![]()
![]()
![]()
CONTRIBUCION AL CONOCIMIENTO DE LOS PARAMETROS BIOLOGICOS DE LA GAMBA BLANCA (Parapenaeus longirostris Lucas, 1846) de la dívísión 34.1.1. de COPACO.
Ignacio Sobrino y Antonio Cervantes
Centro Oceanografico de Fuengirola
1. INTRODUCCION
La gamba blanca (Parapenaeus longirostris Lucas, 1846) constituye uno de los objetivos de la flota de arrastre mixto que faena al fresco en el Atlántico marroqui al norte de Cabo Ghir.
Esta especie se dístribuye por todo el Atlántico Centro Oriental a profundidades que oscilan entre los 50 y los 500 metros, situándose su máximo de abundancia entre los 200 y 300 metros, (FAO. 1986).
El presente trabajo constituye una aportación al conocimiento de los parámetros biológicos de la gamba blanca y los aspectos ligados a su reproducción.
2. MATERIAL Y METODOS
Durante el año 1988 se han tomado muestras procedentes del Atlántico marroqui con frecuencia mensual (excepto en enero y febrero) en los puertos de Algeciras, Málaga y Pto. de Sta. Maria, estando constituida la muestra total anual por 2850 indíviduos. De ellos 895 fueron machos y 1945 hembras. Los datos que se tomaron de cada ejemplar fueron los siguientes: longitud total al milimetro, longitud del cefalotorax a la décima de milímetro, peso a la centésima de gramo y sexo. Asimismo en las hembras se tomó el grado de madurez, en base a una escala “de visu” que comprende tres estados:
I. Virgen/Reposo
II. Desarrollo
III. Puesta
Mediante el tratamiento de estos datos se ha procedido a establecer las siguientes estimaciones:
2.1. Ecuación longitud total - peso:
A partir de los pares de valores de longitud total y peso se ha calculado una expresión del tipo exponencial:
Peso = A* (longitud total)^B
2.2. Ecuación longitud del cefalotorax - peso:
Al igual que en la ocuación longitud total – peso la expresión exponencial será de la forma:
Peso = A*(longioitud del cefalotorax)^B
2.3. Sex - : ratio
A partir de los ejcmplares mupstreados se calcula el % de machos y hembras por intervalo de 0.5 cms de longitud total.
2.4. Epoca de puesta
A partir de los porcentajes de hembras en estado I, II y III se compone una gráfica, cuyos máximos se corresponden con los picos de puesta de la especie.
3. RESULTADOS Y DISCUSION
3.1. Ecuación longitud total - peso
La Tabla I recoge los valores de los parámetros de esta ecuacíón para machos y hembras por separado, incluyéndose igualmente los propuestos por Cervantes y Goñ (1986) para la misma zona.
En las figuras 1 y 2 se representan gráficamente ambas expresiones para machos y hembras respectivamente, observándose algunas diferencias que probablemente respondan a la mejora en la medida del peso al utilizarse para el presente trabajo una balanza de precisión.
3.2. Ecuación longitud del cefalotórax - peso
En la tabla II figuran los parámetros resultantes para esta ecuación además de los obtenidos por Ribeiro et al (1984) en la costa sur de Portugal y por Cervantes y Goñ (1986) en el Atlántico marroqui.
En las figuras 3 y 4 se reflejan las representaciones gráficas de las ecuaciones obtenidas por estos autores para machos y hembras respectivamente. Si bien en el caso de las hiifibras las representaciones son muy similaras, en la correspondiente a los machos propuesta por Cervantes y Goñi (1986) se observan valores menorns del peso para la misma longitud del cefalotórax.
3.3. Sex - ratio
En la tabla III se muestran las sex ratio para cada trimestre y el total anual. Se puede observar en ella como en los dos primeros trimestres se igualan los porcentajes de machos y hembras a partir de los 12 cm; sin embargo, en los trimestres tercero y cuarto los porcentajes se mantienen igualados desde tallas menores. Por otra parte, a partir de los 14 cm de longitud total no aparecen machos lo que constata un crecimíento diferente entre los sexos.
3.4. Epoca de puesta
En la tabla IV se presentan los porcentajes de hembras en estado I, II y III encontrados en el periodo que abarca de Marzo a Diciembre del año 1988.
La representacíón gráfica de estos valores (figura 5 ) muestra una puesta prolongada a lo largo del período estudiado con dos picos en los meses de Junio y Octubre, coincidiendo con la disminución de individuos en estado I.
4. BIBLIOGRAFIA
Arrobas, I., & A. R. Cascalho, 1982. Some aspects of the fishery and biology of P. longirostris (Lucas, 1846) of south Portuguese coast. ICES, C.M. 1982/K:5.
Burukowsky, R.N., 1970. Certains problèmes de l'oogenèse de la crevette rose (P. duorarum). Archives de l'Anatomie Histologie et Embryologie 63(6): 56–66.
Cervantes A. y R. Goñi, 1986. Resumen de los datos de base y parámetros biológicos de la pesquería de gamba en la división 34.1.1. de CECAF.COPACE/PACE SERIES 86/33 : 157–164.
Cervantes, A. y R. Goñi, 1986. Composición por tallas de la captura española de Gamba blanca (Parapenaeus longirostris Lucas, 1846) en el área de CECAF. División 34.1.1. COPACE/PACE SERIES 86/33, pp 240–245.
Establier, R., 1963. Composición química de la gamba (P. longirostris) del Golfo de Cádiz y Marruecos y su variacón estacional. Inv. Pesq. 23: 159–167.
FAO, 1986. Rapport du premier groupe de travail spécial sur les pêcheries de merlus et crevettes profondes dans la zone nord du COPACE. COPACE/PACE SERIES 86/33.
Heldt, J.H., 1954. Les fonds à Parapenaeus longirostris (Lucas, 1846) des mers tunisiennes. Bull. Stat. Ocean. de Salambo, 47: 11–22.
Karlovac, O., 1949. Le Parapenaeus longirostris (Lucas, 1846) de la Haute Adriatique. Acta Adriatica, 3(12): 3–12 (407–416).
Mircenda, A., 1933. Ensayo de un catálogo de los crustáceos decápodos marinos en España y Marruecos Español. Notas y r–esúmenes. Inst. Esp. Ocean. Serie II, no 67, 72 pp.
Ribeiro - Cascalho, A. & I. Arrobas, 1984. Parapenaeus longirostris (Lucas, 1846) from south Portuguese coast: data on its fishery and observations on some biological caracteristics. ICES Shellfish Committee C.M. 1984/K: 29.
Ribeiro - Cascalho, A., 1985. About two trawl surveys on the south Portuguese coast shrimp grounds. ICES. Shellfish Committee.
Vedavysa Rao, P. 1968. Maturation and spawning of the penaeid prawns of the south west coast of India. FAO Fish. Rep. No 57, Vol. 2, pp.: 285–302.
| 10 TRIMESTRE | 20TRIMESTRE | 30 TRIMESTRE | 410 TRIMESTRE | TOTAL AAO 1988 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TOTAL | TOTAL | TOTAL | TOTAL | PORCEIAJES | ||||||
| TAIIA | M | H | M | H | M | H | M | H | M | H |
| 5.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 5.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 6.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 100 |
| 6.5 | 0 | 0 | 0 | 0 | 0 | 0 | 50 | 50 | 50 | 50 |
| 7.0 | 100 | 0 | 0 | 0 | 0 | 0 | 95 | 25 | 76 | 24 |
| 7.5 | 100 | 0 | 0 | 0 | 59 | 41 | 43 | 57 | 44 | 56 |
| 8.0 | 85 | 15 | 100 | 0 | 0 | 100 | 66 | 34 | 57 | 33 |
| 8.5 | 80 | 20 | 99 | 1 | 69 | 31 | 54 | 46 | 57 | 48 |
| 9.0 | 100 | 0 | 98 | 2 | 54 | 46 | 57 | 53 | 55 | 15 |
| 9.5 | 100 | 0 | 98 | 2 | 45 | 55 | 21 | 79 | 47 | 53 |
| 10.0 | 97 | 3 | 94 | 6 | 64 | 36 | 10 | 90 | 44 | 51 |
| 10.5 | 99 | 1 | 89 | 11 | 68 | 32 | 17 | 83 | 58 | 48 |
| 11.0 | 97 | 3 | 93 | 7 | 50 | 50 | 32 | 68 | 63 | 17 |
| 11.5 | 87 | 13 | 78 | 22 | 29 | 71 | 30 | 70 | 53 | 47 |
| 12.0 | 44 | 51 | 46 | 54 | 15 | 85 | 24 | 71 | 32 | 58 |
| 12.5 | 36 | 64 | 25 | 75 | 4 | 96 | 6 | 94 | 17 | 83 |
| 13.0 | 7 | 93 | 16 | 84 | 2 | 98 | 3 | 97 | 9 | 91 |
| 13.5 | 21 | 77 | 9 | 91 | 5 | 95 | 1 | 99 | 8 | 98 |
| 14.0 | 0 | 100 | 11 | 89 | 9 | 91 | 1 | 99 | 6 | 98 |
| 14.5 | 0 | 100 | 15 | 85 | 0 | 100 | 0 | 100 | 4 | 96 |
| 15.0 | 0 | 100 | 0 | 100 | 0 | 100 | 0 | 100 | 0 | 100 |
| 15.5 | 0 | 100 | 0 | 100 | 0 | 100 | 0 | 100 | 0 | 100 |
| 16.0 | 0 | 100 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 100 |
| 16.5 | 0 | 100 | 0 | 100 | 0 | 100 | 0 | 100 | 0 | 100 |
| 17.0 | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 100 |
| 17.5 | 0 | 0 | 0 | 100 | 0 | 100 | 0 | 0 | 0 | 100 |
| 18.0 | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 100 |
| 18.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 19.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 19.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 20.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 20.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 21.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 21.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 22.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 22.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 23.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 23.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 24.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 24.5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 25.0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| AUTORES | ZONA | A | B | R | N | |
|---|---|---|---|---|---|---|
| Crosnier (1970) | Congo | M | 1.88*10^-5 | 2,74 | 0,99 | 157 |
| M | 1.1*10^-5 | 2,85 | 0,99 | 187 | ||
| Cervantes y Gohi (1986) | Altantico aarroqul | M | 9.42*10^-3 | 2,66 | 0,85 | 561 |
| M | 4,67*10^3 | 2,99 | 0,97 | 1119 | ||
| Sobrina y Cervantes (1989) | Altantico arrouqui | M | 6.7*10^-3 | 2,9 | 0,92 | 153 |
| H | 1.03*10^-2 | 2,83 | 0,92 | 537 |
| AUTORES | ZONA | A | B | R | N | |
|---|---|---|---|---|---|---|
| Ribeiro (1984) | Portuqal | M | 7.5 10-3 | 2,19 | 0,99 | 111 |
| H | 5.5*10^-3 | 2,31 | 0,99 | 315 | ||
| Cervantes y Goñi (1986) | Atlántico arroqui | M | 5.61*10^-3 | 2,205 | 0,74 | 599 |
| H | 3.44*10^-3 | 2,42 | 0,9 | 1129 | ||
| Sobrino y Cervantes (1989) | Atlántico arroqui | M | 2.15*10^-3 | 2,56 | 0,95 | 153 |
| H | 9.4*10^-3 | 2,12 | 0,84 | 555 |
| 10 TRIMESTRE | 20TRIMESTRE | 30 TRIMESTRE | 410 TRIMESTRE | TOTAL AAO 1988 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TALLAS (cm) | M | H | M | H | M | H | M | H | M | H |
| 5,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 | 0,00 |
| 5,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 | 0,00 |
| 6,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 100,0 | 0,00 | 100,00 |
| 6,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 50,0 | 50,0 | 50,0 | 50,00 |
| 7,0 | 100,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 75,0 | 25,0 | 75,60 | 24,40 |
| 7,5 | 100,0 | 0,0 | 0,0 | 0,0 | 58,5 | 41,5 | 42,8 | 57,2 | 43,78 | 56,22 |
| 8,0 | 85,1 | 14,9 | 100,0 | 0,0 | 0,0 | 100,0 | 65,9 | 34,1 | 66,75 | 33,25 |
| 8,5 | 80,0 | 20,0 | 98,9 | 1,1 | 68,7 | 31,3 | 53,8 | 46,2 | 57,19 | 42,81 |
| 9,0 | 100,0 | 0,0 | 97,7 | 2,3 | 54,0 | 46,0 | 47,0 | 53,0 | 54,75 | 45,25 |
| 9,5 | 100,0 | 0,0 | 97,6 | 2,4 | 45,4 | 54,6 | 20,7 | 79,3 | 46,84 | 53,16 |
| 10,0 | 97,0 | 3,0 | 93,5 | 6,5 | 64,2 | 35,8 | 10,4 | 89,6 | 49,46 | 50,54 |
| 10,5 | 98,9 | 1,1 | 89,0 | 11,0 | 67,6 | 32,4 | 16,9 | 83,1 | 58,29 | 41,71 |
| 11,0 | 96,9 | 3,1 | 92,5 | 7,5 | 50,0 | 50,0 | 32,2 | 67,8 | 63,26 | 36,74 |
| 11,5 | 87,0 | 13,0 | 78,2 | 21,8 | 28,6 | 71,4 | 29,9 | 70,1 | 53,30 | 46,70 |
| 12,0 | 48,7 | 51,3 | 46,4 | 53,6 | 15,0 | 85,0 | 29,1 | 70,9 | 31,80 | 68,20 |
| 12,5 | 36,1 | 63,9 | 25,3 | 74,7 | 3,5 | 96,5 | 5,8 | 94,2 | 17,10 | 82,90 |
| 13,0 | 7,2 | 92,8 | 16,1 | 83,9 | 2,3 | 97,7 | 3,0 | 97,0 | 8,62 | 91,38 |
| 13,5 | 20,5 | 79,5 | 9,4 | 90,6 | 4,8 | 95,2 | 0,6 | 99,4 | 8,18 | 91,82 |
| 14,0 | 0,0 | 100,0 | 10,7 | 89,3 | 9,3 | 90,7 | 1,1 | 98,9 | 5,54 | 94,46 |
| 14,5 | 0,0 | 100,0 | 15,3 | 84,7 | 0,0 | 100,0 | 0,0 | 100,0 | 4,39 | 95,61 |
| 15,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,00 | 100,00 |
| 15,5 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,00 | 100,00 |
| 16,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 100,00 |
| 16,5 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,00 | 100,00 |
| 17,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 100,0 | 0,0 | 0,0 | 0,00 | 100,00 |
| 17,5 | 0,0 | 0,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 0,0 | 0,00 | 100,00 |
| 18,0 | 0,0 | 0,0 | 0,0 | 100,0 | 0,0 | 100,0 | 0,0 | 0,0 | 0,0 | 100,00 |
| 18,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 19,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 19,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 20,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 20,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 21,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 21,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 22,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 22,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 23,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 23,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 24,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 24,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
| 25,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,00 |
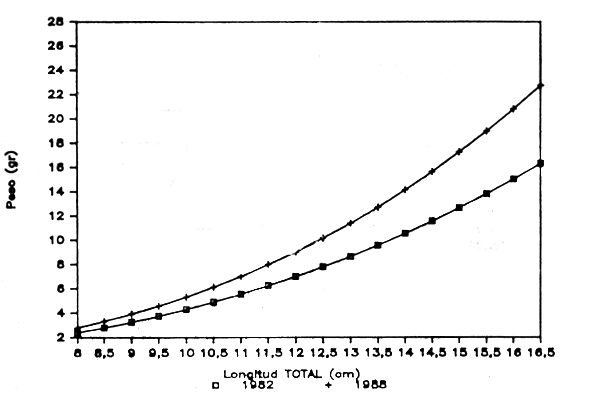
Figura 1.- Reprecentación gráfica de las ecuaciones longitud total-peso obtenidas para la Gamba blanca en 1982 y 1988. División 34.1.1. de COPACE. (MACHOS)
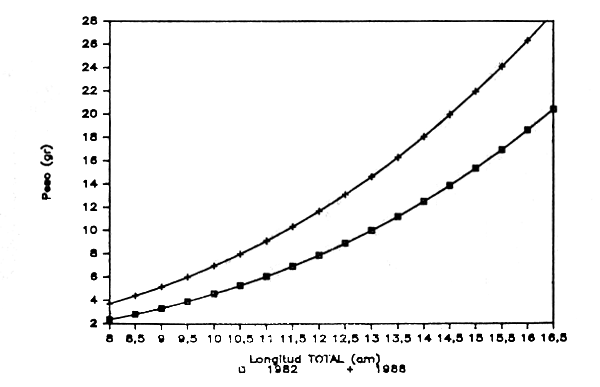
Figura 2.- Reprecentación gráfica de las ecuaciones longitud total-peso obtenidas para la Gamba blanca en 1982 y 1988. División 34.1.1. de COPACE. (HEMBRAS)
| MES | I | II | III | Total |
|---|---|---|---|---|
| 3 | 61,03896 | 37,01298 | 1,948051 | 100 |
| 4 | 51,61774 | 41,91689 | 6,465355 | 100 |
| 5 | 42,19653 | 46,82080 | 10,98265 | 100 |
| 6 | 35,02304 | 39,40092 | 25,57603 | 100 |
| 7 | 69,00958 | 23,64217 | 7,348242 | 100 |
| 8 | 55,79196 | 29,31442 | 14,89361 | 100 |
| 9 | 51,68539 | 38,20224 | 10,11235 | 100 |
| 10 | 46,23655 | 30,10752 | 23,65591 | 100 |
| 11 | 60,11904 | 32,14285 | 7,738095 | 100 |
| 12 | 30,12048 | 60,24096 | 9,638554 | 100 |
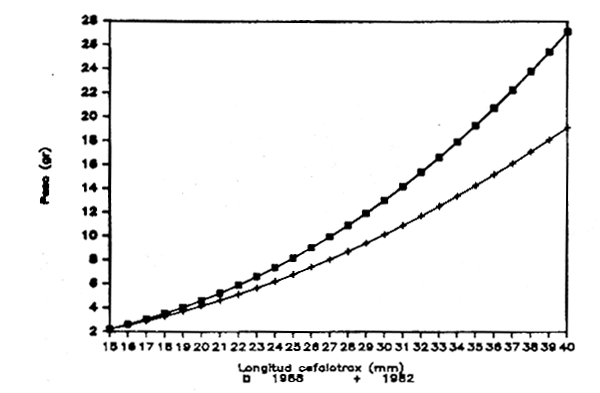
Figura 3.- Reprecentación gráfica de las ecuaciones longitud del cefalotórax-peso obtenidas para la Gamba blanca en 1982 y 1988. División 34.1.1. de COPACE. (MACHOS)
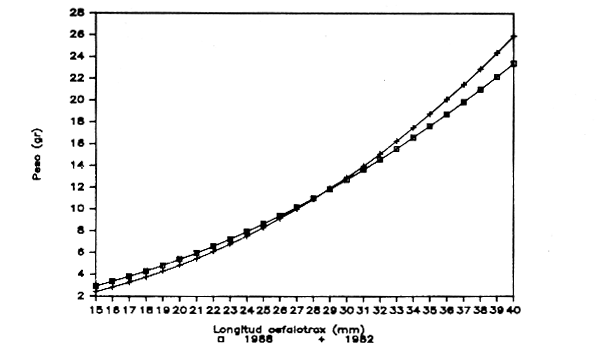
Figura 4.- Reprecentación gráfica de las ecuaciones longitud del cefalotórax-peso obtenidas para la Gamba blanca en 1982 y 1988. División 34.1.1. de COPACE. (HEMBRAS)
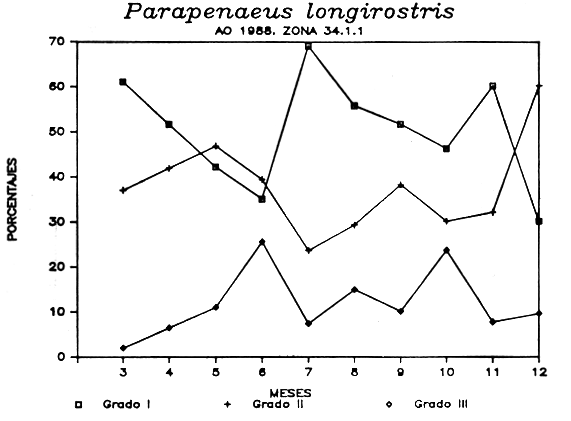
Figura 5.- Evolución mensual de los porcentajes de los diferentes estados de madurez de la Gamba blanca. Año 1988. División 34.1.1. de Copace.
![]()
![]()
![]()