5.1 Changements sensoriels
5.2 Altérations autolytiques
5.3 Changements bactériologiques
5.4 Oxydation et hydrolyse des lipides
Les changements sensoriels sont ceux perçus par les sens, c’est-à-dire: apparence, odeur, texture et goût.
Changements dans le poisson frais cru
Les premières modifications sensorielles du poisson pendant le stockage concernent l’apparence et la texture. Le goût caractéristique des espèces se développe normalement pendant les deux premiers jours de la conservation sous glace.
Le changement le plus important est l’établissement de la rigor mortis. Immédiatement après la mort, le muscle est totalement détendu et la texture élastique et souple dure habituellement quelques heures, après quoi le muscle se contracte. Quand il durcit, le corps se raidit et le poisson est alors en état de rigor mortis. Cet état dure habituellement un jour ou plus et alors la rigor disparaît, ce qui détend le muscle à nouveau et le rend souple mais il n’est plus aussi élastique qu’avant la rigor. Le rapport entre l’apparition et la disparition de la rigor varie d’une espèce à l’autre et est affecté par la température, la manutention, la taille et la condition physique du poisson (Tableau 5.1).
L’effet de la température sur la rigor n’est pas uniforme. Dans le cas du cabillaud, une température élevée crée l’apparition rapide d’une rigor mortis très forte. On doit l’éviter car des fortes tensions pendant la rigor peuvent créer des discontinuités, c’est-à-dire des affaiblissements du tissu conjonctif et une rupture du filet.
On admet généralement que l’apparition et la durée de la rigor mortis sont plus rapides à température élevée mais des observations, surtout sur le poisson tropical, ont montré un effet inverse de la température sur l’apparition de la rigor. Il est évident que, dans ces espèces, l’apparition de la rigor est accélérée à 0°C par rapport à 10°C, ce qui correspond bien à une stimulation des changements biochimiques à 0°C (Poulter et al., 1982; Iwamoto et al., 1987). Cependant Abe et Okuma (1991) ont fourni une explication à ce phénomène en démontrant que l’apparition de la rigor mortis chez la carpe (Cyprinus carpio) dépend de la différence entre la température de l’eau et celle du stockage. Quand la différence est importante, le délai entre la mort et l’apparition de la rigor est court et vice versa.
La rigor mortis s’installe immédiatement ou très rapidement après la mort quand le poisson est affamé et que les réserves de glycogène sont épuisées ou si le poisson est fatigué. La méthode utilisée pour étourdir et tuer le poisson influence également l’apparition de la rigor.Si on étourdit et tue le poisson par hypothermie (le poisson est tué par l’eau glacée), la rigor survient très rapidement, alors qu’un coup sur la tête donne un délai de 18 heures (Azam et al., 1990; Proctor et al., 1992).
La signification technologique de la rigor mortis a une importance majeure si le poisson est fileté avant ou pendant la rigor. Pendant la rigor, le corps du poisson sera complètement raide; le rendement du filetage sera très mauvais et une manutention brutale peut produire des déchirures. Si les filets sont levés pre-rigor, le muscle peut se contracter librement et les filets rétréciront à l’apparition de la rigor. Les muscles rouges peuvent se réduire de 52% et les muscles blancs jusqu’à 15% de leur longueur originale (Buttkus, 1963). Si le poisson est cuit pre-rigor, la texture sera très molle et pâteuse. Au contraire, si le poisson est cuit en état de rigor la chair sera dure mais pas sèche. Post-rigor, la chair deviendra ferme, élastique et succulente.
Tableau 5.1 Apparition et durée de la rigor mortis dans différentes espèces de poissons
Espèce |
Condition |
Température (°C) |
Temps écoulé entre la mort du poisson et l’apparition de la rigidité cadavérique (heures) |
Temps écoulé entre la mort du poisson et la fin de
la rigidité cadavérique |
| Cabillaud (Gadus morhua) |
Fatigué |
0 |
2-8 |
20-65 |
| Mérou (Epinephelus malabaricus) |
Au repos |
2 |
2 |
18 |
| Tilapia bleu (Areochromis aureus) |
Fatigué |
0 |
1 |
|
| Tilapia petite, 60g (Tilapia mossambica) |
Au repos |
0-2 |
2-9 |
26,5 |
| Grenadier (Macrourus whitsoni) |
Fatigué |
0 |
<1 |
35-55 |
| Anchois (Engraulis anchoita) |
Fatigué |
0 |
20-30 |
18 |
| Carrelet (Pleuronectes platessa) |
Fatigué |
0 |
7-11 |
54-55 |
| Rascasse du nord (Sebastes spp.) |
Fatigué |
0 |
22 |
120 |
| Lieu noir (Pollachius virens) |
Fatigué |
0 |
18 |
110 |
| Rouget (Sebastes spp.) | Fatigué |
0 |
22 |
120 |
| Carrelet japonais (paralichthys olivaceus) |
0 |
3 |
>72 |
|
| Carpe (Cyprinus carpio) |
Fatigué Au repos |
0 |
8 |
Sources: Hwang et al., 1991; Iwamoto et al., 1987; Korhonen et al., 1990; Nakayama et al., 1992; Nazar et Magar, 1963; Partmann, 1965; Pawar et Magar, 1965; Stroud, 1969; Trucco et al., 1982.
Les poissons entiers et les filets congelés pre-rigor peuvent donner de bons produits s’ils sont soigneusement décongelés à basse température de façon à donner à la rigor mortis le temps de disparaître pendant que le muscle est encore gelé.
L’évaluation sensorielle du poisson frais sur les marchés et aux débarcadères se fait en vérifiant l’aspect, la texture et l’odeur. Les qualités sensorielles pour le poisson sont présentées au Tableau 5.2. La plupart des systèmes d’évaluation se basent sur les modifications qui se produisent pendant le stockage dans la glace fondante. Il faut rappeler que les changements caractéristiques dépendent des méthodes de stockage. L’aspect du poisson stocké au froid sans glace ne change pas autant que celui du poisson sous glace, mais le poisson s’abîme plus vite et un examen de la flaveur du poisson cuit devient nécessaire. De ce fait, une connaissance des conditions temps/température s’avère nécessaire au débarquement.
Les changements sensoriels caractéristiques dans le poisson post mortem varient considérablement suivant l'espèce de poisson et la méthode de stockage. Les directives de la CEE, élaborées à cet effet donnent une description générale pour l’examen de la qualité du poisson comme le montre le Tableau 5.2. L’échelle utilisée varie de 0 à 3, 3 représentant la meilleure qualité.
L’Association des technologistes du poisson d’Europe de l’Ouest a établi un glossaire polyglotte des odeurs et saveurs qui peut également s’avérer très utile pour la terminologie décrivant l’évaluation sensorielle de la fraîcheur du poisson (Howgate et al.,1992) (Annexe C).
Altérations de la qualité gustative
Les critères de qualité pour du poisson réfrigéré pendant le stockage peuvent être déduits d'un examen sensoriel du poisson cuit. Le Tableau 5.2 présente certaines des qualités sensorielles des poissons et coquillages cuits. Le schéma caractéristique de la détérioration du poisson conservé sous glace se caractérise par quatre phases :
| Phase 1 | Le poisson est très frais avec une saveur douce et délicate d’algues. Le goût peut être légèrement métallique. Dans le cabillaud, le merlan, l’églefin et le flétan, la saveur douce atteint son maximum 2 à 3 jours après capture. |
| Phase 2 | Il y a une perte de l’odeur et de la saveur caractéristiques. La chair devient neutre mais sans arrière-goûts. La texture est encore plaisante. |
| Phase 3 | Des signes de détérioration apparaîssent et un certain nombre de substances volatiles à l’odeur désagréable se forment suivant les espèces de poisson et le type d’altération (aérobie, anaréobie). Un des composants volatiles peut être la triméthylamine (TMA) dérivée de la réduction bactérienne de l’oxyde de triméthylamine (OTMA). La TMA a une odeur de poisson caractéristique. Au début de cette phase, l’arrière-goût peut être légèrement aigre, fruité et légèrement amer surtout dans le poisson gras. Pendant les derniers stades se développent des odeurs doucereuses, de chou, ammoniacales, sulfureuses et rances. La texture devient soit molle et aqueuse, soit dure et sèche. |
| Phase 4 | Le poisson peut être considéré comme altéré et putride. |
Tableau 5.2 Echelle de fraîcheur: Règlement du Conseil (EEC) N° 103/76 OJ N° L20 (28 Janvier 1976) (EEC, 1976)
Critères |
||||
Parties du poisson |
Notes |
|||
3 |
2 |
1 |
0 |
|
Apparence |
||||
| Peau | Pigmentation brillante, iridescente; pas de décoloration; mucus transparent, aqueux | Pigmentation brillante mais non luisante; mucus légèrement trouble | Pigmentation en voie de décoloration et terne; mucus laiteux | Pigmentation terne1 ; mucus opaque |
| Œil | Convexe (gonflé); cornée transparente; pupille noire et brillante | Convexe et légèrement enfoncé; cornée légèrement opalescente; pupille noire et terne | Plat; cornée opalescente; pupille opaque | Concave au centre1; cornée laiteuse; pupille grise |
| Branchies | Couleur brillante; pas de mucus | Moins colorées, quelques traces de mucus clair | En voie de décoloration; mucus opaque | Jaunâtres1, mucus laiteux |
| Chair (de l’abdomen) | Bleuâtre, translucide, lisse et brillante; pas de changement de la couleur initiale | Veloutée, cireuse, terne; couleur légèrement altérée | Légèrement opaque | Opaque1 |
| Couleur le long de la colonne vertébrale | Incolore | Légèrement rosée | Rose | Rouge1 |
| Organes | Les reins et résidus d’autres organes devront être rouges, de même que le sang dans l’aorte | Les reins et résidus d’autres organes devront être rouge terne ; sang en voie de décoloration | Les reins, résidus et sang devront être roses | Les reins1, résidus et sang devront être brunâtres |
Etat physique |
||||
| Chair | Ferme et élastique; surface lisse | Moins élastique | Légèrement molle (flasque), moins élastique, cireuse (veloutée) et surface terne | Molle (flasque)1 ; écailles facilement détachables; surface ridée |
| Colonne vertébrale | Se casse au lieu de se détacher | Adhère | Adhère légèrement | N’adhère pas1 |
| Péritoine | Adhère complètement à la chair | Adhère | Adhère légèrement | N’adhère pas1 |
Odeur |
||||
| Branchies, peau, cavité abdominale | Odeur d’algues | Pas de mauvaise odeur ni d’odeur d’algues | Légèrement aigre | Aigre1 |
1 ou tout autre état d’altération plus avancé.
On peut utiliser une échelle numérique pour l’évaluation sensorielle du poisson cuit comme le montre la Figure 5.1. L’échelle est numérotée de 0 à 10, 10 indiquant une fraîcheur absolue, 8 une bonne qualité, 6 un poisson neutre et sans goût et 4 le seuil de rejet. En utilisant l’échelle de cette façon, on obtient une courbe sigmoïde en forme de S, indiquant une dégradation rapide du poisson pendant la première phase, une vitesse moins rapide en phases 2 et 3 devenant finalement élevée quand le poisson est altéré.
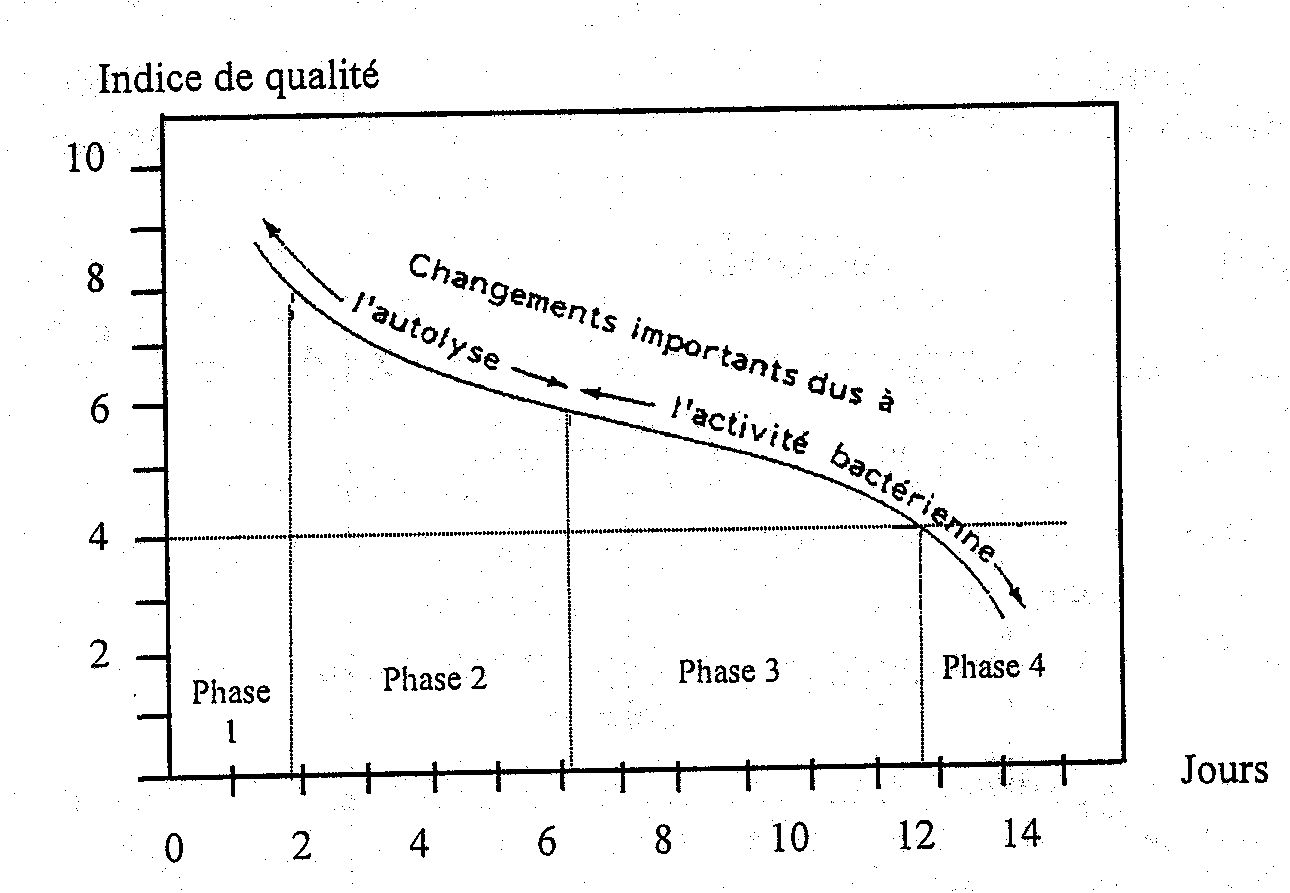
Figure 5.1 Altération des qualités gustatives du cabillaud glacé (0°C) (Huss, 1976)
On peut aussi bien utiliser d’autres échelles qui modifient la forme du graphique. Il est cependant très important de comprendre les résultats attendus de l’analyse sensorielle de façon à poser les questions appropriées au panel sensoriel.
Autolyse signifie "autodigestion". Il a été établi, depuis de nombreuses années, qu’il existe au moins deux types d’altération du poisson : bactérienne et enzymatique. Uchyama et Ehira (1974) ont montré que dans le cabillaud et l’albacore, les changements enzymatiques affectant la fraîcheur du poisson précédaient les changements de qualité microbiologique et étaient sans rapport avec ceux-ci. Dans certaines espèces (encornets, harengs), les altérations enzymatiques précèdent et par conséquent dominent la détérioration du poisson réfrigéré. Chez d’autres espèces, l’autolyse contribue, à des degrés différents, à l'altération de la qualité et s’ajoute au processus de dégradation microbienne .
Production d’énergie post mortem dans le muscle
Au moment de la mort du poisson, l'apport d’oxygène dans le muscle est interrompu du fait que le sang n’est plus pompé par le coeur et ne circule plus au travers des branchies où il est enrichi en oxygène, dans le poisson vivant. Du fait qu’il n’y a plus d’oxygène disponible pour une respiration normale, la production d’énergie à partir d’aliments ingérés est très réduite. La Figure 5.2 illustre la voie normale pour la production d’énergie musculaire dans la plupart des téléostéens vivants (poissons osseux à nageoires). Le glycogène (carbohydrate de réserve) ou la graisse sont oxydés ou "brûlés" par les enzymes des tissus dans une série de réactions qui, en phase ultime, produisent du gaz carbonique (CO2), de l’eau et un composé organique riche en énergie, l’adénosine triphosphate (ATP). Ce type de respiration se réalise en deux étapes: l'une anaréobie et l'autre aérobie. Cette dernière dépend de la présence continue d’oxygène (O2) qui ne peut provenir que du système circulatoire. La plupart des crustacés sont capables de respirer hors de l’élément aquatique en absorbant l’oxygène atmosphérique pendant de courtes périodes.
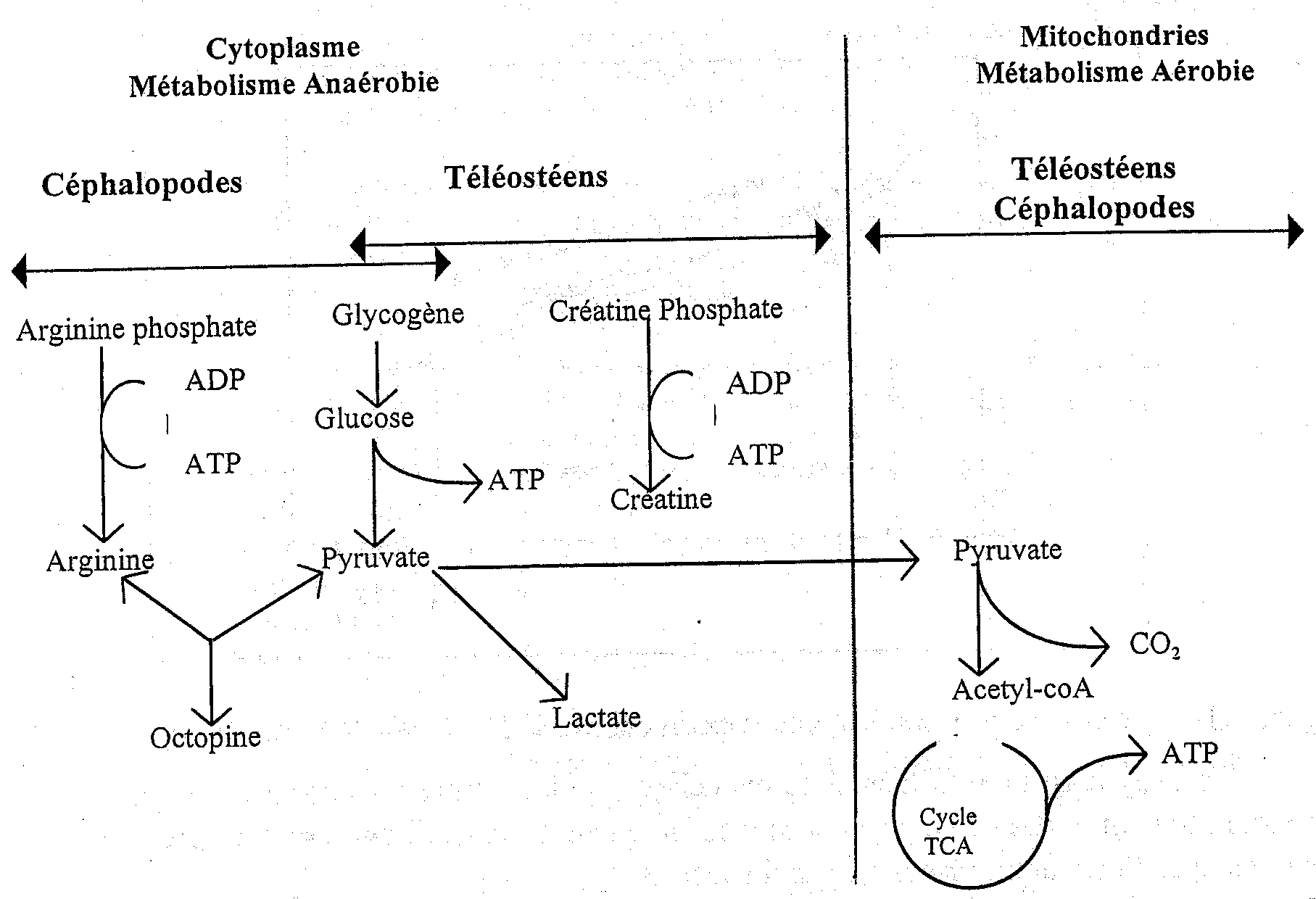
Figure 5.2 Dégradation aérobie et anaréobie du glycogène dans les muscles du poisson
La Figure 5.2 montre également que, dans des conditions d’anaérobiose, l’ATP peut être synthétisée selon deux autres voies importantes à partir de la créatine phosphate ou de l’arginine phosphate. La première source d’énergie est réservée aux muscles des vertébrés (téléostéens) tandis que la dernière est caractéristique de quelques invertébrés comme les céphalopodes (encornets et poulpes). Dans les deux cas, la production d’ATP cesse quand la créatine ou l'arginine phosphates sont épuisés. Il est intéressant de noter que l’octopine est le produit final du métabolisme anaérobie des céphalopodes. Elle n’est pas acide (contrairement au lactate) et, de ce fait, tout changement de pH post mortem dans ces animaux n’est pas relié à la production d’acide lactique à partir du glycogène.
Pour la plupart des poissons téléostéens, la glycolyse est la seule voie possible de production d’énergie après l’arrêt du coeur. Ce procédé particulièrement inefficace a essentiellement comme produits finaux les acides lactique et pyruvique. De plus, la glycolyse produit de l’ATP, mais seulement 2 moles pour chaque mole de glucose oxydé comparée aux 36 moles d’ATP produites pour chaque mole de glucose si les produits finaux glycolytiques sont oxydés en aérobiose dans le mitochondrion de l’animal vivant. Ainsi, après la mort, le muscle anaérobie ne peut pas maintenir son niveau normal d’ATP et, quand le niveau intracellulaire décline de 7 à 10 mmoles/g à £ 1.0mmole/g de tissu, le muscle entre en rigor mortis. La glycolyse post mortem provient de l’accumulation d’acide lactique qui à son tour abaisse le pH du muscle. Dans le cabillaud, le pH diminue de 6,8 à un pH final de 6,1 à 6,5. Chez certaines espèces de poissons, le pH final peut être plus bas: dans les gros maquereaux, le pH final de rigor peut être aussi bas que 5,8-6,0. Il peut même atteindre 5,4-5,6 dans le thon et le flétan. Cependant, des niveaux de pH aussi bas ne sont pas courants dans les téléostéens marins. Ces pH sont rarement aussi bas que ceux observés post mortem dans les muscles des mammifères. Par exemple, le muscle de boeuf atteint souvent des niveaux de pH de 5,1 au momemt de la rigor mortis. La quantité d’acide lactique produite est en rapport avec la quantité de carbohydrate (glycogène) accumulée dans le tissu vivant. En général, le muscle de poisson contient un taux assez faible de glycogène comparé aux mammifères et donc il se forme bien moins d’acide lactique après sa mort. Son état nutritionnel, le stress et la fatigue avant la mort auront également un impact important sur les niveaux du glycogène stocké et par la suite sur le pH final post mortem. En règle générale, un poisson bien reposé et bien nourri contiendra plus de glycogène qu’un poisson épuisé. Une étude récente sur la loche japonaise (Chiba et al.,1991) a montré que quelques minutes seulement de stress avant capture produisaient une baisse de 0,50 unités de pH en 3 heures alors que, dans un poisson qui ne s’est pas débattu, le pH diminuait uniquement de 0,10 unités dans le même temps. Les mêmes auteurs ont également montré que la saignée du poisson réduisait de façon significative la production post mortem d’acide lactique.
La réduction du pH post mortem du muscle du poisson affecte les propriétés physiques du muscle. Quand le pH chute, la charge nette des protéines musculaires est réduite, causant leur dénaturation partielle et la perte d’une partie de leur capacité de rétention d’eau. Le tissu musculaire, en état de rigor mortis, perd son humidité à la cuisson et est particulièrement inapte à un traitement ultérieur par la chaleur, du fait de sa dénaturation par la chaleur qui augmente la perte d’eau. Celle-ci a un effet néfaste sur la texture du muscle du poisson et Love (1975) a montré qu’il y a un rapport inverse entre la dureté du muscle et le pH, des niveaux de dureté inacceptables (accompagnés de pertes d’eau à la cuisson) se produisant à des niveaux très faibles de pH (Figure 5.3).
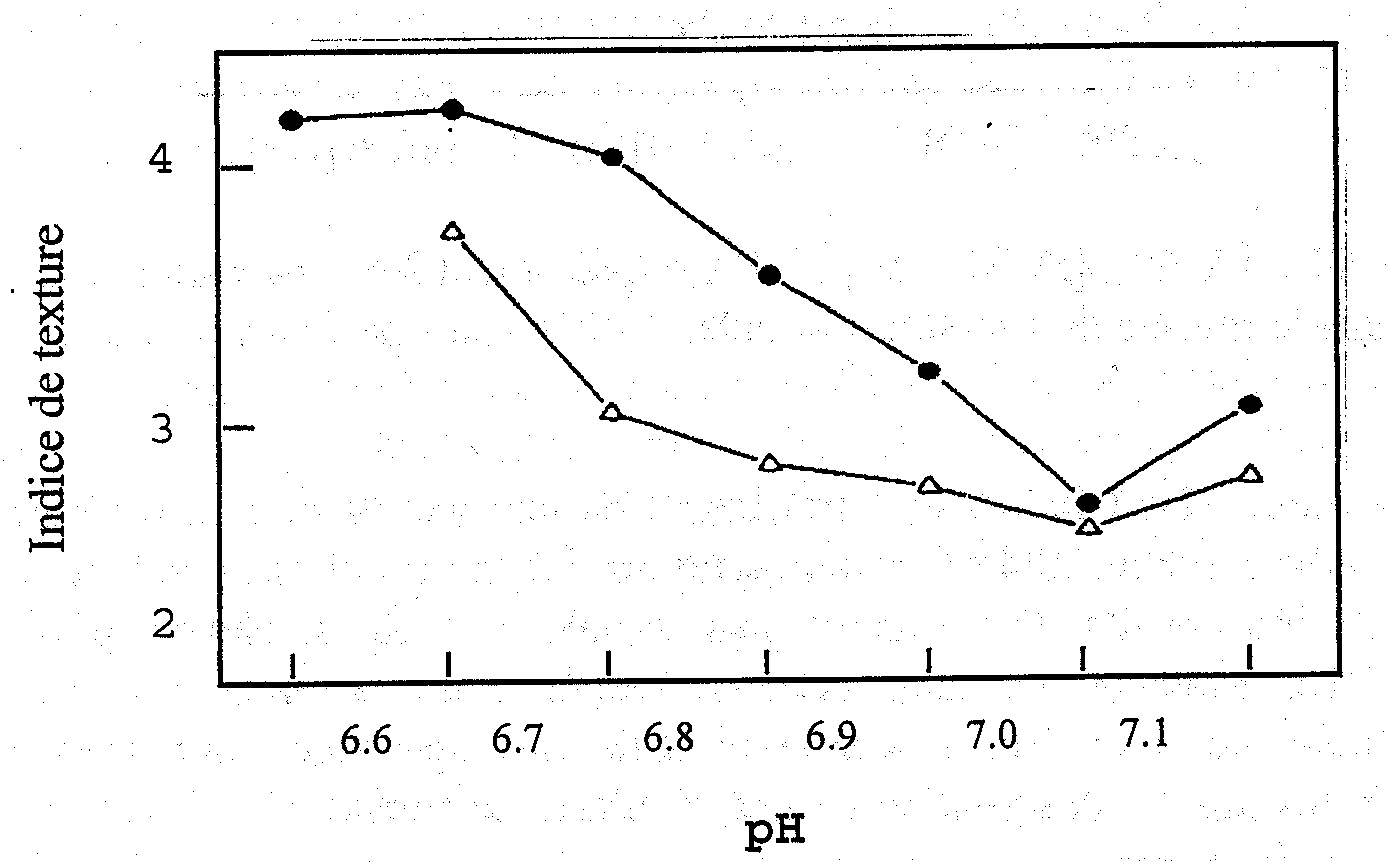
Figure 5.3 Relation entre la texture du muscle de cabillaud et le pH, selon Love (1975). Les points noirs se rapportent à du poisson pris à St Kilda, Océan Atlantique, tandis que les triangles se rapportent à du poisson pris à Fyllas Bank, Détroit de Davis
Autolyse et catabolisme de nucléotide
Comme on l’a signalé auparavant, la rigor mortis survient quand le niveau de l’ATP dans le muscle chute à £ 1,0 mmole/g. L’ATP n’est pas seulement une source d’énergie puissante nécessaire à la contraction musculaire chez l’animal vivant mais elle agit également comme agent plastifiant. La contraction musculaire en soi est contrôlée par le calcium et une enzyme l’ATP-ase que l’on trouve dans chaque cellule musculaire. Quand les niveaux intracellulaires de Ca+2 sont > 1mM, l’ATP-ase activée par le Ca+2 réduit la quantité de l’ATP musculaire libre, ce qui conduit à l’interaction entre les principales protéines contractiles, l’actine et la myosine. Ceci produit finalement la contraction du muscle, le rendant dur et inextensible. Un poisson en état de rigor mortis ne peut normalement pas être fileté ou traité car la carcasse est trop raide pour être manipulée et est souvent déformée, rendant impossible le traitement mécanique (voir également le chapitre 3.2 sur la saignée et le chapitre 5.1 sur les modifications sensorielles).
La disparition de la rigor est un processus qui n’a pas encore été entièrement élucidé mais qui produit toujours le ramollissement (détente) du tissu musculaire. Elle serait reliée à l’activation d’une ou de plusieurs enzymes naturelles du muscle, digérant certains composants du complexe de rigor mortis. Le ramollissement du muscle durant la résolution de la rigor (et éventuellement les processus d’altération) coïncide avec les changements autolytiques. Une des premières modifications qu’on décèle est la dégradation des composés reliés à l’ATP de manière plus ou moins prévisible après la mort. La figure 5.4 illustre la dégradation de l’ATP pour former l’adénosine diphosphate (ADP), l’adénosine monophosphate (AMP), l’inosine monophosphate (IMP), l’inosine (Ino) et l’hypoxanthine (Hx). La dégradation des catabolites de l’ATP s’effectue de la même manière dans la plupart des poissons, mais la vitesse de chaque réaction individuelle (d’un catabolite à l’autre) varie beaucoup d’une espèce à l’autre et elle coïncide souvent avec le niveau perceptible de la dégradation déterminé par des analystes entraînés. Saito et al.(1959) ont été les premiers à observer ce schéma et à développer une formule pour la fraîcheur du poisson basée sur ces changements autolytiques :
où [ATP], [ADP], [AMP], [IMP], [Ino] et [Hx] représentent les concentrations relatives de ces composants dans le muscle du poisson, mesurées à différentes périodes pendant la conservation au froid.
Le paramètre K ou indice de "fraîcheur" donne une estimation de fraîcheur relative basée principalement sur les altérations autolytiques qui se produisent pendant la conservation post mortem du muscle. Par conséquent, plus la valeur de K est élevée, plus le niveau de fraîcheur est bas. Malheureusement, certaines espèces de poissons comme le cabillaud d’Atlantique atteignent une valeur maximum de K de loin supérieure à la durée de conservation déterminée par des juges entraînés. De ce fait, K n’est pas considéré une valeur fiable en tant qu’indicateur de fraîcheur pour tous les poissons de mer. De même, la dégradation des catabolites de nucléotides n’est qu’une coïncidence dans les changements perceptibles de la fraîcheur et n’est pas reliée nécessairement à la cause de la détérioration de la fraîcheur, du fait que l’on considère que seul l’Hx aurait un effet direct sur l’arrière-goût amer du poisson altéré (Hughes et Jones, 1966). Aujourd’hui, on considère généralement que l’IMP est responsable du goût recherché de poisson frais qui n’existe que dans les produits de la mer de première qualité. Aucun des catabolites de nucléotides n’est considéré comme ayant un rapport avec les changements perceptibles de texture pendant le processus autolytique excepté bien sûr l’ATP dont la perte est associée à la rigor mortis.
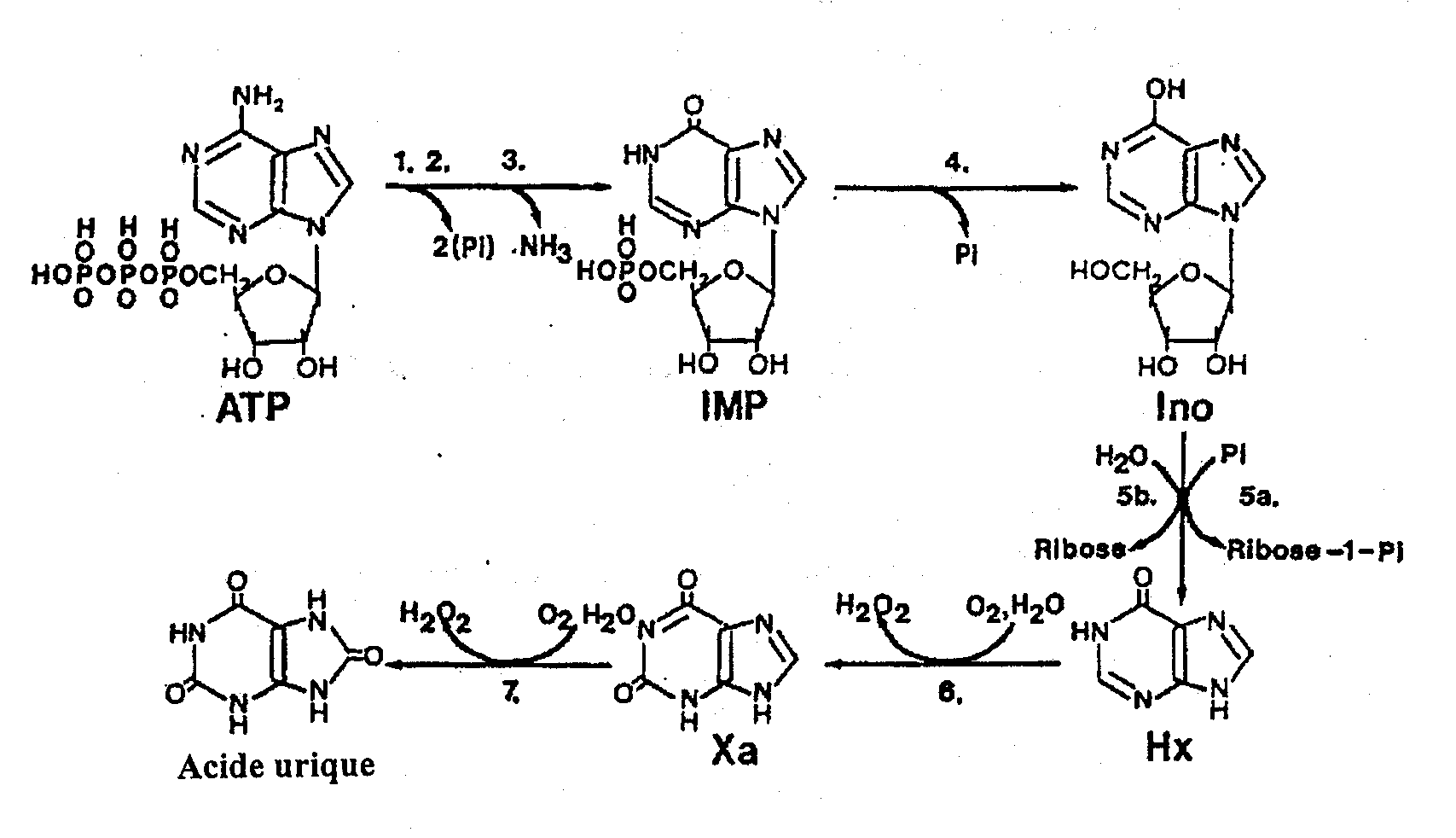
Figure 5.4 Dégradation post mortem de l’ATP dans le muscle de poisson. Les enzymes comprennent: 1. ATP-ase; 2. Myokinase; 3. AMP désaminase; 4.IMP phosphohydrolase; 5a. Nucléoside phosphorylase; 5b. Inosine nucléosidase; 6,7. Xanthine oxydase. Source: Gill (1992)
Surette et al. (1988) ont étudié l’autolyse du cabillaud stérile et non stérile comme l’indiquent les catabolites d’ATP. Les taux de formation et de décomposition de l’IMP étaient les mêmes dans les échantillons de tissus stériles et non stériles de cabillaud (Figures 5.5a et 5.5b) indiquant que la voie catabolique de dégradation de l’ATP donnant l’inosine est entièrement due aux enzymes autolytiques.
La conversion de l’Ino en Hx a été accélérée d’environ 2 jours dans les échantillons non stériles suggérant que la nucléoside phosphorylase bactérienne (enzyme 5a dans la Figure 5.4) joue un rôle majeur dans la production post mortem de Hx dans le cabillaud réfrigéré (voir également chapitre 5.3). Il est intéressant de noter que Surette et al.(1988) n’ont pas été capables d’isoler la nucléoside phosphorylase à partir du cabillaud fraîchement tué mais que Surette et al. (1990) sont arrivés à l’isoler et à la purifier à partir d’une bactérie Proteus isolée de filets de cabillaud avariés. Comme on l’a signalé auparavant, on peut s’attendre à de grandes variations dans les schémas de dégradation des nucléotides d’une espèce à l’autre. Les variations de Hx dans différents types de poissons sont exposées à la Figure 5.6. Ce graphique montre clairement que la détermination de Hx ne serait sans doute pas utile pour des espèces comme l’espadon et la rascasse du Nord.
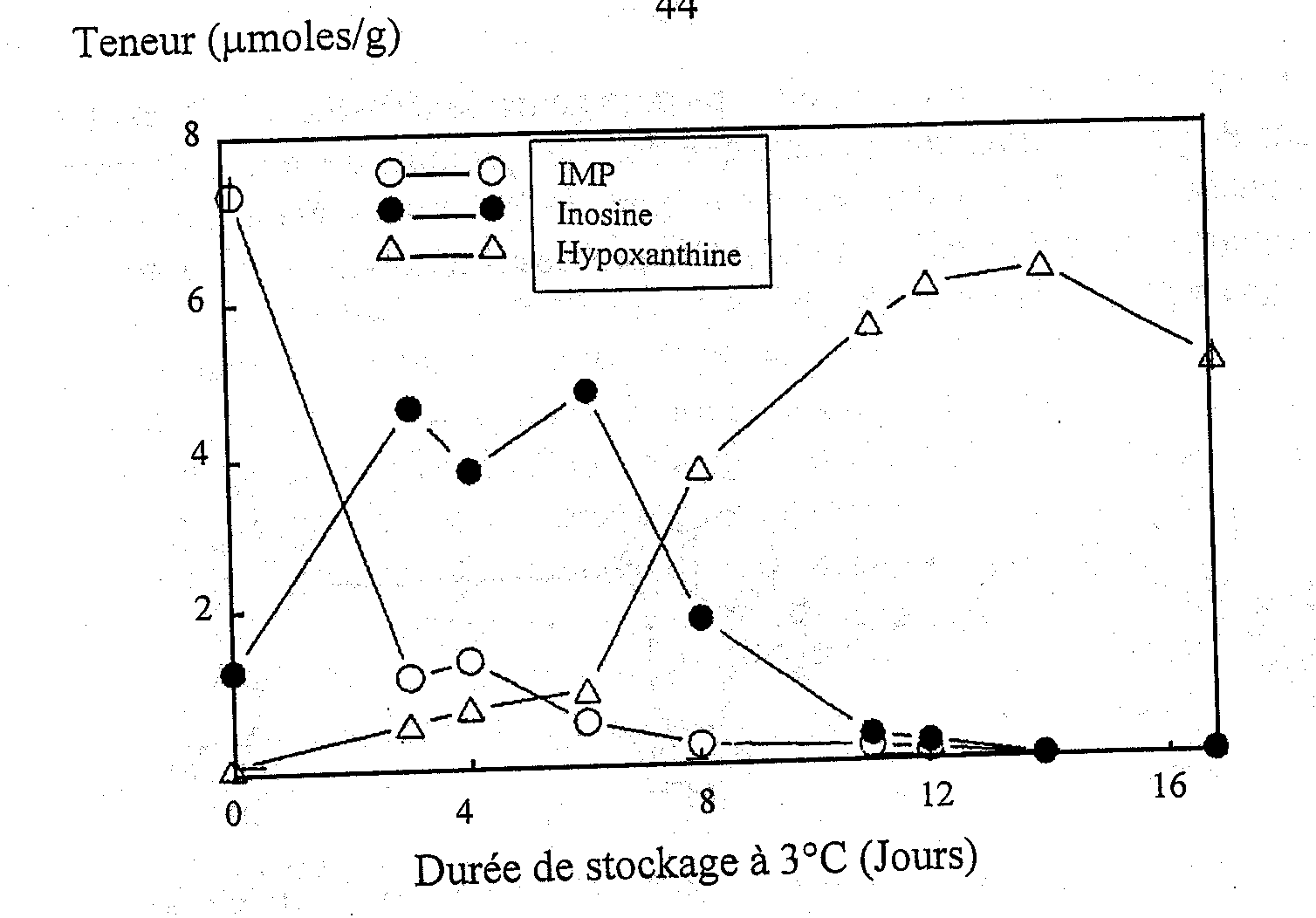
Figure 5.5a Variations de IMP, Ino et Hx dans des filets stériles de cabillaud à 3°C, d’après Gill (1990)
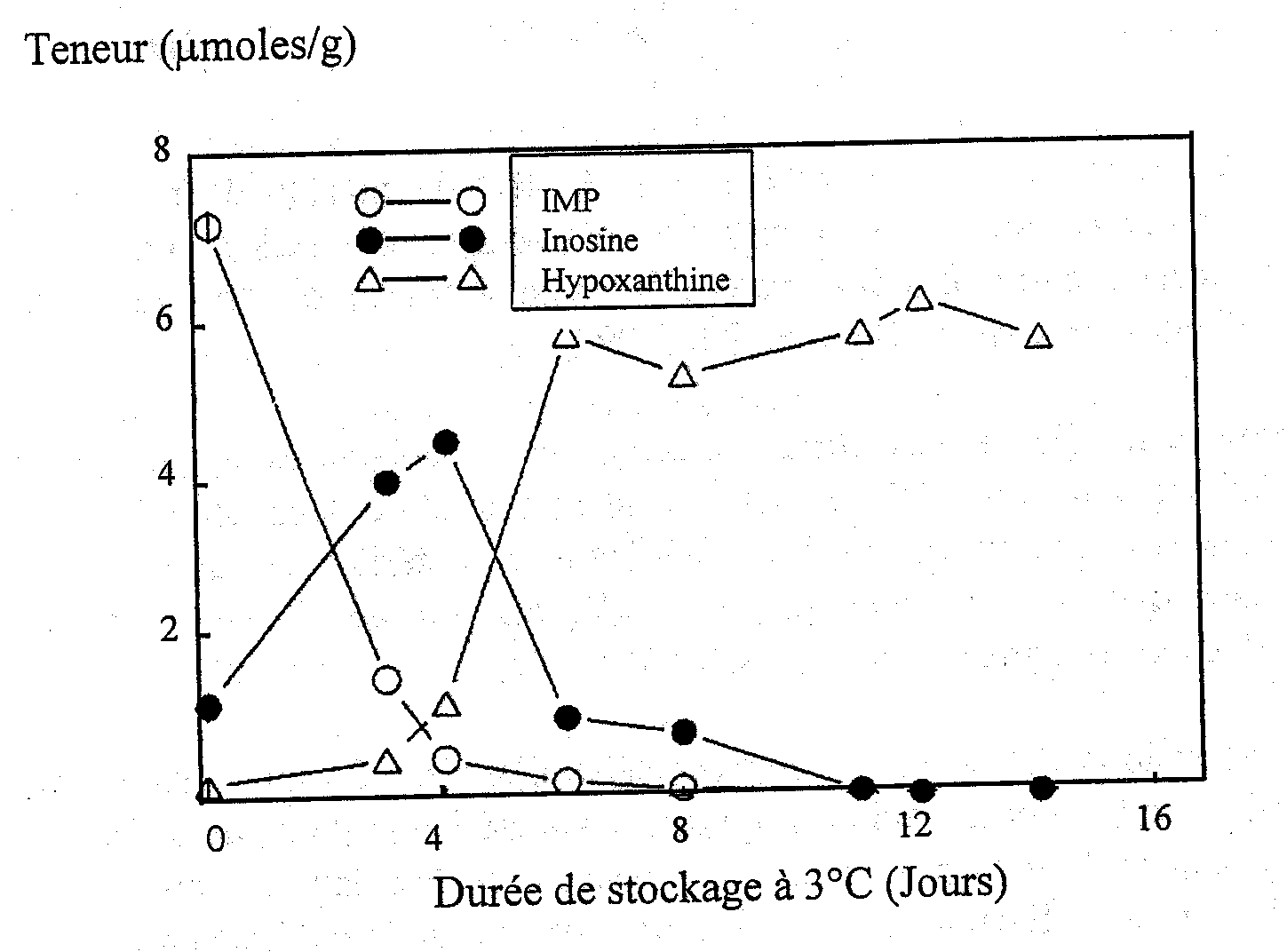
Figure 5.5b Variations de IMP, Ino et Hx dans des filets non stériles de cabillaud à 3° C, d’après Gill (1990)
Il est à peu près certain que la manutention physique accélère les changements autolytiques dans le poisson réfrigéré. Surette et al. (1988) ont trouvé que le taux de détérioration des catabolites de nucléotides est plus élevé dans les filets stériles que dans le cabillaud entier éviscéré non stérile. Ceci n’est peut être pas surprenant car on a démontré que plusieurs des enzymes autolytiques sont compartimentées en paquets enveloppés discrètement dans des membranes qui se déchirent quand elles sont soumises à de mauvais traitements physiques. Il en résulte un mélange intime des enzymes et du substrat. L’écrasement du poisson par la glace ou par d’autres poissons peut sérieusement affecter la comestibilité et les rendements du filetage même pour des poissons qui ont une charge bactérienne faible, montrant ainsi l’importance des processus autolytiques. Le poisson glacé ne devrait jamais être mis dans des caisses d’une profondeur supérieure à 30 cm et il est également important de s’assurer que les caisses à poisson ne puissent pas s’encastrer les unes dans les autres si on veut que l’autolyse soit réduite au minimum. Des systèmes adéquats pour convoyer le poisson et décharger les bateaux doivent être prévus afin d’éviter les mauvais traitements physiques aux tissus fragiles.
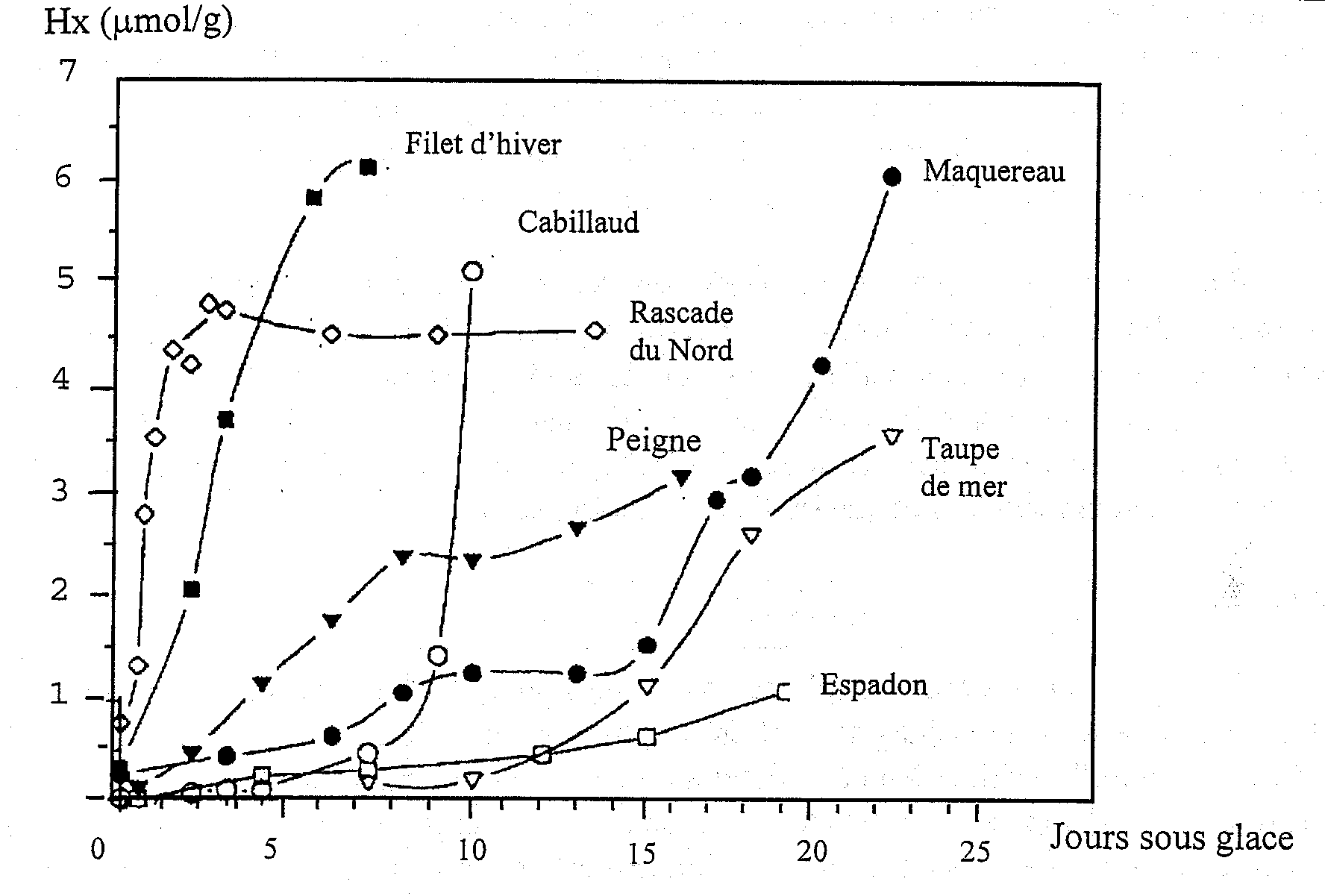
Figure 5.6 Variation du taux d’accumulation de Hx de différentes espèces durant la conservation sous glace. D’après Fraser et al. (1967)
Plusieurs méthodes rapides ont été mises au point pour déterminer les catabolites de nucléotides individuels ou des combinaisons incluant l’indice de fraîcheur. Le lecteur est invité à consulter les deux ouvrages récents de Gill (1990, 1992).
Altérations autolytiques incluant les enzymes protéolytiques
De nombreuses protéases ont été isolées des muscles du poisson et les effets de la dégradation protéolytique sont souvent reliés à un ramollissement considérable du tissu. Un des effets les plus notables de la protéolyse autolytique est peut-être l’éclatement de l’abdomen chez les espèces pélagiques (poissons gras) comme le hareng et le capelan. Ce type de ramollissement du tissu est plus fréquent dans les mois d’été quand les pélagiques mangent beaucoup, particulièrement des "aliments rouges" consistant en copépodes et euphosiidés. Les peptides à faible poids moléculaire et les acides aminés libres produits par l’autolyse des protéines non seulement diminuent l’acceptabilité commerciale des pélagiques, mais, dans les capelans stockés en vrac, on a constaté que l’autolyse accélérait le développement des bactéries d’altération en fournissant un environnement favorable à la croissance de tels organismes (Aksnes et Brekken, 1988). L’introduction de l’altération bactérienne dans le capelan par autolyse a provoqué également la décarboxylation des acides aminés, produisant des amines biogènes et a abaissé la valeur nutritive du poisson de façon significative. Ceci est particulièrement important du fait que l’autolyse et la croissance bactérienne abaissent sensiblement la valeur commerciale des pélagiques utilisés pour la production de farine de poisson.
De la même façon, on a constaté que le hareng, stocké en vrac et utilisé pour la farine de poisson, contient des carboxypeptidases A et B, de la chymotrypsine et de la trypsine; et des études préliminaires ont montré que la protéolyse peut être inhibée par l’addition d’extraits de pomme de terre qui non seulement ralentissent la protéolyse mais ont pour effet de ralentir également la croissance microbienne et de conserver la valeur nutritionnelle de la farine (Aksnes, 1989).
Plus récemment, Botta et al. (1992) ont trouvé que l’autolyse de la cavité viscérale (éclatement abdominal) des harengs était davantage reliée à la manutention physique qu’à des facteurs biologiques comme la taille du poisson, les aliments dans les viscères ou la quantité de rogue. En particulier, on a constaté que pour le hareng, les opérations de congélation et décongélation, le temps de décongélation à 15°C et le temps de stockage sous glace influaient plus sur l’éclatement abdominal que les facteurs biologiques.
Cathepsines
Bien que l’on ait découvert plusieurs enzymes protéolytiques dans les tissus du poisson, ce sont peut être les cathepsines qui ont été le plus souvent décrites. Les cathepsines sont des protéases "acides" habituellement rassemblées dans des organites très petites - infra- microscopiques - appelées lysosomes. On pense que, dans le tissu vivant, les protéases lysosomiques sont responsables de la dégradation des protéines aux endroits lésés. Les cathepsines sont pour la plupart inactives dans les tissus vivants mais sont libérées dans les jus des cellules à la suite d’accidents physiques ou de congélation/décongélation post mortem du muscle.
On croit que les cathepsines D et L jouent un rôle majeur dans la dégradation autolytique des tissus du poisson du fait que la plupart des autres cathepsines ont une plage d’activité étroite de pH bien trop basse pour être significative physiologiquement. Reddi et al. (1972) ont démontré qu’une enzyme, considérée comme étant la cathepsine D dans le flet d’hiver, était active sur une gamme de pH de 3 à 8 avec un pH maximum de près de 4,0 bien que l’on n’ait pas essayé de confirmer l’identité de l’enzyme en utilisant des substrats synthétiques ou des inhibiteurs spécifiques. Néammoins, l’enzyme était nettement moins active en présence d’ATP, ce qui laisse à penser qu’une telle enzyme serait seulement active post mortem dans le muscle de poisson. L’activité de l’enzyme était également fortement inhibée par la présence de sel (Figure 5.7) et l’activité était pratiquement inexistante après une incubation de 25 heures en présence de 5 % de chlorure de sodium. Il est donc improbable que l’enzyme de Reddi ait été active dans le poisson salé.
La cathepsine L a été impliquée dans le ramollissement du muscle de saumon pendant la migration de frai. Il est vraisemblable que cette enzyme contribue davantage à l’autolyse du muscle de poisson que la cathepsine D car elle est plus active à pH neutre et on a constaté qu’elle digérait aussi bien les protéines myofibrillaires (actomyosine) que les tissus conjonctifs. Yamashita et Konogaya (1990) ont apporté des preuves solides de l’action de la cathepsine L plutôt que des autres cathepsines sur le ramollissement du saumon durant la ponte. Ils ont démontré que l’électrophorèse de myofibrilles purifiées traitées avec la cathepsine L donnait des schémas pratiquement identiques à ceux des protéines récupérées sur les muscles des poissons en train de pondre. Par ailleurs, l’activité autolytique de la cathepsine L correspondait bien à la texture du muscle mesurée instrumentalement. La corrélation linéaire entre l’activité de la cathepsine L et la force de rupture du muscle était excellente avec r = 0, 86 et 0,95 respectivement pour des tissus frais et congelés/décongelés. Il est intéressant de constater que, dans tous les cas, la capacité autolytique, mesurée par l’activité de la cathepsine L, a été plus grande dans les tissus congelés/décongelés que dans les tissus frais. La congélation et la décongélation déchirent souvent les membranes des cellules permettant aux enzymes autolytiques enfermées dans les membranes de réagir à leurs substrats naturels. L’enzyme et son inhibiteur naturel ont, par la suite, été étudiés plus en détail par les mêmes auteurs (Yamashita et Konogaya, 1992). La cathepsine L a aussi été associée à la production d’un amollissant du flet ressemblant à de la gelée (Toyohara et al., 1993 a) et à l’amollissement incontrôlable des muscles du merlu du Pacifique qui a été parasité par Myxosporidia (Toyohara et al., 1993 b ).
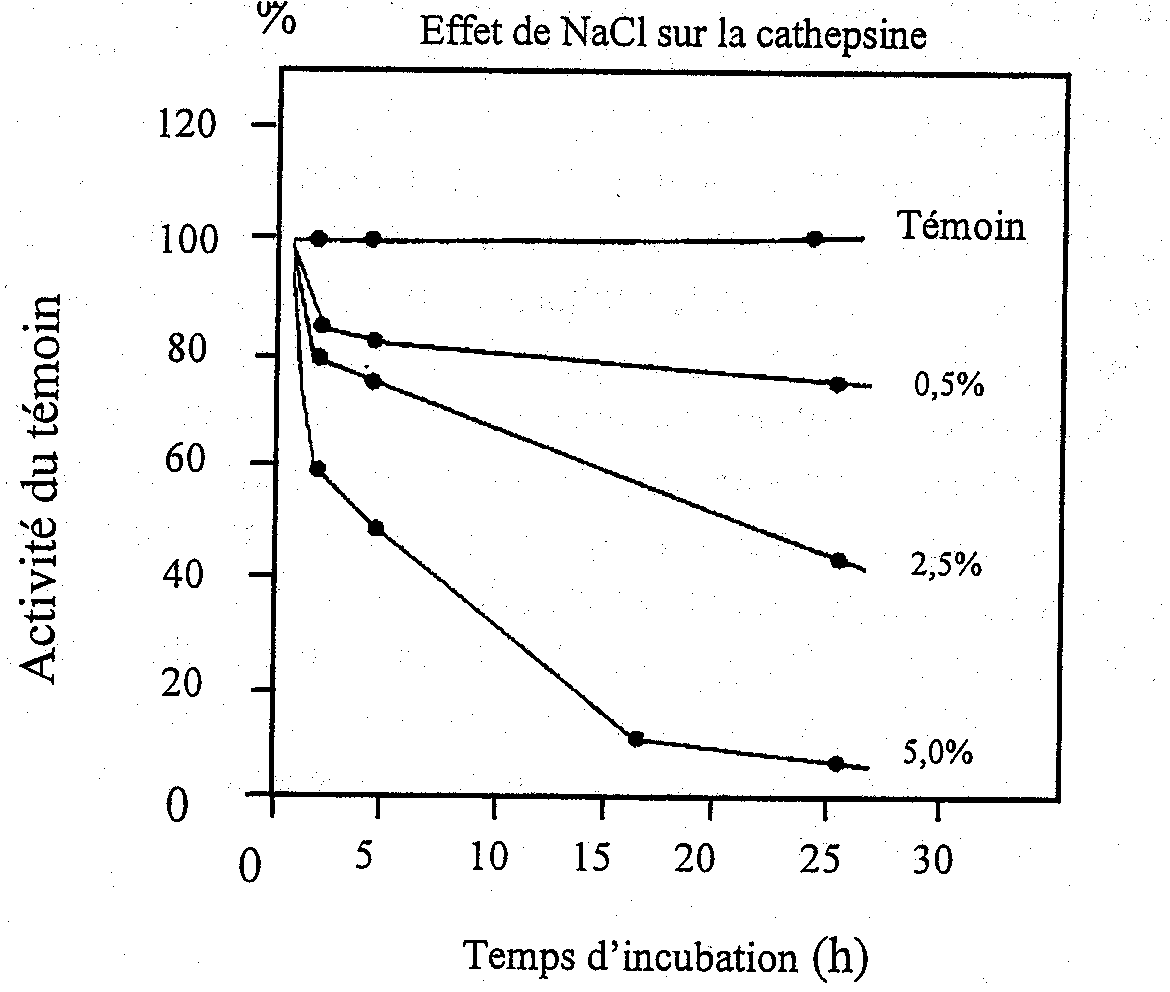
Figure 5.7 Effet du NaCl sur l'activité de la cathepsine. D’après Reddi et al. (1972)
Les tissus de tels poissons infectés n’ont que peu de valeur commerciale, mais, à ce jour, on ne sait pas si c’est le parasite ou son hôte qui secrète les enzymes protéolytiques qui autolysent le muscle.
En plus de leur effet nuisible sur la texture, les enzymes catheptiques causent des altérations autolytiques voulues dans les produits fermentés à base de poisson. On croit, par exemple, que les cathepsines sont responsables des changements majeurs de texture au cours de la fermentation des encornets japonais et de la carpe conservés sous sel (Makinodan et al., 1991, 1993).
Calpaines
Un second groupe de protéases intracellulaires appellées "calpaines" ou "calcium activated factor" (CAF) a récemment été associé à l’autolyse du muscle du poisson; on les trouve dans les viandes, les poissons à nageoires et les crustacés. Il est connu depuis près d'un siècle que la tendreté, probablement le critère le plus important pour la qualité de la viande rouge, est associée à sa maturation post mortem. Les calpaines seraient les premières responsables de l’autolyse post mortem de la viande par la digestion des protéines en Z des myofibrilles. Ces calpaines sont des endopeptidases intracellulaires nécessitant cystéine et calcium; les m-calpaines nécessitent de 5 à 50 mM Ca+2 et les m-calpaines de 150 à 1 000 mM Ca+2. La plupart des calpaines sont actives au pH physiologique et il paraît donc raisonnable de suspecter leur importance dans le ramollissement du poisson pendant la conservation au froid. Alors que la dureté est rarement un problème dans du poisson non congelé, le ramollissement par autolyse est un sérieux problème diminuant sa valeur commerciale.
Des études ont montré que, dans les muscles des crustacés, les calpaines sont associées à des changements de structure résultant de la mue et causant la digestion généralisée non spécifique des protéines myofibrillaires. Cependant on a démontré que des calpaines des muscles de vertébrés peuvent être très spécifiques, digérant d’abord la tropinine-T, desmine, titine et nébuline mais n’attaquant ni actine ni myosine (Koohmaraie, 1992). Par contre, les calpaines du poisson digèrent la myosine (particulièrement la myosine à chaîne longue) pour former un fragment initial avec un poids moléculaire d’environ 150 000 Da (Muramoto et al., 1989). Les mêmes auteurs ont démontré que les calpaines du poisson sont nettement plus actives à basse température que ne le sont les calpaines de mammifères, et que les vitesses de dégradation dépendent de l'espèce, étant très élevés dans le cas des myosines les plus sensibles à la chaleur. De ce fait, les espèces de poisson adaptées aux environnements plus froids sont plus sensibles à l'autolyse par les calpaines que les espèces provenant des eaux tropicales. Bien que l’on ait identifié des calpaines dans plusieurs espèces de poisson y compris la carpe (Toyohara et al., 1985), le tilapia et la crevette (Wang et al., 1993) ainsi que le thon, le maigre, la dorade rose et la truite (Muramoto et al.,1989) pour ne citer que celles-là , à ce jour peu de travaux ont été réalisés pour démontrer une relation de cause à effet entre l’activité des calpaines et la texture telle que mesurée par voie instrumentale.
Collagénases
Jusqu’à présent, tous les changements autolytiques post mortem décrits ont concerné les changements à l’intérieur de la cellule musculaire elle-même. Cependant la chair des poissons téléostéens est divisée en blocs de cellules musculaires séparées en lamelles ou myotomes par un tissu conjonctif appelé myocomme (Figure 3.3). Chaque cellule musculaire ou fibre est enveloppée d’un tissu conjonctif qui se fixe au myocomme, aux extrémités des cellules, au moyen de fines fibrilles de collagène. Pendant la conservation au froid, ces fibrilles se détériorent (Bremner et Hallett, 1985). Plus récemment, on a montré que des mesures instrumentales de la texture du muscle de truite réfrigérée diminuaient quand la quantité de collagène de type V était solubilisée, sans doute du fait de l’action autolytique des enzymes collagénases autolytiques (Sato et al., 1991). Ce sont ces enzymes qui produisent sans doute le "clivage" ou la destruction du myotome durant la conservation de longue durée sous glace ou de brève durée à haute température. Pour le cabillaud de l’Atlantique, on a montré que lorsque l’on atteint 17°C, le clivage est inévitable, sans doute à cause de la dégradation du tissu conjonctif et de la contraction rapide du muscle due à une rigor à haute température.
On a également montré qu'une durée de conservation relativement courte des crevettes réfrigérées due au ramollissement du tissu était due à la présence d’enzymes collagénases (Nip et al., 1985). Ces enzymes proviendraient de l’hépatopancréas (organe de digestion).
Altérations autolytiques pendant la conservation du poisson congelé
La réduction de l’oxyde de triméthylamine (OTMA), un composé osmorégulateur dans de nombreux poissons téléostéens marins, est habituellement due à l’action bactérienne (chapitre 5.3). Mais, dans certaines espèces le tissu musculaire contient une enzyme musculaire capable de dégrader l'OTMA en diméthylamine (DMA) et formaldéhyde (FA):
(CH3)3 NO
(CH3)2 NH + HCHO
Il est important de noter que la quantité de formaldéhyde produite est équivalente à la diméthylamine formée mais possède une importance plus grande du point de vue commercial. Le formaldéhyde provoque des liaisons croisées des protéines musculaires, ce qui durcit le muscle et lui fait perdre rapidement sa capacité de rétention d’eau. L’enzyme responsable du durcissement provoqué par la formaldéhyde est appelée OTMA-ase ou OTMA déméthylase et est très communément trouvée dans les gadidés (famille du cabillaud). La plupart des OTMA déméthylases connues à ce jour sont liées aux membranes et deviennent actives quand les membranes du tissu sont déchirées par la congélation ou artificiellement par dissolution par détergent. Le muscle rouge a un taux d’activité plus élevé que le muscle blanc alors que d’autres tissus comme les reins, la rate et la vessie natatoire sont extrêmement riches en cette enzyme. Il est donc important que le poisson haché soit complètement débarassé des organes tels que les reins chez les gadidés si on veut éviter le durcissement au cours du stockage en congélation. Il est souvent difficile d’avoir la certitude que les reins ont été enlevés avant le désarêtage mécanique car cet organe particulier se trouve tout au long de l’arête dorsale à laquelle il adhère. L’enzyme OTMA-ase a été isolée de la fraction microsomique dans le muscle de merlu (Parkin et Hultin, 1986) ainsi que de la membrane lysosomique dans les tissus rénaux (Gill et al.., 1992). On a démontré que le durcissement du muscle de merlu congelé est lié à la quantité de formaldéhyde produite et que le taux de production de FA est le plus important aux températures les plus fortes de congélation (Gill et al., 1979). De plus, on a constaté que l’importance du durcissement dû au FA est augmenté par la mauvaise manutention infligée au poisson avant la congélation et par les variations de température au cours de la conservation en congelé. Les moyens les plus pratiques pour prévenir la production autolytique de FA sont de conserver le poisson à des températures inférieures à -30°C pour réduire les variations de température dans la chambre froide et d’éviter les manipulations brutales ou l’application de pression physique sur le poisson avant de le congeler. Les altérations autolytiques affectant la comestibilité du poisson frais et congelé sont résumées au Tableau 5.3. Généralement le facteur le plus important affectant l’autolyse est la déchirure physique des cellules musculaires. A remarquer qu'on n’a pas essayé ici de traiter des protéases alcalines associées au ramollissement des produits de surimi cuit et qui font l'objet de l'article écrit par Kinoshita et al. (1990).
Tableau 5.3 Résumé des changements autolytiques dans le poisson réfrigéré
Enzyme(s) |
Substrat |
Modifications produites |
Prévention/Inhibition |
| Enzymes glycolytiques | Glycogène | Production d’acide lactique, chute du pH des tissus, perte de la
capacité de rétention d’eau du muscle. La rigor à haute température peut produire des déchirures. |
On doit laisser le poisson franchir la rigor à des températures
aussi proches que possible de 0°C Le stress pre-rigor doit être évité |
| Enzymes autolytiques impliqués dans la dégradation des nucléotides | ATP ADP AMP IMP |
Perte du goût de poisson frais, production progressive....amertume avec Hx (étapes suivantes) | Comme ci- dessus Une manutention brutale et l’écrasement précipitent l’altération |
| Cathepsines | Protéines, peptides | Ramollissement des tissus rendant le traitement difficile ou impossible | Manutention brutale au stockage et au déchargement |
| Chymotrypsine, trypsine, carboxy-peptidases | Protéines, peptides | Autolyse des parois abdominales chez les pélagiques (éclatement) | Problème amplifié par les congélations/décongélations ou un stockage prolongé au froid |
| Calpaine | Protéines myofibrillaires | Ramollissement dû à la mue chez les crustacés | Elimination du calcium évitant ainsi l’activation |
| Collagènases | Tissu conjonctif | «Déchirures» des filets Ramollissement |
Dégradation du tissu conjonctif en fonction des durée et température de stockage au froid |
| OTMA Déméthylase |
OTMA | Durcissement des gadidés congelés dû au formaldéhyde | Conserver le poisson à une température inférieure à -30°C Mauvais traitements physiques et congélation/décongélation accélèrent le durcissement dû au formaldéhyde |
Flore bactérienne du poisson vivant
Les microo-organismes se trouvent sur toute la surface externe (peau et branchies) et dans les intestins des poissons vivants et fraîchement pêchés. Le nombre varie énormément allant de 102 à 107 UFC (unités formant colonies)/cm² de surface de peau (Liston, 1980) et de 103 à 109 UFC/g de branchies ou d'intestins (Shewan, 1962).
La flore bactérienne du poisson fraichement pêché dépend de l’environnement dans lequel il a été capturé, plus que de l’espèce de poisson (Shewan, 1977). Le poisson pêché dans des eaux propres et très froides a la charge bactérienne la plus faible alors que celle du poisson pêché dans les eaux chaudes est légèrement plus élevée. Des charges importantes, de l'ordre de 107 UFC/cm² sont trouvées sur les poissons provenant d’eaux chaudes polluées. Concernant les espèces de bactéries, on en rencontre plusieurs à la surface du poisson. Elles se classent en psychrotrophes ou psychrophileses. Selon leurs intervalles de température de croissance les psychrothrophes (supportant le froid) sont capables de se développer à 0°C mais avec un optimum aux environs de 25°C. Les psychrophiles (aimant le froid) possèdent une température maximale de croissance aux environs de 20°C et une température optimum à 15°C (Morita, 1975). Les mésophiles sont isolés en nombres importants à partir des poissons des eaux plus chaudes. La microflore des poissons d’eaux tempérées est dominée par les bactéries psychrotrophes à Gram négatif en forme de bâtonnets appartenant aux genres Pseudomonas, Moraxella, Acinetobacter, Shewanella et Flavobacterium. Les membres de la famille des Vibrionaceae (Vibrio et Photobacterium) et des Aeromonodaceae (Aeromonas spp.) sont aussi des bactéries aquatiques courantes et typiques de la flore du poisson (Tableau5.4). Des bactéries à Gram positif comme Bacillus, Micrococcus, Clostridium, Lactobacillus et corynéformes peuvent être également trouvés en quantités variables, mais ce sont généralement les bactéries à Gram négatif qui prédominent. Shewan (1977) avait conclu que les genres Bacillus et Micrococcus à Gram positif dominent dans les poissons des eaux tropicales. Cette conclusion n'a cependant pas été confirmée par plusieurs études ultérieures qui ont trouvé que la microflore des espèces de poissons tropicaux est très semblable à la flore des espèces tempérées (Acuff et al., 1984; Gram et al., 1990; Lima dos Santos 1978; Surendran et al., 1989). Plusieurs études menées en Inde (Surendran et al. 1989) ont permis d'isoler une microflore constituée de Pseudomonas, Acinetobacter, Moraxella et Vibrio à partir de poisson fraîchement pêché. De ce fait, plusieurs auteurs concluent, comme Liston (1980), que la microflore du poisson tropical porte souvent une charge légèrement supérieure en bactéries à Gram positif et entériques, mais est par ailleurs semblable à la flore du poisson des eaux tempérées.
Les Aeromonas sont typiques des poissons d’eaux douces alors que nombre de bactéries ont besoin de sodium pour leur croissance et sont donc typiques des eaux marines. Celles-ci incluent Vibrio, Photobacterium et Shewanella. Cependant bien que l'espèce Shewanella putrefaciens soit caractérisée par son besoin en sodium, on peut également isoler des souches de S. putrefaciens dans des environnements d’eau douce (Di Christina et De Long, 1993; Gram et al., 1990; Spanggaard et al., 1993). Toutefois, S. putrefaciens ne semble pas jouer un rôle important dans l'altération des espèces de poisson d'eaux douces, bien qu'on l'ait isolé d'espèces tropicales de poisson d'eaux douces (Lima dos Santos, 1978; Gram, 1990).
Tableau 5.4 Flore bactérienne du poisson capturé dans des eaux propres non polluées
Gram-négative |
Gram-positive |
Commentaires |
| Pseudomonas Moraxella Acinetobacter Shewnella putrefaciens Flavobacterium Cytophaga Vibrio Photobacterium Aeromonas |
Bacillus Clostridium Micrococcus Lactobacillus Coryneforms |
Vibrio et Photobacterium sont typiquement marins; Aeromonas est typiquement d’eau douce. |
Dans les eaux polluées, on peut trouver des charges élevées d’Enterobacteriaceae. Ces organismes disparaissent toutefois rapidement dans des eaux tempérées propres. Cependant, on a constaté que Escherichia coli et Salmonella peuvent survivre très longtemps dans les eaux tropicales et, une fois introduits, peuvent devenir indigènes de l’environnement (Fujioka et al., 1988).
La taxonomie de S. putrefaciens a souvent prêté à confusion. Cet organisme était à l’origine, associé au groupe Achromobacter puis classé plus tard dans le genre Pseudomonas groupe IV de Shewan. sur la base du pourcentage en guanine + cytosine (GC %), Il a été ensuite transféré dans le genre Alteromonas, puis classé dans un nouveau genre, Shewanella sur la base de l'homologie 5SRNA (Mac Donnell et Colwell, 1985). On a de même suggéré que le genre Aeromonas spp. qui était un membre de la famille des Vibrionaceae soit transféré dans sa propre famille, les Aeromonadaceae (Colwell et al., 1986).
Des études japonaises ont décelé une charge élevée de micro-organismes dans le tractus gastro-intestinal du poisson bien supérieure à celle des eaux environnantes, ce qui indique la présence d’une niche écologique favorable à ces micro-organismes. De même, Larsen et al., (1978) ont trouvé jusqu’à 107 UFC/g de bactéries du type vibrio dans l’appareil intestinal du cabillaud et Westerdahl et al. (1991) en ont isolé un grand nombre dans les intestins du turbot. Photobacterium phosphoreum qui peut être isolé de la surface du poisson, se rencontre également en grand nombre dans l’appareil gastro-intestinal de certaines espèces de poissons (Dalgaard, 1993). Par contre, certains auteurs pensent que la microflore de l’appareil gastro-intestinal est simplement le reflet de l’environnement et de l’alimentation.
Invasion microbienne
La chair du poisson sain, vivant ou fraîchement pêché est stérile car le système immunitaire du poisson empêche les bactéries de se multipliproliférerer dans sa chair (Figure 5.8 aa). A la mort du poisson, le système immunitaire s’effondre et les bactéries peuvent proliférer librement. A la surface de la peau, les bactéries colonisent largement les alvéoles des écailles. Pendant le stockage, elles envahissent la chair en se déplaçant entre les fibres musculaires. Murray et Shewan (1979) ont trouvé que seul un nombre très limité de bactéries envahissent la chair pendant la conservation sous glace. Ruskol et Bendsen (1992) ont montré que les bactéries peuvent être détectées au microscope dans la chair, stockée sous glace ou à température ambiante, quand le nombre d’organismes à la surface de la peau dépasse 106 UFC/cm² (Figure 5.6 b). Aucune différence du mode d'invasion n'a été détectée dans les bactéries spécifiques d’altération (par ex.emple S. putrefaciens) et les autres bactéries.
Du fait que seul un nombre limité d’organismes envahit réellement la chair et que le développement microbien se situe essentiellement à la surface, l’altération s’avère probablement, pour une grande part, être une conséquence de la diffusion des enzymes bactériennes dans la chair et des nutriments à l’extérieur.
Le poisson s’altère à des vitesses très variables (voir aussi chapitre 6.5). Certains auteurs expliquent ce fait par des différences dans les propriétés de la surface du poisson. Les peaux des poissons ont des textures très différentes. Ainsi, le merlan (Merlangius merlangus) et le cabillaud (Gadus morhua) qui ont un tégument très fragile s’abiment rapidement comparés à différents poissons plats comme le carrelet qui possède un derme et un épiderme très robustes. Ces derniers ont de plus une couche épaisse de mucus qui comprend plusieurs substances antibactériennes tels que des anticorps et des enzymes bactériolytiques (Murray et Fletcher, 1976; Hjelmland et al., 1983).
Evolutions de la microflore au cours du stockage et altération/organismes spécifiques d’altération
Les bactéries du poisson pêché dans les eaux tempérées entreront dans la phase de croissance exponentielle presque immédiatement après la mort du poisson. Ceci est également vrai quand le poisson est glacé, probablement parce que la microflore s’est déjà adaptée aux basses températures. Pendant la conservation sous glace, les bactéries vont doubler pratiquement chaque jour et, après 2 à 3 semaines, elles auront atteint 108 - 109 UFC/g de chair ou par cm² de peau. Pendant la conservation à température ambiante, elles atteignent près de 107 -108 UFC/g en 24 heures. Les bactéries des poissons capturés dans les eaux tropicales passeront souvent par une phase de latence de 1 à 2 semaines si le poisson est conservé sous glace, après quoi la croissance exponentielle commence. Au cours de la dégradation, le niveau bactérien des poissons tropicaux est semblable à celui observé sur les espèces de poissons des eaux tempérées (Gram, 1990; Gram et al., 1990).
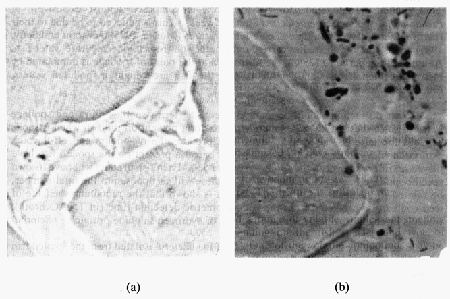
Figure 5.8 Coupe histologique de (a) cabillaud frais et (b) de filets de cabillaud stockés sous glace pendant 12 jours. La coupe a été colorée au Giemsa (Ruskol et Bendsen, 1992)
Si le poisson glacé est conservé dans des conditions anaérobies ou si on le met dans une atmosphère contenant du CO2, le nombre de bactéries psychrotrophes telles que S. putrefaciens et Pseudomonas est souvent bien plus faible, c’est-à-dire 106-107 UFC/g, que sur le poisson conservé en aérobiose. Cependant, le taux de bactéries à caractère psychrophile telles que P. phosphoreum atteint 107-08 UFC/g lorsque le poisson s'altère (Dalgaard et al., 1993).
La composition de la microflore varie également de façon très importante pendant le stockage. Ainsi, dans la conservation sous glace en aérobiose, la flore est composée presque exclusivement de Pseudomonas spp. et S. putrefaciens après 1 à 2 semaines. Ceci serait dû à leur temps de génération relativement court à basse température (Morita, 1975; Devaraju et Setty, 1985) et se vérifie pour toutes les études menées sur du poisson qu'il soit tropical, ou d’eaux tempérées. A température ambiante (25°C), la microflore au stade de l’altération est dominée par les mésophiles Vibrionaceae, et particulièrement, si les poissons sont pêchés dans des eaux polluées, par les Enterobacteriaceae.
On doit faire une distinction claire entre les termes flore d’altération et bactérie d’altération car la première décrit simplement les bactéries présentes sur le poisson quand il s’altère tandis que la seconde constitue le groupe spécifique qui produit les odeurs et les goûts désagréables associés à la dégradation. Une grande partie des bactéries présentes sur le poisson altéré ne joue aucun rôle dans son altération (Figure 5.9). Chaque poisson possède ses propres bactéries spécifiques d’altération dont le nombre à l'opposé du nombre total de bactéries, sera en rapport avec la durée de conservation. La Figure 5.10 montre que l’on peut prédire la durée limite de conservation du cabillaud glacé d’après le temps de détection conductimétrique (dans un bouillon à base d'OTMA) qui est inversement proportionnel au nombre de bactéries produisant du sulfure d'hydrogène.
L'identification des bactéries d'altération du poisson n’est pas une tâche facile. Celà nécessite des études sensorielles, microbiologiques et chimiques poussées. D’abord, les changements sensoriels, microbiologiques et chimiques au cours du stockage doivent être étudiés et quantifiés, y compris la détermination du niveau d’un composé chimique donné, qui présente une bonne corrélation avec l’altération. C'est l’indicateur chimique d’altération.
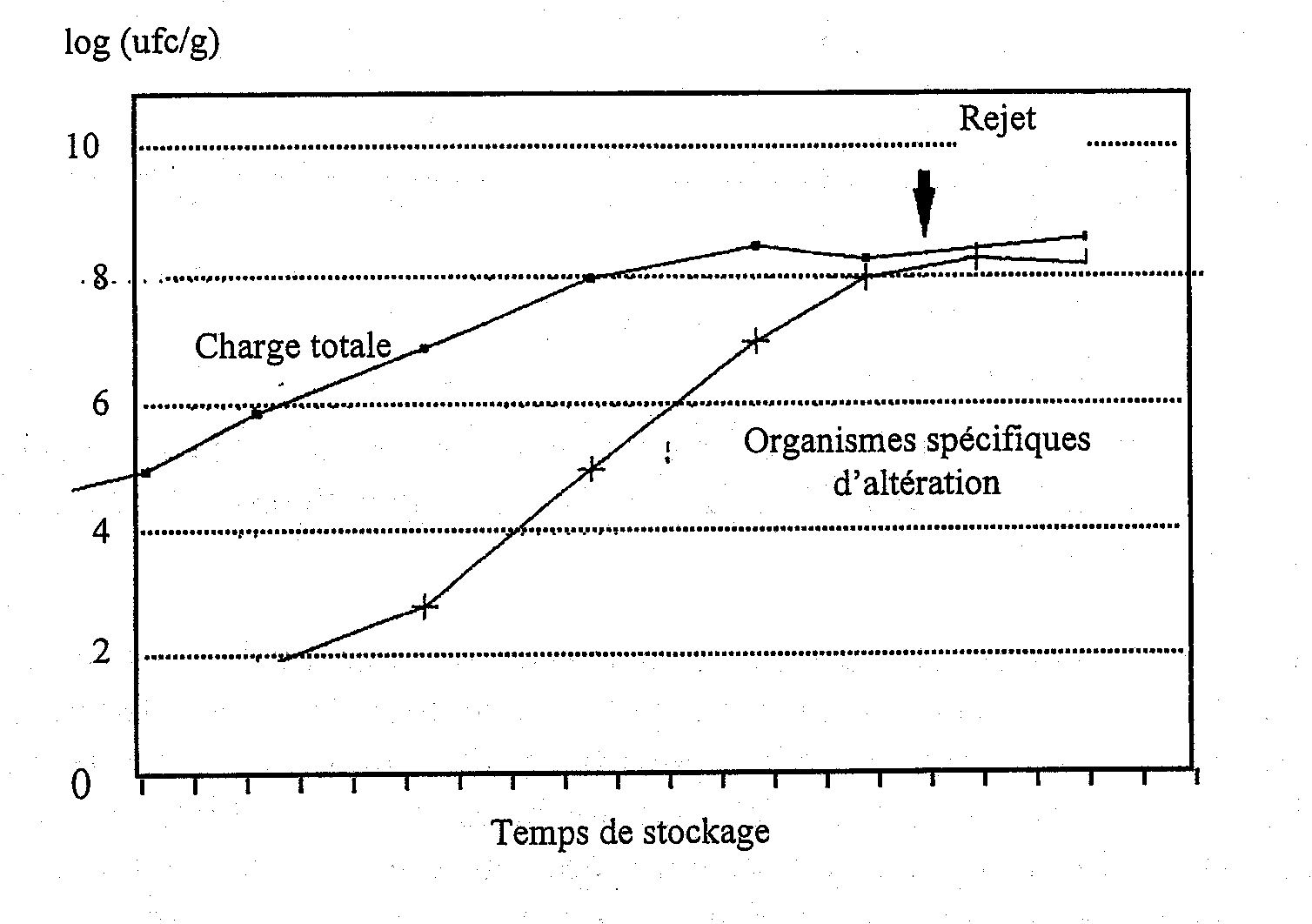
Figure 5.9 Evolutions de la flore totale et des bactéries spécifiques d’altération au cours de la conservation (modifiée d’après Dalgaard, 1993)
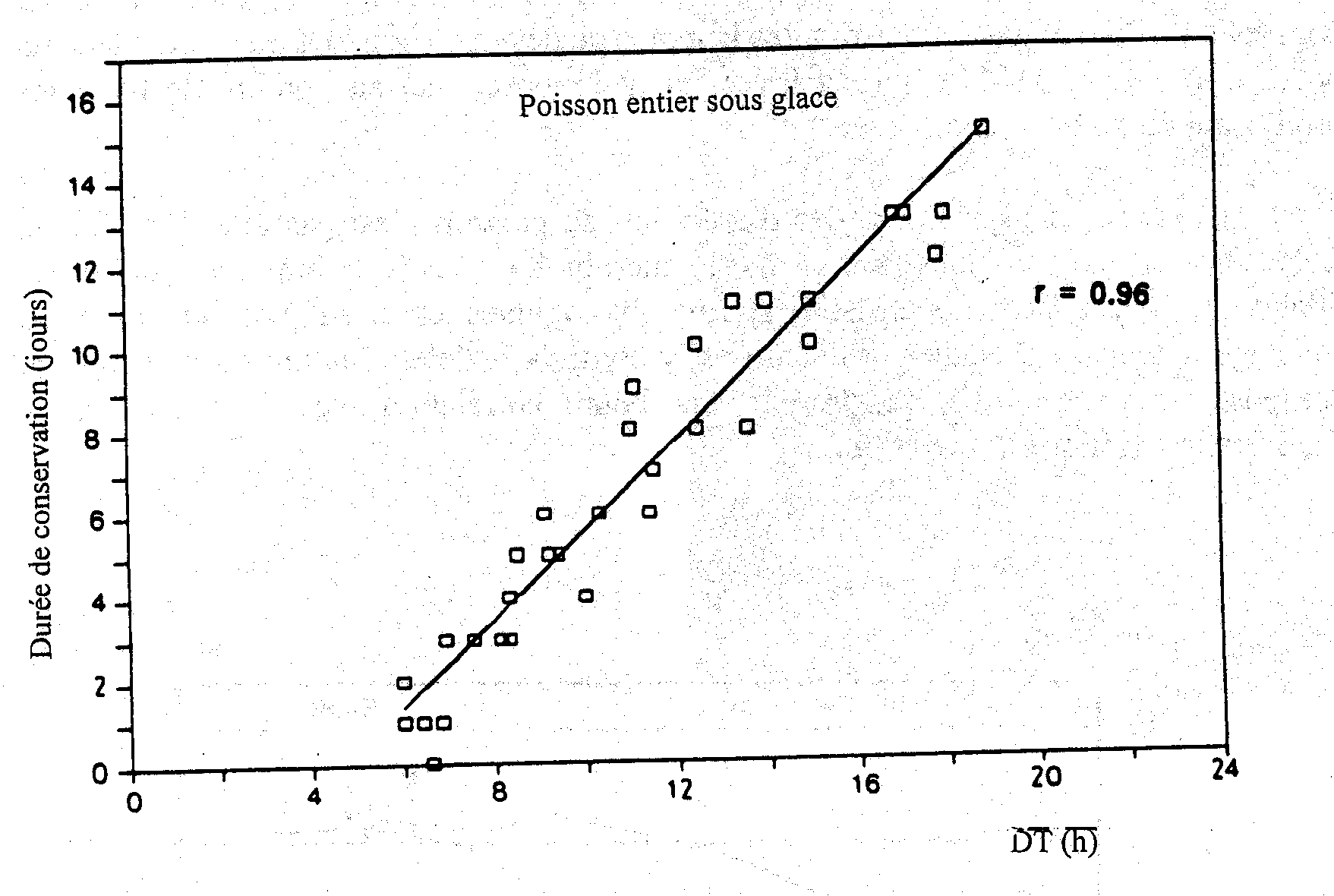
Figure 5.10. Comparaison entre la durée limite de conservation du cabillaud congelé et le temps de détection dans un bouillon de culture à l’OTMA (Jorgensen et al., 1988)
Deuxièmement, les bactéries sont isolées au moment du rejet sensoriel. Des cultures bactériennes pures ou en mélanges sont étudiées dans des substrats stériles de poisson et classées en fonction de leur potentiel d’altération, c’est-à-dire leur capacité à produire des changements sensoriels (odeurs indésirables) et chimiques typiques du produit altéré. Enfin, les souches sélectionnées sont testées pour évaluer leur pouvoir d’altération, c’est-à-dire pour vérifier si leur taux de croissance et leur production qualitative et quantitative d’odeurs indésirables sont similaires aux mesures effectuées sur les produits altérés (Dalgaard, 1993).
Pour sélectionner les bactéries potentielles d’altération dans une flore d’altération, on peut aussi utiliser un essai qualitatif évaluant la capacité des bactéries à produire du H2S et/ou réduire l'OTMA. Un milieu de culture permettant de détecter la réduction de l'OTMA en TMA par un changement de la couleur d’un indicateur redox et la formation de H2S par formation d'un précipité noir de FeS, a été mis au point par Gram et al.(1987).
On a identifié Shewanella putrefaciens comme la bactérie résponsable de l’altération du poisson des eaux tempérées conservé en aérobiose sous glace. Si le produit est emballé sous vide, P. phosphoreum participe à l’altération et devient la bactérie spécifique de l’altération du poisson emballé sous CO² (voir chapitre 6.3). La flore d’altération du poisson des mers tropicales est composée presque exclusivement de Pseudomonas spp. et S. putrefaciens. Quelques espèces de Pseudomonas sont responsables de l'altération des poissons tropicaux d’eau douce stockés sous glace (Lima dos Santos, 1978; Gram et al., 1990) et constituent avec S. putrefaciens les bactéries d'altération des poissons marins tropicaux conservés sous glace (Gillespie et MacRae, 1975; Gram, 1990).
A température ambiante, les aéromonades mobiles sont spécifiquement responsables de l'altération des poissons d’eau douce conservés en aérobiose (Gorzyka et Pek Poh Len, 1985; Gram et al., 1990). Barile et al (1985) ont montré qu’une large proportion de la flore du maquereau conservé à température ambiante consistait en S. putrefaciens, indiquant que cette bactérie peut aussi jouer un rôle dans l’altération.
Le Tableau 5.5 donne un aperçu des bactéries spécifiques d’altération du poisson frais conservé sous glace et à température ambiante.
Changements biochimiques produits par le développement bactérien durant la conservation et l’altération
La comparaison des composés chimiques qui se développent dans le poisson au cours de sa dégradation naturelle et dans le poisson stérile a montré que la plupart des composés volatils sont produits par les bactéries (Shewan, 1962) comme le montre la Figure 5.11. Ceux-ci comprennent la triméthylamine, les composés soufrés volatils, les aldéhydes, les cétones, esters, hypoxanthine et autres composés de faible poids moléculaire.
Tableau 5.5 Microflore dominante et bactéries d’altération identifiées sur du poisson blanc frais (cabillaud) au moment de son altération (d’après Huss, 1994)
Température de stockage |
Atmosphère d’emballage |
Microflore dominante |
Organismes Spécifiques d’Altération (OSA) |
Références |
0°C |
Aérobie |
Psychrotrophe Gram-négative, bâtonnets incapables de fermentation (Pseudomonas Spp., S. putrefaciens, Moraxella, Acinetobacter) | S. putrefaciens Pseudomonas 3 | 2,3,4,9 |
Vide |
Bâtonnets Gram-négatifs; psychhrotrophes ou à caractère psychrophile (S. putrefaciens, Photobacterium) | S. putrefaciens P. phosphoreum |
1,9 |
|
EAM1 |
Bâtonnets capables de fermentation Gram-négatifs à caractère
psychrophile (Photobacterium) Bâtonnets psychrotrophes incapable de fermentation (1 à10% de la flore, Pseudomonas, S. putrefaciens) Bâtonnets Gram-positifs (BL²) |
P. phosphoreum | 1,7 |
|
5°C |
Aérobie |
Bâtonnets psychrotrophes Gram-négatifs; (Vibrionaceae, S. putrefaciens) |
Aeromonas spp. S. putrefaciens |
10 |
Vide |
Bâtonnets psychrotrophes Gram-négatifs (Vibrionaceae, S. putrefaciens) |
Aeromonas spp. S. putrefaciens |
10 |
|
MAP |
Bâtonnets psychrotrophes Gram-négatifs (Vibrionaceae) |
Aeromonas spp. | 6 |
|
20/30°C |
Aérobie |
Bâtonnets capables de fermentation masophyles Gram-négatifs (Vibrionaceae, Enterobacteriaceae) |
Aeromonas spp. Mobile (A. hydrophila) |
2,4,5,8 |
1) EAM: Emballage sous Atmosphère Modifiée (contenant CO2). 2) BL: bactéries lactiques 3) Le poisson capturé dans les eaux tropicales ou en eau douce est surtout altéré par Pseudomonas spp.
References:
1) Barile et al. (1985); 2) Dalgaard et al. (1993); 3) Donald et Gibson
(1992), 4) Gorczyca et Pek Poh Len (1985); 5) Gram et al. (1987); Gram et al.
(1990); 7) Gram et Dalgaard (communication personnelle), 8) Jorgensen et Huss (1989); 9)
Lima dos Santos (1978); 10) Van Spreekens (1977)
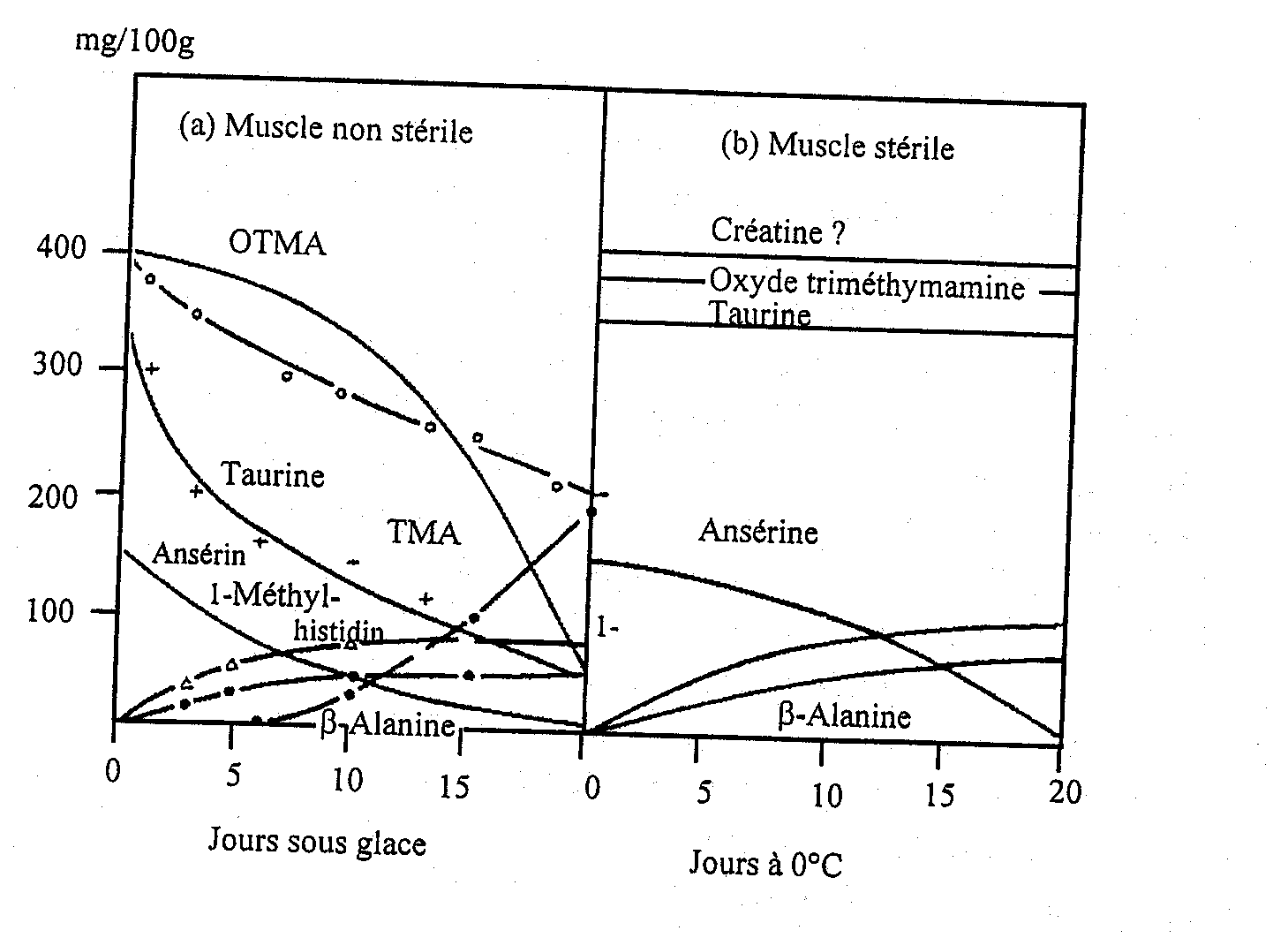
Figure 5.11 Evolution des extraits azotés dans le muscle de cabillaud a) en cours d’altération et b) pendant l’autolyse) (Shewan , 1962)
Les substrats pour la production des composés volatils sont les carbohydrates (par exemple lactate et ribose), les nucléotides (par exemple inosine monophosphate et inosine) et d’autres molécules azotées non protéiques ANP. Les acides aminés sont des substrats particulièrement importants pour la formation des sulfures et de l’ammoniaque.
Les micro-organismes retirent bien plus d’énergie de l’oxydation aérobie que de la fermentation anaérobie. Ainsi, l’oxydation totale d’une mole de glucose (ou autre hexose) par les cycles de Kreb produit 6 moles de CO2 et 36 moles d’ATP, alors que la fermentation d’une mole de glucose donne seulement 2 moles d’ATP et 2 moles d’acide lactique. Le développement aérobie initial dans le poisson est dominé par les bactéries utilisant les carbohydrates comme substrat et l’oxygène comme accepteur final d’électrons avec production également de CO2 et H2O.
Réduction de l'oxyde de triméthylamine (OTMA)
La croissance des bactéries consommant de l’oxygène provoque la formation de niches anaérobies ou micro-aérophiles dans le poisson. Cependant, ceci ne favorise pas nécessairement le développement des bactéries anaérobies. Certaines des bactéries présentes dans le poisson peuvent respirer avec production avantageuse d'ATP en utilisant d’autres molécules comme accepteur d’électrons. C'est ainsi que de plusieurs bactéries spécifiques d’altération du poisson ont la particularité de pouvoir utiliser l'OTMA comme accepteur d’électrons par respiration anaérobie. Le produit de cette réduction, la TMA, est l’un des composants dominants du poisson altéré caractérisé par une odeur typique. Le niveau de TMA, à partir duquel les panels d'évaluation sensorielle rejettent le poisson, varie suivant les espèces mais se situe typiquement autour de 10-15 mg de TMA-N/100 g dans le poisson conservé en aérobiose et à un niveau de 30 mg TMA-N/100 g dans le cabillaud emballé (Dalgaard et al., 1993).
La réduction de l'OTMA est surtout associée au genre de bactéries typiques de l’environnement marin (Alteromonas, Photobacterium, Vibrio et S. putrefaciens), mais est effectuée par Aeromonas et les bactéries intestinales de la famille des Enterobacteriaceae. La réduction de l'OTMA a fait l'objet d'études chez les bactéries anaérobies facultatives et fermentatives comme E. Coli (Sakaguchi et al., 1980) et Proteus spp. (Stenberg et al., 1982) ainsi que par les bactéries non fermentatives de l'espèce S. putrefaciens (Easter et al., 1983; Ringo et al., 1984). Durant la croissance aérobie, S. putrefaciens utilise le cycle de Kreb pour produire les électrons qui sont plus tard canalisés à travers la chaîne respiratoire. Ringo et al.(1984) ont suggéré que pendant la respiration anaérobie, S. putrefaciens utilisait également le cycle complet de Kreb ( Figure 5.12 ), alors que l’on a récemment montré que, dans la respiration anaérobie, une partie seulement du cycle de Kreb est utilisée par S. putrefaciens (Figure 5.13) et les électrons sont également produits par une autre voie métabolique, à savoir la voie de la sérine (Scott et Nealson, 1994). S. putrefaciens peut utiliser plusieurs sources de carbone comme substrat dans sa respiration anaérobie dépendante de l'OTMA, y compris le formate et le lactate. Des composés comme l’acétate et le succinate qui sont utilisés dans la respiration aérobie ne peuvent pas être utilisés quand l'OTMA est l’accepteur final d’électrons (Dichristina et DeLong, 1994). Au contraire, l’acétate est un produit de la réduction anaérobie de l'OTMA (Ringo et al., 1984; Scott et Nealson, 1994).
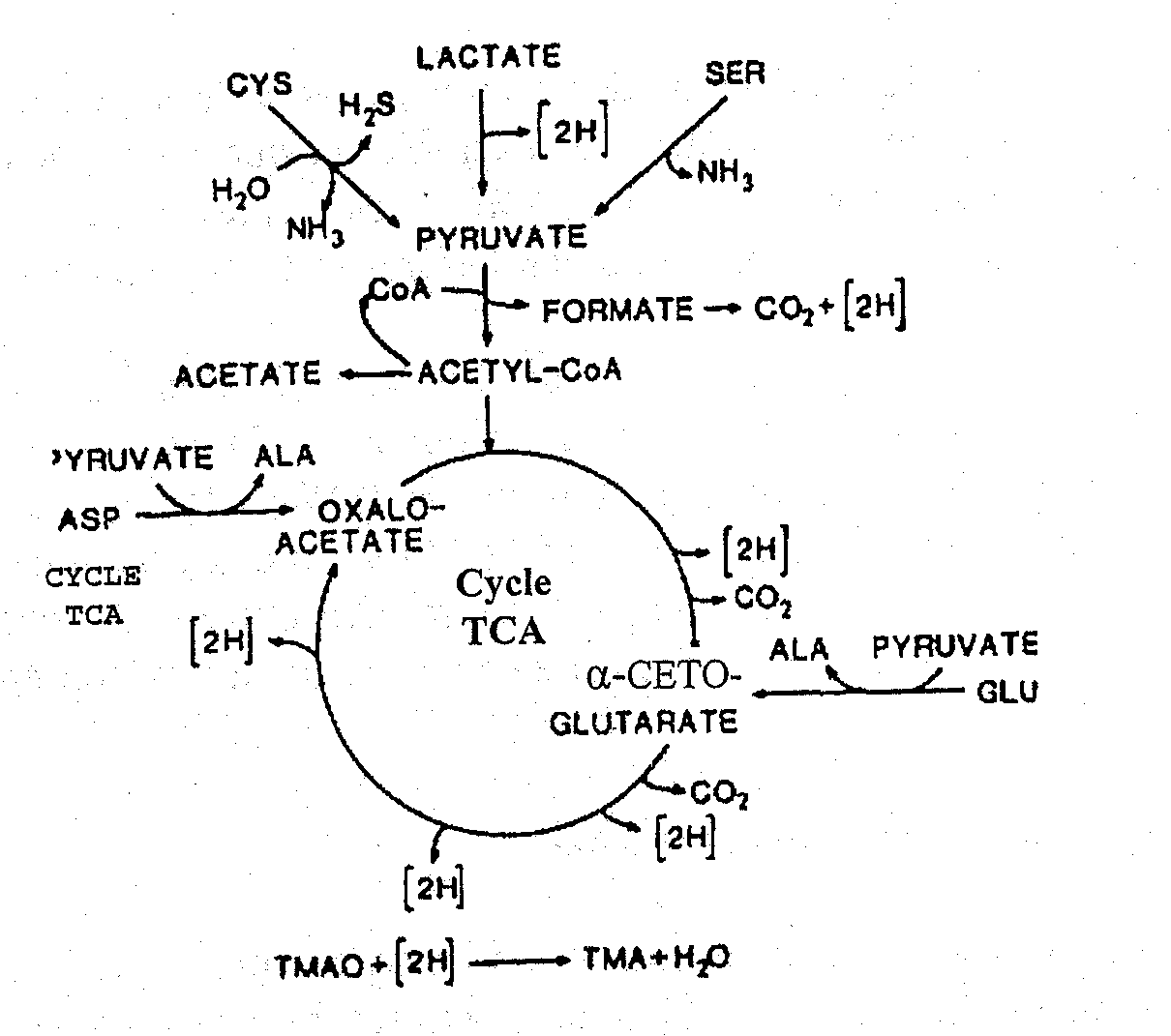
Figure 5.12 Réduction anaérobie de l'OTMA par Shewanella putrefaciens (anciennement Alteromonas) suggérée par Ringo et al.(1984)
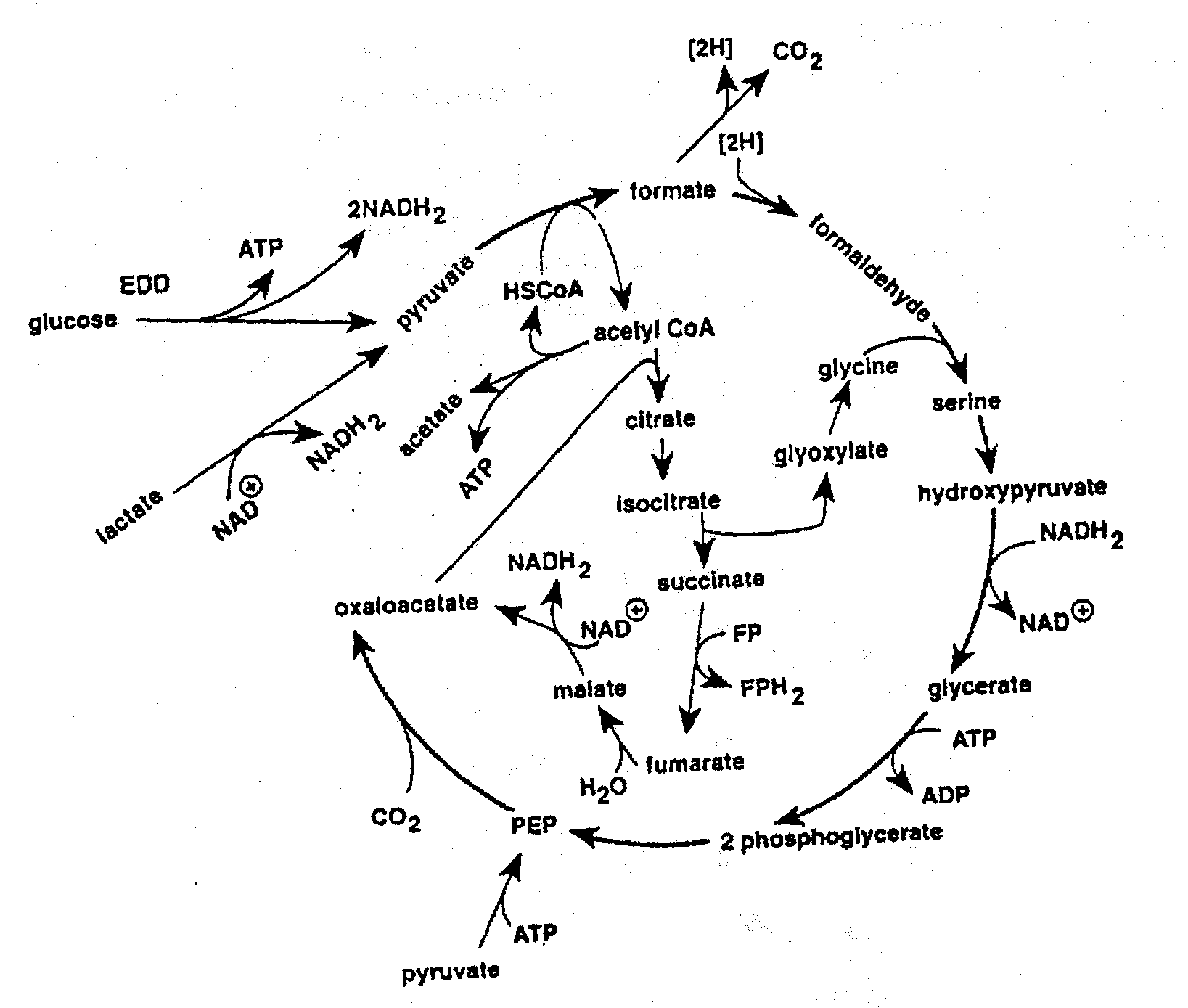
Figure 5.13 Circuit du carbone proposé pendant l’anaérobiose pour S. Putrefaciens (Scott et Nealson, 1994)
A l’opposé, les sucres et le lactate sont les principaux substrats générateurs d'électrons quand Proteus spp. réduit l'OTMA. Cette réduction est accompagnée de production d’acétate principalement (Kjosbakken et Larsen, 1974).
Comme il est mentionné au chapitre 4.4, l'OTMA est un composant typique des poissons marins. Toutefois, il a été récemment rapporté que certains poissons tropicaux d’eau douce contenaient de grandes quantités d’OTMA (Anthoni et al., 1990). Cependant, la TMA n’est pas nécessairement un produit de l'altération de tels poissons puisque celle-ci est due aux espèces de Pseudomonas.( Gram et al., 1990) qui ne réduisent pas l'OTMA.
Le développement de la TMA s'accompagne dans de nombreuses espèces de poissons de la production d’hypoxanthine qui peut, comme on le décrit chapitre 5.2, être formée par la décomposition autolytique des nucléotides. Mais elle peut aussi être formée par les bactéries et le taux de formation bactérienne est plus élevé que celui de l’autolyse. Jorgensen et al. (1988) et Dalgaard (1993) ont constaté une corrélation linéaire entre les teneurs en TMA et en hypoxanthine au cours de la conservation sous glace de cabillaud emballé (Figure 5.14). Plusieurs des bactéries d’altération produisent de l’hypoxanthine à partir d’inosine ou d’inosine monophosphate, y compris Pseudomonas spp. (Surette et al., 1988) S. putrefaciens (van Spreekens, 1977; Jorgensen et Huss, 1989; Gram, 1989) et P. phosphoreum (van Spreekens, 1977).
Dans le cabillaud et autres gadidés, la TMA constitue la plus grande partie de ce que l’on appelle les bases volatiles totales, (également appelée azote basique volatil total ou ABVT) jusqu’à l'altération. Cependant, dans le poisson altéré où l'OTMA est épuisé et où la TMA a atteint son niveau maximal, les niveaux d’ABVT s’élèvent encore du fait de la formation de NH3 et d’autres amines volatiles. De faibles quantités d'ammoniaque se forment aussi au cours des premières semaines de conservation sous glace du fait de l’autolyse. Dans certains poissons qui ne contiennent pas d'OTMA ou dont la dégradation est due à une flore non réductrice de l'OTMA, on observe une faible augmentation d’ABVT pendant la conservation, résultant probablement de la désamination des acides aminés.
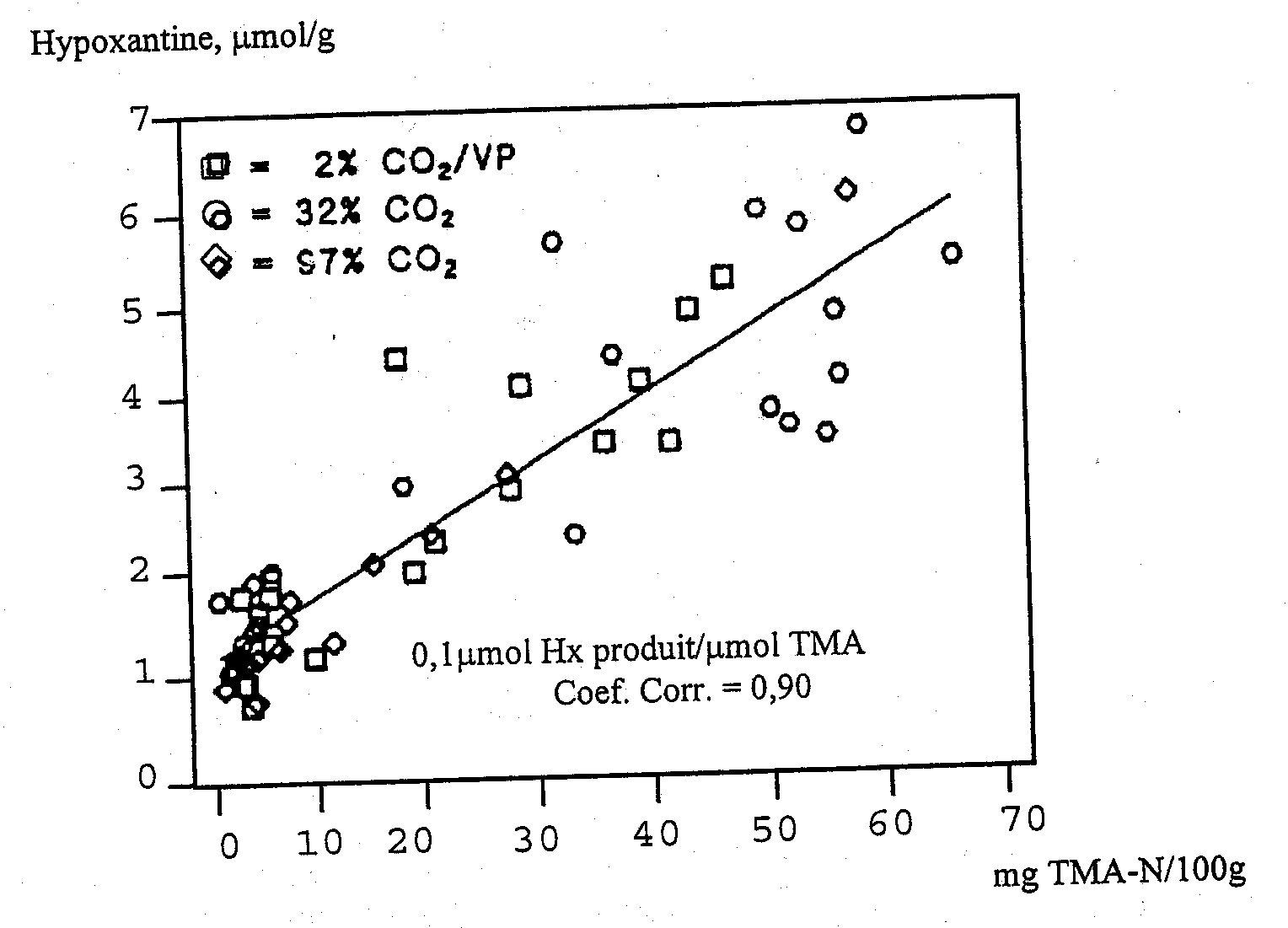
Figure 5.14 Relation entre les taux de triméthylamine (TMA) et d'hypoxanthine (Hx) pendant le stockage de cabillaud emballé sous glace (Dalgaard et al., 1993)
Les composés soufrés volatils sont typiques du poisson en cours d’altération et la plupart des bactéries d’altération produisent un ou plusieurs de ces composés. S. putrefaciens et quelques Vibrionaceae produisent du H2S à partir de la L-cystéine, acide aminé soufré (Stenstroem et Molin, 1990; Gram et al., 1987). Par contre, ni Pseudomonas, ni P. phosphoreum ne produisent de quantités notables de H2S. De ce fait, le sulfure d’hydrogène, qui est typique de l’altération du cabillaud glacé conservé en aérobiose, n’est donc pas produit dans le poisson emballé sous CO2 au cours de son altération (Dalgaard et al., 1993). Le méthylmercaptan (CH3SH) et le diméthylsulfure ((CH3)2 S) sont tous deux formés à partir de l'autre acide aminé soufré, la méthionine. La taurine, qui contient également du soufre, est présente comme acide aminé libre en très forte concentration dans les muscles du poisson, elle disparaît de la chair du poisson au cours du stockage (Figure 5.11) à cause de diffusion plutôt que d’une attaque bactérienne (Herbert et Shewan, 1975). La Figure 5.15 montre les composés qui se forment dans le cabillaud altéré naturellement comparé au muscle stérile.
Les composés soufrés volatils sentent très mauvais et peuvent être détectés à des concentrations très faibles, de l'ordre du ppb, de sorte que même des quantités minimes ont un effet considérable sur la qualité.
Ringo et al. (1984) ont indiqué que la cystéine est utilisée comme substrat dans le cycle de Kreb quand les électrons sont transferés à l'OTMA et les réactions de formation de H2S et de TMA sont donc, dans une certaine mesure, reliées (Figure 5.12).
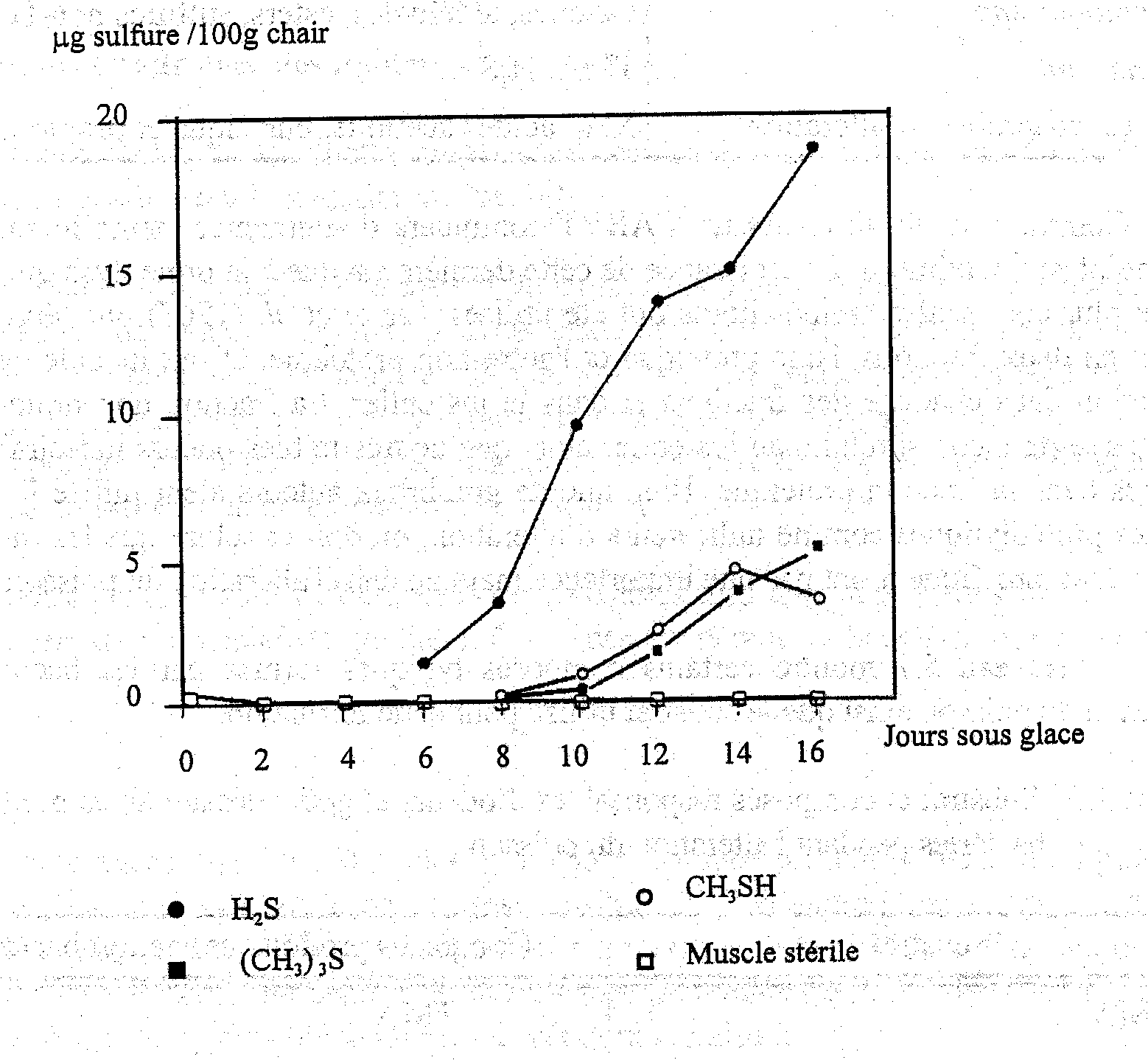
Figure 5.15 Production de H2S, CH3SH et (CH3)2S dans les filets de cabillaud en cours d’altération naturelle et dans les blocs de muscles stériles (Herbert et Shewan, 1976)
Contrairement à la dégradation sous glace par S. putrefaciens et l’altération dans l’air ambiant par les Vibrionaceae qui sont dominées par H2S et TMA, l’altération causée par les espèces de Pseudomonas est caractérisée par l’absence de ces composés (Gram et al., 1989; 1990). Des odeurs et flaveurs fruitées, putrides et sulfydriques sont typiques de l’altération du poisson glacé par Pseudomonas. Ces bactéries produisent plusieurs aldéhydes, cétones, esters et sulfures volatiles (Edwards et al., 1987; Miller et al., 1973a, 1973b). Cependant, les composés responsables des mauvaises odeurs typiques ne sont pas connus (Tableau 5.6). Les mauvaises odeurs fruitées produites par Pseudomonas fragi proviennent des acides aminés mono-aminés et monocarboxyliques.
Tableau 5.6 Composés typiques de dégradation au cours de l’altération de poisson frais conservé en aérobiose et emballé sous glace ou à température ambiante
Organisme spécifique d’altération |
Composés typiques d’altération |
| Shewanella putrefaciens | TMA, H2S, CH3SH, (CH3)2S, Hx |
| Photobacterium phosphoreum | TMA, Hx |
| Pseudomonas spp. | Cétones, aldéhydes, esters, sulfures non-H2S |
| Vibrionaceae | TMA, H2S |
| Bactéries anaérobies d’altération | NH3, acides acétique, burytique et propionique |
Comme il a été dit ci-dessus, l’ABVT continuera d’augmenter même lorsque la
TMA aura atteint son maximum. La croissance de cette dernière est due à la protéolyse
qui commence lorsque plusieurs acides aminés libres ont été utilisés. Lerke et al. (1967)
ont séparé le jus de poisson en deux fractions, l'une protéique et l'autre non
protéique, et ont inoculé des bactéries d’altération dans chacune des fractions
et dans le jus entier. La fraction non protéique du jus s’est altérée de façon
similaire au jus entier alors que de très faibles odeurs indésirables ont été
détectées dans la fraction protéique. Bien que de nombreux auteurs aient utilisé le
nombre de bactéries protéolytiques comme indicateurs d’altération, on doit
conclure que les modifications de la fraction protéique n’ont pas une importance
majeure dans l'altération du poisson frais.
Le Tableau 5.7 montre certains composés typiques formés par les bactéries durant l’altération du poisson ainsi que le substrat utilisé pour cette formation.
Tableau 5.7 Substrat et composés responsables d’odeurs et goûts désagréables produits par les bactéries pendant l'altération du poisson
Substrat |
Composés produits par action bactérienne |
OTMA |
TMA |
| Cystéine | H2S |
| Méthionine | CH3SH, (CH3)2S |
| Carbohydrates et lactate | acétate, CO2, H2O |
| Inosine, IMP | hypoxanthine |
| Amino-acides (glycine, sérine, leucine) | esters, cétones, aldehydes |
| Amino-acides, urée | NH3 |
La formation de TMA s'accompagne d’un dégagement d’ammoniaque pendant la conservation anaérobie de harengs et maquereaux (Haaland et Njaa, 1988). Une conservation en anaérobiose prolongée du poisson donne une importante production de NH3 due à une dégradation plus poussée des acides aminés et à une accumulation d’acides gras plus faibles tels que les acides acétique, butyrique et propionique. Les plus importants producteurs de NH3 se sont révélés être anaérobies strictes appartenant à la famille des bactériodacés, genre Fusobacterium (Kjosbakken et Larsen, 1974; Storroe et al., 1975, 1977). Ces organismes ne se développent que dans l’extrait de poisson altéré et n’ont que peu ou pas du tout d’activité protéolytique dépendant des protéines déjà hydrolysées.
Pendant la conservation sous glace du poisson gras frais, les modifications de la fraction lipidique sont presque exclusivement d'origine chimique, par exemple l’oxydation, alors que l’attaque bactérienne de la fraction lipidique contribue peu à l’altération. Toutefois, l’hydrolyse bactérienne des lipides peut faire partie du processus d’altération pendant le stockage du poisson ayant subi un traitement de préservation léger.
Les deux réactions distinctes impliquant les lipides du poisson et d’intérêt pour l'altération de sa qualité sont l'oxydation et l'hydrolyse.
Il en résulte la production d'une série de substances dont certaines ont une odeur et un goût désagréables (de rance). Certaines peuvent également modifier la texture par des liaisons covalentes avec les protéines des muscles du poisson. Les différentes réactions sont, soit non-enzymatiques, soit catalysées par des enzymes microbiennes ou intracellulaires ou digestives provenant du poisson lui-même. De ce fait, l’importance relative de ces réactions dépend surtout des espèces de poissons et de la température de conservation.
Les poissons gras sont évidemment très sensibles à la dégradation des lipides, ce qui peut créer de sérieux problèmes de qualité, même à des températures de conservation inférieures à 0°C.
Oxydation
De nombreuses fractions d’acides gras polyinsaturés présentes dans les lipides du poisson (voir chapitre 4.2) les rendent très sensibles à l’oxydation selon un mécanisme autocatalytique (Figure 5.16). Ce processus, décrit ci-dessous, est initié par l’extraction d’un atome d’hydrogène du carbone central de la structure pentadiène trouvée dans la plupart des chaînes acyles des acides gras contenant plus d’une double liaison:
-CH =CH-CH2-CH=CH-
Contrairement à la molécule native, le radical lipidique (L.) réagit très rapidement avec l’oxygène atmosphérique formant un radical peroxyde (LOO.) qui, à son tour, peut extraire un hydrogène d’une autre chaîne acyle, ce qui donne un hydroperoxyde lipidique (LOOH) et un nouveau radical L.. Cette préparation se poursuit jusqu’à ce qu’un des radicaux soit éliminé par réaction avec un autre radical ou avec un antioxydant (AH) dont le radical résultant (A.) est beaucoup moins réactif. Les hydroperoxydes produits en quantités relativement importantes pendant la réaction sont sans goût et, par conséquent, il n’est peut-être pas surprenant que l’indice de peroxyde" largement utilisé (chapitre 8.2) n’ait souvent que peu de relations avec les propriétés sensorielles.
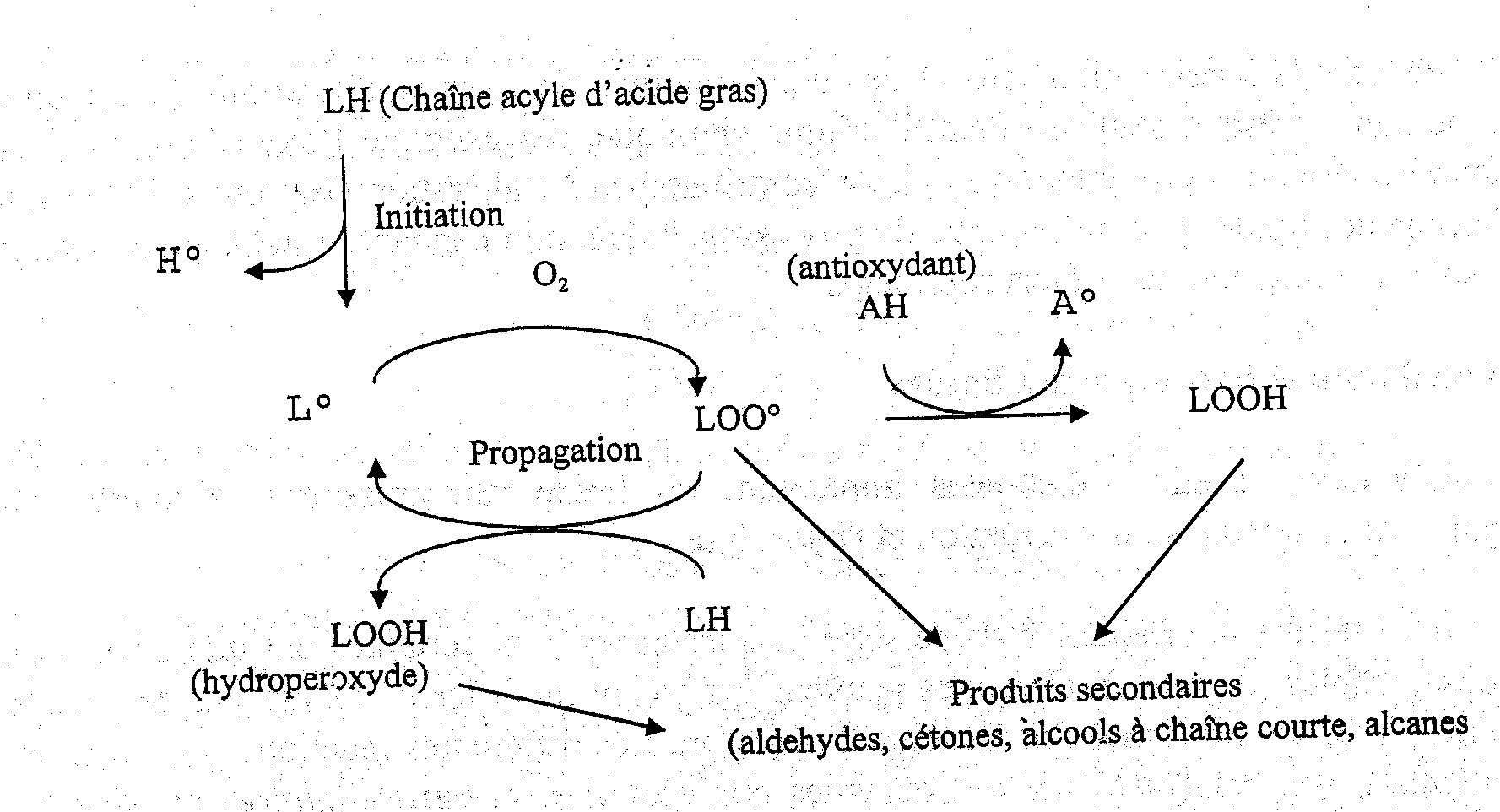
Figure 5.16 Autooxydation des lipides polyinsaturés
Les hydroperoxydes se dégradent facilement. Cette dégradation est catalysée par les ions de métaux lourds, et résulte en produits secondaires d’auto-oxydation de chaîne carbonée plus courte, essentiellement des aldéhydes, des cétones, des alcools, des acides carboxyliques et des alcanes responsables d'un spectre très large d’odeurs et, dans certains cas, d'une coloration jaunâtre. Plusieurs des aldéhydes peuvent être déterminés comme "substances réagissant avec l'acide thiobarbiturique"(chapitre 8.2).
Les ions métalliques jouent un rôle très important durant la première étape de l’auto-oxydation des lipides - l'initiation - en catalysant la synthèse de formes très réactives d’oxygène comme par exemple le radical hydroxyle (OH.). Ce radical réagit immédiatement avec les lipides ou autres molécules sur le site où il est généré. La forte réactivité peut expliquer le fait que les acides gras libres sont plus sensibles à l’oxydation que ceux qui étaient liés, parce que la quantité de fer dans la phase aqueuse est probablement plus importante que celle liée à la surface des membranes cellulaires et aux gouttelettes de lipides.
Les hydroperoxydes d’acides gras peuvent également être formés par voie enzymatique, suite à la catalyse par la lipoxygénase présente en quantités variables dans divers tissus de poisson. On en a constaté une activité relativement élevée dans les branchies et sous la peau de plusieurs espèces. Cette enzyme est instable et est probablement importante pour l’oxydation lipidique uniquement dans le poisson frais. La cuisson ou la congélation/décongélation détruisent assez efficacement son activité.
Les cellules vivantes possèdent plusieurs mécanismes de protection dirigés contre les produits d’oxydation des lipides. Il existe une enzyme, la glutathione peroxydase, qui réduit les hydroperoxydes dans les membranes cellulaires en composés hydroxy correspondants. Cette réaction nécessite un apport de glutathion réduit et par conséquent s’arrêtera post mortem quand cette substance sera épuisée dans la cellule. Les membranes contiennent aussi le composé phénolique a-tocophérol (vitamine E) qui est considéré comme le plus important antioxydant naturel. Le tocophérol peut fournir un atome d’hydrogène aux radicaux L· ou LOO· fonctionnant ainsi comme la molécule AH (Figure 5.16). On estime généralement que le radical tocophéryl résultant réagit avec l’acide ascorbique (vitamine C) à l’interface lipide/eau, régénérant la molécule de tocophérol. D’autres composés, par exemple les caroténoides, peuvent également agir comme antioxydants. La fumée de bois contient des phénols qui peuvent pénétrer la surface du poisson au cours du fumage et donc apporter une protection contre l’oxydation des lipides.
Hydrolyse
Au cours du stockage, une quantité considérable d’acides gras libres (AGL) s'accumule (Figure 5.17). Le phénomène est plus sensible dans le poisson non éviscéré que dans le poisson éviscéré sans doute à cause de l’action d'enzymes digestives. Les triglycérides dans les dépôts de graisses sont scindées par la lipase triglycéride (TL dans la Figure 5.18), issue du tractus intestinal ou excrétée par certains micro-organismes. Les lipases cellulaires peuvent également jouer un rôle mineur.
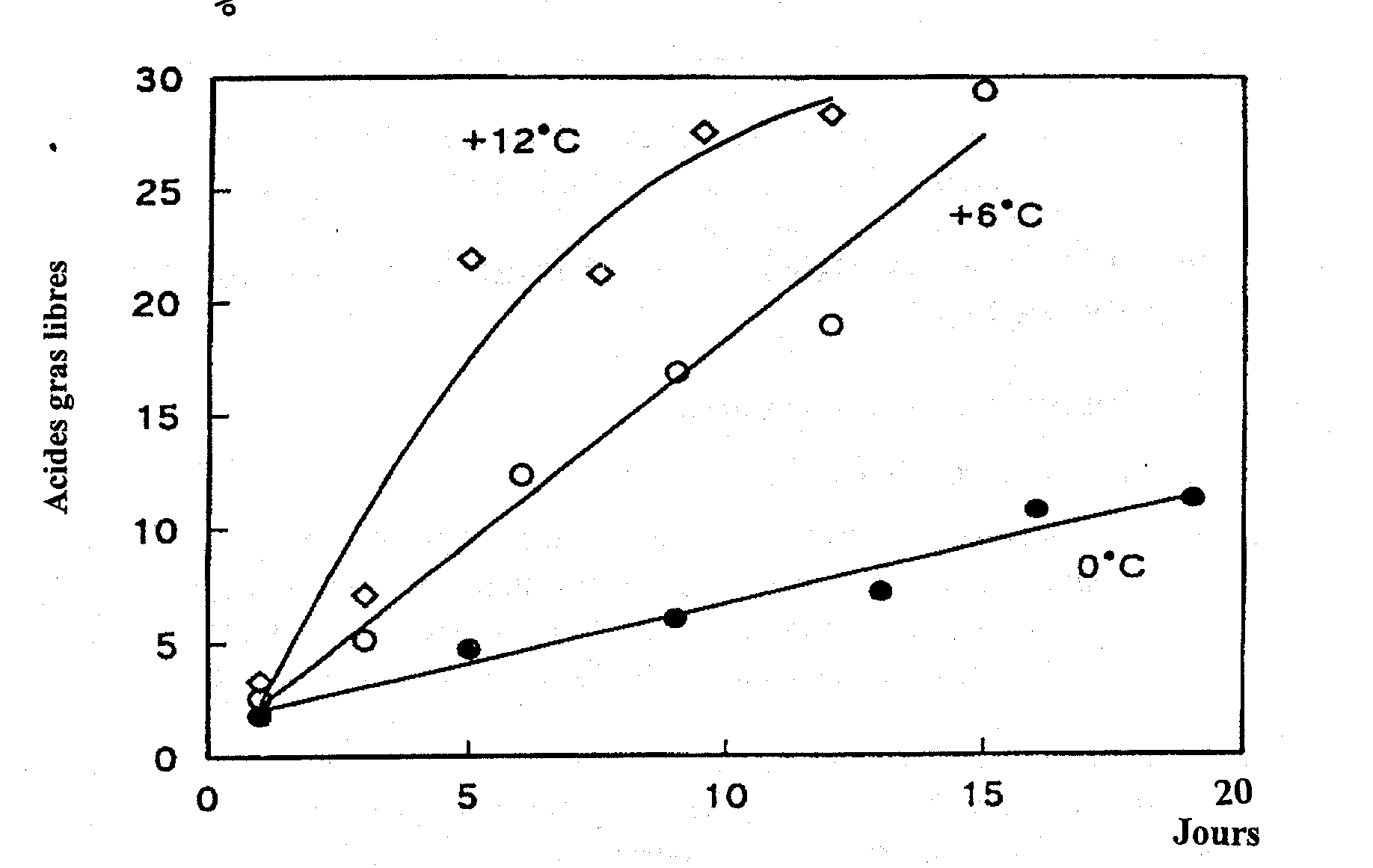
Figure 5.17 Développement des acides gras libres dans le hareng conservé à différentes températures (Laboratoire de Technologie, Ministère des Pêches du Danemark, Rapport annuel, 1971)
Dans le poisson maigre, tel que le cabillaud de l’Atlantique, il y a production d’acides gras libres, même à basses températures. On suppose que les enzymes responsables sont les phospholipases cellulaires - en particulier les phospholipases A2 (PL2 dans la Figure 5.18) - bien que l’on n’ait pas encore établi avec certitude la corrélation entre l’activité de ces enzymes et le taux d’apparition des AGL. Les acides gras liés aux phospholipides au niveau du carbone 2 du glycérol sont surtout du type polyinsaturé, et, par conséquent, l’hydrolyse conduit également souvent à une oxydation plus poussée. De plus, les acides gras eux-mêmes peuvent donner un arrière-goût de savon.
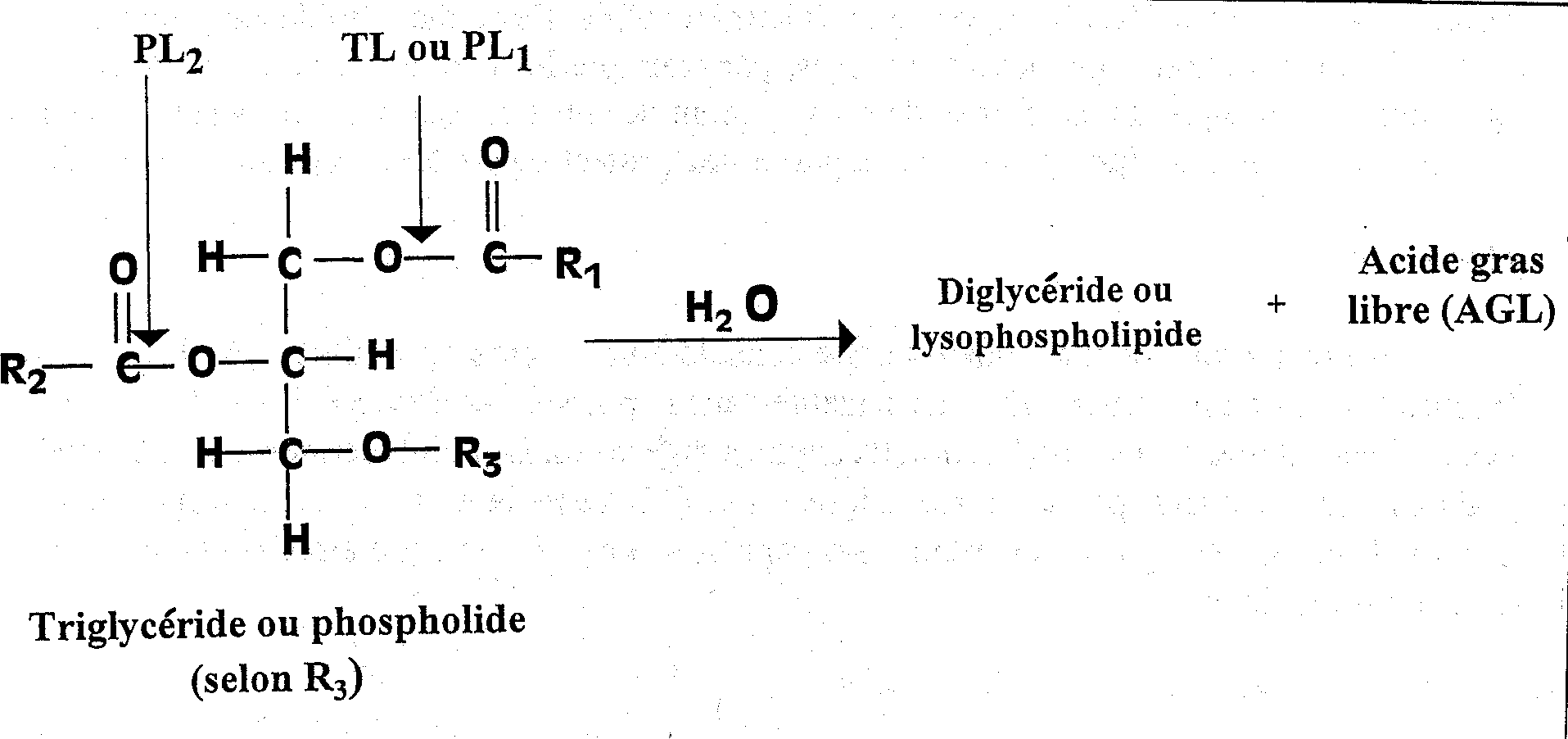
Figure 5.18 Réactions hydrolytiques primaires des triglycérides et phospholipides. Enzymes: PL1 & PL2, phospholipases; TL, lipase triglycéride
![]()
![]()
![]()