2 914 | 27 655 | 11,853 | 1,249 | 34 540 |
| 1975 | 2 932 | 24 556 | 9,373 | 1,076 | 26 412 |
| 1976 | 1 457 | 13 867 | 9,249 | 0,972 | 13 476 |
| 1977 | 2 964 | 31 127 | 13,042 | 1,241 | 38 643 |
| 1978 | 1 837 | 18 419 | 18,367 | 1,832 | 33 740 |
| 1979 | 1 030 | 9 669 | 40,256 | 4,288 | 41 464 |
| 1980 | 1 842 | 20 647 | 30,500 | 3,000 | 64 034 |
| 1981 | 2 375 | 20 099 | 24,455 | 1,995 | 58 081 |
| 1982 | 2 768 | 35 325 | 12,581 | 0,982 | 34 673 |
3. EVALUATION DU STOCK DE CHINCHARD T. TRACHURUS
La méthode de l'analyse des cohortes a été utilisée pour évaluer le stock de chinchard. Les principaux paramètres que nous avons considérés constants et importants au cours de toute la période d'exploitation ont été : la mortalité naturelle et la relation rendement-recrutement.
3.1 Calcul des paramètres
3.1.1 La mortalité naturelle
| METHODE | M | |
|---|---|---|
| 1. | Z des courbes de capture et l'effort de pêche | 0,630 |
| 2. | F de VPA, selon l'effort de pêche | 0,600 |
| 3. | Z des courbes d'âge de VPA, selon l'effort de pêche | 0,560 |
| 4. | Courbes de capture pour les périodes d'effort maximum | 0,401 |
| et minimum | 0,440 | |
| 5. | Méthodes Silliman (1943) | 0,569 |
| 6. | Valeur moyenne | 0,533 |
Pour estimer l'abondance du stock de chinchard, M = 0,5 a été considéré comme valeur suffisamment proche de la réalité.
3.1.2 La relation rendement-recrutement
On a utilisé l'équation du recrutement de Beverton et Holt (1957), ayant la forme suivante :
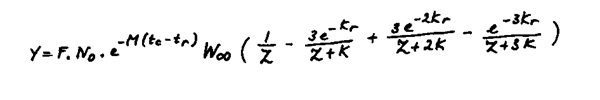
et les paramètres suivants:
| M = 0,5 | to = -1,1613 |
| K = 0,2775 | tr = tc = 1 an |
| W = 760,36 | tmax = 12 ans |
| r = tc - to = 2,1613 |
| 34.1.3 | 34.3.1 | TOTAL | |||||
|---|---|---|---|---|---|---|---|
| T.trachurus | T.trecae | TOTAL | T.trachurus | T.trecae | TOTAL | Captures | |
| 1965 | 1 445 | - | 1 445 | 555 | - | 555 | 2 000 |
| 1966 | 3 492 | - | 3 492 | 308 | - | 308 | 3 800 |
| 1967 | 1 806 | - | 1 806 | 2 994 | - | 2 994 | 4 800 |
| 1968 | 953 | - | 953 | 1 747 | - | 1 747 | 2 700 |
| 1969 | 3 276 | - | 3 276 | 1 324 | - | 1 324 | 4 600 |
| 1970 | 1 774 | - | 1 774 | 526 | - | 526 | 2 300 |
| 1971 | 2 475 | - | 2 475 | 1 325 | - | 1 325 | 3 800 |
| 1972 | 10 671 | - | 10 671 | 3 073 | - | 3 073 | 13 744 |
| 1973 | 22 491 | 6 613 | 29 104 | - | - | - | 29 104 |
| 1974 | 21 862 | 12 678 | 34 540 | - | - | - | 34 540 |
| 1975 | 18 377 | 6 158 | 24 535 | 1 877 | - | 1 877 | 25 412 |
| 1976 | 13 016 | 460 | 13 476 | - | - | - | 13 476 |
| 1977 | 33 391 | - | 33 391 | 5 252 | - | 5 252 | 38 643 |
| 1978 | 21 308 | - | 21 308 | 12 432 | - | 12 432 | 33 740 |
| 1979 | 20 265 | - | 20 265 | 21 199 | - | 21 199 | 41 464 |
| 1980 | 35 524 | - | 35 524 | 28 510 | - | 28 510 | 64 034 |
| 1981 | 9 749 | 25 474 | 35 223 | 9 129 | 13 729 | 22 858 | 58 081 |
| 1982 | 9 852 | 6 442 | 16 294 | 6 262 | 12 117 | 18 379 | 34 673 |
| sexe | AGE | |||||||
|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| Femelles | cm | 20,16 | 23,24 | 26,90 | 30,14 | 33,03 | 35,03 | 36,66 |
| g | 118 | 153 | 247 | 340 | 453 | 542 | 610 | |
| Mâles | cm | 19,11 | 23,28 | 26,76 | 30,00 | 32,76 | 34,21 | 37,00 |
| g | 105 | 155 | 244 | 334 | 438 | 533 | 625 | |
| Année | CLASSES D'AGE | No. d'individus | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0+ | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| 1972 | - | - | 21,04 | 24,71 | 28,61 | 31,34 | 34,00 | 35,29 | 36,25 | 301 | ||
| 1973 | - | - | 21,21 | 24,01 | 28,22 | 30,72 | 33,18 | - | - | 1 659 | ||
| 1974 | 11,37 | 16,85 | 20,88 | 24,78 | 28,83 | 31,17 | 33,65 | - | - | 4 172 | ||
| 1975 | 12,80 | 17,61 | 21,72 | 24,38 | 28,04 | 30,20 | - | - | 2 743 | |||
| 1976 | 13,09 | 17,99 | 21,44 | 23,80 | 28,30 | 31,87 | 33,33 | 35,00 | 2,345 | |||
| 1977 | 11,16 | 17,48 | 21,62 | 24,04 | 28,07 | 31,00 | - | 35,50 | 1 616 | |||
| 1978 | - | 18,09 | 22,13 | 24,86 | 28,57 | 30,32 | 32,30 | 35,20 | 36,12 | 2 162 | ||
| 1979 | - | 18,73 | 22,80 | 25,96 | 28,90 | 30,73 | 33,60 | - | 1 929 | |||
| 1980 | - | 17,62 | 21,82 | 25,23 | 28,38 | 32,05 | 33,80 | - | 3 956 | |||
| 1981 | - | 18,60 | 22,22 | 26,83 | 29,08 | 32,08 | 33,60 | 35,11 | 2 758 | |||
| 1982 | 11,00 | 18,64 | 22,29 | 26,36 | 29,44 | 32,62 | 33,80 | - | 2 426 | |||
| Moyenne | 11,69 | 17,77 | 21,88 | 25,35 | 28,77 | 31,27 | 33,33 | 35,25 | 36,15 | 36,80 | 37,30 | 26 637 |
| Année | CLASSES D'AGE | No. Individus | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0+ | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | ||
| 1972 | - | - | 109,5 | 186,4 | 268,3 | 348,4 | 420,0 | 568,0 | 640,0 | - | - | 301 |
| 1973 | - | - | 118,7 | 166,4 | 207,4 | 343,2 | 407,7 | - | 685,0 | 1 659 | ||
| 1974 | 15,2 | 67,9 | 103,3 | 165,6 | 323,5 | 461,7 | 520,6 | - | 4 742 | |||
| 1975 | 23,2 | 81,6 | 123,1 | 186,6 | 290,3 | 395,3 | - | - | - | - | - | 2 743 |
| 1976 | 18,0 | 68,0 | 119,0 | 174,0 | 310,0 | 434,0 | 515,0 | 560,0 | 2 345 | |||
| 1977 | 21,7 | 57,4 | 98,2 | 169,3 | 286,1 | 361,1 | - | 580,0 | 1 616 | |||
| 1978 | - | 66,3 | 85,0 | 162,4 | 278,3 | 374,4 | 488,5 | 590,0 | 610,0 | 2 162 | ||
| 1979 | - | 62,4 | 117,7 | 164,2 | 254,6 | 339,0 | 400,0 | - | 1 929 | |||
| 1980 | - | 67,0 | 118,0 | 163,0 | 223,0 | 317,0 | 417,0 | - | 3 956 | |||
| 1981 | - | 87,0 | 125,0 | 175,0 | 252,0 | 349,0 | 478,0 | 557,0 | 2 758 | |||
| 1982 | 18,0 | 85,0 | 123,0 | 162,0 | 257,8 | 325,8 | 421,5 | - | - | - | - | 2 426 |
| Moyenne | 16,1 | 73,2 | 111,8 | 167,4 | 252,8 | 356,7 | 442,9 | 574,9 | 608,3 | 640,0 | 685,0 | 26 637 |
| Année | CLASSES D'AGE | No. d'individus | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| 1973 | 18,83 | 22,39 | 26,04 | 29,40 | - | 33,45 | - | - | 796 |
| 1974 | 16,28 | 21,33 | 25,20 | 28,64 | 31,36 | 33,00 | 35,50 | 37,00 | 1 500 |
| 1975 | 16,25 | 21,70 | 25,66 | 28,80 | 31,19 | 32,90 | - | - | 1 313 |
| 1976 | 17,22 | 22,00 | 26,13 | 28,90 | 31,20 | 33,60 | - | - | 1 200 |
| 1981 | 15,38 | 20,57 | 25,42 | 28,66 | 31,45 | 33,76 | 35,21 | 36,33 | 5 439 |
| 1982 | 17,02 | 21,47 | 25,77 | 28,94 | 31,09 | 33,50 | - | - | 1 574 |
| Moyenne | 16,91 | 21,35 | 25,56 | 28,75 | 31,41 | 33,72 | 35,21 | 36,42 | 11 822 |
| Année | CLASSES D'AGE | No. d'individus | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| 1973 | 91,3 | 157,9 | 264,9 | 316,2 | - | 460,0 | - | - | 796 |
| 1974 | 58,5 | 120,6 | 228,5 | 337,7 | 423,6 | 490,0 | 658,0 | 760,0 | 1 500 |
| 1975 | 97,2 | 124,7 | 188,4 | 331,4 | 407,8 | 503,7 | - | - | 1 313 |
| 1976 | 86,5 | 140,5 | 260,2 | 380,5 | 430,6 | 510,2 | - | - | 1 200 |
| 1981 | 48,0 | 109,0 | 171,0 | 315,0 | 415,0 | 520,0 | 621,0 | 723,0 | 5 439 |
| 1982 | 77,3 | 144,7 | 244,6 | 397,5 | 492,0 | 570,0 | - | - | 1 574 |
| Moyenne | 77,8 | 125,8 | 207,6 | 333,6 | 421,2 | 518,3 | 622,0 | 728,8 | 11 822 |
Pour F0.1 = 0,27 on obtiendrait une capture d'environ 300 000 tonnes pour l'année 1982. Dans ces conditions, il y aurait une augmentation dans le recrutement de 6% au début de l'année 1983 par rapport à 1981.
3.2 Etat du stock
Afin d'estimer l'abondance numérique et la biomasse du stock, on a calculé, sur la base des données des recherches pour la période 1972– 1982, les variations en mortalité par pêche, abondance et recrutement.
La mortalité par pêche est résumée au Tableau 9, le recrutement au Tableau 10. Les Tableau 11, 12 et 13 donnent respectivement l'abondance numérique, la biomasse et les captures réalisables des chinchards en 1972–1981.
4. REFERENCES BIBLIOGRAPHIQUES
Allen, K.R., 1966 Some methods for Estimating Exploited Populations. J. Fish. Res.Bd.Canada, 23 (10) : 1553–1574.
Boely, T., 1979 et P. Fréon, Les ressources pélagiques côtières. In “Les ressources halieutiques de l'Atlantique Centre-Est”. Première partie : les ressources du Golfe de Guinée de l'Angola à la Mauritanie. FAO.Doc.tech.No.186 : 13–78.
Gulland, J.A., 1965 Manual of methods for fish stock assessment. Part. I Fish population analysis. FAO Fish.Th.Paper.No. 40.
Maxim, C. et Staicu I., 1980 Evaluation of the horse mackerel stock in the Northwest African area of Atlantic ocean. IRCM, Halieutica nr. 2 (14) : 103–116.
Maxim, C. et I. Staicu, 1982 Assessment of the Atlantic horse mackerel stock in the eastern–central Atlantic area (divisions 34.1.3 et 34.3.1) using provisional 1980 data. Recherches Marines No. 13 : 163–172.
Maxim, C. et I. 1980 Staicu, Assessment of the natural mortality of the horse mackerel (Trachurus trachurus trachurus L.) from the East-Central Area. Halieutica nr. 4 (16) : 79–89.
Maxim, C., 1982 Assessment of stock and calculation of recruitment and total admissible catch on the horse mackerel (Trachurus trachurus trachurus L.) in the East-Central Atlantic Ocean - Mauritanian Waters. Halieutica nr.2 (22) : 75–88.
| CLASSES D'AGE | 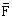 0 0 | 1 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | |||
| 1972 | 0,000 | 0,406 | 0,549 | 0,780 | 0,742 | 0,675 | 0,620 | 0,539 | 0,629 |
| 1973 | 0,000 | 0,513 | 0,535 | 0,540 | 1,005 | 0,910 | 0,620 | 0,589 | 0,687 |
| 1974 | 0,093 | 0,553 | 0,520 | 0,498 | 0,847 | 1,147 | 0,620 | 0,611 | 0,697 |
| 1975 | 0,080 | 0,528 | 0,603 | 0,718 | 0,886 | 0,861 | 0,620 | 0,614 | 0,703 |
| 1976 | 0,165 | 0,427 | 0,335 | 0,517 | 0,844 | 0,924 | 0,620 | 0,547 | 0,611 |
| 1977 | 0,003 | 0,327 | 0,491 | 0,473 | 0,380 | 0,740 | 0,620 | 0,433 | 0,505 |
| 1978 | 0,006 | 0,323 | 0,369 | 0,370 | 0,398 | 0,497 | 0,620 | 0,369 | 0,430 |
| 1979 | 0,000 | 0,316 | 0,312 | 0,404 | 0,439 | 0,555 | 0,953 | 0,426 | 0,497 |
| 1980 | 0,000 | 0,244 | 0,297 | 0,580 | 0,980 | 1,737 | 2,483 | 0,903 | 1,054 |
| 1981 | 0,000 | 0,488 | 0,395 | 0,711 | 1,576 | 1,757 | 2,749 | 1,096 | 1,279 |
| 1982 | 0,1501 | 0,4501 | 0,620 | 0,620 | 0,620 | 0,620 | 0,620 | 0,6202 | 0,6202 |
1 Valeurs estimées selon la capture connue.
2 Valeur moyenne pour deux et plusieurs années.
| Année | Age | B | U | R | W |
|---|---|---|---|---|---|
| 1975 | 0 | 00 | 0 | 0 | 0 |
| 1 | 17,8185 | 0,0436 | 0 | 0 | |
| 3 | 0,4097 | 0,3521 | 0 | 0 | |
| 4 | 0,5549 | 1,0000 | 0 | 0 | |
| 5 | 0,3238 | 1,0000 | 0 | 0 | |
| 6 | 0,5037 | 1,0000 | 0 | 0 | |
| 0 | |||||
| 1976 | 0 | 00 | 0 | - | - |
| 1 | 8,9243 | 0,0881 | 1,0000 | 0,5540 | |
| 2 | 0,4359 | 0,7986 | 0,9856 | 0,2040 | |
| 3 | 0,2550 | 1,0000 | 0,5962 | 0,0256 | |
| 4 | 0,3550 | 1,0000 | 0 | 0 | |
| 5 | 0,4565 | 0,9637 | 0 | 0 | |
| 6 | 0,7152 | 1,0000 | 0 | 0 | |
| 0,7836 | |||||
| 1977 | 0 | 00 | 0 | - | - |
| 1 | 1,6269 | 0,5685 | 0,8029 | 0,3436 | |
| 2 | 0,3458 | 0,8057 | 0,1724 | 0,0603 | |
| 3 | 0,4337 | 1,0000 | 0 | 0 | |
| 4 | 0,4837 | 0,9812 | 0 | 0 | |
| 5 | 0,2150 | 1,0000 | 0 | 0 | |
| 6 | 0,2129 | 1,0000 | 0 | 0 | |
| 0,4039 | |||||
| 1978 | 0 | 00 | 0 | - | - |
| 1 | 203,6211 | 0,0043 | 0,3296 | 0,1226 | |
| 2 | 0,8594 | 0,8930 | 0,1756 | 0,0590 | |
| 3 | 0,5849 | 1,0000 | 0,0011 | 0,0003 | |
| 4 | 0,4940 | 1,0000 | 0 | 0 | |
| 5 | 0,6050 | 1,0000 | 0 | 0 | |
| 6 | 0,5971 | 1,0000 | 0 | 0 | |
| 0,1819 | |||||
| 1979 | 0 | 00 | 0 | - | - |
| 1 | 19,0101 | 0,1300 | 0,5815 | 0,1929 | |
| 2 | 0,6317 | 0,6004 | 0,1247 | 0,0352 | |
| 3 | 0,2248 | 1,0000 | 0 | 0 | |
| 4 | 0,3514 | 1,0000 | 0 | 0 | |
| 5 | 0,2018 | 1,0000 | 0 | 0 | |
| 6 | 0,6259 | 1,0000 | 0 | 0 | |
| 0,2281 | |||||
| 1980 | 0 | - | - | - | - |
| 1 | - | - | - | - | |
| 2 | 0,7576 | 0,8215 | 0,1733 | 0,0276 | |
| 3 | 0,4109 | 0,9892 | 0,0007 | 0,0001 | |
| 4 | 0,5513 | 1,0000 | 0 | 0 | |
| 5 | 0,1281 | 1,0000 | 0 | 0 | |
| 6 | 0,2648 | 1,0000 | 0 | 0 | |
| 0,0277 | |||||
| Années | CLASSES D'AGES | Abondance totale | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | ||
| 1972 | 16 090,4 | 4 360,5 | 1 735,2 | 597,3 | 325,6 | 189,0 | 80,01 | 23 378,1 |
| 1973 | 19 763,9 | 6 273,4 | 1 668,5 | 609,5 | 152,2 | 88,1 | 61,7 | 28 617,3 |
| 1974 | 12 962,4 | 7 670,5 | 2 270,8 | 601,6 | 190,4 | 32,1 | 28,7 | 23 756,5 |
| 1975 | 17 667,1 | 5 935,1 | 2 622,7 | 814,6 | 189,6 | 49,2 | 10,5 | 27 288,8 |
| 1976 | 21 356,4 | 8 497,0 | 2 303,7 | 902,1 | 212,3 | 46,7 | 16,1 | 33 334,3 |
| 1977 | 13 571,2 | 10 224,5 | 3 272,9 | 942,5 | 345,7 | 57,7 | 15,2 | 28 429,5 |
| 1978 | 14 676,7 | 7 116,2 | 4 295,1 | 1 277,5 | 367,4 | 136,4 | 18,8 | 27 988,1 |
| 1979 | 10 000,01 | 7 718,0 | 3 141,4 | 1 815,5 | 519,8 | 140,2 | 41,9 | 23 376,8 |
| 1980 | 8 000,01 | 4 936,8 | 3 437,1 | 1 244,3 | 582,6 | 125,2 | 25,1 | 18 351,1 |
| 1981 | 7 000,01 | 2 395,1 | 2 198,1 | 1 304,0 | 291,2 | 100,8 | 9,9 | 13 299,1 |
| Année | CLASSES D'AGES | Biomasse (tonnes)2 | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | ||
| 1972 | 389 6271 | 293 025 | 190 002 | 111 359 | 87 360 | 65 855 | 33 600 | 981 201 |
| 1973 | 355 7501 | 439 141 | 198 051 | 101 424 | 31 572 | 30 231 | 25 155 | 1 003 574 |
| 1974 | 197 028 | 536 933 | 234 572 | 99 565 | 61 602 | 14 801 | 14 941 | 1 060 914 |
| 1975 | 409 876 | 484 306 | 270 921 | 152 010 | 55 044 | 19 460 | 5 290 | 1 396 907 |
| 1976 | 384 415 | 577 795 | 274 142 | 168 325 | 65 806 | 20 268 | 8 292 | 1 499 043 |
| 1977 | 294 494 | 586 889 | 321 394 | 159 539 | 107 161 | 20 821 | 6 597 | 1 496 895 |
| 1978 | 287 663 | 471 801 | 373 579 | 207 461 | 102 240 | 49 257 | 9 184 | 1 501 185 |
| 1979 | 180 000 | 481 604 | 369 740 | 298 103 | 132 343 | 47 524 | 20 475 | 1 439 789 |
| 1980 | 144 000 | 330 767 | 405 582 | 202 814 | 129 928 | 39 692 | 10 484 | 1 191 267 |
| 1981 | 126 000 | 208 377 | 274 763 | 228 194 | 73 379 | 35 194 | 4 688 | 887 595 |
1 En considérant un poids moyen de 18 g pour la classe d'âge de “O” an
2 50% du nombre d'individus âgés d'un an et 100% de deux années et plus
| Année | Captures réalisables1 (tonnes) | Captures réalisées (tonnes) |
|---|---|---|
| 1972 | 349 084 | 447 100 |
| 1973 | 403 553 | 486 300 |
| 1974 | 424 626 | 500 500 |
| 1975 | 483 611 | 452 700 |
| 1976 | 477 896 | 430 100 |
| 1977 | 427 309 | 491 000 |
| 1978 | 380 488 | 329 600 |
| 1979 | 391 730 | 249 500 |
| 1980 | 385 156 | 489 800 |
| 1981 | 305 312 | 400 0002 |
1 Calculée à partir de M = 0.5 et les montalités par pêche estimées
Paloheimo J.E. and L.M. Dickie, 1964 Abundance and fishing success. Rapp.Procès. Verbaux Réunions, Conseil Perm.Int.Exploration Mer, 155 (28) : 152 –162.
Ricker, W.E., 1965 Computations and Interpretation of Biological Statistics of Fish Populations. J.Fish.Res.Bd.Canada, Bull. 191 : 382 p.
Silliman, R.P., 1943 Studies on the Pacific pilchard or sardine (Sardinops caerula). 5. A method of computing mortalities and replacements. U.S. Fish. Wildl.Serv.Spec.Sci.Rep. No. 24 : 10p.
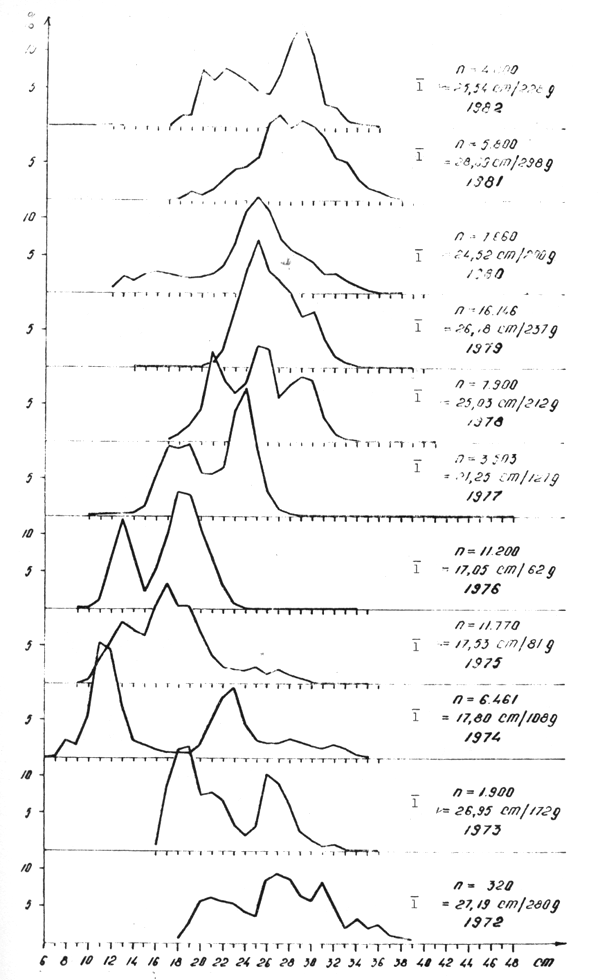
Figure 1 : Composition en longueurs des captures du chinchard Trachurus trachurus trachurus L. pêché dans l'Atlantique Centre-Est en 1972–1982.
Figure 2 : Composition en longueurs des captures du chinchard Trachurus trecae Cadenat pêché dans l'Atlantique Centre-Est en 1973– 1976 et 1981–1982.
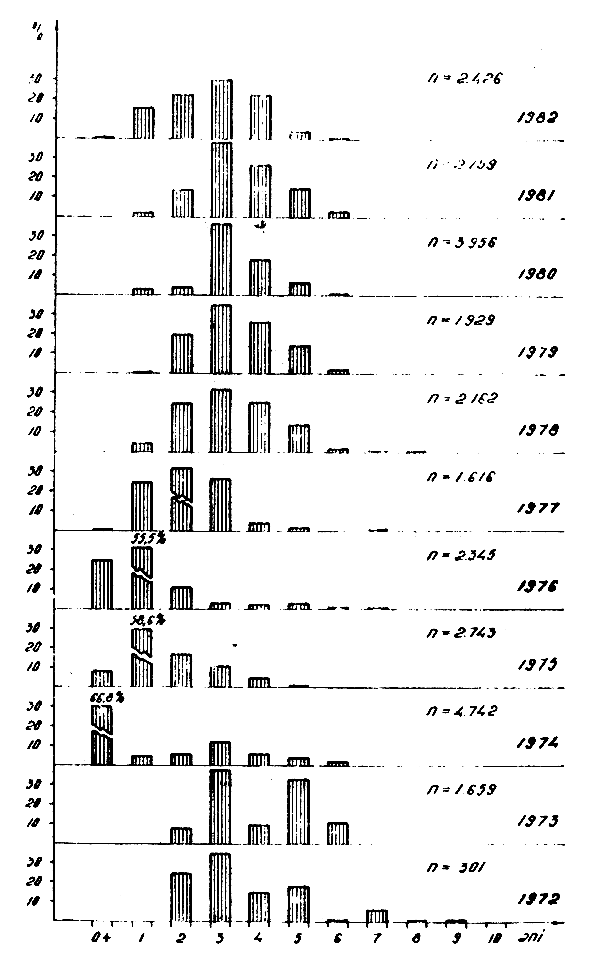
Figure 3 : Composition en classes d'âge des Trachurus trachurus trachurus L. pêchés dans l'Atlantique Centre-Est en 1972–1982.
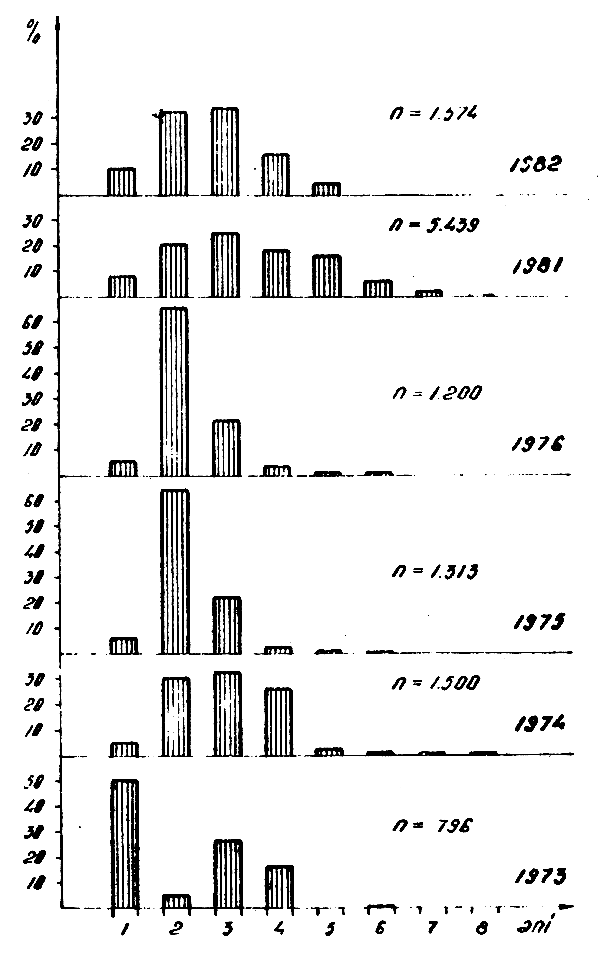
Figure 4 : Structure en âge des captures de Trachurus trecae pêchés dans l'Atlantique Centre-Est en 1973–1976 et 1981–1982
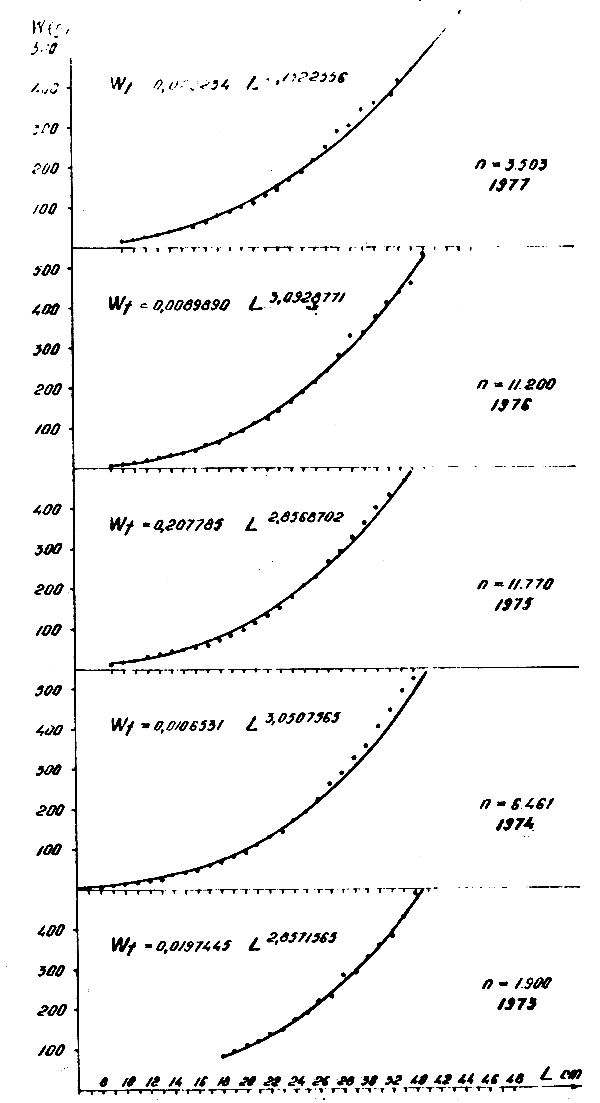
Figure 5 : Relation longueur-poids de Trachurus trachurus en 1973–1977
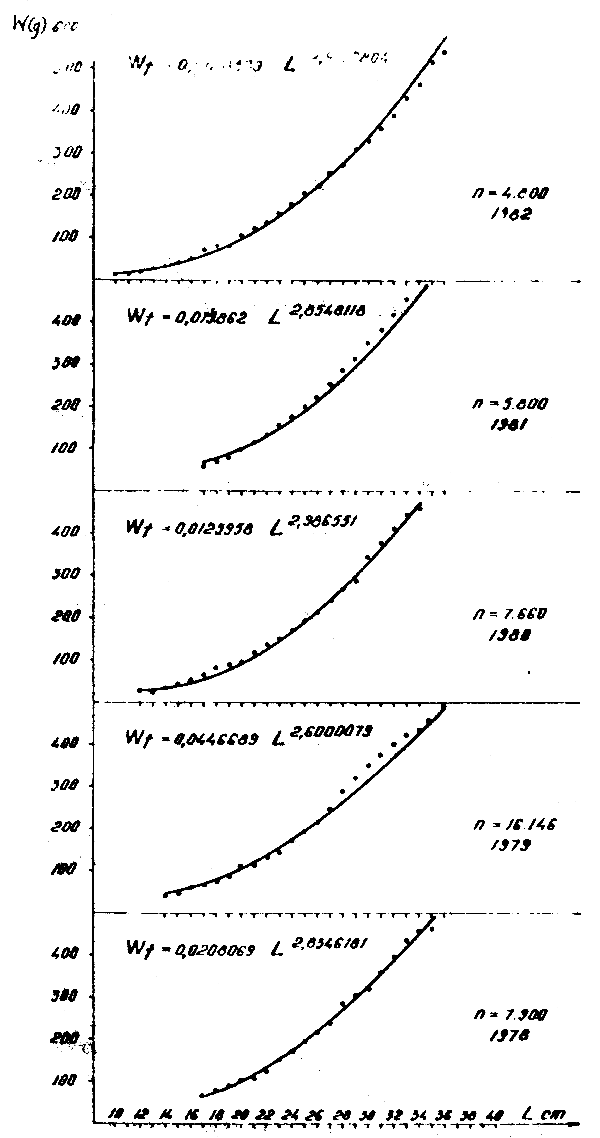
Figure 6 : Relation longueur-poids de Trachurus trachurus en 1978–1982
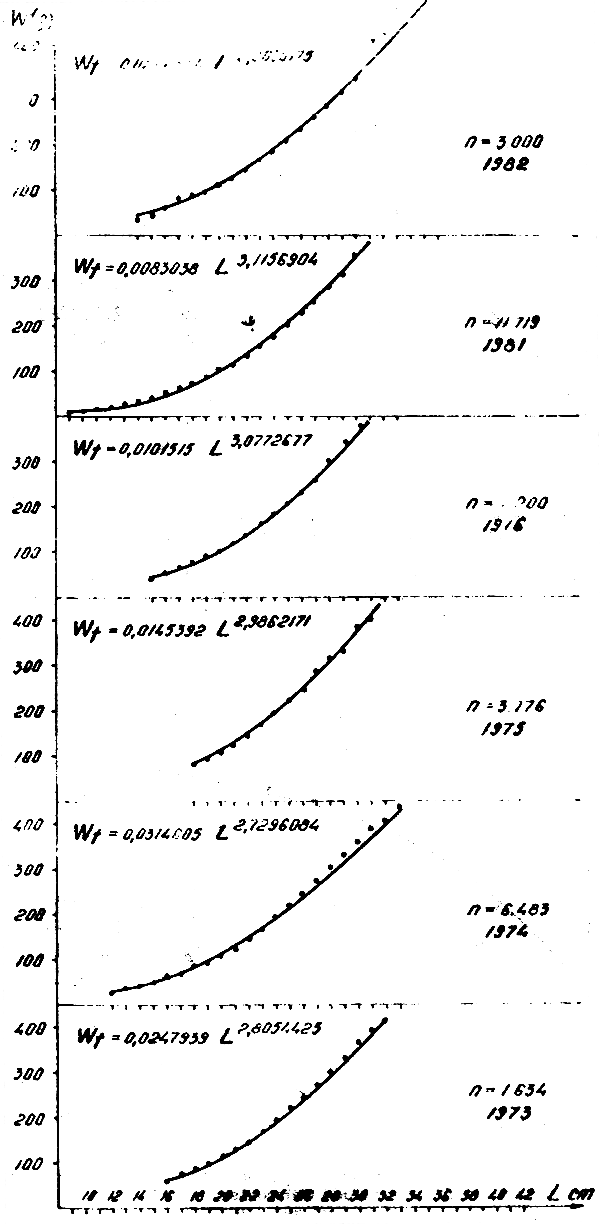
Figure 7 : Relation longueur-poids de Trachurus trecae, 1973–1976 et 1981–1982
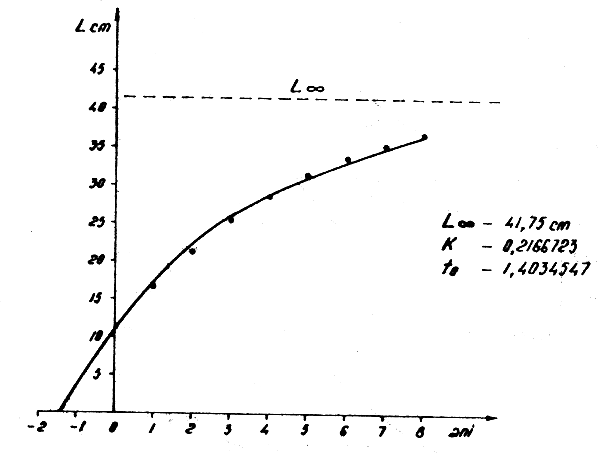
Figure 9 : Relation longueur-âge pour Trachurus trecae Cadenat. dans l'Atlantique Centre-Est en 1973–1976 et 1981–1982
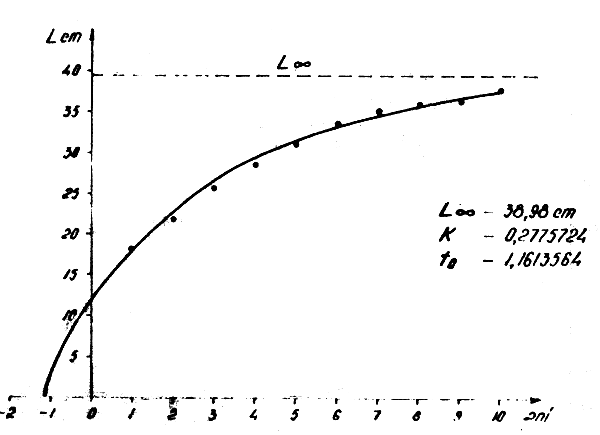
Figure 8 : Relation longueur-âge pour Trachurus trachurus trachurus dans l'Atlantique Centre-Est en 1972–1982.
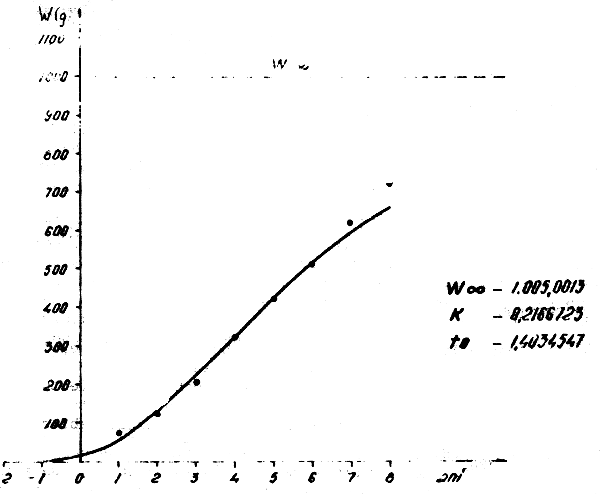
Figure 11 : Relation poids-âge pour Trachurus trecae dans l'Atlantique Centre-Est en 1973–1976 et 1981–1982
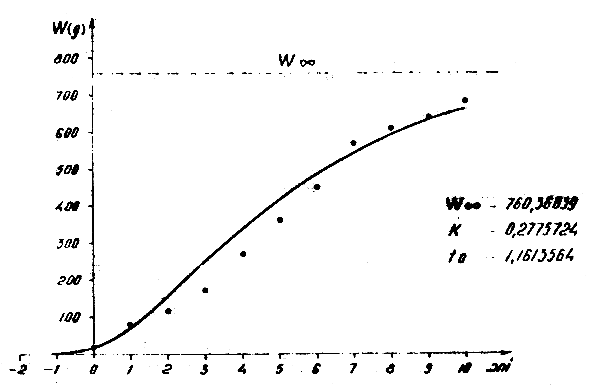
Figure 10 : Relation poids-âge pour Trachurus trachurus trachurus dans l'Atlantique Centre-Est en 1972–1982
RAPPORT SUR LES RESULTATS DES RECHERCHES ROUMAINES DANS LA ZEE
DE LA REPUBLIQUE ISLAMIQUE DE MAURITANIE EN 1982
par I. Staicu et C. Maxim
Institut Roumain de Recherches Marines, Constantza, Roumanie
1. INTRODUCTION
Pendant l'année 1982, les bateaux de pêche roumains ont exploité la zone côtière (navires du type B–410, 25 m) et la zone au large (chalutiers usines) des eaux mauritaniennes. Les recherches effectuées uniquement à bord des navires, ont eu comme objectif de collecter des informations nécessaires à l'évaluation des ressources exploitables.
2. LA ZONE DU LARGE
2.1 Situation de la pêche
Les captures totales roumaines dans la zone au large de la R.I. de Mauritanie ont été de 84 142 tonnes en 1982, ce qui représente une diminution de 1,7% par rapport à l'année 1981, quant on avait réalisé 85 567 tonnes (Tableaux 1 et 4). Entre 1982 et 1981, les captures par unité d'effort ont diminué de 36,0 à 30,0 tonnes/jour ou de 2,9 à 2,3 tonnes par heure de chalutage. Cette tendance est visible à l'intérieur de chacune des deux divisions statistiques du COPACE 34.1.3 et 34.3.1 (Tableaux 2,3 et 5,6). La réduction des captures totales et celle des CPUE ont été particulièrement fortes en 34.1.3, où comme déjà constaté avant on assiste à une diminution continuelle de la biomasse.
L'engin de pêche utilisé dans cette pêcherie a été le chalut pélagique pendant toute la période et pour tous les chalutiers usines.
2.1.1 Les chinchards
Deux espèces de chinchards, Trachurus trachurus et Trachurus trecae ont constitué 42,7% des captures roumaines en 1982. La pêche des chinchards est pratiquée pendant toute l'année, les meilleurs résultats étant obtenus en janvier-février et avril-juin (Tableau 1 et 4).
Tout comme en 1981, en 1982 Trachurus trecae a été l'espèce dominante dans les deux divisions statistiques où ont opéré les chalutiers roumains (Tableaux 2, 3, 5 et 6).
Les rendements moyens journaliers et horaires ont été les plus grands pendant les mois de juin, juillet, août 1982 (Tableau 4) sans que toutefois cette période ait été différente comme c'était le cas en 1981– 82 aux mois de mai-juillet et novembre-janvier.
A part ces deux espèces du genre Trachurus, une autre espèce de Carangidae, Decapterus rhonchus, est capturée en quantité : elle a constitué en 1982 et 1981 respectivement 11,7 et 7,0% des captures (Tableaux 1 et 4). Cette dernière augmentation doit être attribuée à la diminution de l'abondance des Trachurus spp. plutôt qu'une croissance spectaculaire du stock de D. rhonchus. Les meilleurs résultats ont été obtenus en 1982 comme en 1981 dans la division 34.1.3 aux mois de mars et juillet (Tableau 6).
2.1.2 Le maquereau
Le maquereau Scomber japonicus a été en 1982 une espèce secondaire dans les captures roumaines en Mauritanie, contribuant seulement 4 324 tonnes ou 5,1% du total pêché. Les captures ayant été de 3 212 tonnes (Tableau 1) en 1981, on note une augmentation de 34,6% en 1982. Les meilleurs mois ont été mai-juin et septembre-octobre 1982, et juillet 1981. Les CPUE ont évolué dans le même sens : 0,110 t/hr ou 1,35 tonnes/ jour en 1981 contre 0,122 t/hr ou 1,56 t/jr en 1982. Cette augmentation reflète l'amélioration de la situation de stock de maquereaux espagnols.
2.2 Recherches effectuées
2.2.1 Recherches biologiques
2.2.1.1 Trachurus trachurus
Les échantillonnages effectués à bord des navires de pêche
commerciale (4 800 mensurations biométriques et 2 426 déterminations
de l'âge) indiquent que les individus de 19–30 cm (![]() = 25,54 cm)
dominent (Tableau 7), âgés de 3–5 ans (Tableau 8). La génération née
en 1979, âgée de 4 ans en 1982 montrait une nette dominance (30,5%).
= 25,54 cm)
dominent (Tableau 7), âgés de 3–5 ans (Tableau 8). La génération née
en 1979, âgée de 4 ans en 1982 montrait une nette dominance (30,5%).
Le sex ratio a été de 1:1 pour toute la période analysée.
Le rapport taille-poids pour l'année 1982, basé sur les tailles et les poids moyens correspondants (Tableau 7) est :
Y = 0,0128379 X2,9912804
2.2.1.2 Trachurus trecae
En 1982, on a mesuré 3 000 individus et on a déterminé l'âge
de 1 574 exemplaires (tableaux 9 et 10). Les tailles dominantes étaient
comprises entre 22 et 28 cm (![]() = 25,01 cm), l'âge étant de 2–3 ans.
= 25,01 cm), l'âge étant de 2–3 ans.
Le sex ratio était voisin de 1:1, les femelles étant légèrement plus nombreuses (51,7 : 48,3%).
Le rapport taille-poids, calculé à partir des données au Tableau 9 est :
Y = 0,0248344 X2,8090175
2.2.1.3 Decapterus rhonchus
D. rhonchus représente 11,7% des prises, est capturé pendant toute
l'année, avec un maximum en juillet-septembre. Les tailles sont comprises
entre 19 et 40 cm (![]() = 28,25 cm) et l'âge entre 2 et 8 ans (Tableaux
11 et 12).
= 28,25 cm) et l'âge entre 2 et 8 ans (Tableaux
11 et 12).
2.2.1.4 Maquereau espagnol
Chez Scomber japonicus, les tailles ont varié de 14 à 42 cm ;
la plupart étant de 23–26 et 31–37 cm (![]() = 28,61 cm) (Tableau 13). Dans
les captures, les individus nés en 1979 et 1980 représentent plus de
50% du nombre total.
= 28,61 cm) (Tableau 13). Dans
les captures, les individus nés en 1979 et 1980 représentent plus de
50% du nombre total.
3. LA ZONE COTIERE
3.1 Situation de la pêche
La pêche roumaine dans les eaux côtières de la R.I de Mauritanie a eu lieu de mai à octobre 1982, avec des bateaux du type B-140. Deux zones ont été visitées, autour du Cap Timiris et du Cap Blanc. Au Cap Timiris, on a pêché en mai-juin et au Cap Blanc durant tous les mois de mai à octobre (Tableau 15). L'effort de pêche, les captures et les rendements sont présentés aux Tableaux 16–21.
3.2 Composition mensuelle des captures
Les captures ont augmenté régulièrement entre mai-août et diminuent en septembre-octobre, malgré l'augmentation de l'effort de pêche (Tableau 22).
Les prises ont varié en fonction de l'évolution de l'effort de pêche, de la zone de pêche et de leur composition spécifique. Pendant toute la période d'activité, trois principales espèces ont dominé les captures, dans l'ordre suivant : le diagramme gris (Plectorhynchus mediterraneus), le poulpe commun (Octopus vulgaris) et l'émissole lisse (Mustelus mustelus).
Durant les mois de mai à juillet, le diagramme domine nettement au Cap Timiris et au Cap Blanc : 43–57% des captures (Tableau 22). Au mois d'août, son importance diminue : 6% pour augmenter de nouveau à plus de 33% en septembre et à 52% en octobre.
Tout comme le diagramme, le poulpe commun est pêché pendant toute l'année dans les deux secteurs exploités. Son importance relative dans les captures augmente de 2% en mai jusqu'à environ 49% en août.
L'émissole lisse a aussi été abondant au Cap Blanc aux mois d'août et de septembre, représentant respectivement 25 et 17% des captures (Tableau 22).
Le nombre total d'espèces capturées a été de 149, la répartition étant différente dans les deux zones de pêche : au Cap Timiris entre 21 et 48 espèces sont capturées par trait de chalut. Le Tableau 23 montre que le diagramme (56%), les Sparidés (20%), et les mérous (9%) ont été les espèces dominantes. Au Cap Blanc, le nombre d'espéces capturé par trait de chalut est assez réduit : 8–24. Les céphalopodes (poulpe commun + seiche = 34%), le diagramme (29%) et l'émissole lisse (15%) sont dominants. D'autres espèces, également présentes, ont été de moindre importance.
3.3 Recherches effectuées
3.3.1 Recherches biologiques
Les observations effectuées ont mis en évidence l'existence, pendant la période mai-octobre, de saisons et zones de concentration maximale d'un nombre d'espèces : diagramme, mérous, sparidés, poulpe, seiche et émissole. Les autres espèces, dont les plus importantes sont les poissons plats, Decapterus rhonchus, la sardinelle plate, la liche, la courbine, la raie-guitare, les raies, qui apparaissent fréquemment dans les captures, ne forment pas de concentrations suffisamment denses pour être l'objet d'une pêcherie spécialisée. Le reste des espèces capturées, groupées dans la catégorie “autres espéces” sont des espèces secondaires. Les résultats de l'échantillonnage des principales espèces sont présentés par mois dans les tableaux annexés.
3.3.1.1 Diagramme
Plectorhynchus mediterraneus a été l'espèce prédominante aux mois de mai, juin, juillet, septembre, octobre. Au cours de ces trois premiers mois elle était en pleine période de reproduction, formant des concentrations importantes aux profondeurs de 8–23 m au Cap Timiris et 16–22 m au Cap Blanc. Après la reproduction, l'espèce connait une forte dispersion et ne représente que 6% des captures au Cap Blanc en août.
En septembre et surtout en octobre, le diagramme commence à former
de nouvelles concentrations trophiques, à des profondeurs de 18–22 m.
Sa contribution aux captures totales est alors de 50%. Les individus
capturés avaient des tailles comprises entre 17 et 62 cm, les poids
correspondants étant 100–3 730 g. (![]() = 31,1 cm ; p = 557 g.).
= 31,1 cm ; p = 557 g.).
3.3.1.2 Mérous
Les Serranidés ont été représentés par cinq principales espèces : Epinephelus aeneus, E. guaza, E. alexandrinus, E. goreensis et Polyprion americanus, étant relativement importantes au Cap Timiris seulement. Elles y sont capturées tout le long de l'année, avec des pics dans les mois d'été quand E. aeneus, E. alexandrinus et E. guaza se concentrent pour la reproduction dans les eaux froides (10–20°C) sur des fonds sableux, sableux—vaseux et coralliens.
3.3.1.3 Sparidés
Tout comme les mérous, les sparidés constituent un objet de la pêche important au Cap Timiris. Les espèces des genres Sparus (S. caeruleostictus et S. auriga), Spondyliosoma (S. cantharus) et Diplodus (D. vulgaris, D. fasciatus et surtout D. bellottii) dominent nettement dans les captures de Sparidés. Ces espèces, vivant pendant toute l'année aux profondeurs de 8–25 mètres sur des fonds sableux, sableux vaseux, coralliens, vaseux à gravillon et végétation dure abondante sont en début d'été en période de ponte ou de pré-ponte concentrés en bancs relativement grands. Dépendant du moment exact de leur reproduction, la plupart des espèces de sparidés sont plus nombreuses dans les captures pendant la période mai—septembre.
3.3.1.4 Céphalopodes
Les céphalopodes, représentés dans les captures par le poulpe commun (Octopus vulgaris), la seiche (Sepia officinalis) et l'encornet (Loligo vulgaris) ont été pê chés en grandes quantités seulement dans le secteur du Cap Blanc, où 98,5% des captures de céphalopodes ont été réalisées. L'encornet est capturé par individus isolés, la seiche dépasse légèrement 10% et le poulpe représente environ 90% des captures de céphalopodes.
Après une première période de reproduction au mois de mars—début mai, les poulpes font leur apparition au Cap Blanc aux profondeurs de 15–35 m, où ils se nourissent et se reproduisent de nouveau en automne. L'échantillonnage biologique a montré qu'en mi—septembre les femelles sont avancées dans leur maturité sexuelle. Elles disparaissent des captures pendant les semaines suivantes pour aller se réfugier dans les zones à petits fonds qui sont inaccessibles à la pêche par chalutage.
Les plus importantes concentrations du poulpe commun au Cap Blanc ont été observées quand la température de l'eau était de 16 à 18°C, au moment du maximum de la biomasse du zooplancton.
La seiche a été une espèce secondaire pendant toute la période, dans les deux zones de pêche exploitées par les navires B—410. Les mois d'août et de septembre ont été les plus productifs. Sepia se reproduit pendant toute l'année, avec un maximum en mai-août et même septembre, tous les individus au—dessus de la taille de 12 à 13 cm étant matures.
3.3.1.5 Emissole lisse
On a capturé cette espèce aux deux zones de pê che et pendant tous les mois. Pendant les trois derniers mois d'observations, elle a fortement augmenté en abondance à cause de leurs concentrations tropiques sur les petits fonds, surtout ceux de 15–25 m prè s du Cap Blanc.
3.3.1.6 Poissons plats
Les poissons plats, dont un grand nombre d'espèces est présent dans les captures (2–6 espèces par chalutage), représentent 0, 1–0, 7% des prises totales au Cap Timiris et 1,1–2,3% au Cap Blanc. La principale espèce, constituant jusqu'à 99% des captures de poissons plats a été le turbot épineux tâcheté Psettodes belcheri, que l'on rencontre aux profondeurs de 15 à 30 m. Cette espèce pratiquement non—migratrice est capturée en plus grand nombre pendant la saison chaude, surtout en août-septembre, quand elle se concentre aux petites profondeurs pour la ponte.
3.3.1.7 Autres espèces
La courbine (Argyriosomus regius), la liche (Lichia amia), la raie—guitare (Rhinobatos cemiculus et R. rhinobatos) et les autres raies (surtout Raja miraletus et R. picta) ont toujours été des espèces secondaires dans les deux secteurs. Bienqu'elles apparaissent assez fréquement dans les prises, elles constituent rarement plus de 5% dans les captures au chalut.
3.3.2 Evaluation de la densité des poissons démersaux
L'analyse des échogrammes des concentrations des poissons et céphalopodes a montré que seuls le diagramme et Decapterus rhonchus peuvent être distingués par leurs échogrammes. Le diagramme est toujours observé sous forme de petites tâches arrondies, étant dans leurs parties supérieures moins denses que les traces de substrat. Les bancs ainsi observés mesurent 1–3 m de haut et 8–15 m de long.
Decapterus rhonchus par contre, formant des concentrations dans toute la colonne d'eau, est enregistré sous forme de triangles ayant une grande hauteur et une très petite base.
L'occurence d'un grand nombre d'espèces, vivant généralement près du fond, n'a pas permis de reconnaitre d'autres espèces par leurs échogrammes. L'évaluation des biomasses des poissons démersaux a donc été basée sur les densités observées par unité de surface, méthode souvent appliquée lors de l'utilisation du chalut de fond quand la zone de pêche est nouvelle ou peu connue et en absence de statistiques de pêche ou autres données. On a ainsi évalué les quantités de poissons démersaux et de céphalopodes par hectare ; pour l'ensemble des deux zones et pour tous les mois d'activité, la moyenne a été de 19,1 kg/ha, pour le Cap Timiris seul 23kg/ha et pour le Cap Blanc 18 kg/ha. Par mois, les biomasses ont été de 14 kg/ha en mai, 31 kg/ha en juin et juillet, 21 kg/ha en août, 14 kg/ha en septembre et seulement 8 kg/ha en octobre. Ces densités basées sur les captures des navires roumains du type B–410 sont inferieurs à celles calculées par les chercheurs mauritaniens pour la zone Cap Blanc-Saint Louis, évaluées pour l'année 1974 à 40 kg/ha.
Cette différence s'explique par le fait que nos recherches ont été limitées à quelques mois seulement dans une zone plus restreinte. De même, dû aux modifications dans les conditions climatologiques, et hydrologiques et également à l'exploitation intensive des dernières années, les ressources ont constamment diminué depuis 1974 (COPACE/ PACE Séries 78/11).
Nous considéronc donc que les valeurs de biomasses obtenues pour la période mai–octobre reflètent suffisamment bien la réalité pour être utilisées lors de la détermination du potentiel de la zone côtiére.
3.3.3 Recherches hydrologiques, météorologiques et bathymétriques
3.3.3.1 Cap Timiris, 30 mai–21 juin 1982
Pendant cette période, le vent du nord a dominé à 100% avec une vitesse moyenne de 7,95 m/sec. Le degré d'agitation de la mer a été relativement petit (jusqu'au stade III), ce qui a permis de pêcher durant toute la période.
La température de l'air, moyennant 18,4–22°C, étant normale pour cette saison, a pourtant eu des effets défavorables sur la conservation et le traitement des captures, ainsi que sur le bon fonctionnement des installations et appareils des navires B - 140.
La température de l'eau, à la surface a varié de 15,4–20, 5°C. Elle peut influer sur le fonctionnement de l'installation frigorifique.
La nébulosité a été assez réduite en moyenne (1,8 le matin et 1,1 l'après—midi). Elle peut avoir une influence directe sur la pêche. La visibilité, ayant des maxima jusqu'à 7 miles nautiques, n'a pas créé de problèmes pour la navigation et la pêche. Les courants marins de surface prédominants du sud, avec des vitesses jusqu'à 0,6 noeuds, ont un régime relativement constant au cours des mois de mai et juin.
Les fonds ne posent généralement pas de problèmes pour la pêche au chalut, en dehors des zones dangereuses marquées sur la carte.
La position exacte des navires a été difficile à déterminer, la côte étant basse et dépourvue de repères. Etant donné la surface relativement étendue de la zone de pêche, l'orientation des navires B - 140 a été facilitée par la présence du navire collecteur comme point de repère.
3.3.3.2 Cap Blanc, juin – septembre 1982
Le régime des vents est très caractéristique au Cap Blanc. Pour toute la période analysée, les directions nord ont dominé. La proportion des vents dépassant la vitesse de 10m/sec. aété importante; 89% en juin, 68% en juillet, 71% en août et 77% en septembre. L'agitation de la mer a donc souvent dépassé les stades III—IV, quand la pêche est impossible avec les navires B - 140. 63% des jours de pêche ont ainsi été perdus pendant toute la période (66% en juillet et septembre et 58% en août).
La température de l'air a constamment légèrement augmenté jusqu'au mois d'août, suivie d'une faible diminution en septembre. Les moyennes mensuelles (20,0°C en juin, 24,3°C en août et 24,1°C en septembre), et surtout les valeurs de 33°C pendant quelques jours de la période d'observation, ont eu des répercussions négatives sur la durée de conservation des captures et sur le fonctionnement des installations à bord des navires.
La température de l'eau, a été comprise entre 18,93 et 21,72°C sur les zones de pêche, étant plus de 24°C dans la Baie du Lévrier.
La visibilité, généralement réduite, a parfois diminué jusqu'à 0,1 mille nautique, nécessitant un ralentissement des opérations de pêche.
Les courants marins, tout comme les vents dominants, ont eu une direction constante au sud (courant des Canaries) avec des vitesses jusqu'à un noeud. Ces courants, ajoutés au vent, ont déterminé les déviations de la direction du chalutage, surtou quand on a pêché dans les directions autres que nord et sud.
3.3.4 Recherches sur les engins de pêche
Dans la période mai – octobre, on a effectué à bord des navires de pêche côtière des expériences avec quatre type de chaluts de fond de trois constructions différentes et avec quatre variations dans les cables intermédiaires et six dans les cordes du dos. Deux types de panneaux ont également été expérimentés : le panneau universel de 4,5 mp et un panneau de fond du type “V” de 3,5 mp.
On a essayé également des poches de trois constructions différentes et des flotteurs fixes et détachables.
Tous les travaux de modifications des chaluts, de l'armement, de réparations, etc. (comme par exemple, l'imprégnation de latex des deux chaluts et trois poches) ont été effectués sur le navire de base.
Les paramètres résultant des mensurations ou des calculs, par type de chalut, sont présentés au Tableau 24.
3.3.5 Pertes par traitement
Afin de mieux pouvoir valoriser les captures des navires B-140, on a établi les pertes “technologiques” par l'usinage des principales espèces recherchées :
| Espèce | Pertes par usinage (%) |
|---|---|
| Poulpe éviscéré | 15 |
| Diagramme sans tête éviscéré | 45 |
| Sparidés sans tête éviscéré | 50 |
| Mérou éviscéré | 10 |
| Courbine éviscérée | 10 |
| Emissole sans tête éviscéré et sans queue | 55 |
| Raie-guitare sans tête éviscérée | 60 |
| Raies (ailes) | 60 |
| MOIS | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | TOTAL | |
| Effort de pêche | |||||||||||||
| Nombre de navires | 11 | 9 | 8 | 11 | 11 | 12 | 12 | 12 | 12 | 12 | 11 | 10 | 17 |
| Temps de mer (jours) | 274 | 188 | 186 | 263 | 271 | 327 | 342 | 368 | 271 | 369 | 330 | 321 | 3510 |
| Temps de pêche (jours) | 206 | 106 | 156 | 208 | 147 | 246 | 239 | 230 | 172 | 282 | 144 | 239 | 2375 |
| Heures de chalutage | 2258 | 1167 | 1742 | 2606 | 1624 | 2585 | 2784 | 2676 | 1999 | 3358 | 2951 | 3349 | 29099 |
| Tonnes/jour CPUE | 40,3 | 33,2 | 16,3 | 29,9 | 38,9 | 58,1 | 46,4 | 24,3 | 35,9 | 26,8 | 40,1 | 36,1 | 36,0 |
| Tonnes/heure CPUE | 3,68 | 3,02 | 1,46 | 2,38 | 3,52 | 5,53 | 3,98 | 2,09 | 3,08 | 2,25 | 1,96 | 2,58 | 2,94 |
| Captures | |||||||||||||
| TOTAL | 8333 | 3523 | 2591 | 6214 | 5843 | 14299 | 11091 | 5543 | 6166 | 7558 | 5778 | 8628 | 85567 |
| Merluccius senegalensis | - | - | - | - | - | - | 26 | 1 | - | - | 5 | 6 | 38 |
| Dentex macrophthalmus | 273 | 337 | 44 | 108 | 24 | - | - | - | - | 29 | - | 96 | 911 |
| Pagellus erythrinus | 4 | - | 128 | 21 | 30 | - | - | - | 13 | 24 | 3 | 7 | 230 |
| Pagellus spp. | - | - | - | - | - | - | - | - | - | - | 22 | 36 | 58 |
| Spondyliosoma cantharus | 80 | 18 | 107 | 49 | 4 | - | - | - | - | - | - | - | 258 |
| Sparidae | - | - | - | - | - | 2 | 12 | 72 | 39 | 34 | 24 | - | 183 |
| Diplodus spp. | 47 | 4 | 15 | 20 | 2 | - | - | - | - | - | - | - | 88 |
| Brama brama | - | - | - | - | - | - | - | - | 15 | 2 | - | - | 17 |
| Epinephelus spp. | - | - | - | - | - | - | - | 80 | 3 | 57 | - | 1 | 141 |
| Pomatomus saltatrix | - | - | - | - | - | 3 | 71 | 127 | 12 | 64 | 21 | - | 298 |
| Trachurus trachurus | 4778 | 1595 | 423 | 3487 | 1336 | 1313 | 378 | 86 | 138 | 4050 | 208 | 1086 | 18878 |
| Trachurus trecae | 1796 | 1039 | 1481 | 1242 | 4142 | 7760 | 5749 | 1364 | 2000 | 1012 | 5088 | 6530 | 39203 |
| Decapterus rhonchus | - | - | 140 | 423 | - | 2047 | 682 | 853 | 1310 | 563 | - | - | 6018 |
| Sardinella maderensis | - | - | - | - | - | - | - | 98 | - | 2 | 119 | - | 219 |
| Sardinella aurita | 94 | - | 95 | 482 | 10 | 1478 | 1788 | 2191 | 2000 | 479 | - | 36 | 8653 |
| Sarda sarda | - | - | - | - | - | - | 1 | 5 | - | 11 | 1 | 63 | 81 |
| Scomber japonicus | - | - | 1 | 177 | 260 | 317 | 1709 | 42 | 31 | 186 | 150 | 399 | 3212 |
| Trichiuridae | 717 | 433 | 119 | 54 | 5 | 1379 | 675 | 599 | 605 | 996 | 126 | 423 | 6131 |
| Euthynnus alletteratus | - | - | - | - | - | - | - | 9 | - | - | - | 3 | 12 |
| Autres espèces | 544 | 97 | 38 | 151 | 30 | - | - | 16 | - | 49 | 11 | 2 | 938 |
| MOIS | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VIII | XII | TOTAL | |
| Effort de pêche | |||||||||
| Nombre de navires | 11 | 9 | 7 | 10 | 11 | 12 | 12 | 9 | 12 |
| Temps de mer (jours) | 190 | 141 | 100 | 191 | 198 | 260 | 60 | 66 | 1206 |
| Temps de pêche (jours) | 145 | 76 | 82 | 156 | 105 | 208 | 37 | 57 | 866 |
| Heures de chalutage | 1635 | 748 | 931 | 1964 | 1059 | 2204 | 408 | 804 | 9753 |
| Tonnes/jour CPUE | 39,0 | 38,6 | 10,2 | 25,6 | 42,2 | 57,1 | 18,3 | 36,6 | 37,5 |
| Tonnes/heure CPUE | 3,46 | 3,92 | 0,90 | 2,04 | 4,18 | 5,39 | 1,66 | 2,59 | 3,33 |
| Captures | |||||||||
| TOTAL | 5651 | 2939 | 836 | 4001 | 4429 | 11882 | 678 | 2086 | 32502 |
| Merluccius senegalensis | - | - | - | - | - | - | - | 3 | 3 |
| Dentex macrophthalmus | 172 | 337 | 44 | 53 | 24 | - | - | 17 | 647 |
| Pagellus erythrinus | 4 | - | - | 13 | 30 | - | - | 7 | 54 |
| Spondyliosoma cantharus | 80 | 18 | - | 31 | 3 | - | - | - | 132 |
| Sparidae | - | - | - | - | - | 2 | 3 | - | 5 |
| Diplodus spp. | 47 | 4 | - | 13 | - | - | - | - | 64 |
| Pomatomus saltatrix | - | - | - | - | - | 3 | 16 | - | 19 |
| Trachurus trachurus | 3249 | 1330 | 423 | 2197 | 821 | 1069 | - | 40 | 9129 |
| Trachurus trecae | 1147 | 720 | 207 | 530 | 3343 | 5794 | 158 | 1830 | 13729 |
| Decapterus rhonchus | - | - | - | 423 | - | 2047 | 67 | - | 2537 |
| Sardinella maderensis | - | - | - | - | - | - | 98 | - | 98 |
| Sardinella aurita | 94 | - | 95 | 482 | 8 | 1443 | 228 | 4 | 2354 |
| Sarda sarda | - | - | - | - | - | - | - | 9 | 9 |
| Scomber japonicus | - | - | - | 158 | 176 | 296 | 3 | 66 | 699 |
| Trichiuridae | 515 | 433 | 56 | 40 | - | 1228 | 101 | 110 | 2483 |
| Euthynnus alletteratus | - | - | - | - | - | - | 4 | - | 4 |
| Autres espèces | 343 | 97 | 11 | 61 | 24 | - | - | - | 536 |
| MOIS | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | TOTAL | |
| Effort de pêche | |||||||||||||
| Nombre de navires | 9 | 6 | 8 | 10 | 7 | 10 | 12 | 12 | 11 | 12 | 11 | 10 | 17 |
| Temps de mer (jours) | 84 | 47 | 86 | 72 | 73 | 67 | 342 | 308 | 271 | 369 | 330 | 255 | 2304 |
| Temps de pêche (jours) | 61 | 30 | 74 | 52 | 42 | 38 | 239 | 193 | 172 | 282 | 144 | 182 | 1509 |
| Heures de chalutage | 623 | 419 | 811 | 642 | 565 | 381 | 2784 | 2268 | 1999 | 3358 | 2951 | 2545 | 19346 |
| Tonnes/jour CPUE | 44,0 | 19,5 | 23,7 | 42,6 | 33,7 | 63,6 | 46,4 | 25,2 | 35,9 | 26,8 | 40,1 | 36,0 | 35,1 |
| Tonnes/heure CPUE | 4,30 | 1,39 | 2,16 | 3,45 | 2,50 | 6,34 | 3,98 | 2,15 | 3,08 | 2,25 | 1,96 | 2,57 | 2,74 |
| Captures | |||||||||||||
| TOTAL | 2682 | 584 | 1755 | 2213 | 1414 | 2417 | 11091 | 4865 | 6166 | 7558 | 5778 | 6542 | 53065 |
| Merluccius senegalensis | - | - | - | - | - | - | 26 | 1 | - | - | 5 | 3 | 35 |
| Dentex macrophthalmus | 101 | - | - | 55 | - | - | - | - | - | 29 | - | 79 | 264 |
| Pagellus erythrinus | - | - | 128 | 8 | - | - | - | - | 13 | 24 | 3 | - | 176 |
| Pagellus spp. | - | - | - | - | - | - | - | - | - | - | 22 | 36 | 58 |
| Spondyliosoma cantharus | - | - | 107 | 18 | 1 | - | - | - | - | - | - | - | 126 |
| Sparidae | - | - | - | - | - | - | 12 | 69 | 39 | 34 | 24 | - | 178 |
| Diplodus spp. | - | - | 15 | 7 | 2 | - | - | - | - | - | - | - | 24 |
| Brama brama | - | - | - | - | - | - | - | - | 15 | 2 | - | - | 17 |
| Epinephelus spp. | - | - | - | - | - | - | - | 80 | 3 | 57 | - | 1 | 141 |
| Pomatomus saltatrix | - | - | - | - | - | - | 71 | 111 | 12 | 64 | 21 | - | 279 |
| Trachurus trachurus | 1529 | 265 | - | 1290 | 515 | 244 | 378 | 86 | 138 | 4050 | 208 | 1046 | 9749 |
| Trachurus trecae | 649 | 319 | 1274 | 712 | 799 | 1966 | 5749 | 1206 | 2000 | 1012 | 5088 | 4700 | 25474 |
| Decapterus rhonchus | - | - | 140 | - | - | - | 682 | 786 | 1310 | 563 | - | - | 3481 |
| Sardinella maderensis | - | - | - | - | - | - | - | - | - | 2 | 119 | - | 121 |
| Sardinella aurita | - | - | - | - | 2 | 35 | 1788 | 1963 | 2000 | 479 | - | 32 | 6299 |
| Sarda sarda | - | - | - | - | - | - | 1 | 5 | - | 11 | 1 | 54 | 72 |
| Scomber japonicus | - | - | 1 | 19 | 84 | 21 | 1709 | 39 | 31 | 186 | 150 | 273 | 2513 |
| Trichiuridae | 202 | - | 63 | 14 | 5 | 151 | 675 | 498 | 605 | 996 | 126 | 313 | 3648 |
| Euthynnus alletteratus | - | - | - | - | - | - | - | 5 | - | - | - | 3 | 8 |
| Autres espèces | 201 | - | 27 | 90 | 6 | - | - | 16 | - | 49 | 11 | 2 | 402 |
| MOIS | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | TOTAL | |
| Effort de pêche | |||||||||||||
| Nombre de navires | 11 | 11 | 11 | 11 | 11 | 11 | 13 | 11 | 12 | 11 | 10 | 9 | 15 |
| Temps de mer (jours) | 335 | 308 | 277 | 331 | 340 | 328 | 388 | 341 | 347 | 385 | 300 | 290 | 3970 |
| Temps de pêche (jours) | 215 | 230 | 216 | 248 | 249 | 266 | 240 | 209 | 214 | 185 | 253 | 243 | 2768 |
| Heures de chalutage | 2744 | 2988 | 2996 | 3068 | 3815 | 2924 | 2381 | 2732 | 2922 | 2326 | 3139 | 3290 | 35325 |
| Tonnes/jour CPUE | 35,2 | 23,6 | 27,1 | 27,0 | 30,4 | 39,5 | 37,2 | 39,2 | 34,1 | 25,9 | 25,8 | 19,9 | 20,3 |
| Tonnes/heure CPUE | 2,7 | 1,8 | 1,9 | 2,0 | 2,0 | 3,6 | 3,9 | 3,0 | 2,5 | 2,0 | 2,1 | 1,5 | 2,3 |
| Captures | |||||||||||||
| TOTAL | 7567 | 5432 | 5857 | 6692 | 7558 | 10521 | 8716 | 8189 | 7291 | 4953 | 6520 | 4846 | 84142 |
| Merluccius senegalensis | 27 | 7 | 10 | 24 | 113 | 229 | 10 | 44 | - | - | - | 1 | 465 |
| Dentex spp. | - | - | 24 | 87 | 88 | 252 | - | - | - | - | - | - | 451 |
| Pagellus erythrinus | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Autres pageots | 13 | 59 | 129 | 32 | 54 | 13 | 58 | 598 | 292 | 98 | 176 | 94 | 1616 |
| Spondyliosoma cantharus | - | - | - | - | - | - | - | - | - | 38 | - | 38 | |
| Autres Sparides | 51 | 73 | 74 | 70 | - | 54 | 44 | 71 | 168 | 208 | 188 | 141 | 1142 |
| Diplodus spp. | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Brama brama | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Epinephelus spp. | - | - | - | - | - | 2 | 19 | 12 | 33 | 7 | 15 | 3 | 91 |
| Pomatomus saltatrix | - | 1 | 148 | 26 | 6 | 194 | 428 | 80 | 28 | 5 | 28 | 39 | 983 |
| Trachurus trachurus | 4195 | 1532 | 661 | 2396 | 1790 | 2411 | 778 | - | - | - | 31 | 307 | 14101 |
| Trachurus trecae | 1856 | 2174 | 1359 | 1180 | 3161 | 3619 | 951 | 201 | 105 | 1132 | 2546 | 2288 | 20572 |
| Decapterus rhonchus | 133 | 1 | 1365 | 226 | - | 997 | 1888 | 1399 | 1927 | 1243 | 606 | 61 | 9854 |
| Sardinella maderensis | - | - | - | - | - | 530 | 67 | 116 | 83 | 78 | - | - | 874 |
| Sardinella aurita | 53 | 103 | 306 | 744 | 57 | 604 | 2967 | 5070 | 3294 | 623 | 983 | 358 | 15162 |
| Sarda sarda | 114 | 3 | 2 | 1 | 1 | 7 | 77 | 28 | 3 | - | 3 | 10 | 249 |
| Scomber japonicus | 213 | 89 | 137 | 361 | 630 | 587 | 266 | 332 | 662 | 477 | 350 | 220 | 4324 |
| Trichiuridae | 872 | 1190 | 1376 | 919 | 1326 | 401 | 469 | 59 | 584 | 206 | 1487 | 796 | 9685 |
| Euthynnus alletteratus | - | - | - | - | - | 19 | 68 | 22 | 38 | 52 | 34 | 58 | 291 |
| Autres espèces | 40 | 200 | 258 | 626 | 332 | 602 | 626 | 157 | 74 | 786 | 73 | 470 | 4244 |
| MOIS | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | TOTAL | |
| Effort de pêche | |||||||||||||
| Nombre de navires | 10 | 10 | 7 | 11 | 10 | 11 | 13 | 8 | 11 | 11 | 10 | 9 | 15 |
| Temps de mer (jours) | 71 | 136 | 70 | 137 | 228 | 307 | 172 | 71 | 104 | 271 | 115 | 109 | 1791 |
| Temps de pêche (jours) | 48 | 106 | 53 | 106 | 177 | 248 | 109 | 40 | 71 | 118 | 98 | 95 | 1269 |
| Heures de chalutage | 669 | 1433 | 717 | 1390 | 2825 | 2717 | 1021 | 568 | 996 | 1539 | 1335 | 1229 | 16439 |
| Tonnes/jour CPUE | 35,3 | 23,1 | 25,6 | 20,9 | 32,8 | 40,0 | 37,2 | 35,5 | 37,0 | 32,1 | 31,8 | 18,6 | 31,8 |
| Tonnes/heure CPUE | 2,5 | 1,7 | 1,9 | 1,6 | 2,1 | 3,5 | 3,9 | 2,5 | 2,6 | 2,4 | 2,3 | 1,4 | 2,4 |
| Captures | |||||||||||||
| TOTAL | 1696 | 2450 | 1359 | 2216 | 5805 | 9994 | 4088 | 1421 | 2632 | 3948 | 3119 | 1771 | 40499 |
| Merluccius senegalensis | - | - | 10 | - | 103 | 229 | 10 | - | - | - | - | - | 352 |
| Dentex spp. | - | - | 7 | 61 | 83 | 252 | - | - | - | - | - | - | 403 |
| Autres pageots | 2 | 29 | 39 | 15 | 54 | 13 | 25 | 23 | - | - | 41 | 10 | 251 |
| Spondyliosoma cantharus | - | - | - | - | - | - | - | - | - | 26 | - | - | 26 |
| Autres sparidés | - | 33 | - | 2 | - | 54 | 20 | 13 | 52 | 177 | 101 | 72 | 524 |
| Epinephelus spp. | - | - | - | - | - | 2 | 19 | - | 7 | 3 | 3 | - | 34 |
| Pomatomus saltatrix | - | - | 5 | 10 | 2 | 190 | 257 | - | - | 4 | - | 5 | 473 |
| Trachurus trachurus | 1273 | 650 | 397 | 577 | 891 | 2348 | 589 | - | - | - | 31 | - | 6756 |
| Trachurus trecae | 84 | 981 | 691 | 792 | 2928 | 3559 | 139 | - | - | 818 | 885 | 1240 | 12117 |
| Decapterus rhonchus | 34 | - | 25 | 146 | - | 850 | 718 | 389 | 864 | 974 | 606 | - | 4606 |
| Sardinella maderensis | - | - | - | - | - | 530 | 67 | 116 | 83 | 78 | - | - | 874 |
| Sardinella aurita | 15 | 23 | 55 | 161 | 47 | 483 | 1685 | 745 | 839 | 533 | 925 | 35 | 5546 |
| Sarda sarda | 12 | 1 | 2 | - | 1 | 7 | 77 | - | - | - | - | 4 | 104 |
| Scomber japonicus | 18 | 49 | 16 | 24 | 107 | 566 | 79 | 43 | 420 | 405 | 282 | 51 | 2060 |
| Trichiuridae | 249 | 549 | 86 | 375 | 1295 | 394 | 245 | 26 | 76 | 58 | 216 | 89 | 3658 |
| Euthynnus alletteratus | - | - | - | - | - | 15 | - | 6 | 11 | 33 | 7 | 5 | 77 |
| Autres espèces | 9 | 135 | 26 | 53 | 294 | 502 | 158 | 60 | 280 | 839 | 22 | 260 | 2638 |
| MOIS | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | TOTAL | |
| Effort de pêche | |||||||||||||
| Nombre de navires | 11 | 11 | 11 | 10 | 11 | 4 | 12 | 11 | 12 | 9 | 10 | 9 | 15 |
| Temps de mer (jours) | 264 | 172 | 207 | 194 | 112 | 21 | 216 | 270 | 243 | 114 | 185 | 181 | 2179 |
| Temps de pêche (jours) | 167 | 124 | 163 | 142 | 72 | 18 | 131 | 169 | 143 | 67 | 155 | 148 | 1499 |
| Heures de chalutage | 2075 | 1555 | 2279 | 1678 | 990 | 207 | 1360 | 2164 | 1926 | 787 | 1804 | 2061 | 18886 |
| Tonnes/jour CPUE | 35,2 | 24,0 | 27,6 | 31,5 | 24,3 | 30,0 | 35,5 | 40,0 | 32,5 | 15,0 | 22,0 | 20,7 | 29,1 |
| Tonnes/heure CPUE | 2,8 | 1,9 | 1,9 | 2,6 | 1,8 | 2,5 | 3,4 | 3,1 | 2,4 | 1,2 | 1,9 | 1,5 | 2,3 |
| Captures | |||||||||||||
| TOTAL | 5871 | 2982 | 4498 | 4476 | 1753 | 525 | 4655 | 6768 | 4659 | 1005 | 3401 | 3075 | 43668 |
| Merluccius senegalensis | 27 | 7 | - | 24 | 10 | - | - | 44 | - | - | - | 1 | 113 |
| Dentex spp. | - | - | 17 | 26 | 5 | - | - | - | - | - | - | - | 48 |
| Autres pageots | 11 | 30 | 90 | 17 | - | - | 33 | 575 | 24 | 33 | 135 | 84 | 1032 |
| Spondyliosoma cantharus | - | - | - | - | - | - | - | - | - | 12 | - | - | 12 |
| Autres sparodés | 51 | 40 | 74 | 68 | - | - | 24 | 58 | 116 | 31 | 87 | 69 | 618 |
| Epinephelus spp. | - | - | - | - | - | - | - | 12 | 26 | 4 | 12 | 3 | 57 |
| Pomatomus saltatrix | - | 1 | 143 | 16 | 4 | 4 | 171 | 80 | 28 | 1 | 28 | 34 | 510 |
| Trachurus trachurus | 2922 | 882 | 264 | 1819 | 899 | 63 | 189 | - | - | - | - | 307 | 7345 |
| Trachurus trecae | 1772 | 1193 | 668 | 388 | 233 | 60 | 812 | 201 | 105 | 314 | 1661 | 1048 | 8455 |
| Decapterus rhonchus | 99 | 1 | 1348 | 80 | - | 147 | 1170 | 1010 | 1063 | 269 | - | 61 | 5248 |
| Sardinella aurita | 38 | 80 | 251 | 583 | 10 | 121 | 1282 | 4325 | 2455 | 90 | 58 | 323 | 9616 |
| Sarda sarda | 102 | 2 | - | 1 | - | - | - | 28 | 3 | - | 3 | 6 | 145 |
| Scomber japonicus | 195 | 40 | 121 | 337 | 523 | 21 | 187 | 289 | 242 | 72 | 68 | 169 | 2264 |
| Trichiuridae | 623 | 641 | 1290 | 544 | 31 | 7 | 224 | 33 | 508 | 148 | 1271 | 707 | 6027 |
| Euthynnus alletteratus | - | - | - | - | - | 4 | 68 | 16 | 27 | 19 | 27 | 53 | 214 |
| Autres espèces | 31 | 65 | 232 | 573 | 38 | 98 | 495 | 97 | 62 | 12 | 51 | 210 | 1964 |
Fréquence et poids par classe de longueur dans les captures roumaines en 1982, Divisions 34.1.3 et 34.3.1
| Tableau 7 : Trachurus trachurus | Tableau 9 : Trachurus trachurus |
| L (cm) | N | %° |  | L (cm) | N | %° | |
|---|---|---|---|---|---|---|---|
| 10 | 9 | 2 | 12,7 | 14 | 6 | 2 | 38,3 |
| 11 | 12 | 3 | 15,0 | 15 | 30 | 10 | 48,0 |
| 12 | 3 | + | 21,6 | 16 | 67 | 22 | 59,3 |
| 13 | - | - | - | 17 | 141 | 47 | 75,4 |
| 14 | - | - | - | 18 | 189 | 63 | 85,6 |
| 15 | - | - | - | 19 | 65 | 22 | 97,0 |
| 16 | 2 | + | 45,0 | 20 | 44 | 15 | 112,0 |
| 17 | 11 | 2 | 78,6 | 21 | 120 | 40 | 131,2 |
| 18 | 69 | 14 | 80,3 | 22 | 229 | 76 | 147,9 |
| 19 | 209 | 43 | 82,5 | 23 | 315 | 105 | 163,4 |
| 20 | 360 | 76 | 102,0 | 24 | 261 | 88 | 183,0 |
| 21 | 293 | 62 | 121,3 | 25 | 212 | 71 | 224,8 |
| 22 | 369 | 78 | 142,4 | 26 | 207 | 69 | 211,5 |
| 23 | 342 | 71 | 160,5 | 27 | 198 | 66 | 270,1 |
| 24 | 295 | 62 | 180,8 | 28 | 189 | 63 | 300,7 |
| 25 | 225 | 47 | 206,4 | 29 | 139 | 46 | 330,6 |
| 26 | 209 | 43 | 223,6 | 30 | 113 | 38 | 344,8 |
| 27 | 339 | 71 | 250,9 | 31 | 109 | 36 | 405,7 |
| 28 | 547 | 114 | 276,3 | 32 | 141 | 47 | 433,1 |
| 29 | 684 | 142 | 307,4 | 33 | 130 | 43 | 462,8 |
| 30 | 437 | 91 | 331,4 | 34 | 52 | 19 | 494,0 |
| 31 | 148 | 30 | 363,1 | 35 | 26 | 9 | 535,0 |
| 32 | 123 | 26 | 384,0 | 36 | 11 | 4 | 560,0 |
| 33 | 56 | 11 | 431,3 | 37 | 4 | + | 592,5 |
| 34 | 31 | 6 | 465,5 | 38 | 2 | + | 630,0 |
| 35 | 19 | 4 | 520,5 | ||||
| 36 | 8 | 2 | 538,1 | ||||
| TOTAL | 4800 | 1000 | 228,2 | 3000 | 1000 | 233,4 | |
| Age | 1 | 2 | 3 | 4 | 5 | 6 | 7 | TOTAL |
| N | 25 | 396 | 580 | 739 | 579 | 95 | 12 | 2426 |
| %o | 10 | 163 | 239 | 305 | 239 | 39 | 5 | 1000 |
 (cm) (cm) | 11,0 | 19,64 | 22,89 | 27,36 | 30,44 | 32,62 | 34,80 | 25,84 |
| 18,0 | 95,4 | 162,0 | 257,8 | 325,8 | 421,5 | 505,0 | 229,8 |
| Age | 1 | 2 | 3 | 4 | 5 | 6 | TOTAL |
|---|---|---|---|---|---|---|---|
| N | 165 | 518 | 546 | 266 | 78 | 1 | 1574 |
| %o | 105 | 329 | 348 | 169 | 49 | + | 1000 |
| 17,02 | 21,47 | 25,71 | 31,09 | 33,5 | 38,00 | 24,73 | |
| 77,30 | 144,70 | 244,60 | 397,49 | 492,07 | 770,00 | 230,25 |
Fréquences et poids par classe de longueur
dans les captures roumaines en 1982, Divisions 34.1.3 et 34.3.1
| Tableau 11 : Decapterus rhonchus | Tableau 13 : Scomber japonicus |
| L (cm) | N | % | | L(cm) | N | % | |
|---|---|---|---|---|---|---|---|
| 19 | 3 | 1 | 122,7 | 14 | 7 | 3 | 25,7 |
| 20 | 37 | 9 | 128,0 | 15 | 26 | 13 | 31,2 |
| 21 | 100 | 25 | 141,1 | 16 | 70 | 35 | 38,6 |
| 22 | 166 | 41 | 156,3 | 17 | 52 | 26 | 45,6 |
| 23 | 225 | 56 | 178,8 | 18 | 17 | 9 | 54,1 |
| 24 | 248 | 62 | 204,9 | 19 | 4 | 2 | 65,0 |
| 25 | 263 | 66 | 228,3 | 20 | 23 | 12 | 84,8 |
| 26 | 240 | 60 | 254,1 | 21 | 21 | 10 | 96,7 |
| 27 | 236 | 59 | 280,0 | 22 | 23 | 12 | 122,6 |
| 28 | 384 | 96 | 313,7 | 23 | 107 | 53 | 143,1 |
| 29 | 475 | 119 | 356,3 | 24 | 215 | 108 | 156,9 |
| 30 | 470 | 117 | 399,8 | 25 | 173 | 86 | 173,1 |
| 31 | 289 | 72 | 442,3 | 26 | 144 | 72 | 195,1 |
| 32 | 347 | 87 | 498,4 | 27 | 39 | 20 | 220,9 |
| 33 | 270 | 67 | 544,1 | 28 | 29 | 14 | 263,8 |
| 34 | 154 | 38 | 578,7 | 29 | 28 | 14 | 327,9 |
| 35 | 57 | 14 | 616,1 | 30 | 72 | 36 | 370,1 |
| 36 | 23 | 6 | 664,2 | 31 | 128 | 64 | 407,7 |
| 37 | 8 | 2 | 680,0 | 32 | 169 | 85 | 494,6 |
| 38 | 3 | 1 | 736,7 | 33 | 170 | 85 | 498,9 |
| 39 | 1 | 1 | 730,0 | 34 | 121 | 60 | 542,1 |
| 40 | 1 | 1 | 770,0 | 35 | 82 | 41 | 598,2 |
| 36 | 92 | 46 | 680,4 | ||||
| 37 | 93 | 47 | 752,7 | ||||
| 38 | 49 | 24 | 784,1 | ||||
| 39 | 18 | 9 | 874,4 | ||||
| 40 | 20 | 10 | 925,3 | ||||
| 41 | 6 | 3 | 1020,0 | ||||
| 42 | 2 | 1 | 1120,0 | ||||
| TOTAL | 4000 | 1000 | 349,3 | 2000 | 1000 | 362,8 | |
| Age | 2 | 3 | 4 | 5 | 6 | 7 | 8 | TOTAL |
|---|---|---|---|---|---|---|---|---|
| N. | 337 | 252 | 516 | 510 | 349 | 35 | 1 | 2000 |
| %o | 169 | 126 | 258 | 255 | 174 | 17 | 1 | 1000 |
| (cm) | 22,55 | 25,20 | 27,81 | 30,30 | 33,11 | 34,94 | 38,0 | 28,29 |
| 169 | 233 | 313 | 419 | 540 | 648 | 810 | 351,2 |
| Age | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | TOTAL |
|---|---|---|---|---|---|---|---|---|---|
| N. | 174 | 375 | 255 | 152 | 105 | 81 | 46 | 12 | 1200 |
| %o | 145 | 313 | 212 | 128 | 88 | 66 | 38 | 10 | 1000 |
| (cm) | 16,30 | 24,04 | 28,79 | 32,76 | 34,54 | 36,28 | 37,90 | 40,28 | 27,95 |
| 40,8 | 158,8 | 337,1 | 482,3 | 556,0 | 706,8 | 812,9 | 1006,6 | 326,0 |
| Zone | Mai | Juin | Juillet | Août | Septembre | Octobre | TOTAL |
|---|---|---|---|---|---|---|---|
| C.Timiris | 23 | 21 | - | - | - | - | 44 |
| C.Blanc | 9 | 2 | 28 | 81 | 87 | 93 | 300 |
| Zone | Navire | Dates | Jours de pêche | No. de chalutages | Heures de chalut | Heures ch./jour | Prises (kg) | PUE | |
|---|---|---|---|---|---|---|---|---|---|
| kg/h ch. | kg/jour | ||||||||
| Cap | Morunul | 13–31 | 16 | 53 | 101,9 | 6,4 | 19.270 | 189 | 1.204 |
| Timiris | Delfin | 11–19 | 7 | 13 | 30,8 | 4,4 | 1.620 | 53 | 231 |
| TOTAL ZONE | 23 | 66 | 132,7 | 5,8 | 20.890 | 157 | 908 | ||
| Cap | Morunul | 6–10 | 4 | 11 | 18,7 | 4,7 | 880 | 47 | 220 |
| Blanc | Delfin | 6–10 | 5 | 11 | 22,3 | 4,5 | 1.420 | 64 | 284 |
| TOTAL ZONE | 9 | 22 | 41,0 | 4,5 | 2.300 | 56 | 256 | ||
| TOTAL | Morunul | 6–31 | 20 | 64 | 120,6 | 6,0 | 20.150 | 167 | 1.008 |
| MOIS | Delfin | 5–19 | 12 | 24 | 53,1 | 4,4 | 3.040 | 56 | 253 |
| TOTAL MOIS | 32 | 88 | 137,7 | 5,4 | 23.190 | 134 | 725 | ||
| Zone | Dates | Jours de pêche | No. de chalutages | Heures de chalutage | Heures ch/jour | Prises (kg) | PUE | |
|---|---|---|---|---|---|---|---|---|
| kg/h ch. | kg/jour | |||||||
| Cap Timiris | 1–22 | 21 | 62 | 99,8 | 4,8 | 28.190 | 282 | 1.342 |
| Cap Blanc | 27–28 | 2 | 2 | 4,3 | 2,2 | 550 | 127 | 225 |
| TOTAL MOIS | 23 | 64 | 104,2 | 4,5 | 28.740 | 276 | 1.250 | |
| Zone | Navire | Dates | Jours de pêche | No. de chalutages | Heures de chalutage | Heures de ch./jour | Prises (kg) | PUE | |
|---|---|---|---|---|---|---|---|---|---|
| kg/h ch. | kg/jour | ||||||||
| Cap | Morunul | 9–29 | 16 | 39 | 56,3 | 3,5 | 18.190 | 323 | 1.137 |
| Blanc | Delfin | 9–26 | 12 | 26 | 42,4 | 3,5 | 10.310 | 243 | 859 |
| TOTAL MOIS | 28 | 65 | 98,7 | 3,5 | 28.500 | 289 | 1.018 | ||
| Zone | Navire | Date | Jours de pêche | No. de chalutages | Heures de chalutage | Heure de ch./jour | Prises (kg) | PUE | |
|---|---|---|---|---|---|---|---|---|---|
| kg/h ch. | kg/jour | ||||||||
| Morunul | 5–30 | 19 | 43 | 69,5 | 3,7 | 18.140 | 261 | 954 | |
| Cap | Delfin | 5–31 | 20 | 57 | 97,3 | 4,9 | 17.714 | 182 | 886 |
| Blanc | Dorada I | 5–31 | 22 | 78 | 134,8 | 6,1 | 18.103 | 134 | 823 |
| Dorada II | 7–31 | 20 | 64 | 87,3 | 4,4 | 11.702 | 134 | 585 | |
| TOTAL MOIS | 81 | 242 | 388,9 | 4,9 | 65.659 | 169 | 811 | ||
| Zone | Navire | Date | Jours de pêche | No. de chalutages | Heures de chalutage | Heure de ch./jour | Prises (kg) | PUE | |
|---|---|---|---|---|---|---|---|---|---|
| kg/h ch. | kg/jour | ||||||||
| Morunul | 1–26 | 18 | 32 | 71,2 | 3,6 | 6.570 | 92 | 365 | |
| Cap | Delfin | 1–30 | 23 | 65 | 133,5 | 5,8 | 10.185 | 76 | 443 |
| Blanc | Dorada I | 1–30 | 25 | 81 | 151,7 | 6,1 | 21.225 | 140 | 849 |
| Dorada II | 1–30 | 21 | 52 | 92,3 | 4,4 | 19.370 | 210 | 922 | |
| TOTAL MOIS | 87 | 230 | 448,7 | 5,2 | 57.350 | 128 | 659 | ||
| Zone | Navire | Date | Jours de pêche | No. de chalutages | Heures de chalutage | Heure de ch./jour | Prises (kg) | PUE | |
|---|---|---|---|---|---|---|---|---|---|
| kg/h ch. | kg/jour | ||||||||
| Morunul | 3–31 | 24 | 54 | 125 | 5,3 | 4.465 | 36 | 186 | |
| Cap | Delfin | 1–31 | 24 | 71 | 121 | 5,1 | 12.140 | 100 | 506 |
| Blanc | Dorada I | 1–31 | 24 | 70 | 157 | 6,6 | 10.700 | 68 | 446 |
| Dorada II | 1–28 | 21 | 59 | 120 | 5,7 | 12.870 | 107 | 613 | |
| TOTAL MOIS | 93 | 254 | 523 | 5,6 | 40.175 | 77 | 432 | ||
| Groups Commerciaux | Espèces | Mai | Juin | Juillet | Août | September | Octobre | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | kg | % | kg | % | kg | % | kg | % | kg | % | kg | ||
| Céphalopodes | Poulpe | 2,3 | 555 | 2,8 | 820 | 14,3 | 4072 | 48,9 | 32124 | 25,2 | 14385 | 13,7 | 5505 |
| Seiche | 1,4 | 325 | 0,1 | 15 | 1,3 | 371 | 4,4 | 2889 | 4,3 | 2480 | 3,6 | 1455 | |
| Encornet | 0,2 | 45 | 0,2 | 55 | - | - | - | - | - | - | - | - | |
| Diagrammes | 53,1 | 12315 | 43,0 | 12354 | 56,8 | 16800 | 5,9 | 3873 | 33,7 | 19355 | 51,7 | 20770 | |
| Sparides | Dentex | 0,8 | 185 | 0,1 | 20 | - | - | 0,2 | 131 | - | - | - | - |
| Pageot | 0,8 | 190 | 0,5 | 155 | - | - | - | - | - | - | - | - | |
| Pagre | 4,3 | 1000 | 8,3 | 2390 | - | - | - | - | 0,1 | 62 | - | - | |
| Daurade grise | 5,5 | 1230 | 12,1 | 3480 | - | - | - | - | - | - | - | - | |
| Sargue | 1,7 | 395 | 2,1 | 604 | - | - | - | - | - | - | - | - | |
| Poissons plats | 0,1 | 25 | - | - | 0,7 | 195 | 1,1 | 720 | 2,3 | 1330 | 3,9 | 1550 | |
| Mérous | Mérou bronzé | 0,8 | 660 | 4,2 | 1205 | - | - | 0,8 | 525 | 0,8 | 475 | 0,3 | 120 |
| Mérou noir | 0,4 | 95 | 1,4 | 395 | - | - | - | - | - | - | - | - | |
| Badèche | 1,4 | 330 | 4,3 | 1240 | - | - | - | - | - | - | - | - | |
| Cernier | 0,3 | 70 | 0,3 | 80 | - | - | - | - | - | - | - | - | |
| Autres mérous | 0,1 | 35 | 0,2 | 65 | - | - | - | - | - | - | - | - | |
| Autres | D.rhonchus | 11,0 | 2560 | 8,1 | 2330 | 8,4 | 2403 | 4,0 | 2625 | 0,4 | 240 | - | - |
| Sardinelle | - | - | - | - | 3,1 | 900 | - | - | 3,6 | 2070 | - | - | |
| Rouget | 2,0 | 460 | 3,3 | 965 | - | - | - | - | - | - | - | - | |
| Tassergal | 0,1 | 25 | 0,1 | 25 | - | - | 0,1 | 60 | 0,1 | 35 | - | - | |
| Liche | 1,0 | 230 | 0,5 | 145 | 0,4 | 120 | 3,5 | 2295 | 5,5 | 3165 | 9,5 | 3813 | |
| Courbine | 0,1 | 20 | 4,0 | 1170 | 9,5 | 2715 | 1,6 | 1050 | 0,2 | 135 | - | - | |
| Emissole | 2,1 | 493 | 0,4 | 115 | 1,0 | 285 | 24,6 | 16152 | 17,4 | 9955 | 3,3 | 1340 | |
| Raies et guitares | - | - | - | - | 0,7 | 210 | 1,8 | 1180 | 1,9 | 1118 | - | - | |
| Raies | 0,6 | 140 | - | - | 1,2 | 305 | 0,9 | 590 | 0,8 | 450 | - | - | |
| Autres espèces | 7,9 | 1827 | 4,0 | 1112 | 2,6 | 744 | 2,2 | 1445 | 3,7 | 2115 | 14,0 | 5620 | |
| Captures totales | (kg) | 23190 | 28740 | 28500 | 65659 | 57350 | 40175 | ||||||
| Groupes Commerciaux | C. TIMIRIS | C. BLANC | |||||
|---|---|---|---|---|---|---|---|
| % par espèce | % par groupes commerciaux | Quantités pêchées (kg) | % par espèce commerciaux | % par groupes (kg) | Quantités pêchées | ||
| Céphalopodes | Poulpe | 1,0 | 490 | 29,8 | 57.894 | ||
| Seiche | 0,4 | 1,6 | 200 | 3,8 | 33,6 | 7.425 | |
| Encornet | 0,2 | 100 | - | - | |||
| Diagrammes | 56,0 | 56,0 | 27.480 | 28,9 | 28,9 | 56.305 | |
| Dentex | 0,8 | 390 | - | - | |||
| Pageot | 0,7 | 345 | - | - | |||
| Pagre | 6,9 | 19,6 | 3.390 | 0,03 | 0,1 | 62 | |
| Daurade grise | 9,6 | 4.710 | - | - | |||
| Sar | 1,6 | 785 | 0,1 | 195 | |||
| Poissons plats | - | - | - | 2,0 | 2,0 | 3.850 | |
| Mérou bronzé | 3,8 | 1.865 | 0,6 | 1.080 | |||
| Mérou noir | 1,0 | 490 | - | - | |||
| Badèche | 3,2 | 8,5 | 1.570 | - | 0,6 | - | |
| Cernier | 0,3 | 150 | - | - | |||
| Autres mérous | 0,2 | 100 | - | - | |||
| Autres | D.rhonchus | 3,6 | 3,6 | 1.765 | 4,1 | 4,1 | 8.000 |
| Sardinelle | - | - | - | 1,5 | 1,5 | 2.970 | |
| Rouget | 2,9 | 2,9 | 1.425 | - | - | - | |
| Tassergal | - | - | - | 0,1 | 0,1 | 130 | |
| Liche | 1,4 | 1,4 | 685 | 4,7 | 4,7 | 9.210 | |
| Courbine | - | - | - | 2,4 | 2,4 | 4.590 | |
| Emissole | 0,3 | 0,3 | 145 | 14,8 | 14,8 | 28.855 | |
| Raie-guitare | - | - | - | 1,2 | 1,2 | 2.283 | |
| Raies | 0,7 | 0,7 | 345 | 0,6 | 0,6 | 1.130 | |
| Autres espèces | 5,4 | 5,4 | 2.650 | 5,4 | 5,4 | 10.555 | |
| Captures totales | (kg) | 49.080 | 194.534 | ||||
| Type de chalut | Vitesse de chalutage (noeuds) | Ouverture verticale (m) | Ouverture horizontale (m) |
|---|---|---|---|
| 37/50–57 | 2,8–3,0 | 6 | 16–18 |
| 42/48–50 | 3,0–3,4 | 6 | 18–20 |
| 42/48–50 | 3,0 | 8 | 18 |
| 52/58–55 | 3,0–3,2 | 4 | 22 |
| 42/59–57 | 3,0–3,2 | 3 | 22 |
| 52/70–56 | 2,8 | 2 | 26 |
| 47/60–54 | 3,2 | 4 | 24–26 |
LA PECHERIE DE MAQUEREAUX (Scomber japonicus)
DE L'ATLANTIQUE MAROCAIN ENTRE 29°N et 33°N
par H. Belvèze
Institut Scientifique des Pêches Maritimes du Maroc
1. LA FLOTTE DE PECHE
Une flotte de senneurs artisanaux s'est développée au Maroc après la seconde guerre mondiale pour exploiter un abondant stock de sardine à partir des ports de Safi, Essaouira et Agadir. Elle est constituée de navires en bois qui chargent leurs captures en vrac dans des cales non isolées et non réfrigérées. La dégradation rapide du poisson limite l temps de séjour en mer à une douzaine d'heures. Le rayon d'action de ces senneurs est donc faible, et n'excède pas une quarantaine de milles nautiques autour de leur port de débarquement.
Le nombre de bateaux en activité s'élève à un peu plus de 200 unités dont la moitié se trouve à Agadir et l'autre moitié se partage inégalement entre Safi et Essaouira. Si l'évolution en nombre est peu sensible, par contre la taille des bateaux s'accroit de 1973 à 1981 par remplacement progressif des plus petites unités par des plus plus grandes. Les données concernant le nombre et la jauge brute des senneurs sont résumées au Tableau 1.
| Année | SAFI | ESSAOUIRA | AGADIR | TOTAL | ||||
|---|---|---|---|---|---|---|---|---|
| N | J.B.moy. | N | J.B.moy. | N | J.B.moy. | N | J.B.moy. | |
| 1973 | 91 | 36.14 | 19 | 30.81 | 98 | 43.11 | 208 | 38.94 |
| 1974 | 89 | 34.20 | 19 | 30.81 | 96 | 40.53 | 204 | 36.86 |
| 1975 | 83 | 36.63 | 20 | 35.33 | 99 | 46.11 | 202 | 41.15 |
| 1976 | 83 | 37.76 | 25 | 39.19 | 98 | 49.36 | 206 | 43.45 |
| 1977 | 89 | 40.23 | 25 | 40.66 | 101 | 53.74 | 215 | 46.63 |
| 1978 | 90 | 42.40 | 27 | 45.40 | 103 | 56.91 | 220 | 49.56 |
| 1979 | 97 | 44.60 | 30 | 53.39 | 101 | 57.91 | 228 | 51.65 |
| 1980 | 87 | 44.81 | 28 | 49.05 | 105 | 59.82 | 220 | 52.51 |
| 1981 | 85 | 43.58 | 25 | 48.66 | 102 | 59.35 | 212 | 51.76 |
Les bateaux ne sont pas astreints à débarquer leur poisson dans leur port d'attache. Les senneurs de Safi et d'Essaouira exploitent une zone de pêche commune et débarquent leurs prises dans l'un ou l'autre port, en général le plus proche du lieu de capture. A certaines périodes de l'année où le poisson est rare, une partie de la flotte de ces deux ports se rend à Agadir, ou plus récemment même à Tan-Tan, pour y exercer leur activité.
Les senneurs sont en moyenne plus grands à Agadir et plus petits à Safi, mais l'écart entre les tailles extrêmes est important : en 1981 deux bateaux jaugeaient moins de 20 tonneaux et trois plus de 120 tonneaux. Tous les bateaux sont équipés de sondeur vertical et d'un power-block. Le sonar horizontal est en cours d'introduction en 1983.
2. LES CONDITIONS HYDROLOGIQUES DE LA ZONE
Furnestin (1959) a mis en évidence l'existence d'un upwelling se manifestant de Safi au Cap Juby. Son activité est saisonnière : faible ou inexistant en hiver, il se développe au printemps et atteint son maximum à la fin de l'été. Son étude a été approfondi e au cours du programme CINECA par les campagnes du “Jean Charcot”, dont les résultats ont été analysés par Grall et al. (1974), Le Floch (1974), Minas et al. (1982), Le Corre et Tréguer (1976) Thiriot (1976). Huber et al., (1977), Groupe Mediprob (1974), Treguer et Le Corre (1979), Grall et al., (1982). La relation entre le régime des vents alizés et l'upwelling est évidente. Les variations saisonnières de la position de l'anticyclone des Açores engendrent un balancement de la zone d'activité des alizés vers le nord en été et vers le sud en hiver.
La région comprise entre Safi et le Cap Juby se divise en deux zones distinctes (Fig. 1) : la zone nord entre Safi et le Cap Ghir, et la zone sud entre Ifni et le Cap Juby ; des deux zones sont le siège de remontées d'eau profonde originaire de 150 m en hiver et de 250 m en été.
Elles sont séparées par des eaux plus chaudes dans la baie d'Agadir qui sont considérées par Tréguer et Le Corre (1979) comme des eaux de remontée, vieillies et réchauffées.
Dans la zone nord, la source de l'upwelling se situe entre le Cap Sim et le Cap Tafelney. A partir de cette source, la topographie des fonds marins (présence de bancs rocheux a faible profondeur) favorise l'extension vers le sud-ouest d'une langue d'eau froide riche en sels nutritifs dans laquelle se développe progressivement au fur et mesure de sa dérive vers le large, le phytoplancton, puis le zooplancton que Grall et al. (1974) retrouvent encore en abondance à plus de 80 milles de la côte.
Dans la zone sud entre Ifni et le Cap Juby, le plateau continental est plus large et de pente plus faible que dans la zone nord. Les vents y sont plus réguliers mais moins violents qu'entre le Cap Sim et le Cap Tafelney. Les eaux froides de l'upwelling n'apparaissent qu'épisodiquement en surface. Il est fréquent d'observer la présence d'une couche d'eau froide d'origine profonde occupant tout le plateau continental, recouverte en surface par une couche d'eau plus chaude dont l'épaisseur s'accroit de la côte vers le large. Cette couche d'eau froide est le siège d'importants phénomènes de régénération des sels nutritifs ainsi que l'ont montré Le Corre et Tréguer (1976). La production chlorophyllienne est très élevée au niveau du gradient thermique entre les deux couches d'eau, (Grall et al, 1974). Par contre, la couche superficielle chaude dont l'épaisseur n'excéde parfois pas quelques mètres est pauvre en sels nutritifs et en chlorophylle.
La baie d'Agadir qui sépare ces deux zones d'upwelling est également le siége de remontées d'eau profonde mais celles-ci n'atteignent que rarement la surface, occupée par une partie des eaux de l'upwelling de la zone nord qui ont dérivé et se sont réchauffées en vieillissant. Cette caractéristique a une incidence sur la production biologique de la zone, mise en évidence par le rapport ATP/chlorophylle (a) qui permet de définir l'importance du phytoplancton autoplancton autotrophe au sein de la matière vivante. Les Figures 2 et 3, reprises de Grall et al. (1974), montrent les distributions horizontales de la chlorophylle (a) et du rapport ATP/chlorophylle (a), en valeurs inégrées de O à 30 métres. Elles mettent bien en évidence l'extension vers le banc de la conception des eaux de l'upwelling et l'avancée vers la baie d'Agadir d'une langue d'eau moins froide et plus riche en hétérotrophes (zooplancton) qu'en autotrophes (phytoplancton). Cette langue d'eau longe de la bordure du plateau continental de la zone sud où elle est maintenue écartée de la côte par l'upwelling. Cette structure hydrologique particulière est à rapprocher de la répartition des espéces pélagiques montrée à la Figure 1 d'après Chbani (1982). Les sardines occupent les eaux de l'upwelling où la teneur en chlorophylle est élevée tandis que les maquereaux se tiennent plus au large dans des eaux plus chaudes où le rapport ATP/ chlorophyile (a) est plus fort. D'aprés Lamboeuf (comm.pers.) les campagnes de détection acoustique dans les régions d'upwelling du nordouest africain montrent que la distribution du maquereau se situe plus au large que celle de la sardine, dans des eaux de remontée vieillies et réchauffées, oú, d'aprés leur couleur et leur transparence, le développement du zooplancton a considérablement réduit la teneur en phytoplancton. Bien qu'il n'y ait pas réellement de barriére thermique entre les deux espèces, le maquereau se situe en général dans des eaux à plus de 18°C de température. La répartition schématique des trois principales espéces néritiques par rapport à la côte, est représentée à la Figure 4.
3. LES CAPTURES DE MAQUEREAUX
Parmi les petits pélagiques capturés par cette flotte de pêche artisanale, la sardine représente l'espéce largement dominante avec en moyenne 94% des débarquements de 1960 à 1973. Parmi les autres espéces, by-catch de la sardine, le maquereau est la plus importante avec en moyenne 5.6% des débarquements de 1960 à 1973, le chinchard et l'anchois étant négligeables (0.4%). A partir de 1974, les proportions de sardine dans les captures des senneurs diminuent au bénéfice des autres espèces, principalement le maquereau. De 1974 à 1982 les proportions sont en moyenne de 75.7% pour la sardine et de 16.3% pour le maquereau.
Les prises annuelles de maquereaux en tonnes et en pourcentage pour chaque port sont exposées au Tableau 2. Sur l'ensemble de la période 1963/1982, 73% des débarquements ont eu lieu à Agadir et 27% dans les deux autres ports. Mais si on considére les deux périodes : 1963/1973 et 1974/9182, on constate qu'au cours de la premiére, les prises d'Agadir ne représentent que la moitié des prises totales tandis qu'au cours de la seconde elles s'élévent à 81% des prises totales. Il y a eu une augmentation importante des prises de maquereau à partir de 1974 (Fig. 5), provenant essentiellement de la zone de pêche d'Agadir. Il est probable que les particularités hydrologiques de cette zone permettent aux bancs de maquereaux de se concentrer plus prés de la côte et d'être plus accessibles aux senneurs d'Agadir. Les prises annuelles des autres principales espéces capturées par les senneurs d'Agadir, en tonnes et en pourcentage sont exposées au Tableau 3. On constate que les plus fortes captures de maquereau ont lieu les années où la capture de sardine est faible. Ceci est particuliérement net an 1977, 1978 et 1982. Il semble donc qu'il y ait un balancement entre ces deux espéces. Par contre, si le chinchard est également plus abondant dans les captures de 1977 et 1978, il reste à un niveau faible en 1982, Fig. 6).
Cette augmentation des prises de maquereaux à Agadir ne peut s'expliquer ni par un changement de lieu de pêche ni par un changement de cible (prix moins attractif pour cette espéce que pour la sardine). Il est donc vraisemblable qu'elle corresponde à une augmentation de la disponibilité de cette espéce dans le rayon des derniéres années : les vents alizés mesurés à Essaouira, c'est à dire prés de la source de l'upwelling, ont tendance à s'affaiblir depuis deux décennies, mais ont diminué de faç importante entre 1974 et 1976 et sont stabilisés depuis, à un niveau nettement inférieur à la moyenne (Belvéze Erzini et la disponibilité en sardine qui diminue depuis 1974 bien que le stock dans son ensemble soit relativement stable. Il est probable que cet affaiblissement de l'upwelling dans la zone nord, défavorable à la migration des sardines vers les zones de pêche marocaines, puisse en contre partie favoriser un déplacement des bancs de maquereaux, du large vers la côte, hypothése plus réaliste qu'une augmentation réelle du stock de maquereau.
4. LES RENDEMENTS
Ces variations de disponibilité ne permettent pas de considérer les prises par unité d'effort comme des indices d'abondance réelle, mais seulement d'abondance locale dans le rayon d'action des sardiniers.
Les sorties journaliéres des bateaux sont recensées depuis 1968 à Agadir. Nous n'avons pas pu mettre en évidence de corrélation significative entre la jauge brute des senneurs et leur capture moyenne par sortie comme nous l'avons fait pour la sardine, (Belvèze, 1981). Nous nous bornerons donc à analyser le nombre de jourse de pêche des sardiniers et les rendements, c'est à dire la capture moyenne par sortie (cf. Tableau 4).
Les sardiniers ont eu une péridoe d'activité élevée entre 1971 et 1974. Ensuite, le nombre annuel de sorties a diminué jusqu'en 1979, et après avoir réaugmenté en 1980 et 1981, il chute à son niveau le plus bas en 1982. La diminution de l'activité aprés 1974 est liée au renchérissement du prix du carburant et à la diminution de la disponibilité de la sardine qui reste l'espéce cible : certain armateurs ayant des difficultés de trésorerie, ne font sortir leurs bateaux que lorsque la présence de sardine est attestée par les captures des autres bateaux qui continuent à sortir en prenant le risque de rentrer à vide ou avec une espéce moins rénumératrice. En 1982, la baisse de l'activité des sardiniers a également pour origine le transfert d'une partie de la flotte au nouveau port de Tan-Tan pendant certains mois de l'année où la sardine était rare dans la baie d'Agadir.
Il n'y a apparament pas de lien entre l'activité des bateaux et les rendements en maquereau. Malgré des fluctuations inter-annuelles assez prononcées, les rendements par jour de pêche ont tendance à augmenter à partir de 1974 et atteignent leur maximum en 1982 (Fig. 7). L'augmentation des captures n'est donc pas liée à une augmentation de l'effort de pê ou à un chagement de straégie ou de cible, mais bien à une meilleure disponibilité des maquereaux dans la baie d'Agadir.
Les répartitions moyennes des captures et des rendements mensuels de 1974 à 1982 évoluent de façon sensiblement parallèle, (Fig. 8). Les données sont exposées au Tableau 5. Il est intéressant de constater que les deux maxima (juin et août) s’intercalent avec les deux maxima de prise et de capture par unité d’effort de la sardine qui se situent en juillet et en octobre, ce qui confirme l’antagonisme entre les disponibilités de l'autre espèce.
5. DISTRIBUTION DE FREQUENCE DE TAILLE DANS LES CAPTURES
L'échantillonnage des captures de sardine, commencé en 1970 à Safi et en 1971 à Agadir, a fait l'objet d'une extension systématique à toute la flotte à partir de 1973. Dans chaque port, un dixième des bateaux, désigné par tirage au sort est échantillonné tous jours. Primitivement destiné à l'étude du stock sardinier, ce plan d'échantillonnage au hazard a été étendu aux bateaux pêchant du maquereau en 1975, en raison de la part grandissante de cette espèce dans les débarquements.
L'échantillon est prélevé sur une caisse au hazard au cours du déchargement. Il est pesé et tous les poissons sont mesurés à la fourche au demi-centimètre près, avec la règle standard utilisée pour la sardine. La distribution de fréquence de taille est extrapolée à la capture du bateau échantillonnée, puis additionnée aux autres distributions obtenues au cours du mois. Cette somme est ensuite extrapolée à la capture totale mensuelle de maquereau. La distribution de fréquence de taille est alors regroupée en classes de un centimètre pour limiter le nombre de classes (la classe “200 cm” s'étend de 20 à 20.9 cm et son point médian est 20.5 cm).
Ces captures par classe de taille mensuelles peuvent être ensuite regroupées sur une base trimestrielle ou annuelle, ou additionnées port à port. Nous présentons aux Tableaux 6 et 7 les captures annuelles de maquereau, distribuées par classes de taille, de 1975 à 1982, pour Agadir d'une part, et pour l'ensemble Safi-Essaouira d'autre part, et au Tableau 8 les captures regroupées pour l'ensemble des trois ports. Les poids moyens et les tailles moyennes des maquereaux pêchés chaque année figurent au bas de ces Tableaux. Elles montrent (Fig. 9) un rajeunissement rapide des poissons dans les captures de 1975 à 1977, suvi d'un vieillissement progressif à Safi-Essaouira et d’une stabilisation autour de 20 cm de longueur à Agadir.
6. RELATION TAILLE-POIDS
La relation taille-poids a été calculée par régression fonctionnelle au sens de Ricker (1973), puisque les valeurs de X (longueur L) ne sont pas fixées mais sujettes à la variabilité naturelle, les deux variables étant réciproquement fonction l'une de l'autre.
La relation classique W = K.La transformée en valeurs logarithmiques est linéaire : log W = log K + a.log L. sa pente a est égale au rapport de l'écart-type de log W sur l'écart-type de log L.
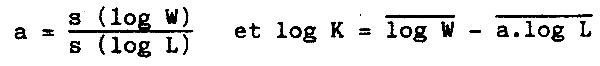
Nous avons utilisé les mesures individuelles de 322 individus dont les taille sont échelonnées entre 9 et 39 cm. Le coefficient de corrélation r = 0.99 est hautement significatif, et la relation taillepoids obtenue s’écrit:
W = 0,00407 L3.30
La représentation graphique (Fig. 10) montre la répartition des poids moyens de chaque classe de taille de un centimètre par rapport à la courbe obtenue.
7. REFERENCES DES AUTEURS CITES
Belvèze, H., 1981 Note sur l'état de la pêcherie sardiniére marocaine dans l'Atlantique en 1980. Trav. Dov. Dév. Pêche Maroc No. 31.
Belvèze, H., 1983 Erzini, K., Influence des facteurs hydroclimatiques sur la disponibilité en sardine (Sardina pilchardus) dans la pêcherie marocaine atlantique. Papier présenté à la Consultation Technique sur les changements d’abondance et de composition spécifique des stocks néritiques, San José, Costa—Rica, 19 avril 1983.
Chbani, M., 1982 Campagne de prospection acoustique de Casablanca au Cap Barbas Octobre 1982. Résultats preéliminaires. Doc. Institut. Scientifique des Pêches Maritimes du Maroc (miméo).
Furnestin, J., 1959 L'hydrologie du Maroc Atlantique de 1949 à 1953. Rev. Trav. Inst. Scient. Tech. Pêch. mar. 23 (I).
Grall, J.R., 1974 Laborde P., Le Corre P., Neveux J., Tréguer, P., Thiriot A., Caractéristiques trophiques et production planctonique dans la région sud de l'atlantique marocain. Téthys 6 (I–2) : II–28.
Grall, J.R., 1982 Le Corre P., et Tréquert P., Short term variability of primary production in coastal upwelling off Morocco. Rapp. P.-V. Réun. Cons. int. Explor. Mer, 180: 221–227.
Groupe MEDIPROD 1974, Généralités sur la campagne CINECA—CHARCOT II Théthys 6 (I–2) : 33–42.
Hugues, P., 1974 and Barton E.D., Physical investigations in upwelling region of north west Africa on RRS Discovery Cruise 48. Téthys 6 (I–2):
Le Corre, P. et Tréguer, P., 1976 Contribution à l'étude des sels nutritifs et de la matière organique dissoute dans l'eau de mer. Caractéristiques chimiques du Golfe de Gascogne et des upwelling côtiers de l'Afrique du nord-ouest. Theèse Doct.Etat, Univ.Brest, 490pp.
Le Floch, J., 1974 Quelques aspects de la dynamique et de l'hydrologie des couches superficielles dans l'ouest marocain. Campagnes CINECACHARCOT I et III. Téthys 6 (I–2) : 53–68.
Minas, H.J., Codispoti, L.A. and Dugdale, R.C., 1982 Nutrients and primary production in the upwelling region off northwest Africa. Rapp. P.-V. Réun.Cons.int.Explor.Mer, 180 : 148.
Ricker, W.E., 1973 Linear regressions in fishery research. J. Fish.Res.Board Can. 30 : 409–434.
Thiriot, A., 1976 Les remontées d'eau (upwelling) et leur influence sur la production pélagique des côtes atlantiques du Maroc. Bull.Inst. Pêche.Marit.Marc 22 : 5–12.
Tréguer, P. et Le Corre, P., 1979 The ratios of nitrate, phosphates and silicates during uptake and regeneration phases of the moroccan upwelling regime. Deep-Sea Res. 26 : 163–184
| SAFI | ESSAOUIRA | AGADIR | TOTAL | ||||
|---|---|---|---|---|---|---|---|
| Tonnes | % | Tonnes | % | Tonnes | % | Tonnes | |
| 1963 | 6 530 | 52.0 | 17 | 0.1 | 6 007 | 47.9 | 12 554 |
| 1964 | 312 | 5.1 | 90 | 1.5 | 5 673 | 93.4 | 6 075 |
| 1965 | 4 714 | 52.4 | 49 | 0.5 | 4 243 | 47.1 | 9 006 |
| 1966 | 1 951 | 37.7 | - | 0.0 | 3 225 | 62.3 | 5 176 |
| 1967 | 2 783 | 36.1 | 7 | 0.1 | 4 914 | 63.8 | 7 704 |
| 1968 | 1 450 | 28.0 | - | 0.0 | 3 732 | 72.0 | 5 182 |
| 1969 | 2 746 | 33.5 | - | 0.0 | 5 444 | 66.5 | 8 190 |
| 1970 | 15 378 | 58.2 | 3 719 | 14.1 | 7 330 | 27.7 | 26 427 |
| 1971 | 2 450 | 32.9 | 810 | 10.9 | 4 170 | 56.2 | 7 430 |
| 1972 | 981 | 22.9 | 365 | 8.5 | 2 942 | 68.6 | 4 288 |
| 1973 | 2 457 | 29.7 | 2 677 | 32.3 | 3 146 | 38.0 | 8 280 |
| 1974 | 2 788 | 13.1 | 1 237 | 5.8 | 17 197 | 81.0 | 21 222 |
| 1975 | 60 | 0.5 | 488 | 4.0 | 11 731 | 95.5 | 12 279 |
| 1976 | 6 823 | 26.1 | 647 | 2.5 | 18 691 | 71.4 | 26 161 |
| 1977 | 4 291 | 9.0 | 2 008 | 4.2 | 41 274 | 86.8 | 47 573 |
| 1978 | 8 421 | 17.7 | 1 520 | 3.2 | 37 669 | 79.1 | 47 610 |
| 1979 | 4 117 | 26.3 | 2 878 | 18.3 | 8 681 | 55.4 | 15 676 |
| 1980 | 3 937 | 12.5 | 4 225 | 13.4 | 23 413 | 74.1 | 31 575 |
| 1981 | 2 730 | 9.0 | 1 212 | 4.0 | 26 420 | 87.0 | 30 362 |
| 1982 | 2 852 | 5.2 | 2 277 | 4.1 | 50 056 | 90.7 | 55 207 |
| SARDINE | MAQUEREAU | CHINCHARD | ANCHOIS | TOTAL | |||||
|---|---|---|---|---|---|---|---|---|---|
| T | % | T | % | T | % | T | % | T | |
| 1963 | 42 684 | 87.4 | 6 007 | 12.3 | 140 | 0.3 | 1 | 0.0 | 48 832 |
| 1964 | 56 706 | 90.6 | 5 673 | 9.1 | 176 | 0.3 | - | - | 62 555 |
| 1965 | 64 237 | 93.2 | 4 243 | 6.2 | 305 | 0.4 | 164 | 0.2 | 68 949 |
| 1966 | 114 649 | 97.0 | 3 225 | 2.7 | 224 | 0.2 | 82 | 0.1 | 118 180 |
| 1967 | 72 150 | 93.2 | 4 914 | 6.4 | 314 | 0.4 | 4 | 0.0 | 77 382 |
| 1968 | 72 324 | 93.8 | 3 732 | 4.8 | 810 | 1.1 | 209 | 0.3 | 77 075 |
| 1969 | 77 878 | 92.6 | 5 444 | 6.5 | 781 | 0.9 | 30 | 0.0 | 84 133 |
| 1970 | 72 763 | 86.8 | 7 330 | 8.7 | 1 250 | 1.5 | 2 438 | 2.9 | 83 781 |
| 1971 | 119 470 | 95.0 | 4 170 | 5.7 | 636 | 0.5 | 1 475 | 1.2 | 125 751 |
| 1972 | 103 783 | 95.0 | 2 942 | 2.7 | 1 264 | 1.2 | 1 242 | 1.1 | 109 231 |
| 1973 | 217 502 | 97.8 | 3 146 | 1.4 | 1 522 | 0.7 | 335 | 0.2 | 222 505 |
| 1974 | 140 023 | 87.9 | 17 197 | 10.8 | - | - | 2 110 | 1.3 | 159 330 |
| 1975 | 90 361 | 87.2 | 11 731 | 11.3 | 1 312 | 1.3 | 194 | 0.2 | 103 598 |
| 1976 | 140 144 | 83.2 | 18 691 | 11.1 | 9 370 | 5.6 | 193 | 0.1 | 168 398 |
| 1977 | 57 010 | 43.1 | 41 274 | 31.2 | 29 945 | 22.7 | 3 924 | 3.0 | 132 153 |
| 1978 | 59 944 | 41.4 | 37 669 | 26.0 | 46 653 | 32.2 | 628 | 0.4 | 144 894 |
| 1979 | 98 017 | 85.0 | 8 681 | 7.5 | 7 713 | 6.7 | 903 | 0.8 | 115 314 |
| 1980 | 128 021 | 80.8 | 23 413 | 14.8 | 6 649 | 4.2 | 342 | 0.2 | 158 425 |
| 1981 | 137 190 | 80.7 | 26 420 | 15.5 | 5 897 | 3.5 | 417 | 0.2 | 169 924 |
| 1982 | 65 699 | 54.4 | 50 056 | 41.4 | 3 856 | 3.2 | 1 184 | 1.0 | 120 795 |
| Année | Capture de maquereaux (tonnes) | Nombre de jours de mer | Capture moyenne/jour de mer (kg) |
|---|---|---|---|
| 1968 | 3 732 | 17 310 | 216 |
| 1969 | 5 444 | 16 056 | 339 |
| 1970 | 7 330 | 19 812 | 370 |
| 1971 | 4 170 | 25 338 | 165 |
| 1972 | 2 942 | 25 069 | 117 |
| 1973 | 3 146 | 25 387 | 124 |
| 1974 | 17 197 | 22 254 | 773 |
| 1975 | 11 731 | 18 717 | 627 |
| 1976 | 18 691 | 21 514 | 869 |
| 1977 | 41 274 | 17 385 | 2 374 |
| 1978 | 37 669 | 17 374 | 2 168 |
| 1979 | 8 681 | 16 955 | 512 |
| 1980 | 23 413 | 20 344 | 1 151 |
| 1981 | 26 420 | 20 280 | 1 303 |
| 1982 | 50 056 | 15 321 | 3 267 |
| MOIS | DEBARQUEMENTS (T) | PRISES PAR JOUR (kg) |
|---|---|---|
| Janvier | 134 | 148 |
| Février | 238 | 87 |
| Mars | 201 | 145 |
| Avril | 872 | 466 |
| Mai | 3 279 | 1 786 |
| Juin | 4 582 | 2 065 |
| Juillet | 3 550 | 1 686 |
| Août | 5 588 | 2 970 |
| Septembre | 2 683 | 1 627 |
| Octobre | 2 174 | 1 380 |
| Novembre | 1 692 | 1 482 |
| Décembre | 1 254 | 1 332 |
| cm | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 | 1982 |
|---|---|---|---|---|---|---|---|---|
| 9 | 39 | |||||||
| 10 | 491 | 106 | 2 420 | |||||
| 11 | 8 749 | 1 215 | 133 | 14 110 | ||||
| 12 | 24 942 | 6 795 | 773 | 27 503 | 168 | |||
| 13 | 4 | 98 519 | 21 649 | 1 411 | 17 752 | 1 696 | 4 175 | |
| 14 | 1 058 | 143 013 | 24 006 | 1 097 | 10 679 | 20 525 | 11 056 | |
| 15 | 862 | 107 432 | 11 560 | 5 741 | 8 903 | 35 601 | 29 003 | |
| 16 | 16 | 883 | 82 363 | 7 336 | 7 103 | 6 439 | 28 759 | 26 856 |
| 17 | 36 | 3 460 | 52 481 | 5 471 | 6 671 | 7 367 | 15 406 | 34 374 |
| 18 | 62 | 6 189 | 34 550 | 8 932 | 7 445 | 13 519 | 15 172 | 49 177 |
| 19 | 118 | 7 901 | 12 149 | 16 960 | 7 253 | 17 617 | 12 758 | 45 942 |
| 20 | 331 | 9 281 | 3 178 | 26 058 | 10 909 | 18 069 | 15 169 | 52 729 |
| 21 | 373 | 8 340 | 1 039 | 32 007 | 7 509 | 19 657 | 16 500 | 57 218 |
| 22 | 519 | 5 860 | 3 242 | 31 852 | 6 585 | 19 380 | 18 886 | 40 105 |
| 23 | 727 | 5 396 | 7 457 | 28 921 | 5 720 | 16 322 | 18 729 | 29 352 |
| 24 | 1 537 | 5 401 | 14 407 | 28 744 | 3 918 | 14 539 | 15 510 | 20 748 |
| 25 | 2 230 | 5 344 | 14 984 | 20 499 | 2 118 | 10 473 | 11 660 | 14 832 |
| 26 | 3 644 | 5 018 | 10 475 | 12 577 | 1 181 | 8 236 | 6 254 | 10 847 |
| 27 | 5 619 | 5 722 | 6 614 | 8 480 | 846 | 4 590 | 3 345 | 8 452 |
| 28 | 5 445 | 5 666 | 3 528 | 5 596 | 635 | 1 953 | 1 581 | 6 179 |
| 29 | 4 193 | 4 829 | 3 073 | 3 442 | 299 | 1 302 | 730 | 3 939 |
| 30 | 2 567 | 2 947 | 1 534 | 1 190 | 468 | 459 | 249 | 3 139 |
| 31 | 2 040 | 7 688 | 683 | 1 110 | 204 | 286 | 197 | 1 838 |
| 32 | 997 | 1 278 | 384 | 122 | 111 | 162 | 46 | 1 447 |
| 33 | 630 | 600 | 518 | 83 | 60 | 20 | 63 | 1 212 |
| 34 | 234 | 503 | 84 | 32 | 47 | 9 | 630 | |
| 35 | 100 | 191 | 3 | 13 | 48 | 2 | 426 | |
| 36 | 46 | 84 | 18 | 4 | 14 | 43 | 5 | |
| 37 | 24 | 107 | 15 | 1 | 36 | |||
| 38 | 32 | 52 | 3 | |||||
| 39 | 18 | 29 | 3 | |||||
| 40 | 7 | 14 | ||||||
| 41 | 5 | 57 | 3 | |||||
| 42 | 2 | |||||||
| 43 | ||||||||
| N | 31 552 | 94 764 | 635 925 | 304 763 | 78 306 | 241 850 | 238 836 | 453 885 |
| C | 11 731T | 18 691T | 41 274T | 37 669T | 8 681T | 23 413T | 26 400T | 50 056T |
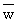 | 372g | 197g | 65g | 124g | 111g | 97g | 111g | 110g |
| 28.1cm | 24.2cm | 16.5cm | 20.9cm | 20.1cm | 19.0cm | 19.7cm | 20.7cm |
| cm | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 | 1982 |
|---|---|---|---|---|---|---|---|---|
| 9 | 3 024 | |||||||
| 10 | 19 012 | |||||||
| 11 | 25 437 | 161 | ||||||
| 12 | 34 080 | 3 777 | 550 | 1 843 | ||||
| 13 | 25 077 | 21 861 | 6 057 | 8 676 | ||||
| 14 | 10 173 | 70 142 | 23 464 | 13 289 | ||||
| 15 | 3 817 | 77 888 | 28 464 | 35 101 | 71 | |||
| 16 | 1 419 | 2 029 | 34 229 | 12 775 | 42 162 | 1 055 | ||
| 17 | 7 646 | 14 550 | 14 262 | 16 081 | 1 070 | 1 635 | ||
| 18 | 16 622 | 203 | 7 268 | 7 287 | 7 250 | 5 089 | 1 193 | |
| 19 | 15 068 | 203 | 2 850 | 5 428 | 2 973 | 2 551 | ||
| 20 | 9 325 | 128 | 1 607 | 2 661 | 2 558 | 963 | 3 650 | |
| 21 | 25 | 8 465 | 23 | 622 | 2 464 | 603 | 1 208 | 4 918 |
| 22 | 79 | 1 270 | 82 | 469 | 4 710 | 1 022 | 2 191 | 3 610 |
| 23 | 15 | 622 | 603 | 580 | 2 861 | 3 077 | 2 911 | 4 068 |
| 24 | 56 | 673 | 1 230 | 160 | 746 | 3 539 | 2 794 | 5 928 |
| 25 | 48 | 898 | 1 732 | 109 | 588 | 6 056 | 3 350 | 4 506 |
| 26 | 69 | 1 126 | 2 023 | 66 | 494 | 2 790 | 2 570 | 2 454 |
| 27 | 68 | 1 186 | 2 355 | 6 | 517 | 2 689 | 1 582 | 1 873 |
| 28 | 170 | 877 | 1 715 | 23 | 200 | 95 | 1 185 | 562 |
| 29 | 355 | 621 | 1 920 | 469 | 801 | 488 | ||
| 30 | 286 | 462 | 963 | 197 | 279 | 54 | ||
| 31 | 252 | 388 | 693 | 124 | 198 | |||
| 32 | 185 | 145 | 410 | 27 | 39 | 100 | ||
| 33 | 125 | 78 | 304 | 19 | ||||
| 34 | 20 | 48 | 189 | 39 | ||||
| 35 | 4 | 31 | 81 | |||||
| 36 | 8 | 2 | 70 | |||||
| 37 | 31 | |||||||
| 38 | 11 | |||||||
| 39 | ||||||||
| 40 | ||||||||
| 41 | ||||||||
| 42 | ||||||||
| N | 1 765 | 67 023 | 137 607 | 236 368 | 114 345 | 146 831 | 29 261 | 38 716 |
| C | 548T | 7 470T | 6 299T | 9941T | 6 995T | 8 162T | 3 942T | 5 129T |
| 310g | 111g | 46g | 42g | 61g | 56g | 135g | 132g |
| 29.6cm | 20.3cm | 14.1cm | 15.5cm | 17.0cm | 17.3cm | 23.1cm | 22.9cm |
| cm | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 | 1982 |
|---|---|---|---|---|---|---|---|---|
| 9 | 3 024 | 39 | ||||||
| 10 | 19 503 | 106 | 2 420 | |||||
| 11 | 34 186 | 1 376 | 133 | 14 110 | ||||
| 12 | 59 022 | 10 572 | 1 323 | 29 346 | 168 | |||
| 13 | 4 | 123 596 | 43 510 | 7 468 | 26 428 | 1 696 | 4 175 | |
| 14 | 1 058 | 153 186 | 94 148 | 24 561 | 23 968 | 20 525 | 11 056 | |
| 15 | 862 | 111 249 | 89 448 | 34 205 | 44 004 | 35 601 | 29 074 | |
| 16 | 16 | 2 302 | 84 392 | 41 565 | 19 878 | 48 601 | 28 759 | 27 911 |
| 17 | 36 | 11 106 | 52 481 | 20 021 | 20 933 | 23 448 | 16 476 | 36 009 |
| 18 | 62 | 22 811 | 34 753 | 16 200 | 14 732 | 20 769 | 20 261 | 50 370 |
| 19 | 118 | 22 969 | 12 352 | 19 810 | 12 681 | 17 617 | 15 731 | 48 493 |
| 20 | 331 | 18 606 | 3 306 | 27 665 | 13 570 | 20 627 | 16 132 | 56 379 |
| 21 | 398 | 16 805 | 1 062 | 32 629 | 9 973 | 20 260 | 17 708 | 62 136 |
| 22 | 598 | 7 130 | 3 324 | 32 321 | 11 295 | 20 402 | 21 077 | 43 715 |
| 23 | 742 | 6 058 | 8 060 | 29 501 | 8 581 | 19 399 | 21 640 | 33 420 |
| 24 | 1 593 | 6 074 | 15 637 | 28 904 | 4 664 | 18 078 | 18 304 | 26 676 |
| 25 | 2 278 | 6 242 | 16 716 | 20 608 | 2 706 | 16 529 | 15 010 | 19 338 |
| 26 | 3 713 | 6 144 | 12 498 | 12 643 | 1 675 | 11 026 | 8 824 | 13 301 |
| 27 | 5 687 | 6 908 | 8 969 | 8 486 | 1 363 | 7 279 | 4 927 | 10 325 |
| 28 | 5 615 | 6 543 | 5 243 | 5 619 | 835 | 2 048 | 2 766 | 6 741 |
| 29 | 4 548 | 5 450 | 4 993 | 3 442 | 768 | 1 302 | 1 531 | 4 427 |
| 30 | 2 853 | 3 409 | 2 497 | 1 190 | 665 | 459 | 528 | 3 193 |
| 31 | 2 292 | 8 076 | 1 376 | 1 110 | 328 | 286 | 395 | 1 838 |
| 32 | 1 182 | 1 423 | 794 | 122 | 138 | 162 | 85 | 1 547 |
| 33 | 755 | 678 | 822 | 83 | 60 | 20 | 82 | 1 212 |
| 34 | 254 | 551 | 273 | 32 | 47 | 9 | 39 | 630 |
| 35 | 104 | 222 | 84 | 13 | 48 | 2 | 426 | |
| 36 | 54 | 86 | 88 | 4 | 14 | 43 | 5 | |
| 37 | 24 | 107 | 46 | 1 | 36 | |||
| 38 | 32 | 63 | 3 | |||||
| 39 | 18 | 29 | 3 | |||||
| 40 | 7 | 14 | ||||||
| 41 | 5 | 57 | 3 | |||||
| 42 | 2 | |||||||
| N | 33 317 | 161 787 | 773 532 | 541 131 | 192 651 | 388 681 | 268 097 | 492 601 |
| C | 12 279T | 26 161T | 47 573T | 47 610T | 15 676T | 31 575T | 30 362T | 55 207T |
| 369g | 162g | 62g | 88g | 81g | 81g | 113g | 112g |
| 28.1cm | 22.6cm | 16.0cm | 18.6cm | 18.3cm | 18.4cm | 20.0cm | 20.9cm |
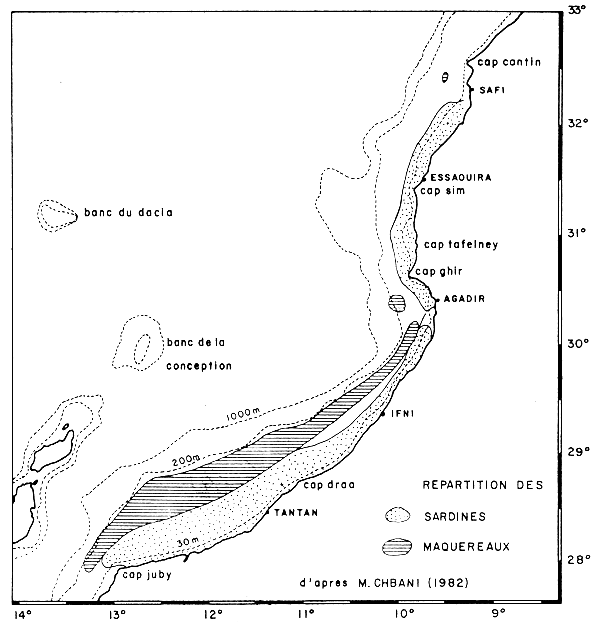
Fig. I : Répartition géographique des sardines (Sardina pilchardus) et des maqueraux (Scomber japonicus) dans la pêcherue marocaine en Octobre 1982. (d'après Chbani, 1982)
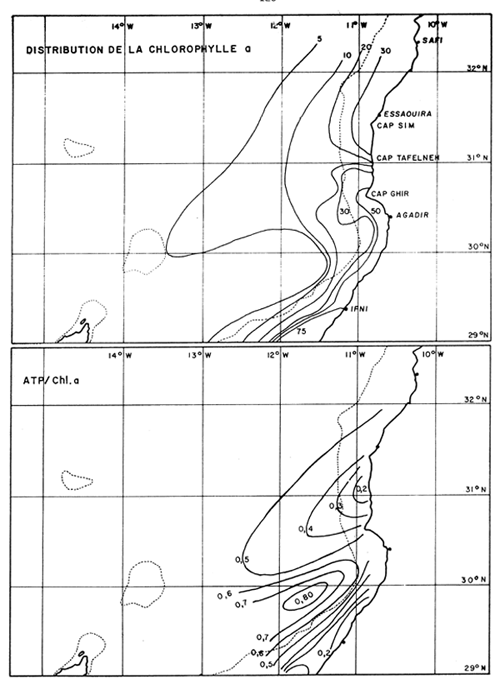
Fig. 2 et 3 : distribution de la chlorophylle (a) et du rapport ATP/chlorophylle (a) caractèrisant les eaux de l'upwelling dans la zone de pêche marocaine. (d'après Grall et al., 1974)
Fig. 4 : Répartition schèmatique des sardines, maquereuux et chinchards dans les eaux de remontée du Nord-Ouest Africain. (D'après Lamboeuf, comm. pers.)
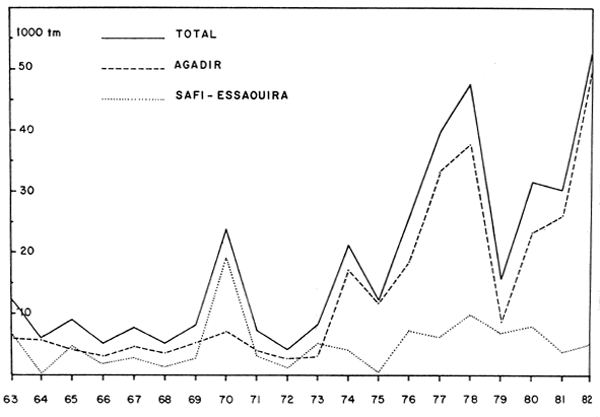
Fig. 5 : Captures de maquereau )Scomberr japonicus) dans la pêcherie marocaine de 1963 à 1982
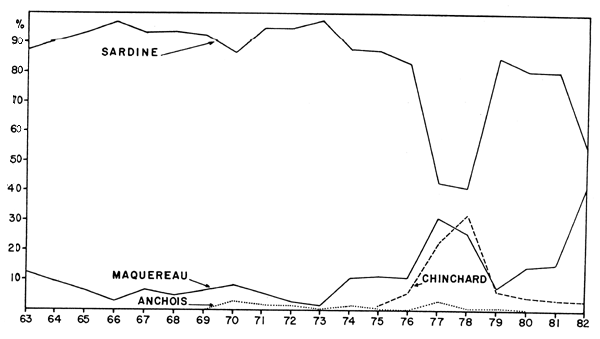
Fig. 6 : Proportions relatives de sardine, maquereau, chinchard et anchois dans les débarquements des senneurs à Agadir, de 1963 à 1982.
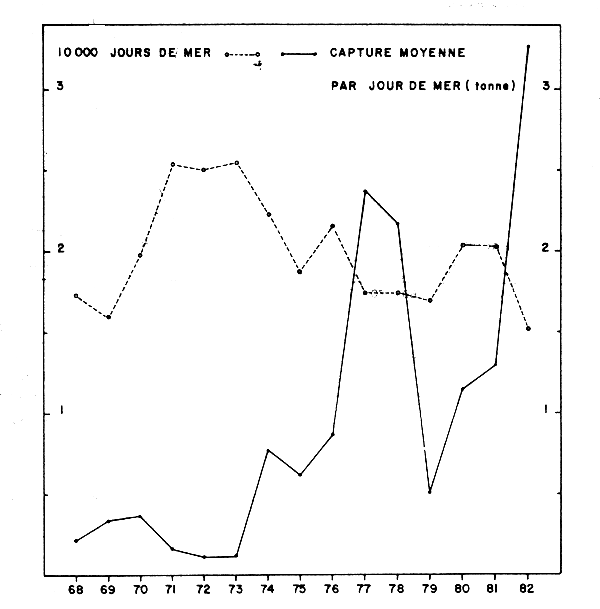
Fig. 7 : Activité de la flotte de senneur d'Agadir (nombre de jours de mer) et prises moyennes annuelles de maquereau par jour de mer (en tonnes) de 1968 à 1982.
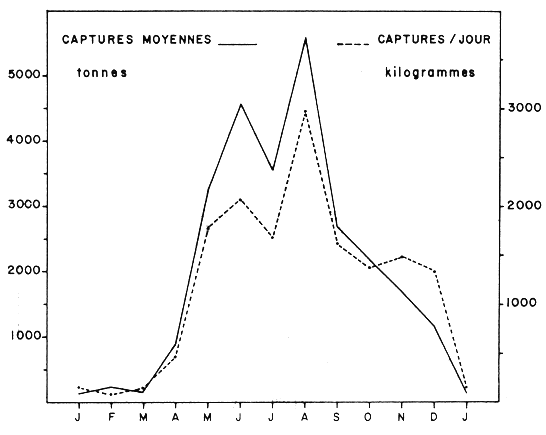
Fig. 8 : Moyennes mensuelles des captures (tonnes) et des captures par jour de mer (kilogrammes) des senneurs d'Agadir pendant la période 1974–1982.
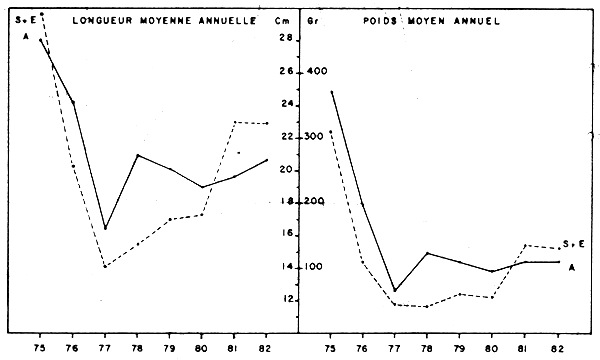
Fig. 9 : Evolutions de la longueur moyenne annuelle (cm) et du poids moyen annuel (g) des maquereaux débarqués à Agadir (A) et dans l'ensemble Safi-Essaouira (S + E) de 1975 à 1982.
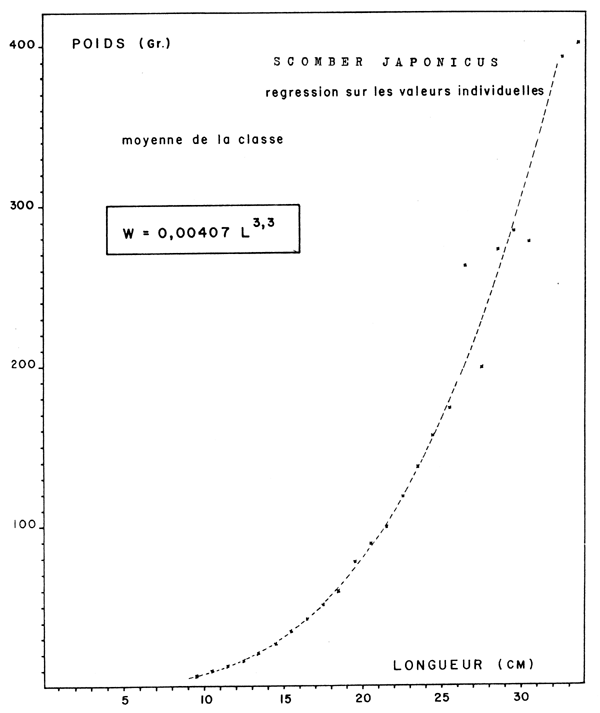
QUELQUES DONNEES SUR LE STOCK DU CHINCHARD DE L'ATLANTIQUE MAROCAIN
par Mehdi El Ouairi, ISPM, Casablanca
1. INTRODUCTION
Trois espéces du genre Trachurus sont présentes sur le plateau continental marocain depuis Cap Spartel jusqu'au sud du cap Bojador (Cap Barbas) : Trachurus trachurus (saurel), représentant plus de 70% des captures de Carangidae, et répandue d'une manière discontinue entre Cap Spartel et Cap Bojador. Les jeunes individus forment des bancs pélagiques prés de la Côte où ils sont capturés avec la sardine par la senne. Les adultes vivent plus au large et sont pêchés par le chalut. Trachurus picturatus, espèce purement pélagique vivant au large. Trachurus trecae, vivant au sud du Cap Bojador où elle est pêchée mélangée avec le saurel. Sa répartition s'étend jusqu'à 15° sud et son comportement est analogue à celui de T. trachurus.
Dans les statistiques de captures, ces espèces ne sont pas distinguées. Par conséquent, la désignation collective de “chinchard” est utilisée dans cet article pour les trois espèces.
2. DISTRIBUTION GEOGRAPHIQUE ET BATHYMETRIQUE DES CONCENTRATIONS
L'analyse des prospections acoustiques et par chalutage montre que le chinchard est largement distribué le long de la côte marocaine entre Cap Spartel et Cap Barbas. De fortes concentrations ont été rencontrées au niveau de Larache (35°10'N) entre Cap Ghir et Essaouira aux profondeurs de 60 à 100 m et au nord du Cap Barbas oú des rendements de 12 tonnes par heure ont été obtenus par Domanevsky (1968).
Le chinchard est rencontré entre la côte et les profondeurs de 300 m. De fortes concentrations sont souvent localisées dans la zone de transition entre la côte et le large, à 100–150 m de profondeur. Les campagnes de prospection par chalutage ont signalé le chinchard dans les secteurs suivants : (a) entre 28°N et 35°N en mai-juin, dans la zone de 20–300 m, avec une forte concentration à 150 m, (b) entre Cap Drâa et Cap Juby, pendant la même pèriode, entre 50 et 75 m de profondeur, (c) entre 28°N et 29°10 N et au niveau d'Agadir en décembre-janvier à 35–65 m.
3. ZONES ET PERIODES DE REPRODUCTION
Avant la ponte, les reproducteurs se concentrent au printemps et au début de l'été dans les profondeurs de 75–200 m. Ils se rapprochent ensuite plus de la côte (50–75 m), où la ponte se fait. Les juvéniles restent quelque temps dans cette zone et y passent leur vie pélagique avant de descendre dans les eaux plus profondes (jusqu'à 300 m) pour commencer leur vie démersale.
Quatre zones de ponte ont été identifiées le long du plateau continental marocain : (1) Safi -Cap Ghir (32°45'-30°30'N), (2) Sidi Ifni-Oued Massa (29°50-29°10N), (3) Cap Drâa-Cap Juby (28°50-28°N), (4) Dakhla-Cap Blanc (Mauritanie) (24°-20°40'N), la dernière zone étant de loin la plus importante.
4. CAPTURES
Le chinchard est capturé par la senne et par le chalut. Dans les débarquements des chalutiers marocains, une augmentation réguliére a été enregistrée depuis 1956 : pendant la période 1956–1965, les captures ont été de 3 000 tonnes en moyenne, en 1966–1975, d'environ 5 000 tonnes et de 1976–1982 près de 10 000 tonnes. Cette augmentation peut s' expliquer par le fait de la diminution des rejets de cette espèqui trouve de plus en plus de débouchés sur le marché intérieur marocain.
Aux captures des chalutiers s'ajoutent les captures des senneurs qui varient considérablement d'une année à l'autre ou même d'un mois à l'autre de la même année. Les individus capturés à la senne sont souvent des juvéniles de tailles inférieures à celles des chinchards pêchés par le chalut de fond.
Les captures les plus importantes de chinchards et aussi des maquereaux ont été obtenues en 1977 et 1978, période où la disponibilité de la sardine a été plus faible.
Les captures marocaines ne permettent pas à elles seules d'évaluer le niveau d'exploitation du stock de chinchard entre Cap Spartel (36°N) et Cap Juby (28°N), zone où la flotte espagnole exerce son activité et dont on ne connait ni les captures ni le volume de rejects des chalutiers autorisés à pêcher, dont l'effort de pêche est supérieur à celui des chalutiers nationaux.
5. PARAMETRES DE CROISSANCE
Les données sur la croissance ne sont pas très récentes puisqu'elles datent de 1976–1977 pour la région comprise entre Tanger et Agadir et de 1978/1979 pour la région Tanger-Cap Juby.
La première courbe de courbe de croissance est celle de Boundi (1977), obtenue selon la méthode d'Allen (1966) et basée sur la lecture d'otolithes de 754 individus :
1t = 61,36 (1-e-0,1011 (t + 0,85))
Une seconde équation a été obtenue lors des campagnes de recherche maroco-soviétiques en 1978/1979 :
lt = 43,39 (1-e-0,19 (t + 1,31))
La différence entre les deux courbes devient plus petite si l'on applique la méthode de Ford-Walford aux données de Boundi :
lt = 53,3 (l-e-0,12 (t + 0,3))
La clé âge-longueur fournie par Boundi est donné au tableau 4.
6. RELATION TAILLE-POIDS
Une relation taille-poids est donnée par Boundi (1977), basée sur 2 500 mensurations d'individus pêchés entre Tanger et Agadir. Cet auteur a séparé des chinchards de moins de 10 cm des plus grands et trouve alors deux équations :
pour le premier groupe : P = 0,023 L2,505;
et pour le second : P = 0,005 L3,12
La relation calculée sur les tailles et poids moyens obtenus lors des campagnes de chalutage exploratoire maroco-soviétique est :
P = 0,019 L2,876
Cette relation est intermédiaire entre celles trouvées par Boundi, bien qu'étant calculée sur des poissons de plus de 15 cm. Comme elle est calculée sur des moyennes par classe de longueur et non sur des valeurs individuelles, elle, est probablement biaisée, cat cette méthode donne un poids équivalent aux classes faiblement ou abondamment représentées.
7. REFERENCES BIBLIOGRAPHIQUES
Allen, K.R., 1966 A method of fitting growth curves of the von Bertalanffy 1966 type to observed data. J.Fish.Res.Bd.Canada (23(2) : 163–179.
Boundi, A., 1977 Contribution à l'étude de la biologie du chinchard (Trachurus trachurus) de l'Atlantique marocain. Mém. 3e cycle, pp. 16 et 29–30
Domanevsky, L.N., 1968 cité par Boundi (1977)
| ANNEE | DEBARQUEMENTS TOTAUX | DEBARQUEMENT DES SENNEURS | DEBARQUEMENT DES CHALUTIERS |
|---|---|---|---|
| 1978 A | 49 659.081 | 44.198.117 | 4.460.964 |
| S | 827.749 | 566.864 | 260.885 |
| E | 3 070 698 | 2 376.076 | 694.622 |
| 1979 A | 9 882 117 | 7 713 194 | 1.168.923 |
| S | 1 623 399 | 1 525 486 | 97 913 |
| E | 2 501 907 | 1 510 823 | 991 084 |
| 1980 A | 6 650 411 | 6 624 407 | 26 004 |
| S | 1 082 168 | 820 803 | 261 365 |
| E | 2 117 940 | 1 539 427 | 638 513 |
| 1981 A | 8.089.707 | 5.896.535 | 2 193.172 |
| S | 174.358 | 150.465 | 23.893 |
| E | 478.677 | 446.414 | 32.263 |
| Année | 1978 | 1979 | 1980 | 1981 | 1982 |
|---|---|---|---|---|---|
| Mois | |||||
| Janvier | 210.586 | 1.764.066 | 4.063.297 | 5.768.741 | 3.267.599 |
| Février | 4.597.251 | 634.823 | 6.509.735 | 10.060.381 | 2.624.481 |
| Mars | 3.528.664 | 503.761 | 10.998.350 | 15.108.165 | 3.398.282 |
| Avril | 3.323.787 | 13.883.043 | 16.713.913 | 14.659.448 | 9.477.080 |
| Mai | 22.413.365 | 9.135.934 | 10.210.371 | 20.406.488 | 4.366.582 |
| Juin | 15.993.145 | 12.638.475 | 18.019.187 | 23.375.716 | 19.339.115 |
| Juillet | 24.271.610 | 18.715.343 | 19.463.482 | 17.364.503 | 18.840.824 |
| Août | 13.287.117 | 15.463.510 | 10.966.399 | 16.684.448 | 23.344.446 |
| Sept. | 23.179.502 | 7.440.312 | 25.542.591 | 21.635.827 | 8.788.758 |
| Oct. | 19.218.491 | 15.574.274 | 26.232.700 | 16.968.298 | 3.288.374 |
| Nov. | 11.513.674 | 10.982.449 | 7.241.843 | 3.531.779 | 16.409.666 |
| Déc. | 3.355.461 | 11.200.330 | 2.167.673 | 4.358.637 | 7.653.145 |
| TOTAL ANNUEL | 114.892.653 | 117.936.320 | 158.129.541 | 169.922.431 | 120.798.352 |
| QUARTIERS ET PORTS | CHALUTIERS COTIERS | CHALUTIERS SARDINIERS | SARDINIERS | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Nbre | TJB | Tonn.moyen | Nbre | TJB | Tonn.moyen | Nbre | TJB | Tonn.moyen | |
| LARACHE | 13 | 926 | 71 | - | - | - | 17 | 303 | 18 |
| ACILAH. | - | - | - | - | - | - | - | - | - |
| KENITRA. | 12 | 444 | 37 | 2 | 110 | 55 | 3 | 35 | 12 |
| RABAT. | - | - | - | - | - | - | 1 | 12 | 12 |
| CASABLANCA. | 29 | 1772 | 61 | 17 | 1126 | 66 | 32 | 1022 | 32 |
| MOHAMEDIA. | 1 | 60 | 60 | 1 | 21 | 21 | 14 | 157 | 11 |
| EL JADIDA | 1 | 26 | 26 | 14 | 200 | 14 | 6 | 70 | 12 |
| SAFI. | 22 | 1296 | 59 | 60 | 3137 | 52 | 57 | 2182 | 38 |
| ESSAOUIRA. | 3 | 106 | 35 | 11 | 1077 | 98 | 25 | 1102 | 44 |
| AGADIR. | 56 | 2800 | 50 | 7 | 505 | 72 | 113 | 9368 | 83 |
| LAAYOUNE. | - | - | - | - | - | - | - | - | - |
| TOTAL | 137 | 7430 | 542 | 112 | 6176 | 55.14 | 268 | 14251 | 53.1 |
| Taille (cm) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | TOT. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 | 1 | 1 | |||||||||||
| 7 | 1 | 1 | |||||||||||
| 8 | 3 | 3 | |||||||||||
| 9 | 3 | 1 | 4 | ||||||||||
| 10 | 1 | 1 | |||||||||||
| 11 | 14 | 10 | 24 | ||||||||||
| 12 | 11 | 48 | 59 | ||||||||||
| 13 | 3 | 34 | 5 | 42 | |||||||||
| 14 | 20 | 1 | 21 | ||||||||||
| 15 | 9 | 2 | 11 | ||||||||||
| 16 | 5 | 2 | 7 | ||||||||||
| 17 | 8 | 7 | 1 | 16 | |||||||||
| 18 | 13 | 27 | 40 | ||||||||||
| 19 | 8 | 58 | 7 | 3 | 76 | ||||||||
| 20 | 5 | 46 | 13 | 64 | |||||||||
| 21 | 2 | 37 | 18 | 2 | 59 | ||||||||
| 22 | 21 | 13 | 7 | 1 | 42 | ||||||||
| 23 | 15 | 20 | 10 | 2 | 47 | ||||||||
| 24 | 10 | 24 | 13 | 2 | 49 | ||||||||
| 25 | 2 | 15 | 10 | 2 | 29 | ||||||||
| 26 | 1 | 5 | 4 | 4 | 14 | ||||||||
| 27 | 1 | 8 | 13 | 5 | 27 | ||||||||
| 28 | 1 | 5 | 5 | 1 | 12 | ||||||||
| 29 | 3 | 8 | 8 | 3 | 22 | ||||||||
| 30 | 7 | 12 | 6 | 25 | |||||||||
| 31 | 2 | 10 | 6 | 18 | |||||||||
| 32 | 2 | 2 | 1 | 1 | 1 | 7 | |||||||
| 33 | 1 | 3 | 2 | 6 | |||||||||
| 34 | 3 | 5 | 8 | ||||||||||
| 35 | 1 | 1 | |||||||||||
| 36 | 3 | 1 | 4 | ||||||||||
| 37 | 1 | 1 | 2 | ||||||||||
| 38 | 3 | 1 | 4 | ||||||||||
| 39 | 2 | 1 | 1 | 4 | |||||||||
| 40 | - | ||||||||||||
| 41 | - | ||||||||||||
| 42 | 1 | 2 | 3 | ||||||||||
| 43 | 2 | 2 | 2 | 6 | |||||||||
| 44 | - | ||||||||||||
| 45 | 1 | 1 | |||||||||||
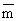 | 11,28 | 14,58 | 20,63 | 23,51 | 26,25 | 29,99 | 32,46 | 37,83 | 40,5 | 43,5 | 42,5 | 42,16 | |
| Ec.ty | 1,68 | 2,63 | 1,97 | 2,41 | 3,13 | 3,50 | 2,83 | 5,03 | 4,94 | 2,30 | |||
| I.C | 0,54 | 0,40 | 0,25 | 0,41 | 0,65 | 0,83 | 1,06 | 5,69 | 4,33 | 2,61 | |||
| N | 37 | 163 | 230 | 128 | 87 | 67 | 27 | 3 | 5 | 2 | 2 | 3 |
I.C. = intervalle de confiance
THE INDUSTRIAL FISHERY FOR COASTAL PELAGICS OFF NW AFRICA
by G.V. Everett and F.C. Roest
UNDP/FAO CECAF Project
North of Cape Blanc
212 Moroccan purse seiners are based at the three main fishing ports of Agadir, Safi and Essaouira. Their target species is sardine, but catches include as first by-catch substantial amounts of mackerel. Since 1974, the quantities of mackerel caught have considerably increased because of their greater availability in relation with hydrographical changes (Belvèze, 1983). The purse seine and trawl catches of horse mackerel and relatively unimportant (Belvèze 1983, El Ouairi, 1983).
Spanish purse seiners from Arrecife, Fuerteventura and Las Palmas also prefer to catch sardine. Their annual by catches of mackerel and horse mackerel vary between 1 000 and 8 000 tons. The activities of these Spanish vessels in 1975 are detailed in the report of the 3rd Session of Working Party on Resource Evaluation (Bravo de Laguna et al., 1976). Up to 31 vessels fished at that time in statistical division 34.1.1.
Details on the fishing agreement between Morocco and Spain are given in Everett et al., (1982).
The Soviet Union operates the largest number of pelagic freezer trawlers in the sub-region, and it seems reasonable that their Atlantik and Mayakovskiy type trawlers of about 2 500 GRT catch 25 tons per day on average. Since 1976 a number of Superatlantik trawlers (3 900 GRT) have joined the fleet and are probably taking a higher catch per day. Soviet flag trawlers are not authorised to fish for pelagics North of Dakhla. The species composition of the USSR catch in the Sahara Division since 1976 was:
| 1976 | 1977 | 1978 | 1979 | 1980 | 1981 | |
|---|---|---|---|---|---|---|
| Total catch (t) | 1.178.988 | 831.037 | 505.455 | 325.980 | 548.973 | 542.715 |
| Hake | 5% | 2% | 1% | 1% | 1% | 1% |
| Sardinella | 4% | 6% | 5% | 3% | 5% | 3% |
| Mackerel | 8% | 11% | 4% | 18% | -- | 15% |
| Horse mackerel | 28% | 30% | 30% | 32% | 26% | 27% |
| Sardine | 43% | 38% | 28% | 34% | 31% | 34% |
| other species | 12% | 13% | 32% | 12% | 37% | 20% |
Target species are therefore the European sardine and the horse mackerels, together contributing 60–70% of the total catch.
The Polish fleet, operating in the Sahara area, caught in the period 1965–1974 35,000–45,000 tons of pelagics per year. Before 1973, mackerel, horse mackerel and hairtails were the dominant species. The sardine becaume target species in 1973 (45% of the catch), and new fishing techniques allowed the sardine catches to increase to over 100,000 tons in 1976 and 1977, when the total catches reached 118,000 and 158,000 tons. The Polish fishing fleet in this region comprised nine trawlers. The activities ceased in early 1978. The average species composition was:
| 1968–1972 | 1975–1977 | |
|---|---|---|
| Sardine | 2% | 87% |
| Horse mackerel | 30% | 5% |
| Mackerel | 20% | 1% |
| Hairtails | 5% | 0 |
| Bluefish | 7% | 0 |
| Sparids | 4% | 0 |
| Sardinellas | 6% | 3% |
In 1969–1978, Bulgaria used to have 4–8 vessels fishing in the Sahara area, their total catches remaining below 50,000 tons. in 1976, the sardine become the dominant species in their catches. Sardine catches increased to 30,000 t in 1977. The average composition shows the change in target species, the period 1974–75 being transitional.
| 1969–1973 | 1976–1978 | |
|---|---|---|
| Sardine | 3% | 63% |
| Horse mackerel | 32% | 27% |
| Mackerel | 33% | 4% |
| Sardinella | 4% | 0% |
| Marine fishes NEI | 23% | 5% |
From 1968 to 1977 it is understood that four Egyptian pelagic freezer trawlers of 1,762 GRT were based on Las Palmas, catching up to 15,000 tons of unspecified fish.
Mauritania
Up to mid 1979 the Soviet pelagic and cephalopod trawler fleet operated in Mauritanian waters. They then stopped until mid April 1980 when the joint venture MAUSOV was formed. Their operations ceased in August 1980. The Company restarted in October 1982 with 20 pelagic trawlers (in addtion to 4 cephalopod trawlers) of which 10 were Superatlantiks and 10 were a Mayakovski /Atlantik/Puskin trawlers, to aim for an annual catch of 200,000 tons. Another Mauritanian company, SIGP, is supplied with fish by Soviet vessels since July 1982. A total of 35 Soviet pelagic trawlers, ranging from 2 100–3 385 GRT, were licensed to fish in the last six months of 1982, but they were not fishing at the same time.
In 1982, Bulgaria had four trawlers of about 2 500 GRT fishing in Mauritania.
In the 1970's, the Romanian annual pelagic catch in the Sahara/ Mauritania waters was of the order of 40,000–80,000 tons. Since 1980 their operations have been restricted to the Mauritanian waters, within the framework of a joint venture with the Mauritanians called SIMAR (see also paragraph 2.3.2 and annex 5 for a detailed description of the Roamnian activities). In 1981–1982, 15–16 pelagic trawlers were operating (11–13 ×3 163 GRT, 3–4 × 2 156 GRT). The total pelagic catches were respectively 77,509 ; 85,567 ; and 84,142 t in 1980–1982, composed of 83%, 68% and 41% of horse mackerel.
The German Democratic Republic fished for sardine in the Sahara area until 1977 (maximum catch 13,000 t), and from 1978 their target species is horse mackerel in the Mauritanian waters (see2.3.1). They operate via the joint venture companies MSP and JTL, using 5 large pelagic trawlers (3 170 GRT) and 12–13 medium-sized ones in 1981–1982. In 1980 and 1981, their catches totalled 87,090 and 103,904 tons, consisting of:
| 1980 | 1981 | |
|---|---|---|
| Horse mackerel | 78% | 86% |
| Sardine | 12% | |
| Sardinella | 7% | 8% |
| Mackerel | 2% | 1% |
According to CNROP files, Irak fishes in 1981 and 1982 with 7 trawlers and 1 seiner in the Mauritanian waters. Spanish vessels fished under an agreement with Mauritania until early 1980. Spain then apparently had 14 seiners in 1981, and only 2 seiners and 1 trawler in 1982 fishing for local companies. The Portuguese Government had an agreement with the Government of Mauritania from 1976 to 1982. Portuguese demersal trawlers were allowed to fish in Mauritanian waters, and 6–8 Portuguese seiners were licensed to catch small pelagic fish for landing to the Nouadhibou fish meal factory. The seiners only worked in 1976–1979 and never caught more than 10,000 tons in any one year. In 1982, 3 Portuguese demersal trawlers were active (until August). See also paragraph 2.3.4 and Table 6.
In addition to the numerous freezer trawlers of distant water nations fishing in the subregion, there were a number of fish meal factory vessels working with a fleet of small purse seiner catching vessels, that operated in Mauritanian waters. “Interpêche” began as a whale factory-ship, the Norwegian “Kosmos V”, and was then bought by South Africans in 1966 who had her converted to a meal factory renamed “Suiderkruis”. Later, the “Suiderkruis” changed name and nationality to “Interpêche”, registered in Bermuda and until 1979 operating from Santa Cruz de Tenerife in the Canary Isles. In 1979, “Interpêche” moved its base to London. All activities ceased in Mauritanian waters after December 1981, when the operations were moved to Chile. The “Willem Barentsz” fished only during 1970 in the area before being returned to South Africa, for sale to South Korea. The “Astra” started fishing in May 1970, and the “Norglobal” in December 1970 whilst “Astra”, under an agreement with the Government of Mauritania, continued operations in the area. “Astra” sank off Guinea on Easter day 1974 and “Norglobal” is believed to have ceased operations in the area at the end of 1975 ; the vessel has since been sold to the Icelandic Government, but was bought by Interpêche for the 1979 season, and called “Interpêche 2”.
Maigret et Brulhet (1973) describe the operations of Interpêche in 1971 and 1972, although these are believed to have started in 1970. The same authors indicate that the catches were 142,436 t in 1971 and 159,620 t in 1972. The company itself declared the following catches : in 1973 161,627 t, in 1974 104,106 t, in 1975 167,943 t and in 1976 81,992 t. The vessel processed about 38,000 t of raw material between Nouakchott and Dakar from March to July 1971. In February and from September to December, the vessel processed about 104,000 t in the vinicity of Nouadhibou. During 1972, the vessel fished off Nouadhibou in January-March and September, November and December catching about 59,000 tons, and off Nouakchott in May, June, July and October to catch about 101,000 tons. In 1973, the vessel fished off Nouadhibou in February-March, October and November to catch about 88,000 tons, and off Nouakchott in May-July and part of September and December to catch about 73,000 tons. Only in June and July 1974 were catches made consistently off Nouakchott whereas the catches off Nouadhibou in February-May and September-December totalled 70,000 tons. Fishing operations off Guinea Bissau started for the first time in 1975 from March to June (see also p.142) yielding a catch of 38,811 tons whereas off Nouadhibou the catch only reached 15,060 tons in two short periods of fishing in September and November. From August to December most fishing was done off Nouakchott where 114,141 tons were caught. In 1976, the vessel fished off Guinea Bissau from January to April where 17,632 tons were caught, and after a refit in Santander (Spain), the vessel fished off Nouadhibou in November and December to catch 64,630 tons. In 1977, 218,490 tons were caught, including 43,697 t in Guinea Bissau. The 1978 catches were slightly less : 196,000 t (of which 19,300 t in Guinea Bissau). In 1979, only 108,300 t were caught in Mauritania (and none in Guinea Bissau), in 1980 a record 274,112 t and in 1981 113,419 t (nine months) (Bergerard and Cheikh, 1981, Bergerard, Domalain and Cheikh 1982).
In 1973 and 1974, “Interpêche” appears to have caught twice as much Sardinella maderensis as S. aurita, whereas in 1975 and 1976, the composition of the catch is reversed. At times between December and June, the vessel has caught Sardina pilchardus off Nouadhibou.
The vessel had freezing facilities for 30,000–40,000 tons per year, preferably for horse mackerel and mackerel. The fish meal factory vessel is of 18,000 GRT with an intake potential of 2,000 tons per day. The actual yearly intake was of the order of 200,000 tons. Three wooden purse seiners of below 200 GRT operated since 1970, the order 11–12 seiners, ranging from 220 to 400 GRT were of Norwegian style, operating on contract for certain months. According to the data provided by Maigret and Brulhet (1973), the catch per boat per fishing day was about 30 tons in 1971 and 1972. Data from the company indicate that the catch per boat per day was 55 tons in 1973 and 42 tons in 1974, increasing to 76 tons in 1975 and decreasing slightly to 67 in 1976. For the first three months of 1977, the catch per boat per fishing day off Mauritania was 71 tons.
The “Astra” catches have been 90,762 tons in 1970, 108,186 tons in 1971, 114,796 tons in 1972 and 66,392 tons in 1973 (until July). The “Norglobal” catches were 92,818 tons in 1971 (including 5,290 tons in December 1970) and 64,857 tons in 1972 (until September). Total Nowegian catches in 1973, 1974 and 1975 were (areas 34.1.3 and 34.3.1) respectively 168,930 tons, 79,377 tons and 23,429 tons. The two vessels tended to fish the same areas at the same time. For example, in March, April and May of 1971 and 1972, the vessels were off the Gambia and/or the Casamance and they were generally stationed off Nouakchott and Nouadhibou for the remaining months. Detailed data need to be collected for the period 1973–1975, until the vessels ceased operating in 1974 and 1975.
Each year, from 1970 through 1974, two other fish meal factory vessels “Devonshire Bay” and “Warwickshire Bay” are believed to have processed 20,000 tons of fish.
North Atlantic purse seiners have often been contracted to catch and land small pelagic for the fish meal factories in Nouadhibou, IMAPEC and SOMIP. The latter was operated by the “Astra” company since May 1974. At that time a number of Canary Island seiners landed to the plant. In 1975 the company closed for a year partially due to departure of management and partially due to law fish meal prices. Then in 1976 it was taken over by the Portuguese as the company COMAPOPE. This closed in July 1982. The closure has been a serious discouragement to establishment of the local purse seiner fleet. There are now only three small (Canary style) purse seiners with Mauritanian flag, which would prefer to sell all Sardinella to the meal factory. Since the meal factory does not function these seiners occasionally go out to look for mackerel and croaker which can be sold to the freezing plants.
Since 15 November 1982, the Mauritanian Government requires that all fish caught in Mauritanian waters be landed or transhipped at Nouadhibou. This allows better control of quantities exported, and the control of foreign exchange earnings. Prior to this date some fish was being landed at Las Palmas. Now a tax is levied on the export of products. The quantity of frozen fish (mostly some pelagics) officially exported from Nouadhibou was 62,000 tons in 1980, 95,000 tons in 1981, and 167,000 tons (provisional) in 1982. Interpêche and some other vessels always transhipped at sea so did not come under official control.
Senegal
Ten (1978) to eighteen (1980) small purse seiners operate from Dakar, each catching on the average 2,400–1,500 tons per year (since 1976). These vessels are usually 20 meters long, have an engine of 300 HP and a net of 450 × 45 meters. The average catch in recent years has been 10–13 tons per 10 hour trip (occasionally two trips may be made per day).
DATA ON DAKAR PURSE SEINERS
| Year | Total catch (t) | Sardinella catch (t) | Number of vessels |
|---|---|---|---|
| 1976 | 31,011 | 26,729 | 13 |
| 1977 | 26,381 | 22,398 | 12 |
| 1978 | 20,901 | 17,965 | 10 |
| 1979 | 25,007 | 22,313 | 15 |
| 1980 | 27,509 | 23,796 | 18 |
| 1981 | 31,363 | 26,769 | 15 |
Source : Centre de Recherches Océanographiques de Dakar-Thiaroye (CRODT)
From 1976 to 1980, Polish vessels fished for pelagics in the Senegalese waters. From 1978, their fishing operations were restricted to Senegal. Their catches increased from 11,400 tons in 1976 to about 76 000 tons in 1980, and consisted in 1977–1980 for 70–80% of Sardinella.
Substantial analysis of the fishing sector has been undertaken with world Bank funds through a consulting company, and long-term alternative actions for development of the sector have been evaluated. One of the options is to reduce the emphasis given to development of secondary ports, and to stimulate utilization of Dakar port facilities. Another option is to encourage fishing of small pelagic species at certain times of the year (this would fish the species which were normally caught by the Polish - 76 000 tons in 11 months of 1980 -by twelve trawlers each employing 20 Senegalese) and for this purpose a Faroe vessel undertook a six month fishing trial starting in December 1981. The vessel “Tollakson” of 47 m operated under the management of the company Senegal Seafood in Senegal waters. The purse seine vessel initially caught mackerel and sardinella at a reasonably high catch rate. However, the reception facilities at Dakar port were not adequate to receive large quantities of small pelagic fish at one time and much time was spent in port between trips. The fish was mostly sold at 45 FCFA for freezing and later sale to Ivory Coast. It was hoped to improve shore facilities so that larger quantities can be landed quickly and the product sold in bulk to such a market as Nigeria. In fact the trial by the Faroe vessel terminated in May 1983 (unconfirmed reasons being low catch rate, declining Ivorian demand for imported sardinella, etc.).
PROJECTED CATCH BY SEASON OF THE SENEGAL SEAFOOD SEINER IN 1983
| 1/1-31/3 | 1/4-30/6 | 1/7-30/9 | 1/10-31/12 | TOTAL | |
|---|---|---|---|---|---|
| Horse mackerel, white | 600 t | 600 t | 1 200 t | ||
| Horse mackerel, black | 3 600 t | 3 600 t | 7 200 t | ||
| S. aurita, big | 600 t | 600 t | 300 t | 300 t | 300 t |
| S. aurita, normal | 600 t | 600 t | 600 t | 600 T | 2 400 t |
| S. maderensis, big | 300 t | 300 t | 1 800 t | 1 800 t | 4 200 t |
| S. maderensis, normal | 3 300 t | 3 300 t | 6 600 t | ||
| Sompat (P. jubelini) | 60 t | 60 t | 120 t | ||
| Fish for fish meal | 1 800 t | 1 800 t | 1 800 t | 1 800 t | 7 200 t |
| Various | 240 t | 240 t | 240 t | 240 t | 960 t |
| TOTAL LANDINGS | 7 740 t | 7 740 t | 8 100 t | 8 100 t | 31 680 t |
Source : CECAF Project Information
The Gambia
In the Gambia, one fishing company specialises in purse seining for Sardinella spp. Since 1975, 11 steel purse seiners have annually taken part in this fishery. The majority of the vessels are 33 m long, have a 300 HP engine, and measure 113 GRT. In 1975, half the trips made were of a day's length, a third of two days, the remainder of the trips being of longer length (Scheffers, 1976).
The catches of this company increased from 3 900 tons in 1975 (7 vessels) to 14 000 tons in 1978 (11 vessels) after which they dropped as the number of vessels was reduced to 7 in 1980, and 6 in 1981. This fleet seldom captures mackerels and horse mackerels (Taylor-Thomas, personal communication).
The recently published “Yearbook of fisheries statistics, The Gambia, 1981” presents the annual total industrial catch for all fishing methods combined, without species breakdown or effort data.
Guinea Bissau
Purse seining for pelagic fish in Guinea Bissau started in 1975 with four Norwegian and ten Dutch owned purse seiners operating for the factory ship 1'Interpêche. They fished until 1978. In 1977, 3 Ghanaian and 1 French seiners joined the fishery, which was greatly expanded in 1978 when 42 Russian and 3 further French seiners arrived. The Russian seiners subsequently reduced in number to 27 in 1979 and 9 in 1980, as can be seen from the following Table :
| Country | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | |
|---|---|---|---|---|---|---|---|
| France | 1 | 4 | 3(Jan) | ||||
| USSR | 42 | 27 | 9 | ||||
| Ghana | 3 | 3 | |||||
| Norway/Holland | 14 | 14 | 14 | 14 | |||
| TOTAL | 14 | 14 | 18 | 63 | 30 | 9 | |
| 50 | 2 | 2 | 2 | 2 | |||
| 100 | 1 | 1 | 1 | 1 | |||
| GRT Class | |||||||
| 150 | 5 | 5 | 8 | 8 | |||
| 200 | 6 | 6 | 6 | 13 | |||
| 500 | 1 | 39 | 30 | 9 | |||
The numbers of pelagic trawlers seem to have been very limited : in 1978, 1979 and 1980 only 2, 3 and 1 French trawlers were reported. It seem reasonable, however, to assume that other unspecified “trawlers” spent at least part of their time trawling for pelagic species. In 1978 and 1979 respectively 43 and 25 large Russian trawlers fished in Guinea Bissau waters.
Catch data available for Interpêche indicate that Sardinella maderensis (60% of the catches in 1975, 17% in 1976 and 48% in 1977), the horse mackerels (30% in 1975, 83% in 1976, and 7% in 1977), and Sardinella aurita (32% in 1977) were the dominant species. No data are available for 1978. The total catches were respectively in 1975, 1976, 1977 and 1978 : 38,811 t, 17,632 t, 43,697 tand 19,284 t.
The catch data of Soviet vessels, not landing at Bissau for the 12 Months period March 1978-February 1979 indicate that of their total catch of 87,540 tons, 27,307 t were transformed into fish meal (apparently yielding 5,261 t.), while 59,000 t were frozen. Of the latter, 33,706 t were pelagic species (Sardinella 55%, Trachurus 38% Decapterus 7%, Caranx 0,4%, mackerel 0,2%).
According to data CECAF files, during the first eleven months of 1982, the USSR fleet caught 118,200 t, of which not less than 94,000 t were of (semi-) pelagic species :
| Balistes | 59,673 t | Caranx | 909 t |
| Trachurus | 27,249 t | Vomer | 788 t |
| Sardinella | 4,944 t | Lichia | 228 t |
The number of vessels taking part in the fishery ranged monthly from 7 to 27, the total number of fishing days being 2,987.
Finally, out of a total of 1,691 tons, the Japanese bottom trawler Taiyo Maru 67 (not specializing in Cephalopods) caught between April 1978 and March 1979, 339 t of pelagic fish (20%). Eight Korean cephalopod trawlers-also fishing in Guinea Bissau waters - landed between March and August 1978, 9 133 tons of horse mackerels in Las Palmas, which represented 3,7% of their total catch;
Cape Verde
In Cap Verde, small pelagic fish (horse mackerel, etc.) are caught to be used as live bait for the pole and line tuna boats. Some data on quantities of unused bait is given in Table 5 of annexe 4.
NORTHWEST AFRICAN SMALL PELAGIC INDUSTRIAL OPERATIONS : Indicative data
| Numbers of vessels Trawlers | Information on target species and name of management company (joint venture) | |||
|---|---|---|---|---|
| Morocco/Sahara | Seiners | Large | Small | |
| Local (Agadir, etc.) | 212 | 0 | 60? | Sardine fishery (Belèze, 1983) |
| USSR | 10? | 20? | 0 | Prefer sardine |
| S. Leone/Morocco | 0 | 10 | 0 | Soviet vessels, prefer mackerel |
| Spain | ||||
| Canary Isles | 31 | 0 | 0 | Lanzarote, Arrecife, Fuerteventura, |
| Las Palmas, 1975. Prefer Sardine | ||||
| Mauritania | ||||
| Romania | 0 | 15 | 0 | JTL, 1982, (SIMAR) |
| Nigeria | 2 | 1 | 0 | 1981 only |
| GDR | 0 | 20 | 0 | SACT, CAM, 1982 (MSP and JTL) |
| USSR | 0 | 33 | 0 | SOCOPA, CAM (July-Dec., 1982) |
| (MAUSOV, SIGP) | ||||
| Bulgaria | 0 | 4 | 0 | SACT |
| Spain | 2 | 0 | 1 | SMTC |
| Portugal | 0 | 0 | 3 | SMTC, SMAIT, SOMAT/G (COMAPOPE- |
| stopped in 7/82) | ||||
| Mauritania | 3 | 0 | 0 | SMTC, SMAIT, SOMAT/G (COMAPOPE - |
| stopped in 7/82) | ||||
| Interpêche | 15 | 0 | 0 | stopped in October 1981 |
| Iraq | 0 | 8 | 0 | SOMACAT (SAMIP) |
| Senegal | ||||
| Local | 14 | 0 | 0 | Small seiners |
| Senegal Seafood | 1 | 0 | 0 | Jan-Dec. 1982 |
| Polish | 0 | 9 | 0 | Until Nov. 80 |
| The Gambia | ||||
| Seagull/Ghana | 6 | 0 | 0 | 6–11 in 1975–1981 |
| Guinea Bissau | ||||
| USSR | 27 | 26 | 0 | 1979 data |
| Interpêche | 14 | 0 | 0 | stopped in 1978 |
REFERENCES
Belvèze, H., 1983 La pêcherie de maquereaux (Scomber japonicus) de l'Atlantique marocain entre 29°N et 33°N, Annex 7 to this report.
Bergerard, P., and M.O. Cheikh, 1981 Estimation des captures totales dans la ZEE mauritanienne en 1980. Bull. CNROP 9 (2) : pag.var.
Bergerard, P., G. Domain, and M.O. Cheikh, 1982 Pêche industrielle : la pêche internationale. Bull.CNROP 10 (2) : 11–108.
Bravo de Laguna, J., M.A.R. Fernandez, E. Santaella, J.C. Santana and J. Gomez. 1976 Report on the activity in the CECAF area of the Spanish fleet operating from the Canary Island in 1975. FAO. Fish.Rep. (183) : 87–110.
El Ouairi, M., 1983 Quelques données sur le stock du chinchard de l'Atlantique marocain. Annex 8 to this report.
Everett, G.V., M. Ansa-Emmim, M.A. Robinson and F.C. Roest, 1982 Recent trends in CECAF fisheries. CECAF/TECH/82/42 : 59p.
Maigret, J., and J. Brulhet, 1973 Informations sur la pêche des côtes mauritaniennes. Nouadhibou, Bull.Lab. Pêches 2 : 113–122.
Scheffers, W.J., 1976 The fishery resources of The Gambia. Project Technical Paper No. 1. Development of Inshore Fishery, The Gambia, Project GAM/72/006, 24p. (mimeo).
EXPECTED F.O.B. PRICES FOR FROZEN SMALL PELAGICS (November 1982, Dakar)
| Species | CFA per kilo |
|---|---|
| Horse mackerel, white | 165 |
| Horse mackerel, black | 135 |
| S. aurita, big | 115 |
| S. aurita, normal | 107 |
| S. maderensis, big | 110 |
| S. maderensis, normal | 100 |
| Sompat (Pomadasys jubelini) | 240 |
| Various frozen fish | 130 |
N.B. : Freezing, handling and packing costs are about 75 FCFA per kilo. Fish for meal (fresh, quayside) is sold for 8 FCFA per kilo.
Annual catch by country in the Eastern Central Atlantic
Metric tonnes
| Year | ||||
|---|---|---|---|---|
| 1977 | 1978 | 1979 | 1980 | |
| Angola | 400 | 400 | 400 | 400 |
| Benin | 4 378 | 4 852 | 4 852F | 4 852F |
| Bulgaria | 48 454 | 31 153 | - | 49 811 |
| Cameroon | 20 168 | 18 996 | 19 421F | 19 421F |
| Cape Verde | 8 331 | 8 331 | 7 476F | 8 354F |
| Congo | 15 364 | 16 297 | 19 630 | 19 926 |
| Cuba | 20 700 | 7 200 | 7 500 | 9 258 |
| Egypt | 15 000 | - | - | - |
| Equatorial Guinea | 4 000F | 4 000F | 4 000F | 2 500 |
| France | 67 242 | 65 270 | 57 317 | 65 229 |
| Gabon | 5 656 | 12 950 | 13 203 | 26 417 |
| Gambia, The | 15 027 | 11 979 | 9 058 | 10 255 |
| German Democratic Republic | 18 820 | 8 524 | 46 635 | 87 118 |
| Ghana | 210 715 | 235 299 | 195 315 | 186 606 |
| Greece | 26 956 | 18 065 | 20 185 | 20 195F |
| Guinea | 8 120F | 9 000F | 17 453 | 17 453F |
| Guinea Bissau | 3 758 | 3 653 | 2 013 | 5 234 |
| Italy | 26 234 | 20 241 | 28 282 | 29 825 |
| Ivory Coast | 79 307 | 74 908 | 71 613 | 61 896 |
| Japan | 67 796 | 63 217 | 44 836 | 27 676 |
| Korea (Republic oF) | 99 043 | 94 338 | 92 720 | 80 263 |
| Liberia | 6 190F | 6 812 | 9 484 | 7 791 |
| Mauritania | 21 170F | 21 170F | 21 170F | 21 170F |
| Morocco | 221 678 | 258 390 | 254 768 | 307 194 |
| Nigeria | 263 850 | 272 581 | 275 803 | 292 390 |
| Poland | 203 429 | 49 302 | 67 703 | 78 797 |
| Portugal | 20 301 | 15 294 | 11 023 | 15 664 |
| Romania | 78 431 | 49 202 | 45 507 | 77 509 |
| Sao Tome and Principe | 1 400F | 1 500F | 1 500F | 1 500F |
| Senegal | 278 857 | 294 052 | 250 259 | 254 951 |
| Sierra Leone | 51 452 | 47 580 | 46 237 | 34 205 |
| Spain | 442 064 | 493 548 | 376 625 | 427 585F |
| Togo | 7 623 | 12 139F | 4 598 | 4 598F |
| USSR | 1 134 433 | 769 500 | 526 011 | 942 334 |
| USA | 10 616 | 5 789 | 2 764 | 2 650 |
| Zaire | 1 155 | 741 | 693 | 693F |
| Other NEI | 225 540F | 206 760F | 206 990F | 207 114F |
| TOTAL | 3 733 658 | 3 213 033 | 2 763 044 | 3 408 834 |
Source : FAO Yearbook oF Statistics, Vol. 50 and CECAF estimates
![]()
![]()
![]()