![]()
![]()
![]()
Lake Malawi is the southernmost and third largest of the East African Great Lakes. It is about 560km long and has a greatest width of about 75km. Maximum depth is about 700m. The lake lies between 9°30'S and 14°30'S, 33°50'E and 35°20'E at an altitude of about 500m in a tropical climate. However it lies far enough south of the equator to experience marked seasonal variations in wind, temperature, and precipitation (Eccles 1974). It is to a large extent delimited by geological faults, particularly to the north and on the eastern coast. As a result these shores are steep and depths in excess of 200m are found close inshore. At the southern extremely and along the southern half of the west coast the shoreline is more gently shelving (Eccles 1974).
Lake Malombe which is situated between 14°30'S and 15°S, 35°E and 35°30'E, is an impoundment of the outflow from Lake Malawi via the Upper Shire river. It is 30km long and has a maximum width of 15km. The depth averages about 7m with a maximum of about 17m. It has the same climate regime as the southern part of Lake Malawi.
Lake Malawi is oligotrophic and the production of phytoplankton which is the base of the food chain is limited by the exhaustion of nutrients which are mostly lost to an almost permanent stratified water layer at a depth below 250m. This layer is anoxic (Eccles, 1962).
During the summer months (December to March) in Lake Malawi, a stable thermocline occurs between 50m and 100m (see Figure 3.1). High temperatures and the absence of strong or undirectional winds enhance the stability of the thermocline, which persists through April. Seasonal southeasterly trade winds (Mwera) begin to blow in late April or early May, marking the onset of the dry and cool winter season (May to August). Air temperatures decline, humidity decreases and the trade winds blow fairly continuously, often with considerable force. Local topography channels these winds along the main axis of the lake. Under the force of persistent winds and lower air temperatures, heat is lost from the lake, the thermocline is driven to 250m and the mixolimnion becomes (nearly) isothermal. Nutrients are recirculated into the euphotic zone (Harding, 1963) and phytoplankton (predominantly diatoms) bloom (Schmidle, 1904; Talling 1969; Degnbol and Mapila, 1982). In shallower southern portions of the lake, winter mixing drives the thermocline to the bottom, and the entire water column mixes throughout this region of the lake. The seasonal trade winds decline in strength by September, accompanying an increase in air temperatures. Under these conditions, the mixolimnion begins to restratify. In October, November and December, locally strong winds, associated with thunderstorms that mark the onset of the rainy season, produce temporary and localized mixing and a period of weak stratification. By late December, even these localized winds become infrequent. High temperatures encourage the formation of a stable thermocline, completing the annual cycle (Twombly, 1983).
Rainfall contributes trivially to the total water balance of the lake and has little influence on cycles of stratification, mixing or the redistribution of nutrients. Surface runoff associated with rains from December to April does not provide the lake with a nutrient supply sufficient to produce blooms in phytoplankton populations (Twombly, 1983).
An internal wave (seiche) that varies seasonally in period and amplitude, local upwelling and profile-bound density currents complicate seasonal patterns of mixing and stratification in Lake Malawi (Mortimer, 1950; Eccles, 1962, 1974).
Lake Malombe is polymictic (fully mixed); nutrients are well recycled and the productive zone extends to the bottom. It is believed that rainfall runoff contributes significantly to the productivity of the lake.
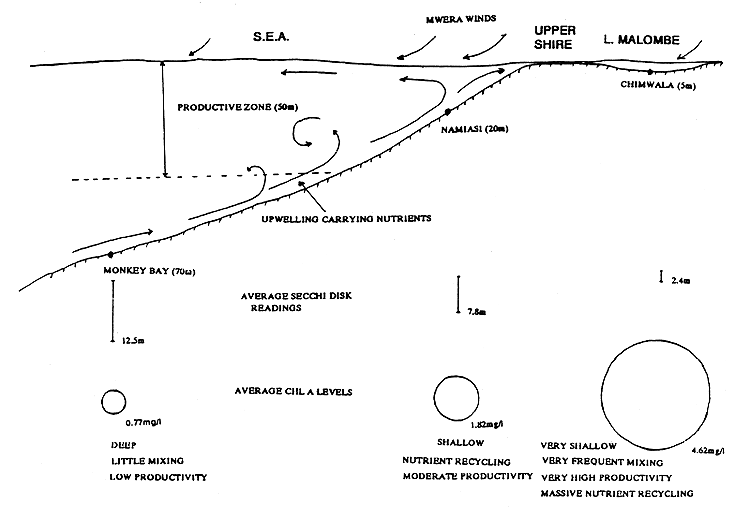
The south-east arm of Lake Malawi supports a much richer fishery than any other area. This is largely due to the upwelling of nutrient rich water under the influence of the strong southerly winds. The existence of internal waves also help in the mixing of the water and hence to maintain the fertility of the south east arm where nutrients sedimented to the bottom are directly returned to the water column (Eccles, 1962).
The trophic gradient of the south-east arm and Lake Malombe was investigated by monthly sampling from March 1991 to March 1992.
Two sampling stations were established in the south-east arm: Monkey Bay (at 60m bottom depth) and Namiasi (at 20m), and one station, Chimwala (4m), in Lake Malombe. Integrated water samples were collected weekly at Monkey Bay (to depth of 40m) and fortnightly at both Namiasi (18m) and Chimwala (4m), filtered through 0.45 μm glass-fibre filter and was later analysed for chlorophyll a. Transparency was determined with a Secchi disk.
Secchi disk readings indicate very low productivity at Monkey Bay station (mean = 12.5m), moderate productivity at the shallower and more mixed Namiasi (7.75m) and very high productivity at the much shallower Chimwala (Malombe) station (2.4m). Mean chlorophyll a values are highly correlated to the Secchi disk readings and show the same trend of productivity: Monkey Bay, 0.77mg/l; Namiasi, 1.82mg/l; and Chimwala, 4.62mg/l.
The limnology of Lake Malombe has not been previously investigated, however from the chlorophyll a and Secchi disk (indicators of productivity) measurements it is evident that it is much richer than the south-east arm (Figure 3.2)
Figure 3.1 Lake Malawi profile, illustrating stratification and mixing.
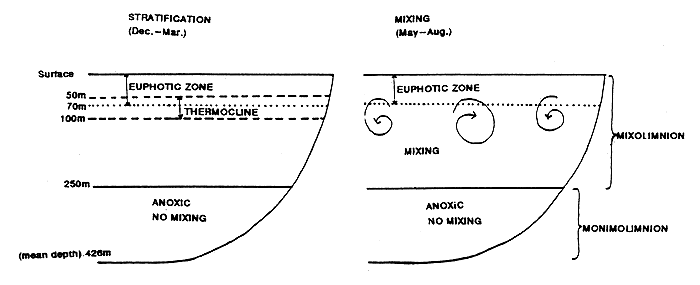
Figure 3.2 Schematic profile of project area.
Chambo feed both on plankton and benthos (Turner, et al. 1991 a) hence a study was conducted to elucidate the plankton and benthic composition. Only preliminary plankton results will be presented and benthic samples are yet to be analysed.
Integrated phytoplankton samples were collected at each station (concurrent with the chlorophyll a samples). Zooplankton samples were collected using an 80 μm mesh net. Phytoplankton samples were preserved in 2% formalin and 1% Lugol's solution and zooplankton in 5% formalin. Phytoplankton samples were settled in a settling chamber for 48 hours, counts and measurements of algae cells was carried out on an inverted microscope.
The phytoplankton (Table 3.1) at Monkey Bay is dominated by Aulacoseira, Surirella, Stephanodiscus and Mougeotia whereas the Namiasi plankton is almost exclusively Aulacoseira. Surirella comprises almost 50% of the phytoplankton for Lake Malombe followed by Aulacoseira and Closterium. Monkey Bay and Namiasi zooplankton (Table 3.1) is dominated by large copepods (Diaptomus and Mesocyclops), whereas Lake Malombe zooplankton is mostly composed of the small cladoceran Bosmina.
Table 3.1 Percent volume phytoplankton and zooplankton composition.
| a. Phytoplankton | Monkey Bay % | Namiasi % | Lake Malombe % |
| Aulacoseira | 43.3 | 93.0 | 34.1 |
| Surirella | 20.1 | 2.7 | 48.7 |
| Stephanodiscus | 18.7 | 3.2 | - |
| Mougeotia | 15.8 | - | - |
| Cymatopleura | 2.0 | - | - |
| Closterium | - | 1.0 | 12.6 |
| Synedra | - | - | 2.8 |
| Staurastrum | - | - | 1.8 |
| b. Zooplankton | % | % | % |
| Diaptomus | 58.8 | 44.4 | 4.3 |
| Mesocyclops | 19.0 | 22.8 | 39.4 |
| Diaphanosoma | 12.9 | 12.1 | 4.4 |
| Nauplii | 8.8 | 14.8 | 3.5 |
| Bosmina | 0.5 | 6.0 | 48.4 |
![]()
![]()
![]()