![]()
![]()
by
Guerlorget et al., 1986
| OCEANOLOGICA ACTA 1986 - VOL. 9 - No1 |  |
| Contribution à l'étude écologique des lagunes septentrionales de la baie d'Amvrakia (Grèce) | Écologie Sédimentologie Lagunes Actuel Grece Ecology Sedimentology Lagoons Present Greece |
Groupe d'étude du domaine paralique : Olivier GUELORGETa , Guy-François FRISO-NIb, Dominique MONTIa, Jean-Pierre PERTHUISOTc
a Laboratoire d'Hydrobiologie Marine, Université des Sciences et Techniques du
Languedoc, Place Eugène Bataillon, 34060 Montpellier Cedex.
b CEMAGREF, Domaine de la Valette, 34000 Montpellier Cedex.
c Laboratoire de Biogéologie et Biostratigraphie, Université de Nantes. 2. chemin de
la Houssinière, 44072 Nantes Cedex.
Reçu le 4/9/84, révisé le 16/7/85. accepté le 22'7/85.
RÉSUMÉ
Les bassins lagunaires du delta du Louros et de l'Arachtos sont étudiés quant à leur hydrochimie, leur sédimentologie et leurs peuplements phytoplanctoniques et benthiques.
Le complexe lagunaire comprend deux ensembles, l'un sursalé à tendance subévaporitique en été (Logarou), l'autre sous-salé à tendance dulçaquicole constante (Tsoukalio-Rhodia), en fonction de leur bilan en eau douce.
Ces deux ensembles présentent une même zonation biologique en fonction du champ de confinement avec des peuplements typiques du proche paralique sauf dans les zones distales sursalées ou sous-salées, à la limite du Paralique lointain.
Les dépôts sédimentaires évoluent latéralement en fonction de l'organisation hydrochimique et biologique, notamment en ce qui concerne les phases chimiques et biogéniques (carbonates, matière organique).
Cette organisation biogéologique est conforme au schéma du domaine paralique méditerranéen proposé par Guelorget et Perthuisot (1983).
Oceanol. Acta, 1986, 9, 1, 9–17.
ABSTRACT
Contribution to the ecological study of the northern lagoons of the Gulf of Amvrakia (Greece)
The hydrochemistry, sedimentology and phytoplanktonic and benthic populations of the lagoonal basins of the Louros and Arachtos delta are studied. The lagoonal complex comprises two systems: Logarou lagoon has higher salinities than Amvrakia Bay in summer; Tsoukalio and Rhodia lagoons are more subject to fresh water inputs throughout the year.
These two restricted systems display the same biological zonation according to the confinement field, and support near paralic populations, with the exception of the farthest parts, which tend to hypersaline or fresh water middles (Far paralic).
The sedimentary deposits depend on the hydrochemical and biological organization. especially where chemical and biogenic phases (carbonates, organic matter) are concerned.
This biogeological organization fits with the schema for the Mediterranean paralic realm as proposed by Guelorget and Perthuisot (1983).
Oceanol. Acta, 1986, 9, 1, 9–17.
INTRODUCTION
La présente étude s'inscrit dans la logique des recherches pluridisciplinaires entreprises par le GREDOPAR (Groupe d'étude du domaine paralique), qui visent à comprendre l'organisation biogéologique des bassins paraliques, c'est-à-dire des bassins situés entre mer et continent (Guerloget, 1985), enclavés dans le domaine continental mais en relation plus ou moins directe avec la mer.
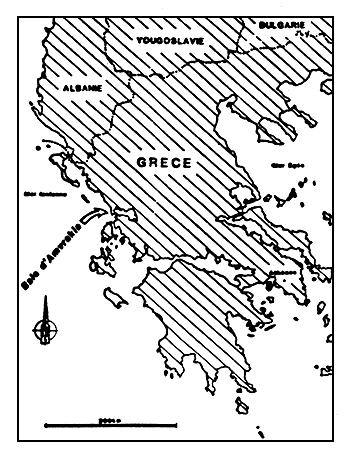
Figure 1
Situation géographique de la baie d'Amvrakia.
Location of the Gulf of Amvrakia.
Dans ces milieux, notamment ceux qui sont le plus directement influencés par la mer (Proche paralique), le confinement, qui peut s'exprimer comme le temps de renouvellement des éléments d'origine marine en un point donné, est le facteur directeur de la zonation biologique. Dans le Paralique lointain, à tendances évaporitiques ou dulçaquicoles, les peuplements sont originaux et différents en fonction de la salinité du milieu (Guélorget, Perthuisot, 1983).
CADRE GÉOGRAPHIQUE ET GÉOLOGIQUE
La baie d'Amvrakia s'ouvre sur la côte Nord-Ouest de la Gréce et communique avec la Mer Ionienne par un chenal étroit près de Preveza (fig. 1 et 2). Ce golfe est enserré dans une zone montagneuse au relief parfois accusé.
Les deux principaux tributaires du golfe sont les fleuves Louros et Arachtos, qui se jettent dans sa partie nord et forment un delta commun abritant un complexe lagunaire constitué par trois lagunes formant deux ensembles, Tsoukalio et Rhodia d'une part, Logarou d'autre part.
Aperçu de l'environment géologique
L'arrière-pays du delta, ainsi que les quelques pointements du substratum qui émergent de la plaine deltaïque, sont constitués d'unités charriées appartenant à la «zone ionienne», c'est-à-dire à la partie externe de l'édifice orogénique hellénique (Aubouin, 1959).
Le delta est constitué d'une plaine alluviale qui enserre les lagunes, celles-ci n'étant séparées de la baie d'Amvrakia que par des cordons littoraux interrompus par des passes. Cette physiographie est liée à 3 phénoménes principaux (Szijj, 1981):
Ainsi, la plaine deltaïque est constituée de cônes alluviaux juxtaposés et son état actuel dérive d'un delta de type «fluvial» (Galloway, 1975) à morphologie digitée; les cordons qui limitent le complexe lagunaire vers le sud, constitués de sables coquillers, sont ancrés soit sur d'anciennes barres transversales sous-marines soit sur des pointements de substratum.
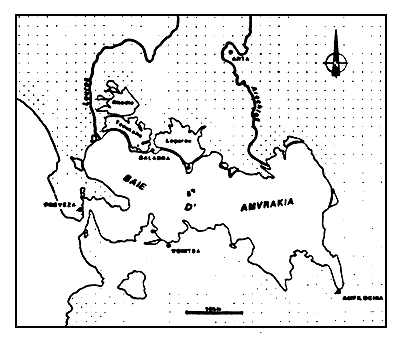
Figure 2
La baie d'Amvrakia et ses annexes paraliques
The Gulf of Amarakia and its paralic dependencies
Figure 3
Les concentrations des eaux de la baic d'Amvrakia et
des lagunes du delta du Louros en avril 1982. Lignes
pointillées: anciens cours du Louros. Lignes tiretóes:
canaux de drainage du delta.
Salt concentrations of waters in the Gulf of Amvrikia and
the Louros delta lagoons in April 1982. Dotted lines: earlier
velleys of the Louros. Hatched lines: drainage canals of
the delta.
Données climatiques
Le climat de cette région est typiquement méditerranéen:
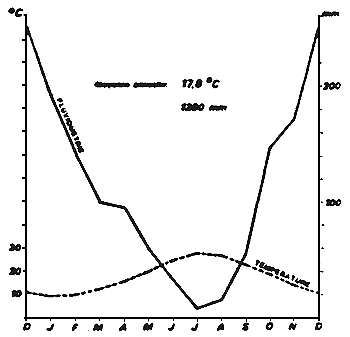
Figure 4
Régime des pluies et des températures à Arta (1930–1975).
Mean rainfalls and temperatures at Arta. (1930–1975).
Principaux tributaires: le Louros et l'Arachtos
Fleuves méditerranéens, courts et à forte pente, le Louros et l'Arachtos ont des vitesses d'écoulement élevées et des débits trés variables; ceux-ci sont maximaux à la fonte des neiges.
Il n'existe pas de donnée précise sur les régimes de ces deux fleuves, mais il semble que le débit moyen de l'Arachtos soit le plus important. C'est sans doute lui qui a participé de façon prépondérante à l'édification du delta et dont les apports sédimentaires massifs sont responsables du déplacement d'Est en Quest du cours du Louros sur la plaine alluviale.
LES LAGUNES DU DELTA DU LOUROS ET DE L'ARACHTOS
Le complexe lagunaire est séparé de la baie d'Amvrakia par un mince lido coupé de quelques passes qui assurent des échanges restreints entre les lagunes et la baie. Ainsi, il constitue une transition brutale entre un milieu marin et un écosystème très confiné habituellement beaucoup plus éloignés l'un de l'autre.
Le complexe lagunaire du delta comprend deux ensembles principaux séparés par un canal débouchant sur la baie dans la région de Salaora (fig. 5). Le premier se compose d'une seule lagune, Logarou, 4900 ha. Le deuxième comporte deux lagunes reliées par une passe étroite: Tsoukalio (2900 ha) et Rhodia (2600 ha).
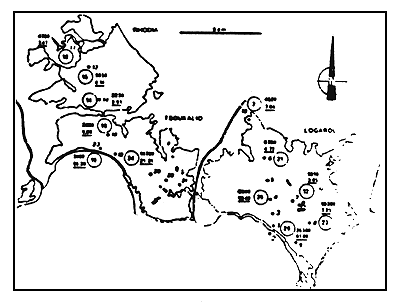
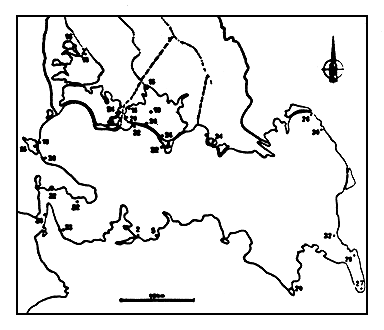
Figure 5
Situation des stations de prélévement dans le complexe lagunaire en
juin 1982.
Location of sampling points in the lagoonal complex in June 1982
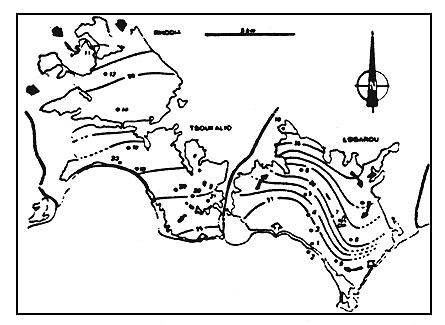
Figure 6
Les concentrations des eaux (en g/l) du complexe lagunaire en juin
1982.
Salt concentrations (g/l) in waters of the lagoonal complex (June 1982)
La tranche d'eau est faible (1 à 2 m) sauf dans le centre de la lagune de Rhodia où elle atteint 3 m. Le fond des lagunes est constitué de sédiments fins. vaseux, parfois organiques; à proximité du lido, le sédiment s'enrichit en éléments plus grossiers de la taille des sables.
Les eaux
Les concentrations
En juin 1982, époque de la présente étude, les concentrations observées sont comprises entre 18,5 et 35 g/l. A cette saison, les deux ensembles lagunaires présentent des gradients de concentration inverses, l'un négatif vers la soussalure, l'autre positif vers la sursalure (fig. 6). Notons que dans le cas de Logarou, cette sursalure ne se manifeste qu'á la belle saison, comme en témoignent des mesures de salinité effectuées au printemps précédent (fig. 3).
Ces situations estivales, diamétralement opposées, s'expliquent par la balance en eau douce de chaque bassin:
La qualité des eaux
Lagune de Logarou
Les températures relevées le 17 juin 1982 entre 9 et 12 h s'inscrivent entre 22,5°C et 31°C. Elles mettent en évidence la surchauffe des eaux périphériques et l'influence des eaux fraiches en provenance de la baie. suivant un schéma voisin de celui des concentrations.
Le pH varie entre 8,2 et 9,1 (station 8) avec un gradient positif vers les confins lagunaires. Le taux d'oxygènc dissous atteint 13 ppm dans les herbiers de bordure, et suit sensiblement la même évolution sauf à la station 8 où, malgré un fort pH, indice d'une activité photosynthétique élevée, le taux d'oxygène dissous reste modéré à 7,8 ppm : ceci est sans doute en relation avec une forte activité bactérienne liée à l'abondance de matière organique dans le sédiment (cf. infra).
Lagunes de Tsoukalio et Rhodia
Les paramétres physico-chimiques sont relativement homogénes sur l'ensemble des deux bassins. Les températures relevées le 19 juin 1982 entre 9 et 12 h s'inscrivent entre 26, 1°C et 26, 9°C; le pH entre 8,5 et 8,6; le taux d'oxygène dissous entre 7,2 ppm et 8,4 ppm.
Seul le point 11 est original: les trois paramètres y présentent une légère baisse, en s'inscrivant respectivement à 25,2°C, 8,3 et 5,9 ppm. Ces chiffres traduisent l'influence des eaux continentales, et expliquent la présence d'une roselière dans cette zone.
Quelques profils verticaux effectués dans la lagune de Rhodia, la plus profonde, mettent en évidence un léger gradient qui se traduit, de la surface vers le fond, par un abaissement de la température de 0,5°C et d'une augmentation de la conductivité de 0,3 mmho/cm.
Le phytoplancton
Dans l'ensemble Tsoukalio-Rhodia, la biomasse s'organise selon un gradient positif depuis les zones méridionales vers les confins septentrionaux (fig. 7). Localement, le gradient est perturbé par la présence de blooms bien visibles, sous la forme de taches d'eaux vertes ou jaunes de taille hectométrique.
Dans Logarou, le maximum de biomasse est enregistré dans la partie centrale de la lagune avec 1,9 mg Chl a/m3; à partir de cette zone, la biomasse décroît vers la périphérie du bassin.
Les valeurs du taux de phéophytine dans les pigments indiquent qu'il s'agit d'un plancton sénescent. Ce caractère est particulièrement marqué à la station 11 sous influence continentale dominante dans Rhodia, et aux stations 7 et 8 de Logarou: cette zone, et notamment la station 8 correspond à un ombilic hydraulique (Guélorget et al., 1984), où aboutissent les eaux aprés leur périple dans le bassin. Elles sont alors fortement chargées en phytoplancton sénescent.
En revanche, les eaux fraîchement entrées dans le bassin sont plus productives et comportent un phytoplancton jeune relativement pauvre en phéophytine: c'est ce qu'on observe dans la zone centrale de Logarou (tab. 1).
Par rapport au phytoplancton de la baie d'Amvrakia étudié par Szijj (1981), les lagunes du delta présentent, avec une vingtaine d'espèces, une richesse taxonomique équivalente. Certaines espèces se trouvent dans les deux milieux, comme Peridinium trochoïdeum, P. depressum, P. granii, P. leonis. Toutefois, il n'a pas été recensé, dans les lagunes, d'espèces typiquement néritique, comme Thalassiotrix frauenfeldi que Szijj signale dans le golfe. De même, les péridiniens néritiques Prorocentrum micans, Exuviella baltica et E. marina sont remplacés dans les lagunes par des espèces typiquement paraliques comme Prorocentrum scutellum, Exuviella compressa et Gymnodinium nelsoni.
Le peuplement phytoplanctonique des lagunes est dominé par les cryptophycées, mais dans les zones les plus reculées de Logarou (station 10), le peuplement est partagé entre les péridiniens, les diatomées pennées et les cyanophycées qui proviennent peut-être des tapis algaires qui colonisent ces zones. En revanche, dans les zones confinées de Rhodia, les chlorophycées apparaissent dans le nannoplancton: elles semblent indiquer ici l'influence des apports d'eaux continentales.
Les peuplements phytoplanctoniques, relativement homogènes sur l'ensemble du complexe lagunaire, sont typiquement paraliques. Leur état physiologique en juin 1982 est assez dégradé, sans que l'on atteigne cependant le stade de la dystrophie. Les valeurs de biomasse chlorophyllienne sont assez faibles par comparaison avec d'autres bassins du pourtour méditerranéen (Guélorget, Perthuisot, 1983). Enfin, les différences de composition taxonomiques des zones les plus confinées du complexe lagunaire traduisent l'opposition entre un bassin à tendance évaporitique (Logarou) et un ensemble fortement influencé par les peuplements dulçaquicoles (Tsoukalio-Rhodia), matérialisant ainsi les deux pôles hydrochimiques du domaine paralique.
Figure 7
Répartition de la biomasse phytoplanctonique dans le complexe
lagunaire en juin 1982 (en mg de Chl a/m3).
Distribution of the phytoplanctonic biomass in the lagoonal complex,
June 1982 (in mg of Chl a/m3).
Le beathos
Les différentes stations se situent le long de transects sensiblement NS, qui recoupent tous les termes de passage entre la baie et les reculées les plus continentales.
La macroflore
Elle est représentée principalement par 3 espèces de macrophytes qui se répartissent différemment le long des transects et ce, en fonction du confinement. Zostera moltii domine dans les zones voisines du lido. Au-delà apparaît Ruppia spiralis, formant habituellement des herbiers mixtes avec Chara sp. En allant vers les zones les plus confinées, les herbiers disparaissent et cèdent la place, soit aux tapis cyanobactériens dans les zones à tendance évaporitique (bordure Nord de Logarou), soit aux chlorophycées Ulva sp. et Enteromorpha sp. (bordure Nord de Tsoukalio et lagune de Rhodia). La zone centrale profonde de Rhodia est dépourvue de macrophytes.
En résumé, l'étude qualitative des peuplements végétaux montre qu'à partir de la baie, les milieux lagunaires limitrophes se confinent très vite: Zostera noltii, espèce indicatrice d'un confinement déjà bien marqué, colonise seulement la frange méridionale du complexe lagunaire, alors que Ruppia spiralis, éspéce strictement paralique, indicatrice de confinement très avancé, colonise la majeure partie des lagunes de première ligne.
Tableau 1
Biomasse phytoplanctonique du complexe paralique du delta de Louros et de l'Arachtos (juin 1982).
Phytoplanktonic biomass in the lagoonal complex of the Louros and Arachtos delta.
| Stations | Chlorophyllc à mg/m3 | Phéophytine mg/m3 | Pourcentage de phéophytine | Total |
| Logarou | 0.87 | 0.74 | 46% | 1.61 |
| 4 | 1.94 | 1.57 | 45% | 3.51 (Bloom) |
| 5 | 0.82 | 0.40 | 33% | 1.22 |
| 6 | 0.72 | 0.80 | 53% | 1.52 |
| 7 | 0.53 | 0.55 | 51% | 1.08 |
| 8 | 0.47 | 0.44 | 48% | 0.91 |
| 10 | ||||
| Tsoukalio-Rhodia | 0.40 | 0.30 | 43% | 0.70 |
| 21 | 0.78 | 0.78 | 50% | 1.56(Bloom) |
| 20 | 1.10 | 0.91 | 45% | 2.01(Bloom) |
| 20 | 0.83 | 0.69 | 45% | 1.52 |
| 18 | 0.49 | 0.59 | 55% | 1.08 |
| 14 | 0.97 | 0.52 | 35% | 1.49 |
| 12 | 1,21 | 1,39 | 53% | 2,60 |
| 11 |
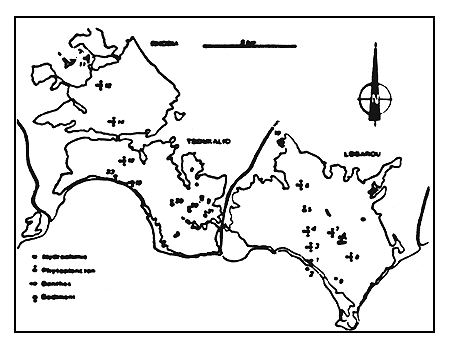
Figure 8
Richesse spécifique (chiffre cerclé), densité et biomasse
(chiffre souligné) de la macrofaune benthique dans le
complexe lagunaire en juin 1982.
Specific richness (circled member), density and biomass
(underlined member) of the benthic macrofauna of the
lagoonal complex, June 1982.
Notons enfin, dans la partie médiane de la frange Sud de Tsoukalio, l'abondance d'Acetabularia mediterranea, formant localement des tapis homogènes dont l'implantation est favorisée par l'abondance des valves de pélécypodes morts.
La macrofaune
Dès le passage du lido, les peuplements se composent en majorité d'espéces strictement paraliques caractéristiques des zones IV et V (Guélorget, Perthuisot, 1983). Par exemple, à la station 1 les espèces dominantes sont toutes des espèces paraliques: Gammarus insensibilis, Corophium insidiosum, Sphaeroma rugicauda, Capitella capitata, Nereis diversicolor, Cerastoderma glaucum, Brachydontes marioni, Abra ovata, des Hydrobiidae, des larves de Chironomidae. Seules quelques espèces mixtes caractéristiques de zone III subsistent, comme Glycera convoluta, Gibbula adamsoni et Venerupis decussata, mais elles disparaissent rapidement dès que l'on pénètre plus profondément dans la lagune.
Dans les zones les plus confinées, on aboutit soit à un peuplement quasiment monospécifique á Pirenella conica dominante et quelques Hydrobia acuta dans les confins subévaporitiques de Logarou (station 10) colonisés par un recouvrement cyanobactérien, soit à un peuplement plurispécifique mais peu diversifié où se mèlent aux espèces paraliques des représentants d'associations dulçaquicoles (oligochètes, trichoptères, odonates, Cricotopus, genre de Chironomidae différent de Chironomus strictement paralique) aux confins septentrionaux de Rhodia (fig. 8).
La richesse spécifique (tab. 2, fig. 9) diminue significativement du Sud vers le Nord, c'est-à-dire vers les confins lagunaires. Cette diminution graduelle est très prononcée à Logarou, tandis que dans l'autre système, il existe une diminution brutale dans Tsoukalio au-delà de laquelle la richesse spécifique reste à peu près constante à cause de l'intrusion d'espèces dulçaquicoles.
Tableau 2
Densité (individus/m2) et biomasse (g/m2 - poids sec après décalcification) des peuplements benthiques des trois lagunes du delta.
Density (individuals/m2) and biomass (g/m2) of the benthic fauna in the three delta lagoons.
| Station | 1 (Logarou) | 4 (Logarou) | 6 (Logarou) | 7 (Logarou) | ||||
| Densité | Biomasse | Densité | Biomasse | Densité | Biomasse | Densité | Biomasse | |
| Pelecypodes | 4910 | 42,060 | 920 | 3,198 | 2800 | 3,748 | 650 | 1,541 |
| Gastropodes | 300 | 0,739 | 340 | 4,245 | - | - | 350 | 1,298 |
| Crustacés | 12000 | 14,469 | 3390 | 0,967 | 3140 | 0,837 | 150 | 0,140 |
| Polychètes | 4440 | 2,985 | 4290 | 1,987 | 620 | 0,108 | 160 | 0,075 |
| Chironomides | 2610 | 0,433 | - | - | 30 | 0,009 | - | - |
| Divers | 2120 | 0,398 | - | - | 190 | 0,021 | - | - |
| Total | 26380 | 61,084 | 8940 | 10,397 | 6780 | 4,723 | 1310 | 3,054 |
| Station | 8 (Logarou) | 10 (Logarou) | 11 (Rhodia) | 12 (Rhodia) | ||||
| Pelecypodes | 4430 | 2,780 | - | - | 480 | 5,130 | 2010 | 2,611 |
| Gastropodes | 760 | 1,385 | 4520 | 7,856 | 3490 | 1,562 | 2410 | 0,713 |
| Crustacés | 2990 | 1,744 | - | - | 180 | 0,066 | 280 | 0,068 |
| Polychétes | 1540 | 1,000 | - | - | 560 | 0,687 | 950 | 0,745 |
| Chironomides | 170 | 0,010 | - | - | 30 | 0,006 | - | - |
| Divers | 410 | 0,328 | - | - | 20 | 0,019 | - | - |
| Total | 10300 | 7,247 | 4520 | 7,856 | 4760 | 7,470 | 5650 | 4,137 |
| Station | 14 (Rhodia) | 18 (Rhodia) | 19 (Tsoukalio) | 23 (Tsoukalio) | ||||
| Pelecypodes | 790 | 3,253 | 250 | 0,213 | 4230 | 8,328 | 13220 | 19,612 |
| Gastropodes | 240 | 0,057 | - | - | 2080 | 3,903 | 10 | 0,064 |
| Crustacés | 590 | 0,133 | 2620 | 0,734 | 480 | 0,550 | 320 | 0,399 |
| Polychètes | 590 | 0,466 | 490 | 0,716 | 580 | 1,593 | 850 | 1,221 |
| Chironomides | 20 | 0,005 | 60 | 0,007 | - | - | 160 | 0,006 |
| Divers | - | - | 40 | 0,017 | 30 | 0,010 | 140 | 0,004 |
| Total | 2230 | 3,914 | 3460 | 1,687 | 7400 | 14,384 | 14700 | 21,306 |
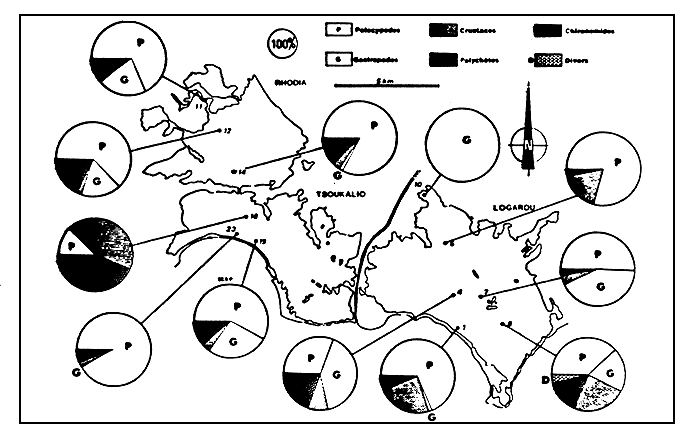
Figure 9
Dominances pondérales des divers groupes taxonomiques.
Ainsi, la lagune de Logarou couvre pratiquement la totalité de l'échelle de confinement du «proche paralique» c'est-à-dire du domaine macrofaunistique (Perthuisot et al., 1983), sans atteindre le paralique lointain évaporitique. Il en va de même pour l'ensemble Tsoukalio-Rhodia, sauf à son extrémité septentrionale où l'on touche presque au paralique lointain dulçaquicole.
L'étude des densités et des biomasses montre également l'établissement de gradients de même sens que ceux de la richesse spécifique et qui aboutissent aux mêmes conclusions.
Les valeurs de biomasse situent ces lagunes en position moyenne entre les étangs très productifs comme l'étang du Prévost (Guélorget, Michel, 1976) et les lagunes plus pauvres comme la Bahiret el Biban (Guélorget et al., 1982).
L'analyse de l'indice de diversité de Shannon (fig. 10) conforte les conclusions tirées des paramètres qualitatifs et quantitatifs précédemment évoqués.
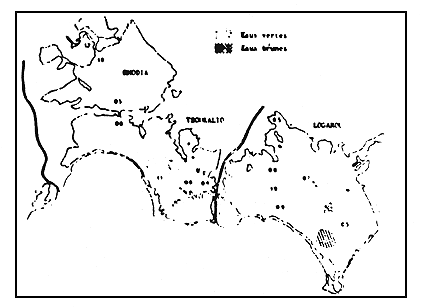
L'ensemble de ces données permet de dresser une carte des zones de confinement dans le complexe paralique du delta du Louros et de l'Arachtos (fig. 11):
Classification of taxonomic groups by weight.
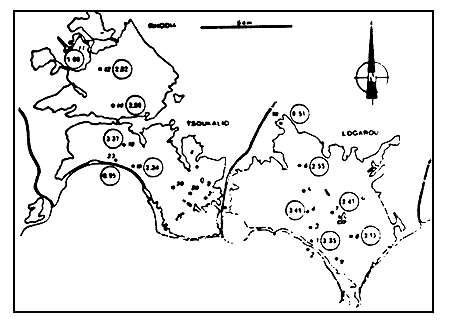
Figure 10
Valeurs de l'indice de diversité de Shannon de la macrofaune
benthique.
Valeues of Shannon's diversity index for benthic macrofauna.
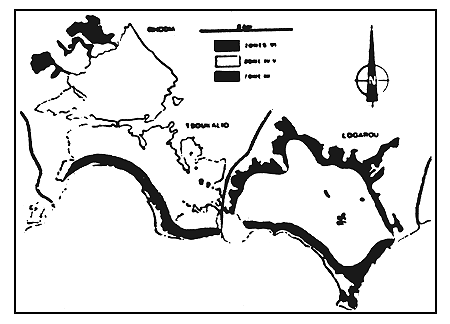
Figure 11
La zonation biologique (zones de confinement) dans le complexe
lagunaire.
Biological zonation (confinement zones) of the lagoonal complex.
Les sédiments actuels
Les sédiments actuels du complexe lagunaire ont été prélevés en 11 points. Les analyses ont porté sur leur composition minéralogique et leur teneur en matière organique.
Les éléments terrigènes sont essentiellement constitués de quartz, de feldspaths et d'argile en faible proportion. Les classes granulométriques vont des sables fins en bordure de la baie aux vases fines des confins lagunaires.
Les carbonates, tous calciques à faiblement magnésiens, sont d'origine locale, biodétritiques, chimiques ou biochimiques.
Seule la fraction carbonatée de la matrice des sédiments (fraction inférieure à 40μ) a fait l'objet de dosages quantitatifs (fig.12). La calcite très faiblement magnésienne, est sans doute pro parte d'origine détritique, mais aussi biodétritique. La calcite magnésienne est généralement d'origine biodétritique et provient de la désagrégation mécanique de tests de mollusques, mais peut aussi provenir de l'activité bactérienne. L'aragonite est le produit à la fois de l'activité biologique du milieu et de son activité chimique. Ainsi, à Logarou, les relativement fortes teneurs en aragonite des confins lagunaires sont vraisemblablement liées à la forte évaporation et à l'élévation de température qui se produisent à la belle saison. L'anomalie positive de la station 8 est sans doute à mettre sur le compte d'un mélange d'eaux confinées et d'eaux en fraiche provenance de la mer. Un tel processus a déjà été évoqué dans le cas d'une lagune tunisienne sursalée (Medhioub. Perthuisot, (1981).
Dans le cas de Rhodia, les trés faibles teneurs en aragonite sont vraisemblablement à mettre en relation avec une transition précoce de l'aragonite en calcite sous l'effet de la dilution du milieu par des eaux continentales par ailleurs calciques.
Le taux de matière organique est en général faible dans les sédiments actuels du complexe lagunaire (fig. 13). ce qui apparaît comme une anomalie par rapport aux autres lagunes méditerranéennes, mais s'explique par la modération de la production biologique globale de ces bassins. Cependant, d'une façon générale, la teneur en matière organique augmente vers les confins lagunaires. où la production biologique est relativement plus élevée et la conservation meilleure. En outre, il peut y avoir des apports d'origine continentale, comme c'est sans doute le cas dans la partie centrale de Rhodia (station 12).
Enfin, le maximum de matiére organique observé à la station 8 s'explique par la présence en ce point de l'ombilic hydraulique de Logarou et d'une abondante sédimentation de plancton sénescent.
Ainsi, les caractéristiques sédimentologiques du complexe paralique du delta du Louros et de l'Arachtos sont en parfaite concordance avec ses caractéristiques hydrochimiques et biologiques.
CONCLUSION
Le complexe paralique du delta du Louros et de l'Arachtos offre une organisation sédimentologique et biologique conforme au schéma du domaine paralique méditerranéen proposé par Guélorget et Perthuisot (1983).
Toutefois, une des originalités de ce complexe réside dans l'étroitesse des zones de confinement modété où dominent les espèces mixtes; cette caractéristique essentielle est liée à la présence d'un mince lido qui limite les échanges entre la baie et les lagunes adjacentes et induit un confinement brutal, court-circuitant ainsi les zones à confinement modéré les plus productives du domaine paralique.
L'autre originalité de ce complexe est sa division en deux ensembles, l'un s'organisant vers le pôle dulçaquicole, l'autre vers le pôle évaporitique (tout au moins en été). Le premier possède une organisation longitudinale Sud-Nord, le deuxième, plus compliqué, présente une zone d'ombilic hydraulique liée à la rotation des eaux dans le bassin. Cette caractéristique hydrologique se retrouve dans bon nombre de bassins paraliques: étang du Prévost (Guélorget, Michel, 1976), lac Melah en Algérie (Guélorget et al., 1984), Bahiret et Biban (Guélorget et al., 1982), lagune de Guemsah en Égypte (Ibrahim, 1983).
La présence d'un mince lido préserve la baie d'Amvrakia
d'une évolution rapide dans l'échelle de
confinement: elle apparaît ainsi comme
Figure 12
Teneurs en aragonite (en %) de la matrice carbonatée des
sédiments du complexe lagunaire.
Aragonite percentage in the carbonate matrix of present sediments
of the lagoonal complex.
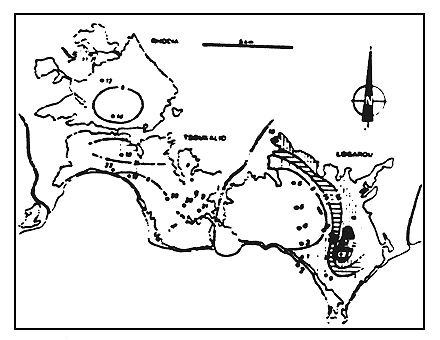
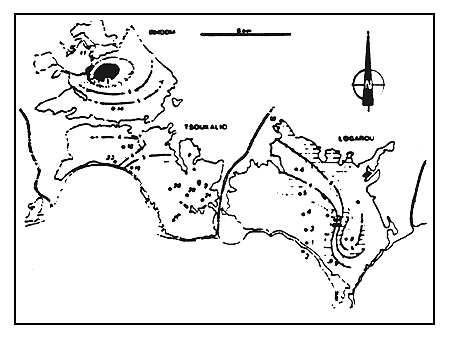
Figure 13
Teneur en matiére organique des sédiments actuels des lagunes
du delta (pourcentages du poids soc).
Organic matter (dry weight) percentage of present sediments
of the lagoonal complex.
RÉFÉRENCES
Aubomin J., 1959. Contribution à l'étude géologique de la Gréce septentrionale: les confins de l'Épire et de la Thessalie, Ann. Geol Pays Hell., 10,526 p.
Galloway W.E., 1975. Process framework for describing the morphologic and stratigraphic evolution of deltaic depositional systems, in: Deltas, edited by M.L. Broussard, Houston Geological Society, 87–98.
Goélorget O., 1985. Entre mer et continent. Contribution à l'étude du domaine paralique, Thése Doct. État, Univ. Sci. Tech. Languedoc, Montpellier, 721 p.
Goélerget O., Michael P., 1976. Recherches écologiques sur une lagune ssumâtre méditerranéenne, l'étang du Prévost (Hérault). I: Le milieu; II: Les peuplements benthiques, Thése 3'cycle, Univ. Sci. Tech. Languedoc, Montpellier, 217 p.
Goélerget O., Perthmisot J.P., 1983. Le domaine paralique. Expressions géologiques, biologiques et économiques du confinement, Trav, Lab. Geol. ENS. 16, 136 p.
Goélerget O., Frisoni G.F., Perthmisot J.P., 1982. Contribution à l'étude biologique de la Bahiret el Biban, lagune du SE tunisien, Mèm. Soc. Géol. Fr., 61, 144, 173–186.
Goélorget O., Frisoni G.F., Ximenes M.-C., Perthuisot J.-P., 1984 a. Diagnose écologique du lac. Melah (Algérie) pour l'évaluation de ses potentialités halieutiques et aquacoles, Rapp. FAO, 130 p.
Goélorget O., Frisoni G.-F., Ibrahim A., Jauzeia A., Medhioub K., Clastère J., Mauria A., Reuchy J.-M., Perthoisot J.-P., 1984 b., Ombilics et chenaux, zones singulières des bassins paraliques. Leurs caractéristiques biologiques et aédimentaires, 5. Congr. Eur. Sédim., Marseille, 202–203.
Ibrahim A., 1983. Contribution à l'étude hydrochimique, biologique et sédimentologique de la lagune de Guemsah (Égypte), Thése Univ., P. et M. Curie, Paris, 126 p.)
Medhioub K., Perthuisot J.-P., 1981. The influence of peripheral sabkhas on the geochemistry and sedimentology of a tunisian lagoon Bahiret el Biban, Sedimentology, 28, 679–688.
Perthuisot J.-P., Goélorget O., Frisoni G.-F., Mauria A., 1983. Expressions hydrochimiques, sédimentologiques et biologiques du gradient de confinement dans un partènement salinier abandonné: le Triangle des Bermudas (Santo Pola, Espagne), J. Rech. Oceanogr., 8, 1. 38–56.
Sriji J., 1981. Ecological assessment of the delta area of the rivers Louros and Arachtos of the Gulf of Amvrakia. Trav. Univ. Essen. 2 vol. 346 p.
![]()
![]()