![]()
![]()
![]()
P. Quézel, F. Médail, R. Loisel y M. Barbero
Pierre Quézel, Frédéric Médail, Roger Loisel
y Marcel Barbero trabajan en el Instituto Mediterráneo de Ecología y Paleoecología, Marsella, Francia.
Evaluación de la diversidad biológica de los bosques mediterráneos y los problemas que plantea su conservación.
La diversidad vegetal de los bosques mediterráneos, mucho más acusada que la de los bosques europeos, se explica por factores paleogeográficos (Verlaque et al., 1997) e históricos pero también por criterios ecológicos actuales (Quézel, 1985). Por otra parte, el mundo mediterráneo, más que cualquier otra región del mundo, presenta por su flora y por sus paisajes principales estrechas interrelaciones con las actividades humanas que le han dado forma desde hace casi diez milenios (Thirgood, 1981; Pons y Quézel, 1985). De hecho, la biodiversidad vegetal mediterránea resulta, para muchos, de una utilización tradicional y en último término armónica del medio por el hombre. Desde fines del siglo pasado este equilibrio ha sido perturbado en muchas partes, ya se trate de superexplotación o por el contrario de menosprecio, cuyos resultados, sin ser idénticos, no son menos nocivos desde el punto de vista de la conservación de las especies y de los hábitats. Analizando las especies forestales principales o las asociadas a éstas, se examinarán: i) la riqueza en plantas leñosas de las dos zonas mediterráneas del hemisferio norte (California y cuenca del Mediterráneo); ii) el origen biogeográfico de las especies endémicas; iii) el valor patrimonial y las amenazas que pesan sobre las especies y los bosques de la región mediterránea.
Fijación de dunas cerca de Teherán, República Islámica del Irán
Un rodal natural de Quercus suber en Portugal
La riqueza de la biodiversidad del matorral natural en Chipre
RIQUEZA EN ESPECIES FORESTALES: CALIFORNIA Y LA CUENCA DEL MEDITERRÁNEO
Una de las características principales de los bosques mediterráneos, frente a los bosques europeos, es su riqueza en especies arborescentes (árboles o arbustos) (Quézel, 1976). Sin embargo, no se ha hecho un balance preciso de estas especies y su realización plantea algunos problemas, pues es discutible el valor de diversas fanerógamas pequeñas en cuanto especies forestales. Tal es el caso en particular de diversas genistas de gran tamaño (por ejemplo, Cytisus ssp.), ciertas Tamarix o Salix, incluso las representantes de los géneros Cotoneaster, Rhus y Withania. Sólo se han considerado en este inventario las leñosas que pueden alcanzar por lo menos 2 m de altura y de aspecto arborescente. Otra dificultad se presenta cuando se debe decidir la inclusión de ciertas especies entre las preferentes de los bosques de tipo mediterráneo o de tipo europeo, toda vez que un número bastante elevado de árboles existen a la vez en ambos tipos de bosques, sin que sea fácil determinar su medio ecológico óptimo (por ejemplo, Quercus pubescens, Q. pyrenaica y Q. cerris). Se han dejado de lado ciertos taxones ambiguos o de valor discutido, en especial en los géneros Quercus, Tamarix o Salix, siendo las referencias principales Flora Europaea (Tutin et al., 1968-1993) y Med-Cheklist (Greuter, Burdet y Long, 1984-1989).
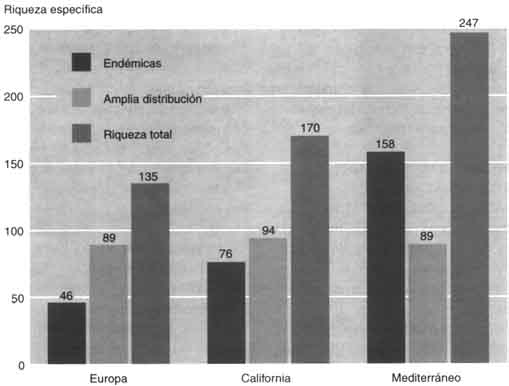
El resultado es que 158 especies y subespecies son exclusivas o muy preferenciales de los bosques mediterráneos, 46 exclusivas o preferenciales de los bosques europeos y 89 están presentes en ambos tipos de bosques. La riqueza en leñosas es pues claramente el doble en los bosques mediterráneos que en los europeos (247 contra 135). Esta disparidad es también perceptible a nivel de los géneros, ya que 34 están presentes solamente en los bosques mediterráneos, contra 7 en Europa; sólo entre las rosáceas ofrecen los bosques europeos un balance específico más elevado que los mediterráneos (con Pyrus, Malus, Sorbus, Mespilus, Crataegus y Prunus). Las fanerógamas (plantas con flores) propias de las riveras lluviales ofrecen las mismas particularidades, siendo los valores anotados respectivamente de 22 leñosas mediterráneas (de ellas una quincena de Tamarix), 7 propias de la región europea y 7 comunes a los dos conjuntos.
El balance de las leñosas de California se ha establecido de acuerdo con The Jepson manual: higher plants of California (Hickman, 1993), pese a ciertas dificultados relativas a la distinción entre árboles y arbustos, en especial en el género Arctostaphyllos. Existen 76 leñosas endémicas de la región mediterránea californiana, mientras que 94 taxones están más ampliamente repartidos. La comparación de los balances de las dos regiones mediterráneas del hemisferio norte muestra que las afinidades ecológicas, biogenéticas e históricas son grandes (Mooney y Dunn, 1970: Quézel y Taylor, 1984). En efecto, los grandes fondos vegetales resultan análogos, sobre todo en cuanto a las leñosas: de 58 géneros de California, 34 están igualmente presentes en torno al Mediterráneo. Además, las perturbaciones climáticas cuaternarias han hecho que desaparezcan del ámbito del Mediterráneo diversas especies todavía presentes en California: Sequoia, Sequoiadendron, Umbellularia, Tsuga, etc. (Roiron, 1992). A las diferencias de superficie entre estos dos conjuntos (2 300 000 km2 en Europa frente a 324 000 km2 en California) corresponde una discordancia entre los balances vegetales (respectivamente 25 000 y 4 400 especies según Quézel) (1985), pero éstos son muy comparables sise cotejan superficies análogas (Médail y Quézel, 1997), por ejemplo Marruecos mediterráneo (300 000 km2 y 4 200 especies) y California mediterránea (324 000 km2 y 4 400 especies). Sin embargo, la región circum-mediterránea presenta una riqueza en elementos arbóreos, incluidos los endémicos, mucho más elevada que los otros dos conjuntos (véase la figura). Sólo los géneros Dendromecon, Sequoia, Sequoia-dendron y Umbellularia son endémicos de California, contra 14 endémicos o subendémicos en la región circum-mediterránea: Argania, Argyrocytisus, Cedrus, Ceratonia, Chamaerops, Fontanesia, Gonocytisus, Hesperola-burnum, Petteria, Podocytisus, Púnica, Securinega, Tetraclinis y Warionia.
VALOR BIOGEOGRÁFICO DE LOS ELEMENTOS FORESTALES ENDÉMICOS DE LA CUENCA DEL MEDITERRÁNEO
La existencia de diversos conjuntos biogenéticos y biogeográficos importantes (Quézel, 1985 y 1995) constituye uno de los factores esenciales para explicar la riqueza de las especies forestales de la región mediterránea. Recientemente se ha dedicado un trabajo a esta cuestión (Barbero et al., 1999), y aquí se ofrece sólo un cuadro general del valor biogeográfico de las principales especies forestales de gran difusión (Cuadro 1), mientras que las especies endémicas se analizan con mayor detalle por su alto valor biogeográfico y las amenazas a menudo muy graves que pesan sobre varias de ellas.
Las especies endémicas de origen eurasiático
Las especies de origen septentrional han dado lugar a algunas endémicas de escaso valor taxonómico como Alnus viridis subsp. suaveolens, en Córcega (Gamisans, 1991), Betula aetnensis neoendémica individualizada en Sicilia a partir de Betula pendula, Betula celtiberica en España centrooccidental y Betula fontqueri en Andalucía y el Rif, diferenciadas respectivamente a partir de B. pubescens y de B. pendula. Quercus petraea subsp. huguetiana se localiza en España septentrional, mientras que otras varias encinas caducifolias, de taxonomía a menudo imprecisa, existen en Sicilia, en Italia meridional, en la región egea o en el Cercano Oriente. Además, buen número de leñosas asociadas, individualizadas en todo el entorno mediterráneo, se encuentran en particular dentro de los géneros Acer, Cotoneaster, Prunus, Pyrus, Salix, Sorbus.
Los elementos endémicos de ámbito restringido
Las endémicas de origen mediterráneo o mesogeo de ámbito restringido son bastante numerosas y ricas en significación biogeográfica (Barbero, Loisel y Quézel, 1995). El caso más notable está constituido por los abetos (Quézel, 1998) con varias especies afines, presentes desde el es trecho de Gibraltar hasta el Líbano (Abies pinsapo, A. marocana incl. A. tazaotana, A. numidica, A. cephalonica incl. A. borisii-regis, A. cilicica), pero también en Anatolia septentrional con los abetos pónticos. Entre las demás coníferas, cabe citar Cupressus atlantica, residual en el Alto Atlas occidental, y Pinus nigra subsp. dalmatica. Entre las frondosas hay que señalar Quercus afares del Maghreb nororiental, Q. euboica en Eubea, Q. vulcanica en Anatolia central y Arbutus pavarii en Cirenaica. Liquidambar orientalis, notable reliquia preglaciar, se localiza en Turquía suroccidental (Akman et al., 1993) y en las islas del Dodecaneso, igual que Quercus aucheri, variante de la encina verde.
Algunos elementos forestales endémicos de origen africano desempeñan un papel marginal (Quézel, 1995), salvo en el suroeste de Marruecos, donde Argania spinosa y Acacia gummifera ocupan un lugar apreciable, asociadas localmente a Olea marocana y a Dracaena draco subsp. ajgal del Antiatlas occidental (Benabid y Cuzin, 1997). Las demás especies endémicas de origen meridional son poco numerosas: Phoenix theophrastii, próxima a Phoenix dactylifera, presente en Creta y en Anatolia suroccidental (Yaltirik y ]Boydak, 1991); Securinega tinctoria se encuentra en el suroeste de la península Ibérica, mientras que Argyrocytisus, Hesperolaburnum y Warionia se limitan a Marruecos. Por último, el género Tamarix individualiza numerosas especies de distribución restringida en situación halohigrófila.
Las islas acogen numerosas leñosas endémicas, en especial en Sicilia: Abies nebrodensis (Morandini, Ducci y Menguzzato, 1994) y Rhamnus lojaconoi, localizadas en los montes de la Madonia, Celtis aetnensis muy próxima a Celtis tournefortii, en el Etna, y Zelkova sicula, endémica residual del monte Lauro (Di Pascale, Garfi y Quézel, 1992). En Creta, Zelkova abelicea está todavía presente en los tres principales macizos montañosos, en poblaciones exiguas. En las rocas verdes de Chipre, Cedrus brevifolia vegeta en el nivel mesomediterráneo (entre 900 y 1 200 m), y Quercus alnifolia se extiende entre 1 200 y 1 900 m.
Inestabilidad genética de las especies constitutivas de los bosques
Gran número de especies forestales mediterráneas se caracterizan por una inestabilidad genética que revela la actividad en ellas de procesos evolutivos y suele conducir a incertidumbres taxonómicas difíciles de dilucidar. De ahí que, entre las encinas esclerófilas, no esté definida la situación precisa de los complejos Quercus ilex (incluido Q. rotundifolia) o Q. coccifera (incluido Q. calliprinos), pese a numerosos estudios bioquímicos o genéticos (Michaud, Lumaret y Romane, 1992). Lo mismo ocurre con los géneros Juniperus, Pinus (Barbero et al., 1998), y Abies (Bergmann y Gregorius, 1992), por no citar más que las gimnospermas en cuyo estudio se ha avanzado más. A esta inestabilidad se deben también las facilidades a menudo considerables que ofrecen muchas especies forestales mediterráneas para la hibridación o la introgresión, en particular en los géneros Quercus, Pinus y Abies. Son así legión los híbridos naturales de Quercus a veces abundantes y dinámicos: por ejemplo Q. streimii (Q. pubescens x Q. sessiliflora) en las riberas de la región mediterránea francesa. Se ha evocado por otra parte la hibridación entre Pinus brutia y P. halepensis, así como entre P. sylvestris y P. uncinata (Barbero et al., 1998). Respecto alas especies o los géneros de ámbito discontinuo, el problema es todavía más evidente y las reforestaciones con abetos mediterráneos originan fenómenos masivos de contaminación genética con los tipos indígenas (Quézel, 1998). Estos fenómenos aparecen por lo demás en muchos otros géneros (Acer, Fagus, Prunus, Sorbus, etc.), y conviene tenerlos en cuenta al introducir especies alóctonas próximas en el plano taxonómico, para evitar graves pérdidas del capital biológico; tal es el caso por ejemplo de las últimas poblaciones de pino negro cevenol (Pinus nigra subsp. salzmanii), profundamente hibridadas a raíz de la introducción masiva de otros tipos de pino negro.
BIODIVERSIDAD Y VALOR PATRIMONIAL DE LAS COMUNIDADES FORESTALES
En un trabajo reciente (Barbero et al., 1999) se ha considerado globalmente este problema, y aquí sólo se destacarán brevemente los resultados más significativos.
Los bosques mediterráneos, sobre todo en las formaciones caducifolias supramediterráneas de África del Norte (Numidia), Córcega y Grecia, presentan una importante riqueza vegetal total. Los valores de 100 especies se superan en particular en buena parte de los bosques de coníferas montañosas mediterráneas, en especial de Cedrus y Pinus nigra. Las únicas formaciones mesomediterráneas esclerófilas que alcanzan esos valores son las estructuras de Quercus calliprinos del Cercano Oriente. Inversamente, las riquezas más débiles se encuentran en particular en las comunidades forestales de las grandes islas mediterráneas: bosques de Acer sempervirens y Cupressus sempervirens de Creta (25 taxones), bosques de Quercus infectoria subsp. veneris de Chipre (27 taxones), o bien en bioclima árido: estructuras de Acacia gummifera del suroeste de Marruecos (25 taxones). La riqueza en fanerógamas puede ser también elevada, en especial en ciertas formaciones caducifolias: castañares corsos (29), encinares caducifolios ligures de Fraxinus ornus (29), de Quercus frainetto de Grecia (26), pero también bosques de cedros y abetos del Cercano Oriente (24-25 taxones).
La presencia de plantas endémicas en los bosques mediterráneos es en general limitada y el porcentaje de endémicas se sitúa entre él 5 y el 10 por ciento. Sin embargo, en algunas regiones pueden encontrarse una veintena de taxones endémicos forestales y el máximo absoluto corresponde a las poblaciones de Juniperus foetidissima en las cimas de las rocas verdes del Troodos de Chipre, donde 20 de los 31 taxones observados son endémicos.
ESPECIES Y COMUNIDADES VEGETALES RARAS O AMENAZADAS
En los países de la parte septentrional de la cuenca del Mediterráneo, el hundimiento del sistema agrosilvopastoral multisecular acarrea profundas modificaciones en la estructura y la arquitectura de las comunidades de plantas forestales y preforestales, y generalmente una maduración de las poblaciones forestales (Barbero et al., 1990). En África del Norte, los trastornos ecológicos derivados de las deforestaciones y el pastoreo excesivo engendran disfunciones graves que afectan en particular a los bosques altos. Pese a las diferencias de los procesos ecológicos y socioeconómicos según las regiones, las disfunciones ocasionadas conducen a consecuencias ecológicas análogas: i) alteraciones de los ciclos naturales de perturbaciones y desequilibrios ecológicos, que generan regímenes de tipo catastrófico en gran escala; ii) homogeneización de las estructuras y arquitecturas de las comunidades vegetales; iii) pérdida neta de diversidad biológica y banalización de la flora, y iv) extensión de plantas exógenas, testigos de la artificialización de los paisajes, que compiten con los vegetales indígenas, incluso en el sotobosque.
Diversos trabajos recientes han tratado de evaluarlos riesgos para el conjunto de la flora mediterránea (Gomez-Campo, 1985; Médail y Quézel 1997), y la Unión Internacional para la Conservación de la Naturaleza (IUCN) ha publicado un balance general relativo a la flora mediterránea según el cual están amenazadas el 53 por ciento de las especies, es decir 1 529 taxones (excluidas la República Árabe Siria, Líbano y Turquía). No obstante, los datos sobre las plantas forestales amenazadas en la región mediterránea son todavía muy fragmentarios. Un trabajo reciente de la UICN (Oldfield, Lusty y McKinven, 1998) establece la lista de las leñosas amenazadas en todo el planeta, pero este inventario no es completo y, por ejemplo, no se toman en consideración numerosos árboles de África del Norte (Olea marocana, Pyrus div. sp., Pinus nigra subsp. mauretanica, etc.), lo mismo que en la 1997 IUCN red list of threatened plants (Walter y Gillett, 1998); el conjunto de estas dos síntesis de la UICN censa 45 leñosas mediterráneas amenazadas. En este contexto, se ha realizado una primera lista de los árboles amenazados de la cuenca del Mediterráneo en la que entran 71 leñosas. La cantidad de endémicas (48 taxones) pone de manifiesto la importancia de la conservación de estos árboles, entre las cuales hay que señalar igualmente el número notable de especies raras progenitoras de vegetales cultivados o que constituyen un fondo genético interesante (Malus, Olea, Phoenix, Prunus, Pyrus).
CUADRO 1. Origen biogeográfíco de las especies forestales mediterráneas y de las principales fanerógamas asociadas1
|
ELEMENTOS FORESTALES DE ORIGEN EUROPEO Muy extendidos (mayoría de las especies) Márgenes septentrionales o dispersos Grandes islas ELEMENTOS FORESTALES DE ORIGEN MEDITERRÁNEO Circum-mediterráneos Mediterráneos occidentales estrictos Mediterráneos occidentales preferenciales Ibero-mauretanos preferenciales Mauretanos Ibéricos Tirrenos Surmediterráneos Mediterráneos orientales estrictos Mediterráneos orientales en sentido amplio Balcánicos Italo-balcánicos Greco-anatolios Cercano-orientales |
1
Excluidas las endémicas; los géneros se han indicado sólo cuando son monoespecíficos en la región mediterránea.
Las comunidades forestales amenazadas de la cuenca del Mediterráneo no han sido objeto hasta ahora de una síntesis precisa, pese a algunos avances (Quézel y Barbero, 1990; Quézel, 1991). Los bosques pueden verse amenazados por razón de: i) sus exigencias ecológicas particulares, en especial la índole del sustrato, que hacen de ellos unos sistemas naturalmente exiguos (como en el caso de ciertos grupos propios de terrenos yesosos o serpentinosos); ii) su situación marginal en el plano geográfico (límites de extensión superficial, grupos aislados), que se explica a menudo por compensaciones bioclimáticas (comunidades abisales) o razones históricas (refugios pleistocenos), como en el caso de la población de Dracaena drago del suroeste de Marruecos (Médail y Quézel, 1999); iii) sobre todo, los efectos intensos y generalizados de la acción del hombre y de los animales sobre estas comunidades, aunque originalmente estuvieran muy repartidos.
En el Mediterráneo septentrional, las amenazas más graves se ciernen sobre las poblaciones residuales de Abies nebrodensis y Zelkova sicula (entre 200 y 250 ejemplares) de Sicilia, mientras que las formaciones de Abies pinsapo, Quercus petraea subsp. huguetiana de España, de Pinus nigra subsp. dalmatica en la ex Yugoslavia, aunque localizadas, parecen algo menos amenazadas. En el litoral mediterráneo francés, los conjuntos termomediterráneos preforestales (formaciones de Ceratonia siliqua y Chamaerops humilis, de Olea y Euphorbia dendroides) y forestales (encinares verdes maduros de Laurus nobilis o Cyclamen balearicum, encinar pubescente de Cyclamen repandum), retroceden por culpa de la urbanización o de la fragmentación, responsable de extinciones locales de especies o del aislamiento de las poblaciones.
Al sur del Mediterráneo, la mayor parte de los sistemas forestales están amenazados en diverso grado; en el Maghreb (Quézel y Barbero, 1990; Quézel, 1991), la situación es particularmente preocupante páralos bosques de abetos de Abies maroccana y A. numidica, las poblaciones de Pinus nigra subsp. mauretanica, Cupressus atlantica, Betula pendula subsp. fontqueri, Olea marocana, Laurus azorica, Quercus afares, Q. faginea subsp. tlemcenensis, e incluso ciertas formaciones de Argania spinosa, Cedrus atlantica, Tetraclinis articulata y Juniperus thurifera.
En el Mediterráneo oriental, la gestión forestal más estricta (sobre todo en Turquía) y la participación de los habitantes locales en los beneficios de la explotación determinan un mejor estado de conservación de los bosques (Quézel y Barbero, 1990); no obstante, ciertas comunidades merecen una atención sostenida: bosques turcos de Abies nordmanniana subsp. equi-trojani, Quercus aucheri, Liquidambar orientalis, Fagus orientalis, formaciones de Cedrus brevifolia y Quercus alnifolia de Chipre, poblaciones difusas de Zelkova abelicea y Phoenix theophrasti de Creta, poblaciones de Abies borisii-regis, Aesculus hippocastanum y Pinus heldreichii.
CONSERVACIÓN Y GESTIÓN PATRIMONIAL
La conservación de los bosques y de las plantas forestales de la cuenca del Mediterráneo constituye un problema complejo, dada la heterogeneidad de las situaciones ecológicas existentes y en vista de los usos múltiples y de las presiones humanas ejercidas por las diversas entidades culturales. La gestión de las diversas fases dinámicas de los sistemas forestales necesita considerar no sólo las comunidades y especies estrictamente forestales, sino también las asociadas a los linderos y a los claros de los bosques. La repartición bastante vasta de la mayoría de las especies forestales lleva a pensar que estas plantas no sufren sino reducidas amenazas potenciales. Ello no obstante, hay que tener también en cuenta la marginalidad corográfica (especies en límite de ámbito de distribución o en espacios aislados) y/o ecológica (especies vinculadas a biotopos raros) de las diversas poblaciones (Rameau y Olivier, 1991). Además, numerosas leñosas constituyen especies clave cuyo mantenimiento es esencial para la perpetuación de los ciclos biogeoquímicos.
Los múltiples regímenes de perturbación han condicionado profundamente la estructura y las sucesiones ecológicas de los bosques mediterráneos, y es preciso garantizar la persistencia, en particular en las zonas protegidas, de tales secuencias de perturbación para asegurar el mantenimiento de un mosaico de comunidades bióticas y de una biodiversidad óptima. Muchas leñosas pueden subsistir en poblaciones aisladas sobre territorios muy restringidos durante varios decenios, incluso más de cien años, mientras persistan las condiciones bióticas. Pero las poblaciones aisladas geográficamente y reducidas son más vulnerables que cualesquiera otras y están expuestas al impacto de la aleatoriedad ambiental. De ahí que Frankel, Brown y Burdon (1995) propongan que el número mínimo de árboles necesarios para mantener una población forestal viable a largo plazo se sitúa en torno a 500. En cambio, la variabilidad genética parece menos vinculada al tamaño de las poblaciones de leñosas. Así, una población muy reducida como la del Abies nebrodensis (29 adultos y 20 planteles según el censo reciente de Morandini, Ducci y Menguzzato, 1994) posee una variabilidad genética análoga a las analizadas en poblaciones dinámicas de Abies alba de Calabria, en situación ecológica similar (Vicario et al., 1995). En realidad, la gran precariedad de esta población residual está en el déficit de producción de semillas, que tienen además una viabilidad muy reducida, y en la naturaleza del biotopo, desfavorable a la regeneración del abeto de Nebrodes (Ducci, Proietti y Favre, 1998).
Rodal de Pinus pinea protegido en el parque nacional de Arabida, en Portugal
El problema de la conservación in situ de los recursos genéticos de las especies forestales mediterráneas gracias a las reservas naturales existentes reside en el hecho de que estas últimas muy rara vez se crean desde esta perspectiva. La designación y la delimitación de las estructuras territoriales de conservación se basan todavía casi siempre en las oportunidades locales o en criterios subjetivos de orden estético, más bien que en análisis rigurosos a diversos niveles de la biodiversidad de un territorio. En el marco de la conservación duradera de las poblaciones de plantas forestales, se observa en general que un archipiélago de pequeñas reservas, aunque puedan contener más especies, no es la solución ideal. En efecto, ciertos taxones como las especies forestales internas se mantendrán únicamente en zonas suficientemente amplias para que su ciclo de vida se desarrolle de manera óptima, sin ser alterado por los efectos de lindero. Sólo las grandes reservas pueden garantizar el desarrollo de ciclos normales de perturbaciones y asegurar el mantenimiento de las poblaciones de grandes vertebrados que garanticen la persistencia de las heterogeneidades del paisaje y de la dispersión de las plantas. Así, para minimizar las pérdidas de diversidad genética y garantizar las potencialidades evolutivas de las especies forestales, parece preferible instaurar sistemas que abarquen vastas reservas integrales (varios cientos de hectáreas como mínimo) rodeadas de zonas aislantes, en mosaico con parcelas gestionadas de manera diferente (Frankel, Brown y Burdon, 1995). Tales estructuras se establecen teóricamente en las «Reservas de la biosfera» de la red El hombre y la biosfera de la UNESCO.
Una estrategia integrada de conservación de los taxones forestales amenazados o considerados de interés primordial requiere pues ante todo un buen conocimiento de la ecología propia y de la biología de las especies, lo que rara vez se da en la actualidad (desconocimiento casi total, por ejemplo, de las especies clave). Debe prestarse la mayor atención a las especies endémicas, por su unicidad y su distribución restringida, y a las poblaciones marginales. Pero el mantenimiento de la capacidad de adaptación de las especies por la presión de selección implica obligatoriamente una conservación in situ que presupone una gestión dinámica de los mosaicos del paisaje forestal que deban integrar las eventuales operaciones de silvicultura.
Bibliografía
Akman, Y., Quézel, P., Ketenoglu, O. y Kurt, I. 1993. Analyse syntaxonomique des forêts de Liquidambar orientalis en Turquie. Ecologia mediterranea, 19: 49-57.
Barbero, M., Bonin, G., Loisel, R. y Quézel, P. 1990. Changes and disturbances of forest ecosystems caused by human activities in the western part of the Mediterranean Basin. Vegetatio, 87: 151-173.
Barbero, M., Loisel, R., Médail, F. y Quézel, P. 1999. Signification biogéographique et biodiversité des forêts du bassin méditerranéen. Bocconea, en prensa.
Barbero, M., Loisel, R. y Quézel, P. 1995. Les essences arborées des îles méditerranéennes: leur rôle écologique et paysager. Ecologia mediterranea, 20 (1/2): 53-69.
Barbero, M., Loisel, R., Quézel, P., Richardson D.M. y Romane, F. 1998. Pines of the Mediterranean Basin. En D.M. Richardson, ed. Ecology and biogeography of Pinus, p. 153-170. Cambridge, Reino Unido, Cambridge University Press.
Benabid, A. y Cuzin, F. 1997. Populations de dragonnier (Dracaena draco L. subsp. ajgal Benabid et Cuzin) au Maroc: valeurs taxinomique, biogéographique et phytosociologique. C. R. Acad. Sci. Paris, Sciences de la vie, 320: 267-277.
Bergmann, F. y Gregorius, H.R. 1992. Ecogeographical distribution and thermostability of isocitrate deshydrogenase (IDH) alloenzymes in European silver fir (Abies alba). Biochem. Syst. Ecol., 21: 597-605.
Di Pascale, G., Garfi, G. y Quézel, P. 1992. Sur la présence d'un Zelkova nouveau en Sicile sud-orientale (Ulmaceae). Biocosme Mésogéen, 8(4)-9, (1): 401-409.
Ducci, F., Proietti, R. y Favre, J.M. 1998. Le genre Abies en Italie: écologie générale, gestion sylvicole et ressources génétiques. Forêt médit., 19(2): 153-164.
Frankel, O.H., Brown, A.H.D. y Burdon, J.J. 1995. The conservation of plant biodiversity. Cambridge, Reino Unido, Cambridge University Press. 299 pp.
Gamisans, J. 1991. La végétation de la Corsé. En D. Jeanmonod y H.M. Burdet, eds. Compléments au prodrome de la flore corse, annexe 2. Ginebra, Conserv. et Jard. bot. Genève. 391 pp.
Gomez-Campo, C., ed. 1985. Plant conservation in the Mediterranean area. Geobotany 7. Dordrecht, Países Bajos, W. Junk. 269 pp.
Greuter, W., Burdet, H.M. y Long, G. 1984-1989. Med-Checklist. 3 vols. Ginebra, Conserv. et Jard. bot. Genève.
Hickman, J.C., ed. 1993. The Jepson manual. Higher plants of California. Berkeley y Los Angeles, Estados Unidos, University of California Press.
Médail, F. y Quézel, P. 1997. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean basin. Ann. Missouri Bot. Gard., 84: 112-127.
Médail, F. y Quézel, P. 1999. The phytogeographical significance of S.W. Morocco compared to the Canary Islands. Plant Ecol., en prensa.
Michaud, H., Lumaret, R. y Romane, F. 1992. Variation in the genetic structure and reproductive biology of holm oak populations. Vegetatio, 99/100: 107-113.
Mooney, H.A. y Dunn, E.L. 1970. Convergent evolution of Mediterranean-climate evergreen sclerophyll shrubs. Evolution, 24: 292-303.
Morandini, R., Ducci, F. y Menguzzato, G. 1994. Abies nebrodensis (Lojac.) Mattei, inventario 1992. Ann. Ist. Sperim. Selvic. Arezzo, 22: 5-51.
Olfield, S., Lusty, C. y McKinven, A. 1998. The world list of threatened trees. Cambridge, Reino Unido, IUCN Press. 650 pp.
Pons, A. y Quézel, P. 1985. The history of the flora and vegetation and past and present human disturbance in the Mediterranean region. En C. Gomez-Campo, ed. Plant conservation in the Mediterranean area. Geobotany 7, p. 25-43. Dordrecht, Países Bajos, W. Junk.
Quézel, P. 1976. Les forêts du pourtour méditerranéen. En Forêts et maquis méditerranéens: écologie, conservation et aménagements. Note technique MAB, 2:9-33. París, UNESCO.
Quézel, P. 1985. Definition of the Mediterranean region and the origin of its flora. En C. Gomez-Campo, ed. Plant conservation in the Mediterranean area. Geobotany 7, p. 9-24. Dordrecht, Países Bajos, W. Junk.
Quézel, P. 1991. Structures de végétation et flore en Afrique du Nord: leurs incidences sur les problèmes de conservation. En M. Rejdali y V.H. Heywood, eds. Conservation des ressources végétales. Rabat, Actes Editions, Institut agronomique et vétérinaire Hassan II.
Quézel, P. 1995. La flore du bassin méditerranéen: origine, mise en place, endémisme. Ecologia mediterranea, 20(1/2): 19-39.
Quézel, P. 1998. Diversité et répartition des sapins sur le pourtour méditerranéen. Forêt médit:, 19(2): 93-104.
Quézel, P. y Barbero, M. 1990. Les forêts méditerranéennes. Problèmes posés par leur signification historique, écologique et leur conservation. Acta Bot. Malacitana, 15: 145-178.
Quézel, P. y Taylor, H.C. 1984. Les fruticées sempervirentes des régions méditerranéennes de l'Ancien Monde. Essai comparé d'interprétations des structures biologiques et des données historiques. Bot. Helv., 94: 133-144.
Rameau, J.C. y Olivier, L. 1991. La biodiversité forestière et sa préservation. Intérêt patrimonial de la flore, de la végétation et des paysages forestiers. Rev. For. Fr., 43, num. sp. 1991: 19-27.
Raven, P.H. y Axelrod, D.I. 1978. Origin and relationships of the California flora. Univ. California Public. Bot., 72: 1-134.
Roiron, P. 1992. Flores, végétations et climats du Néogène méditerranéen: apports de macroflores du sud de la France et du nord-est de l'Espagne. Univ. Montpellier II. 296 pp. (tesis).
Thirgood, J.V. 1981. Man and the Mediterranean forest. Nueva York, Academic Press. 194 pp.
Tutin, T.G. et al. 1968-1993. Flora Europaea. Vols 1-5. Cambridge, Reino Unido, Cambridge University Press.
Verlaque, R., Médail, F., Quézel, P. y Babinot, J.F. 1997. Endémisme végétal et paléogéographie dans le bassin méditerranéen. Geobios, mém. sp. 21: 159-166.
Vicario, F., Vendramin, G.G., Rossi, P., Lio, P. y Giannini, R. 1995. Allozyme, chloroplast DNA and RAPD markers for determining genetic relationships between Abies alba and the relict population of A. nebrodensis. Theor. Appl. Genet., 90: 1012-1018.
Walter, K.S. y Gillett, H.J., eds. 1998 1997 IUCN Red List of threatened plants. Compilado por World Conservation Monitoring Centre. Gland, Suiza y Cambridge, Reino Unido, IUCN-The World Conservation Union.
Yaltirik F. y Boydak, M. 1991. Distribution and ecology of the palm Phoenix theophrasti (Palmae) in Turkey. Bot. Chron., 10: 869-872.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}