![]()
![]()
![]()
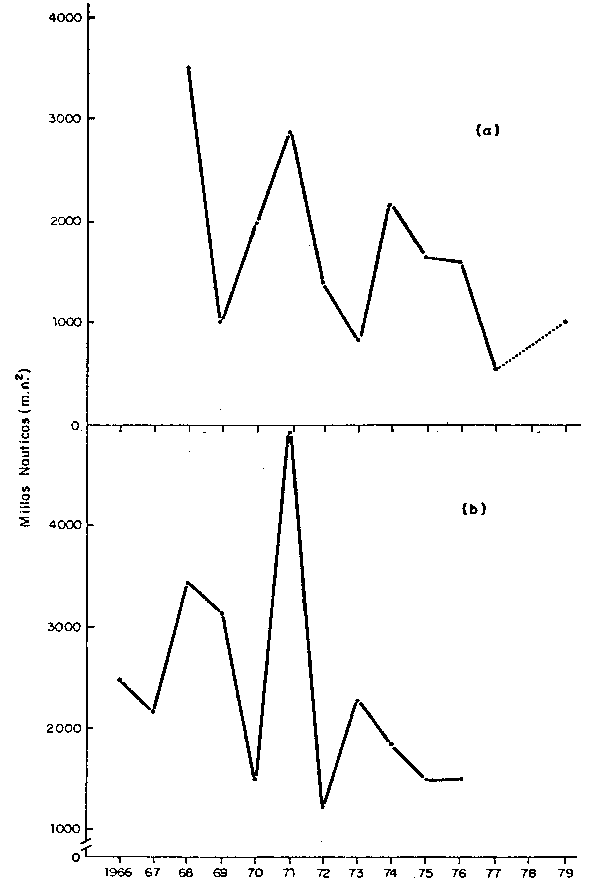
Fig. 30 Cambios en la extension del area habitoda por la Anchoveta, 1966-79: (a) Verano, (b) Inviero (m.n.2. promedio por grodos de latitud).
Fig. 30 Changes in the extent of area inhabitated by Achoveta, 1966-79 (a) Summer, (b) Winter (averoge by latitude degree)
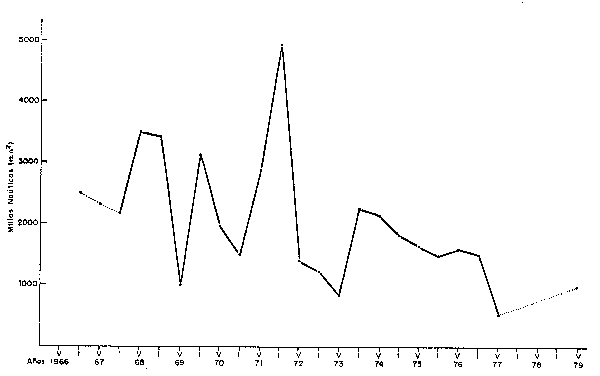
Fig. 31 Cambios en la extensión del área habltado por la Anchoveta, 1966-1979. (m.n.2 promedlo por grodos de latitud) Changes in the extent of the area inhabitated by Anchoveta, 1966-79 (average by latitude degree).
Para calcular el índice de concentración se han empleado los datos de abundancia de las prospecciones Eureka considerando las categorías disperso, denso y muy denso (categorías 2-4) que según Johannesson y Robles (1977) corresponden a densidades de 430, 1550 y 5300 TM/mn2 para las categorías mencionadas, respectivamente, relacionándolas con la extensión de dichas áreas, tal como se muestra en la Fig. 32 y Tabla 1. Los puntos graficados en la parte superior izquierda indican por lo tanto, los meses de máxima concentración y que corresponden a períodos con calentamientos apreciables; en contraste, los puntos situados en la parte inferior derecha indican los meses de mínima concentración de cardúmenes de anchoveta, que corresponden a los meses fríos y normales. En la Fig. 33 se han graficado los valores de estos índices de concentración en secuencia de tiempo.
Johannesson y Vílchez (1981) empleando datos de ecointegración indican que en el crucero de febrero 1973, el 55% de la biomasa estuvo concentrada en sólo 13% del área total de distribución y en el crucero de marzo 1976, el 36% de la biomasa se concentró en sólo 3.4% del área total.
Los índices altos de concentración generan un incremento en la densidad total, produciendo una mayor interacción entre las mismas anchovetas y entre la anchoveta y sardina que comparten el mismo habitat, creando un estado de tensión que a su turno conducen a una declinación del potencial reproductivo, activando a su vez un conjunto de reacciones corporales.
Las implicancias de este fenómeno así como los posibles efectos dependientes de la densidad, han sido discutidas por Csirke (1980), Sharp (1981), MacCall (1981), Tsukayama y Alvares (1981), Ware y Tsukayama (1981).
La dramática reducción en el peso corporal de las anchovetas adultas registrada en El Niño de 1982-83 (Fig. 34), en un 27% con respecto al período 1961-64, cuando la magnitud de la población era alta y las condiciones ambientales estables, evidencia una vez más el estado de intenso stress en que se encuentra el stock, por efecto de factores tanto intrínsecos como extrínsecos.
En Niños anteriores se observó también reducción en el peso, como una respuesta a un ambiente inapropiado, que fueron de 22% en marzo de 1976, 15% en octubre del mismo año y 11% en abril de 1977, atribuíbles a una insuficiente cantidad de alimento disponible, bajo situaciones de altas concentraciones del stock (IMARPE 1977).
Otra reacción observada, es la referente a los altos contenidos grasos que exhibieron las anchovetas, meses antes del desarrollo pleno de los Niños 1972-73 y 1982-83, aún cuando bajo circunstancias diferentes. Previo el Niño 1972-73 (agosto y setiembre 1981) el alto contenido de grasa estuvo relacionado con una actividad reproductiva reducida (IMARPE, 1972), mientras que previo al Niño 1982-83 (febrero-marzo 1982) la actividad reproductiva era apreciable (IMARPE, 1982). Los mecanismos fisiológicos de este proceso no están totalmente esclarecidos, pero estas observaciones pueden interpretarse como una reacción anticipada del stock frentè a condiciones desfavorables del ambiente.
Cambios en la Composición por Especies
En la década de los años 1950 y 1960 y hasta antes de la ocurrencia del Niño 1972, la anchoveta fue el recurso pesquero nerítico pelágico dominante del gran complejo de la corriente peruana. Esto es evidente del análisis de las descargas de la pesca comercial que operó con redes de cerco (boliche), donde la proporción de esta especie constituyó más del 95% del total capturado de especies pelágicas en la costa peruana. Los análisis de restos de peces en los sedimentos marinos sugieren también una dominancia histórica de la anchoveta en la comunidad de peces pelágicos frente al Perú (De Vries y Pearcy, 1982).
Es a partir de 1972 cuando se aprecia tanto en las capturas comerciales, así como en las prospecciones efectuadas por IMARPE, una disminución en la proporción de anchoveta y un paulatino incremento de las otras especies pelágicas, constituyendo capturas mixtas.
Como se ha mostrado también en las Figs. 28 y 29 y a fin de cuantificar estos cambios en la composición por especies, se han usado datos de las prospecciones científicas, los cuales han sido agrupados tanto a nivel Perú como por zonas (Figs. 35, 36, 37, 38 y 39). En todas las zonas es notoria la disminución en la proporción de anchoveta y un incremento en la sardina; para el jurel y la caballa este incremento también se da, aunque es menos conspicuo, en parte debido a los aparejos de pesca usados en estas prospecciones.
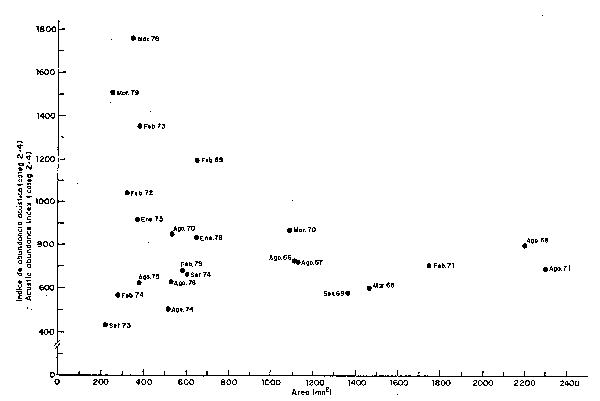
Fig. 32 Relaclón entre la abundancia y extensión de las áreas, con cardúmenes dispersos, densos y muy densos (categ 2-4) Relationship between anchoveta abundance and area extent inhabitated by disperse, dense and very dense shoals (categ. 2-4)
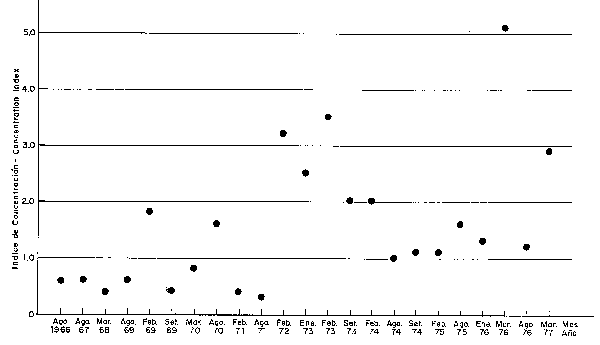
Fig. 33 Variaciones en el indice de concentración del stock de Anchoveta durante los años 1966-1977. Canges in the concetration index of the Achoveta stock during 1966-1977.
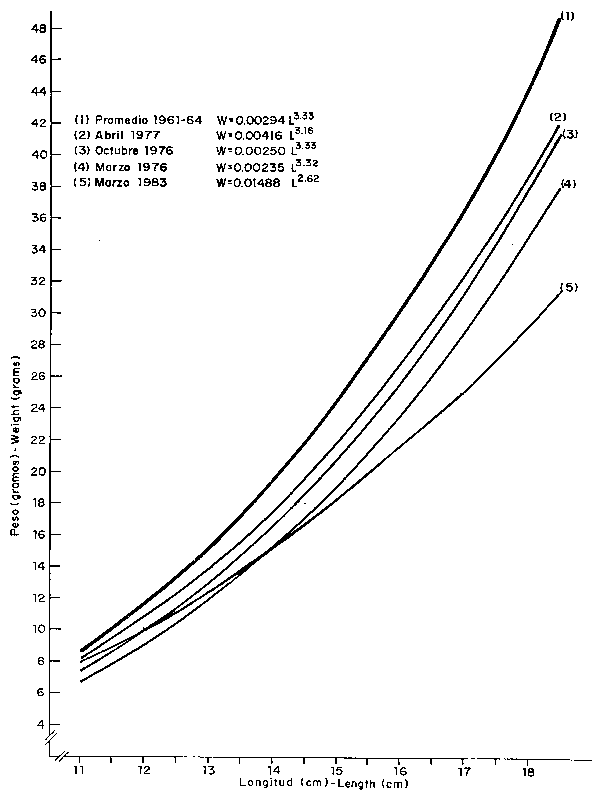
Fig. 34 Variaciones en la relación longitude-peso de Anchoveta. Variationes in the Anchoveta lenght-weight relationship.

Fig. 35 Composición por especies pelágicas (%) por estaciones y años, en la costa peruana. (Datos de calas de
cruceros acústico-pesqueros y eurekas.)
Pelagic species composition (%) along peruvian coast by seasons and years. (Data from hauls made during Eurekas and Cruises
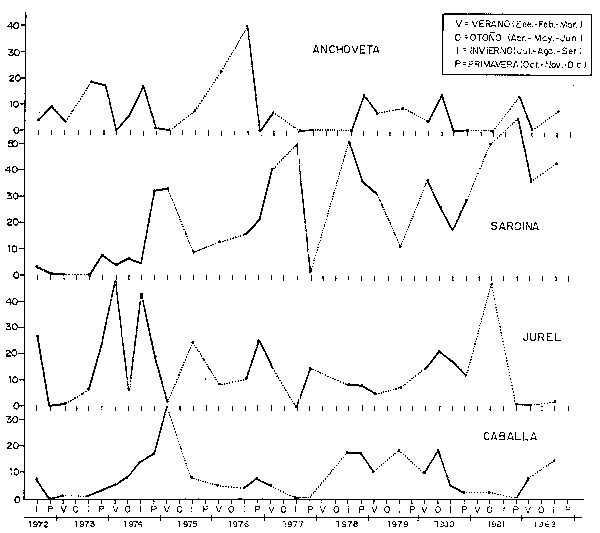
Fig. 36 Composición por especies pelagicas (%) por estaciones y años, en la zona comprendida entre 3°y 7°S:Frontero Norte a Pimentel. (Dotos de calas de cruceros ocústico-pesqueous y eurekas). Pelagic species composition (%) by seasons and years at 3°-7°S: Frontera Norte-Pimentel. (Data from hauls made during Eurekas and Cruises.)
El cerco anchovetero empleado, proporciona informaciones más representativas de la anchoveta y sardina que habitan fundamentalmente en las capas superficiales, formando agregaciones densas y altamente vulnerables a estas redes, mientras que el jurel y la caballa se encuentran también en capas más profundas y conformando cardúmenes más dispersos y en consecuencia, menos vulnerables a dichas redes.
En la Tabla 2, se puede apreciar que la disminución en la proporción de anchoveta fue más drástica en el área de Pimentel-Huarmey (7-10°S) en los años 1972-77, con un valor de 29%, siendo de 43% y 31% para las áreas de Huarmey-Pisco (10-14°S) y Pisco-Frontera sur (14-19°S), respectivamente. En el período 1978-82 continúa el descenso en la proporción de anchovetas con valores de 26%, 27% y 16% para las tres áreas mencionadas, respectivamente. Cabe mencionar que el área más norteña de la costa peruana, tradicionalmente no ha sido productiva una zona de reproducción de anchoveta por las características ambientales que imperan en esa región. Las zonas comprendidas entre los 6-14°S (Paita-Pisco), características por las de la plataforma continental y los intensos y permanentes afloramientos son los que propiciaron que una altísima producción de esta especie, como se detalla en las secciones anteriores.
El incremento de la proporción de sardina es notoria en todas las zonas y aún cuando ya se aprecia desde 1972, es más conspicua en el período 1978-1982. La sardina antes del Niño 1972 ocupó principalmente el área septentrional de la costa peruana, aunque en proporciones muy reducidas, por esta razón dicha área se constituyó en la base de las actividades de la flota artesanal y conservera, cuyo radio de acción comprendió los puertos de Paita, Chicama y Chimbote.
A nivel de toda la costa, la proporción de anchoveta ha disminuido del 32% en el período 1972-77 al 20% en los años 1978-82; en estos mismos períodos la sardina, jurel y caballa se incrementaron de 6 a 23%, de 11 al 16% y del 5 al 8%, respectivamente.
Los cambios en la proporción de especies que aquí se presentan, guardan correspondencia con las informaciones dadas por Santander (1981) quien indica que en el período 1961-71 los huevos y larvas de anchoveta dominaron en el ictioplancton con una proporción que varió entre el 65-90%; en 1972-73 disminuye al 47-50%, y desde 1974 a 1980 la proporción ha variado notablemente, registrándose sólo un 20% de huevos de anchoveta en septiembre de 1979.
Cambios en la Abundancia de las Poblaciones Pelágicas
Con respecto a la anchoveta: Los estimados de biomasa de anchoveta efectuados por IMARPE a lo largo de la costa peruana en el período 1963-81, fueron sumarizados por Tsukayama (1983, en prensa), donde se aprecia que esta población ha sufrido grandes fluctuaciones en el período en mención, con un máximo de 22 millones de toneladas métricas en 1967 y mínimos de 2-3 millones de toneladas en los últimos años (Fig. 40).
La declinación más drástica ocurrió en 1972 coincidente con el Niño de ese año, en que de 15.0 millones de toneladas que se tuvo como promedio en los años previos de 1970-71, desciende hasta 9.0 millones de toneladas a comienzos de 1972 y a 4.1 a fines del mismo año. En los años 1973-75 la biomasa promedio es de 5.6 millones de toneladas; a comienzos de 1976 se incrementó a 10.6 millones de toneladas y a fines de 1976 y comienzos de 1977 con la ocurrencia nuevamente de transtornos en el ambiente, la población disminuye aún más, hasta alrededor de 3.0 millones de toneladas; en 1978-81 la biomasa promedio es de 2.7 millones de toneladas.
A fin de explicar esta dramática reducción del stock de anchoveta e inferir en qué medida ello se ha producido como consecuencia de la presión de la pesquería de los cambios en el ambiente durante este período, Tsukayama (1983, en prensa) estimó tasas de explotación calculadas de la proporción entre las capturas (en peso) y las biomasas. Los valores promedios obtenidos fueron de 0.47 para los años 1963-66, 0.55 en los años 1967-69 y 0.77 en los años 1970-71 (Tabla 3).
Esta altísima tasa de explotación en los años inmediatamente antes del Niño 1972, operó fundamentalmente sobre las anchovetas más pequeñas que se hacían altamente vulnerables a la pesca durante los meses de verano que corresponden al período de reclutamiento. Los datos presentados por IMARPE (1972) y que se refieren al seguimiento de las clases anuales 1962 a 1971 durante su fase explotable, indican que la flota capturó proporciones cada vez mayores de ejemplares en su primer año de vida, en relación con el rendimiento total de la respectiva clase anual. Así por ejemplo, la clase anual 1962 rindió 6.48 millones de toneladas durante su paso por la pesquería, de los cuales el 46% fue de ejemplares con menos de un año de edad; en los años 1970 y 1971 dicha proporción se incrementó a 94% y 80%, respectivamente. Estas evidencias sugieren que en los anos previos al Niño 1972, las altas tasas de explotación provocaron una "sobrepesca del crecimiento" en el stock de anchoveta, eliminando la oportunidad que los peces pequeños logren su potencial de crecimiento, a diferencia de lo sugerido por Clark (1977) citado por Bakun y Parrish (1981), quien expresa "la posibilidad que el colapso del stock de anchoveta, el cual coincidió con El Niño 1972, pudo deberse a una insuficiente pesca en 1971, así como a una excesiva pesca en 1972".
| COMPOSICION FOR ESPECIES (I) | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ZONA | ASOS | VERANO | OTRO | INVIERNO | PRIMAVERA | VER.+ OTO. + INV. + PRIM. | |||||||||||||||||||||
| Anch. | Sard. | Jur. | Cab. | Otroas | Anch. | Sard. | Jur. | Cab. | Otroas | Anch. | Sard. | Jur. | Cab. | Otroas | Anch. | Sard. | Jur. | Cab. | Otroas | Anch. | Sard. | Jur. | Cab. | Otroas | |||
| Frontara Morte-Pimentel: | 1972 | - | - | - | - | - | - | - | - | - | - | 4 | 3 | 27 | 7 | 59 | 9 | 0 | 0.1 | 0 | 91 | 5 | 2 | 20 | 5 | 68 | |
| 3-7°S | 1973 | 3 | 0 | 0.1 | 0.4 | 96 | - | - | - | - | - | 19 | 0.1 | 6 | 0.1 | 74.8 | 18 | 7 | 24 | 3 | 48 | 12 | 2 | 2 | 1 | 78 | |
| 1974 | 0 | 4 | 48 | 5 | 43 | 6 | 6 | 6 | 8 | 74 | 17 | 5 | 43 | 14 | 21 | 0.7 | 32 | 19 | 17 | 69 | 6 | 16 | 19 | 12 | 47 | ||
| 1975 | 0 | 33 | 2 | 35 | 30 | - | - | - | - | - | 8 | 9 | 24 | 8 | 51 | - | - | - | - | - | 7 | 13 | 20 | 13 | 47 | ||
| 1976 | 23 | 13 | 8 | 5 | 51 | - | - | - | - | - | 40 | 16 | 10 | 4 | 30 | 0 | 21 | 25 | 7 | 47 | 25 | 15 | 10 | 5 | 45 | ||
| 1977 | 6 | 41 | 16 | 5 | 32 | - | - | - | - | - | 0 | 50 | 0 | 0 | 50 | 0 | 2 | 14 | 0 | 84 | 3 | 30 | 13 | 3 | 51 | ||
| 1978 | - | - | - | - | - | - | - | - | - | - | 0 | 51 | 8 | 17 | 24 | 14 | 36 | 8 | 17 | 25 | 12 | 38 | 8 | 17 | 25 | ||
| 1979 | 7 | 32 | 5 | 10 | 46 | - | - | - | - | - | 9 | 11 | 7 | 18 | 55 | - | - | - | - | - | 8 | 20 | 6 | 15 | 51 | ||
| 1980 | 4 | 37 | 14 | 10 | 35 | 14 | 26 | 21 | 18 | 21 | 0 | 18 | 17 | 5 | 60 | 0.7 | 29 | 12 | 2 | 56 | 4 | 28 | 16 | 9 | 43 | ||
| 1981 | - | - | - | - | - | 0 | 51 | 47 | 2 | 0 | - | - | - | - | - | 14 | 60 | 0.3 | 0.1 | 26 | 11 | 58 | 10 | 0.4 | 20 | ||
| 1982 | 0.5 | 36 | 0.8 | 7 | 56 | - | - | - | - | - | 8 | 43 | 2 | 14 | 33 | - | - | - | - | - | 3 | 38 | 1 | 9 | 49 | ||
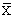 | 1972-77 | 10 | 12 | 7 | 4 | 68 | 6 | 6 | 6 | 8 | 74 | 14 | 6 | 19 | 5 | 56 | 7 | 14 | 16 | 7 | 56 | 11 | 10 | 14 | 5 | 60 | |
| 1978-82 | 4 | 35 | 7 | 9 | 45 | 10 | 33 | 28 | 14 | 15 | 6 | 19 | 9 | 14 | 52 | 12 | 42 | 6 | 9 | 31 | 7 | 31 | 9 | 11 | 41 | |
| Pimentel-Huarmey: | 1972 | - | - | - | - | - | - | - | - | - | - | 17 | 1 | 5 | 4 | 73 | 3 | 0 | 0 | 0 | 97 | 16 | 1 | 5 | 4 | 74 | |
| 7-10°S | 1973 | 0.6 | 0.1 | 0.2 | 2 | 97 | - | - | - | - | - | 70 | 8 | 0.5 | 0.5 | 21 | 64 | 4 | 5 | 4 | 23 | 37 | 4 | 1 | 2 | 56 | |
| 1974 | 50 | 7 | 8 | 8 | 27 | 15 | 22 | 13 | 10 | 40 | 11 | 10 | 24 | 9 | 46 | 41 | 0 | 25 | 2 | 32 | 22 | 10 | 17 | 7 | 39 | ||
| 1975 | 41 | 0.6 | 3 | 6 | 49 | - | - | - | - | - | 43 | 0.4 | 4 | 10 | 42 | - | - | - | - | - | 42 | 0.5 | 3 | 7 | 47 | ||
| 1976 | 47 | 1 | 4 | 1 | 47 | - | - | - | - | - | 48 | 3 | 2 | 0.5 | 46 | 8 | 25 | 9 | 2 | 56 | 39 | 6 | 5 | 1 | 49 | ||
| 1977 | 10 | 23 | 20 | 23 | 24 | - | - | - | - | - | 0 | 67 | 9 | 2 | 22 | 8 | 17 | 16 | 12 | 52 | 8 | 21 | 17 | 15 | 39 | ||
| 1978 | - | - | - | - | - | - | - | - | - | - | 36 | 13 | 23 | 16 | 12 | 69 | 8 | 3 | 8 | 12 | 62 | 9 | 7 | 10 | 12 | ||
| 1979 | 63 | 11 | 0.2 | 3 | 23 | - | - | - | - | - | 4 | 13 | 18 | 30 | 35 | - | - | - | - | - | 29 | 12 | 10 | 18 | 31 | ||
| 1980 | 4 | 64 | 5 | 13 | 14 | 21 | 41 | 5 | 8 | 25 | 1 | 36 | 10 | 14 | 39 | 17 | 38 | 14 | 10 | 21 | 10 | 45 | 10 | 11 | 24 | ||
| 1981 | - | - | - | - | - | 6 | 61 | 21 | 3 | 9 | - | - | - | - | - | 41 | 45 | 0.1 | 5 | 9 | 28 | 51 | 8 | 4 | 9 | ||
| 1982 | 16 | 24 | 7 | 7 | 46 | - | - | - | - | - | 11 | 34 | 16 | 4 | 35 | 3 | 29 | 14 | 16 | 38 | 12 | 30 | 13 | 7 | 38 | ||
| 1972-77 | 25 | 4 | 5 | 6 | 60 | 15 | 22 | 13 | 10 | 40 | 35 | 7 | 7 | 4 | 47 | 29 | 9 | 13 | 6 | 57 | 29 | 7 | 8 | 5 | 51 | |
| 1978-82 | 30 | 35 | 4 | 8 | 23 | 12 | 54 | 15 | 5 | 14 | 9 | 23 | 16 | 18 | 34 | 40 | 28 | 7 | 9 | 16 | 26 | 30 | 10 | 11 | 23 | |
| Huarniy-Piscos: | 1972 | - | - | - | - | - | - | - | - | - | - | 24 | 8 | 13 | 11 | 44 | 9 | 0 | 11 | 18 | 62 | 21 | 7 | 10 | 12 | 47 | |
| 10-14°S | 1973 | 46 | 2 | 0.03 | 5 | 53 | - | - | - | - | - | 77 | 6 | 2 | 0.7 | 14 | 51 | 12 | 0 | 4 | 31 | 61 | 5 | 1 | 1 | 30 | |
| 1974 | 45 | 6 | 12 | 6 | 31 | 38 | 24 | 18 | 8 | 12 | 65 | 8 | 6 | 3 | 18 | 66 | 0 | 3 | 2 | 29 | 53 | 11 | 10 | 5 | 21 | ||
| 1975 | 37 | 3 | 14 | 4 | 42 | - | - | - | - | - | 21 | 4 | 4 | 3 | 68 | - | - | - | - | - | 32 | 3 | 11 | 4 | 50 | ||
| 1976 | 35 | 2 | 10 | 5 | 48 | - | - | - | - | - | 44 | 24 | 7 | 1 | 24 | 39 | 27 | 10 | 3 | 21 | 37 | 11 | 9 | 5 | 38 | ||
| 1977 | 26 | 14 | 36 | 9 | 15 | - | - | - | - | - | 37 | 11 | 9 | 1 | 42 | 33 | 10 | 7 | 4 | 46 | 30 | 12 | 20 | 6 | 32 | ||
| 1978 | - | - | - | - | - | - | - | - | - | - | 20 | 9 | 48 | 5 | 18 | 39 | 39 | 12 | 2 | 8 | 34 | 31 | 21 | 3 | 11 | ||
| 1979 | 27 | 28 | 18 | 5 | 22 | - | - | - | - | - | 28 | 26 | 12 | 8 | 26 | - | - | - | - | - | 28 | 27 | 15 | 6 | 24 | ||
| 1980 | 19 | 29 | 16 | 5 | 31 | 2 | 21 | 18 | 3 | 56 | 43 | 16 | 20 | 2 | 19 | 22 | 18 | 21 | 13 | 26 | 23 | 21 | 19 | 7 | 30 | ||
| 1981 | - | - | - | - | - | 25 | 37 | 11 | 8 | 19 | - | - | - | - | - | 43 | 3 | 19 | 3 | 32 | 36 | 17 | 16 | 5 | 26 | ||
| 1982 | 21 | 18 | 3 | 2 | 56 | - | - | - | - | - | 23 | 21 | 15 | 4 | 37 | 0 | 27 | 23 | 8 | 42 | 18 | 21 | 11 | 4 | 46 | ||
| 1972-77 | 38 | 5 | 12 | 6 | 39 | 38 | 24 | 18 | 8 | 12 | 50 | 8 | 6 | 4 | 32 | 42 | 10 | 5 | 5 | 38 | 43 | 8 | 9 | 5 | 35 | |
| 1978-82 | 23 | 26 | 14 | 4 | 33 | 11 | 27 | 15 | 5 | 42 | 30 | 19 | 21 | 5 | 25 | 30 | 26 | 17 | 7 | 20 | 27 | 24 | 17 | 5 | 28 | |
| Piece-Frontara Sur: | 1972 | - | - | - | - | - | - | - | - | - | - | 14 | 2 | 21 | 1 | 62 | 50 | 0.1 | 17 | 0 | 33 | 24 | 1 | 20 | 1 | 54 | |
| 14-19'S | 1973 | 20 | 2 | 0.3 | 0 | 75 | - | - | - | - | - | 42 | 4 | 19 | 0.06 | 65 | 52 | 2 | 6 | 0 | 40 | 35 | 3 | 8 | 1 | 53 | |
| 1974 | 48 | 4 | 3 | 1 | 44 | 57 | 4 | 4 | 8 | 27 | 37 | 7 | 10 | 1 | 45 | 29 | 0 | 2 | 5 | 64 | 45 | 5 | 6 | 4 | 40 | ||
| 1975 | 26 | 3 | 23 | 8 | 40 | - | - | - | - | - | 20 | 3 | 14 | 0.7 | 62 | - | - | - | - | - | 24 | 3 | 20 | 5 | 48 | ||
| 1976 | 34 | 1 | 26 | 12 | 27 | - | - | - | - | - | 52 | 1 | 10 | 2 | 35 | 64 | 0 | 7 | 1 | 28 | 44 | 1 | 19 | 8 | 28 | ||
| 1977 | 16 | 7 | 27 | 6 | 44 | - | - | - | - | - | 16 | 8 | 32 | 1 | 43 | 16 | 10 | 21 | 1 | 52 | 16 | 8 | 26 | 4 | 46 | ||
| 1978 | - | - | - | - | - | - | - | - | - | - | 29 | 12 | 30 | 3 | 26 | 23 | 22 | 9 | 4 | 42 | 25 | 18 | 16 | 4 | 37 | ||
| 1979 | 10 | 15 | 26 | 6 | 43 | - | - | - | - | - | 14 | 21 | 30 | 8 | 27 | - | - | - | - | - | 12 | 18 | 27 | 7 | 36 | ||
| 1980 | 9 | 13 | 26 | 13 | 39 | 22 | 18 | 24 | 6 | 30 | 23 | 15 | 29 | 5 | 28 | 26 | 27 | 22 | 4 | 21 | 21 | 20 | 25 | 7 | 27 | ||
| 1981 | - | - | - | - | - | 17 | 13 | 18 | 8 | 44 | - | - | - | - | - | 21 | 22 | 14 | 1 | 42 | 19 | 18 | 16 | 4 | 43 | ||
| 1982 | 3 | 4 | 7 | 6 | 80 | - | - | - | - | - | 4 | 5 | 8 | 1 | 82 | 16 | 21 | 24 | 3 | 36 | 6 | 8 | 11 | 4 | 71 | ||
| 1983 | 4 | 33 | 24 | 6 | 33 | ||||||||||||||||||||||
| 1972-77 | 26 | 3 | 16 | 6 | 49 | 57 | 4 | 4 | 8 | 27 | 26 | 4 | 18 | 1 | 51 | 43 | 3 | 12 | 1 | 41 | 31 | 3 | 15 | 3 | 48 | |
| 1978-82 | 8 | 12 | 21 | 8 | 51 | 20 | 16 | 22 | 7 | 35 | 16 | 15 | 26 | 5 | 38 | 23 | 24 | 18 | 3 | 32 | 16 | 17 | 22 | 6 | 39 | |
| Toda la coastal: | 1972 | - | - | - | - | - | - | - | - | - | - | 16 | 3 | 16 | 5 | 60 | 31 | 0.1 | 12 | 4 | 52.9 | 19 | 2.5 | 15 | 5 | 58 | |
| 1973 | 22 | 1 | 0.2 | 3 | 76.8 | - | - | - | - | - | 59 | 5 | 6 | 0.42 | 29.6 | 50 | 6 | 6 | 2 | 36 | 42 | 4 | 4 | 2 | 48 | ||
| 1974 | 46 | 5 | 9 | 5 | 35 | 38 | 13 | 10 | 8 | 31 | 36 | 8 | 15 | 5 | 36 | 38 | 7 | 12 | 6 | 37 | 39 | 9 | 11 | 6 | 35 | ||
| 1975 | 32 | 3 | 16 | 7 | 42 | - | - | - | - | - | 23 | 4 | 11 | 4 | 58 | - | - | - | - | - | 28 | 3 | 14 | 6 | 49 | ||
| 1976 | 36 | 3 | 11 | 6 | 44 | - | - | - | - | - | 46 | 12 | 7 | 2 | 33 | 32 | 19 | 10 | 3 | 36 | 37 | 8 | 10 | 5 | 40 | ||
| 1977 | 18 | 15 | 28 | 10 | 29 | - | - | - | - | - | 21 | 21 | 17 | 1 | 40 | 19 | 10 | 14 | 6 | 51 | 18.5 | 14 | 21 | 7 | 39 | ||
| 1978 | - | - | - | - | - | - | - | - | - | - | 25 | 15 | 32 | 8 | 20 | 40 | 26 | 8 | 7 | 19 | 36 | 23 | 14 | 7 | 20 | ||
| 1979 | 21 | 20 | 18 | 6 | 35 | - | - | - | - | - | 14 | 18 | 19 | 15 | 34 | - | - | - | - | - | 17.6 | 19 | 18 | 10 | 35 | ||
| 1980 | 10 | 33 | 17 | 10 | 30 | 17 | 22 | 20 | 7 | 34 | 20 | 20 | 21 | 6 | 33 | 22 | 28 | 19 | 8 | 23 | 17.5 | 26.5 | 19 | 8 | 29 | ||
| 1981 | - | - | - | - | - | 14 | 35 | 20 | 6 | 25 | - | - | - | - | - | 30 | 33 | 8 | 2 | 27 | 24 | 33.5 | 13 | 14 | 25 | ||
| 1982 | 9 | 15 | 5 | 5 | 66 | - | - | - | - | - | 11 | 21 | 11 | 4 | 53 | 10 | 24 | 22 | 7 | 37 | 10 | 19 | 10 | 5 | 56 | ||
| 1972-77 | 28 | 5 | 11 | 6 | 50 | 38 | 13 | 10 | 8 | 31 | 34 | 6 | 12 | 3 | 45 | 35 | 8 | 11 | 4 | 42 | 32 | 6 | 11 | 5 | 46 | |
| 1978-82 | 15 | 23 | 14 | 7 | 41 | 16 | 27 | 20 | 7 | 30 | 16 | 18 | 20 | 10 | 36 | 28 | 28 | 14 | 6 | 24 | 20 | 23 | 16 | 8 | 33 | |
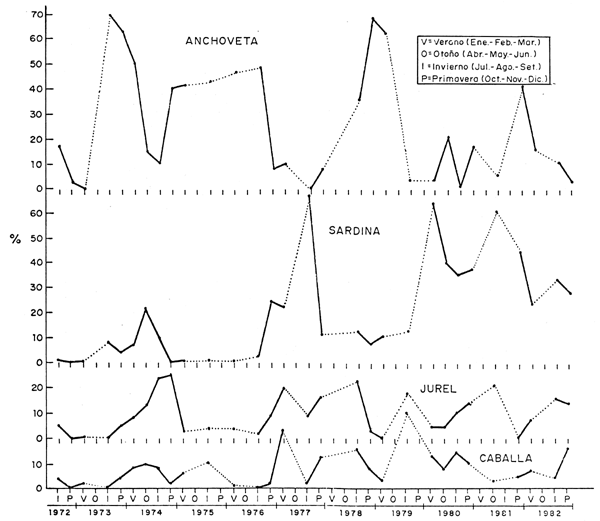
Fig. 37 Composición. por especies pelágicas (%) por estaciones y años en la zona compredido entre 7° y 10°S: Pimentel a Huarmey. (Datos de calas de cruceros acústico-pesqueros y eurekos.) Pelagic species composition (%) by seasons and years at 7° - 10° S, Pimentel-Huarmey. (Data from hauls made during Eurekas and Cruises).
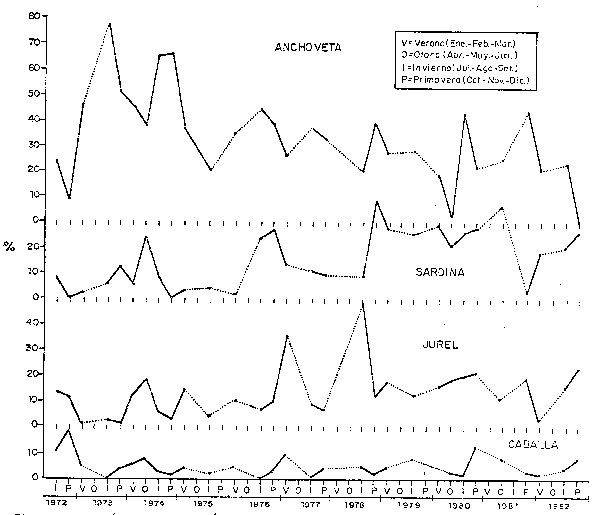
Fig. 38 Compositiógicas (%) por estaciones y anos, en la zona comprendida entre 10°y 14°S: Huarmey a Pisco. (Datos de calas de cruceros acóstico-pesqueros y Eurekas.) Pelagic species composition (%) by seasons and years at 10°-14°S: Huarmey-Pisco (Data from hauls made during Eureka and Cruises.)
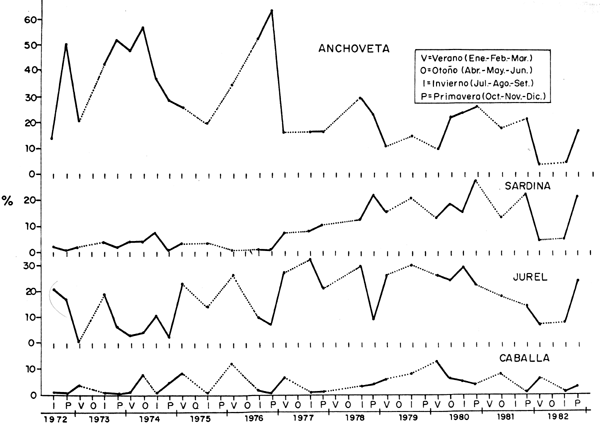
Fig. 39 Composición por espies pelagicas (%) por estaciones y anos, en la zona compredida entre 14°y 19°S: Pisco a la Frontero Sur. (Datos de calas de cruceros acustico-pesqueros y eurekas.) Pelagic species composition (%) by seasons and years at 14°-19°S: Pisco-Frontera Sur (Data from hauls made during Eurekas and Cruises.)
| ANOS | BIOMASA PROMEDIO | DESEMBARQUES | TASA DE EXPLOTACION |
|---|---|---|---|
| Years | Mean biomass | Landings | Exploitation rate |
| 106TM | 103 TM | C/ = F/F+M (1-exp-z) = F/F+M (1-exp-z) | |
| 1963 | 13.4 | 6423 | 0.48 |
| 1964 | 14.7 | 8863 | 0.60 |
| 1965 | 18.6 | 7233 | 0.39 |
| 1966 | 18.5 | 8529 | 0.46 |
| 1967 | 22.0 | 9824 | 0.45 |
| 1968 | 16.8 | 10262 | 0.61 |
| 1969 | 14.2 | 8960 | 0.63 |
| 1970 | 16.0 | 12277 | 0.77 |
| 1971 | 13.4 | 10282 | 0.77 |
| 1972 | 6.6 | 4448 | 0.67 |
| 1973 | 4.9 | 1848 | 0.38 |
| 1974 | 5.4 | 3583 | 0.66 |
| 1975 | 6.8 | 3071 | 0.45 |
| 1976 | 8.3 | 3870 | 0.47 |
| 1977 | 3.0 | 700 | 0.23 |
| 1978 | 3.9 | 1207 | 0.31 |
| 1979 | 3.4 | 1294 | 0.38 |
| 1980 | 1.8 | 566 | 0.31 |
| 1981 | 3.0 | 1167 | 0.39 |
| 1982 | - | 1700(**) | 0.6 - 0.8 ? |
(*) Tomado de Tsukayama, 1983 (en prensa)
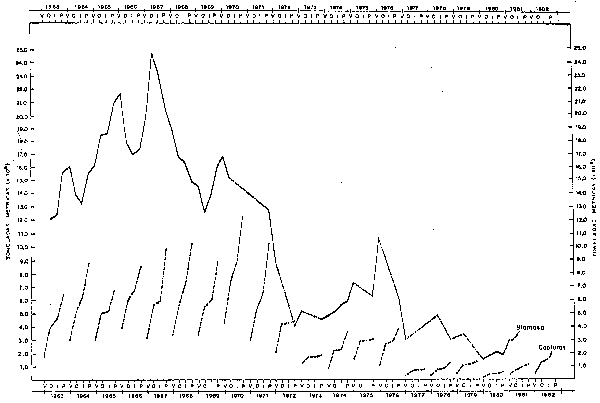
Fig. 40 Fluctuaclones estaclonales e interanuales de la biomasa de Achoveta en la costa peruana y capturas anuoles (acumulados por estaciones) durante 1963-82. Seasonal and annual variations in Anchovelta biomass along peruvian coast and annual catches (accumulated by seasons) during 1963-82. (De Tsukayama 1983, En prensa)
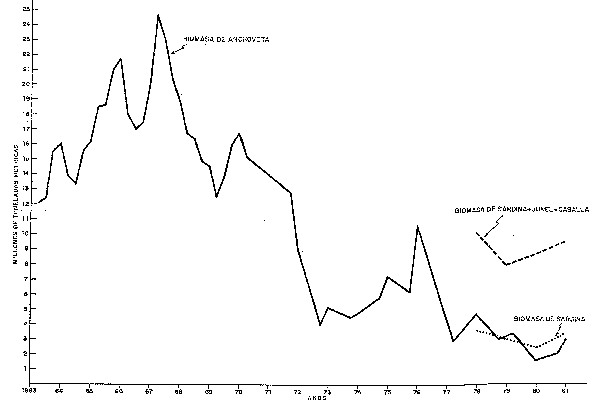
Fig. 41 FLUCTUACION DE LA BIOMASA DE "ANCHOVETA" EN EL PERIODO 1963-1981 Y DE "SARDINA" "JUREL" Y "CABALLA" EN EL PERIODO 1978-1981. ANCHOVETA BIOMASS FLUCTUATION DURING 1963-81 AND SARDINA, JUREL, CABALLA DURING 1978-81.
En estas circunstancias, ocurre El Niño 1972 que precipitó la caída del stock al incidir fundamentalmente en una pobre sobrevivencia de los primeros estados de vida de esta especie, generando a su turno el reclutamiento más pobre registrado hasta esa fecha, que representó 1/7 del reclutamiento promedio de los años 1961-71. Durante 1961-71 en que las condiciones ambientales fueron más estables y la biomasa de adultos fue alta, el reclutamiento varió por un factor de tres veces.
La situación de inestabilidad se acentuó en los años siguientes, en que la población ha continuado sometida a intensa explotación, disminuyendo de esta manera las posibilidades de restablecerse y haciéndola a su vez cada vez más sensible a los cambios aún sutiles del ambiente, con la consecuencia de generar reclutamientos muy pobres que han variado por un factor de 18 veces en el período 1972-78. Esta situación de stress se ha incrementado con la creciente expansión de otras poblaciones pelágicas las cuales exhiben un mayor rango de tolerancia frente a las variaciones del ambiente.
En el período 1978-81 la tasa de explotación es de 0.37. que se consideraba elevada para el nivel poblacional de esos años, cuya biomasa se situó en las 2-3 millones de toneladas, lo cual ha mermado aún más el stock desovante. En efecto, el stock desovante en 1981 ha sido muy reducido, entre 0.6 a 2.0 millones de toneladas, en las regiones norte y central (J. Alheit, comunicación personal).
En 1982, con la ausencia casi total de reclutas provenientes de los desoves de 1981 y capturas de alrededor de 1.7 millones de toneladas, la tasa de explotación debe haber superado la cifra de 0.60. La ocurrencia del Niño 1982-83 en circunstancias aún más críticas que los de 1972-73 y 1976-77, inducen a pensar que las posibilidades de recuperación del stock, tomará un período largo, aún prohibiendo totalmente su explotación.
Con respecto a las otras especies pelágicas: Los estimados de biomasa de la población de sardina, jurel y caballa se han efectuado a partir de 1978, indicando valores de aproximadamente 3.0 millones de toneladas de sardina, 4.5 de jurel y 1.5 millones de toneladas de caballa; haciendo un total de alrededor de 9 millones de toneladas para las tres especies en conjunto (Fig. 41).
Aún cuando no se disponen de estimaciones para años anteriores, las evidencias mostradas con respecto al aumento en su área de distribución y en la composición por especies, permiten inferir que el aumento de la sardina se ha producido a partir de 1972, con la declinación del stock de anchoveta.
Asimismo, las informaciones de desove dadas por Santander (1981) indican que en los años 1966-68 el desove de sardina fue muy pobre y distribuido en el área de Pta. Aguja-Huarmey (6-10°S); a partir de 1969 se percibe un cierto incremento en el número de huevos y a partir de 1972 se hace más abundante, al mismo tiempo que se amplia su área de desove.
Estos desoves incrementados en 1971-72, aunados a un ambiente propicio, se vieron reflejados en las capturas de sardina mezcladas con la anchoveta, realizados por la flota anchovetera en 1973. Las capturas de sardina que en los años anteriores no sobrepasó las 10 000 toneladas, alcanza en 1973 la cifra de 132 000 toneladas constituidas predominantemente por ejemplares de 12-18 cm y edades aproximadas de 1-2 años (Tabla 4).
| AÑO | ANCHOVETA | SARDINA | JUREL | CABALLA |
|---|---|---|---|---|
| 1951 | 12 | 1 | - | 1 |
| 52 | 16 | 0.4 | - | 3 |
| 53 | 37 | 0.1 | - | 2 |
| 54 | 43 | - | - | 4 |
| 55 | 59 | - | - | 1 |
| 56 | 119 | - | - | 4 |
| 57 | 326 | - | - | 9 |
| 58 | 737 | 2 | 0.2 | 13 |
| 59 | 1909 | 4 | 0.4 | 9 |
| 1960 | 2944 | 3 | 0.3 | 9 |
| 61 | 4579 | 3 | 0.2 | 12 |
| 62 | 6275 | 3 | 0.7 | 13 |
| 63 | 6423 | 2 | 2 | 8 |
| 64 | 8863 | 10 | 2 | 2 |
| 65 | 7233 | 7 | 3 | 4 |
| 66 | 8529 | 2 | 4 | 8 |
| 67 | 9824 | 2 | 3 | 13 |
| 68 | 10262 | 2 | 3 | 7 |
| 69 | 8960 | 1 | 4 | 7 |
| 1970 | 12277 | 0.4 | 5 | 9 |
| 71 | 10282 | 6 | 9 | 10 |
| 72 | 4448 | 6 | 19 | 9 |
| 73 | 1848 | 132 | 43 | 65 |
| 74 | 3583 | 73 | 129 | 63 |
| 75 | 3071 | 63 | 37 | 24 |
| 76 | 3870 | 175 | 54 | 40 |
| 77 | 700 | 871 | 505 | 46 |
| 78 | 1207 | 1244 | 387 | 112 |
| 79 | 1294 | 1675 | 147 | 108 |
| 1980 | 566 | 1480 | 124 | 59 |
| 81 | 1167 | 1163 | 68 | 42 |
| 82 | 1707 | 1496 | 72 | 19 |
Fuente de información: Instituto del Mar del Perú.
Ministerio de Pesquería.
| PAISES | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 |
|---|---|---|---|---|---|---|
| Chile | 164.9 | 327.2 | 620.8 | 732.8 | 1618.9 | 1772.7 |
| Peru | 62.9 | 174.7 | 870.9 | 1257.9 | 1/27.2 | 1480.4 |
| Rusia | - | - | - | ø | ø | 0.2 |
| TOTAL | 227.8 | 501.9 | 1491.7 | 1990.8 | 3346.2 | 3253.3 |
JUREL (Trachurus symmetricus murphy)
| PAISES | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 |
|---|---|---|---|---|---|---|
| Bulgaria | - | - | - | - | 4.7 | 15.1 |
| Chile | 261.2 | 342.3 | 340.8 | 586.7 | 597.5 | 562.3 |
| Cuba | - | - | - | - | - | - |
| Japón | - | ø | 2.3 | 1.7 | 0.1 | - |
| Korea | - | - | - | 0.8 | - | - |
| Perú | 37.9 | 54.2 | 505.0 | 462.5 | 150.9 | 123.2 |
| Polonia | - | - | - | - | 1.2 | 0.5 |
| URSS | - | - | - | 49.2 | 532.2 | 494.4 |
| TOTAL | 299.1 | 396.5 | 848.1 | 1100.9 | 1286.6 | 1195.5 |
CABALLA (Scomber japonicus)
| PAISES | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 |
|---|---|---|---|---|---|---|
| Bulgaria | - | - | - | - | 0 | 0.1 |
| Chile | 15.2 | 52.7 | 141.2 | 182.7 | 89.1 | 102.3 |
| Cuba | - | - | - | - | 0.1 | 2.8 |
| Japón | - | - | - | * | * | - |
| Korea | - | - | - | - | - | - |
| Perú | 23.6 | 40.2 | 46.1 | 97.2 | 118.0 | 59.0 |
| Polonia | - | - | - | - | - | - |
| URSS | - | - | - | 1.8 | 5.8 | 41.9 |
| TOTAL | 38.8 | 92.9 | 187.3 | 281.7 | 213.0 | 206.1 |
Fuente de información: Anuario Estadístico de FAO.
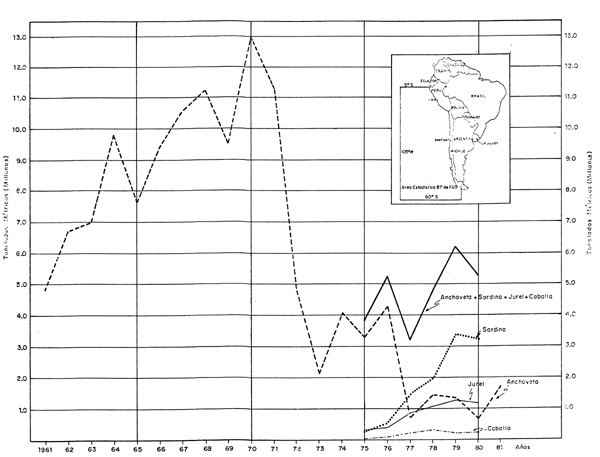
Fig. 42 Captura de Anchoveta, Sardina, jurel y Caballa en el área 87 de FAO. Catch of Anchoveta, Sardina, jurel and Caballa in the FAO Statistical Area 87.
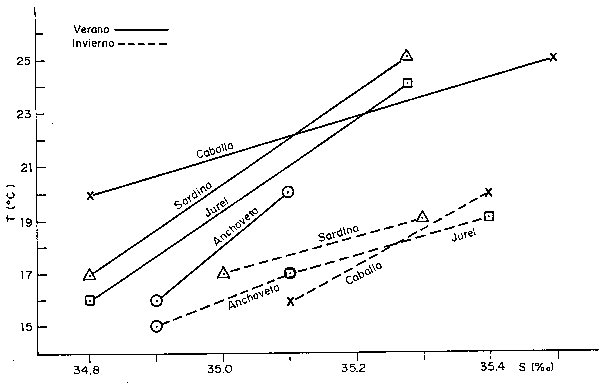
Fig. 43 Diagrama T-S y rango preferencial tentativo de la Anchoveta (A), Sardina (S), Jurel (J) y Caballa (C).
El número de huevos de sardina graficados por Santander (1981) muestra un mayor incremento en los años 1976-80, lo que aunado a las evidencias mostradas en las secciones anteriores, con respecto a la ampliación de las áreas de distribución y el incremento en la proporción de esta especie, independiente a los datos de desembarque que también se han incrementado sustancialmente, sustentan el hecho que se ha producido un real incremento de la población de sardina.
Para el jurel y la caballa, las evidencias mostradas en las secciones anteriores también indican que se ha producido un sustancial incremento poblacional, no obstante los desembarques de estas especies en la costa peruana son reducidas.
Merece destacarse la tendencia creciente en las capturas de sardina, jurel y caballa en la costa peruana y principalmente en el área 87 de FAO, simultáneamente con la declinación del stock de anchoveta (Figs. 1 y 42, y Tablas 4 y 5). Las fluctuaciones en las poblaciones de sardina, jurel y caballa, dada su amplia distribución, deben observarse a través de todo el Pacífico Oriental usando en lo posible datos independientes de la pesquería. A este respecto, cabe destacar la ejecución del Proyecto Regional "Evaluación de los Recursos Sardina, Jurel y Caballa en el Pacífico Suroriental" con la participación de Ecuador, Perú y Chile, bajo la coordinación del SELA y el apoyo económico del Banco Interamericano de Desarrollo.
DISCUSION
Uno de los puntos que merece destacarse en el patrón de distribución de las especies estudiadas, de condiciones ambientales diferenciables dentro de un espectro más o menos amplio de las variables ambientales, es la evidente yuxtaposición o traslape. Este hecho se considera de particular importancia en la etapa actual que podríamos denominar de transición, ya que aquella especie que posea e imponga un mayor grado de adaptación o tolerancia a las fluctuaciones ambientales y detente una mayor dispersión que le permita comparativamente tener una menor vulnerabilidad a la pesca, ocupará en el mediano o largo plazo, un lugar preponderante en el ecosistema pelágico, aun cuando no alcance el mismo nivel poblacional o características de la anchoveta de distinto nivel trófico en la cadena alimenticia del mar.
La intensidad y el tipo de interacción generados por la coincidente distribución observada, evidentemente serán elementos definitorios en la estabilidad a largo plazo de determinadas especies en la comunidad pelágica del Pacífico Sur-oriental.
O'Brien (1972) al hacer un análisis estadístico de los vientos del Pacífico, observa un cambio en el patrón de distribución de estos vientos en la década del setenta, en la que se han presentado además dos eventos El Niño (1972 y 1976). Esto podría estar indicando cambios de macro-escala en el sistema océano-atmósfera que podrían haber conducido al evento 1982-83 de características singulares pero también diferentes en muchos aspectos a fenómenos previos y que tuvo como precedente el período 1977-81 de tendencias promedio.
Es probable que estos cambios hayan posibilitado algunas variaciones en los ecosistemas marinos, particularmente en los de régimen costero, lo que a su vez podría explicar en cierta medida la influencia del ambiente en la magnitud y velocidad del incremento de las poblaciones de sardina, jurel y caballa, especialmente estas dos últimas, que no son propiamente de régimen de afloramiento costero como la anchoveta.
A escala local, las fronteras hídricas, los remolinos de dimensiones variables y el flujo de la corriente costera peruana en forma de meandros, establecen un panorama complejo en la distribución de los recursos, reflejado en la conformación de núcleos de concentración de cardúmenes que propician la mayor interacción de especies.
Por otro lado, los períodos cálidos (El Niño) parecen favorecer la competencia de especies en detrimento de los recursos de régimen netamente costero como la anchoveta, y contrariamente, los períodos fríos (anti-Niño) favorecen una expansión de los recursos costeros y la disminución de la disponibilidad para la pesca.
Este hecho nos permite llamar la atención sobre la necesidad de revisar la estrategia de monitoreo dando énfasis a los estudios en series de tiempo, así como a las observaciones simultáneas de ambiente y recurso, dentro de un plan sustentado en hipótesis de trabajo como la que estamos presentando en la Fig. 43, en la que combinamos conocimientos teóricos con observaciones preliminares para graficar curvas del rango ambiental preferencial de las especies anchoveta, sardina, jurel y caballa, tanto para verano como para invierno.
Finalmente, las condiciones descritas también incrementan la interacción a nivel de los primeros estadios de vida, ya sea a través del canibalismo o de la predacción de huevos y larvas, hecho que a su turno determinará en gran medida la fuerza del reclutamiento.
Siendo el reclutamiento el parámetro fundamental del cual depende la magnitud de las poblaciones y al mismo tiempo el que ha mostrado alta variabilidad como producto de las interacciones provocadas por el ambiente, se hace necesario monitorizar la abundancia de los pre-reclutas a fin de coadyuvar a una planificación adecuada de la administración de los recursos pelágicos.
AGRADECIMIENTOS
Expresamos nuestro reconocimiento a la Srta. María Elena Nakama, a los Sres. Manuel Flores F., Augusto Haro y David Camposano, por su eficente colaboración en el procesamien- to de los datos. Igualmente nuestro agradecimiento a la Sra. Isabel Delgado de Risso, Adalinda Rosales, por el mecanografiado de los manuscritos. De igual forma reconocemos la colaboración del Sr. Guillermo Gracey en la elaboracion de los gráficos.
REFERENCIAS
Bakun, A. y R.H. Parrish. 1981. Environmental inputs to fishery population models for eastern boundary currents. pp. 67-79. In Workshop on the effects of environmental variation on the survival of larval pelagic fishes (G.D. Sharp, ed.). IOC Workshop Report No.28. IOC/UNESCO, Paris.
Brink, K.H., B.H. Jones, J.C. Van Leer, C.N.K. Mooers, D.W. Stuart, M.R. Stevenson, R.C. Dugdale y G.W. Heburn. 1981. Physical and biological structure and variability in an upwelling center off Peru near 15oS during March 1977. pp. 473-495. In Coastal upwelling (F.A. Richards, ed.). Amer.Geophys.Union, Washington, D.C.
Brockmann, C., E. Fahrbach, A. Huyer and L. Smith. 1980. The poleward undercurrent along the Peru coast: 5o to 15oS. Deep-Sea Res. (27A):847-856.
Busalacchi, A.J., Jr. 1982. Linear, wind-driven variability of the Tropical Pacific and Atlantic Oceans. Technical Report, Dpt. of Oceanography, Florida State University.
Cochrane, J.D. y S. Zuta. 1968. Equatorial currents east of the Galapagos Islands in February-March 1967. Texas A & M University, Department of Oceanography, Ref. 67-5-T.
Codispoti, L.A. y T.T. Packard. 1980. Denitrification rates in the eastern tropical South Pacific. J.Mar.Res. 38(3):453-477.
Csirke, J. 1980. Recruitment in the Peruvian anchovy and its dependence on the adult population. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:307-313.
Chirichigno, N. 1974. Clave para identificar los peces marinos del Perú. Informe Inst.Mar Perú-Callao 44:1-387.
De Vries, T.J. y W.G. Pearcy. 1982. Fish debris in sediments of the upwelling zone off central Peru: a late quaternary record. Deep-Sea Res. 28(1A):87-109.
Enfield, D.B. 1980. El Niño Pacific eastern boundary response to interannual forcing. In Resource management and environmental uncertainty (M.H. Glantz and J.D. Thompson, eds). Wiley, New York.
Estudio Regional Fenómeno del Niño. 1983. (MS). Informe resumido del Programa Biológico del ERFEN. Tercera Reunión del Comité Cientifico del ERFEN. Cali-Colombia, 16-19 febrero 1983.
Flores, R. y W. Urquizo. (En prensa). Informe del Crucero 8108 del SNP-1, 6 ago-4 set 1981.
Guillén, O. y R.I. de Rondán. 1974. Productividad de las aguas costeras frente al Perú. I Parte: Medio ambiente y producción primaria. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-148.
Instituto del Mar del Perú. 1972a. Regulación de la pesquería de anchoveta para el año calendario 1972. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-97.
Instituto del Mar del Perú. 1972b. La anchoveta en relación con el fenómeno de "El Niño 1972". Serie de Informes Especiales Inst.Mar Perú-Callao. IM-104.
Instituto del Mar del Perú. 1973a. Operación Eureka XXVI, 20-23 enero 1973. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-122.
Instituto del Mar del Perú. 1973b. Operación Eureka XXVII, 23-26 setiembre 1973. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-141.
Instituto del Mar del Perú. 1973c. Operación Eureka XXVIII, 12-13 nov 1973 y la pesca de comprobación, 14-17 nov 1973. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-146.
Instituto del Mar del Perú. 1974a. Informe de la cuarta sesión del panel de expertos de la evaluación del stock de anchoveta peruana. Bol.Inst.Mar Perú-Callao. 2(10):605-671.
Instituto del Mar del Perú. 1974b. Informe del Cateo No4 (Prospección sinóptica tipo Eureka), 26-28 febrero 1974. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-156
Instituto del Mar del Perú. 1974c. Operación Eureka XXIX, 28-30 mayo 1974. Serie de Informes Especiales Inst.Mar Peru-Callao. IM-158.
Instituto del Mar del Perú. 1974d. Operación Eureka XXX, 7-8 agosto 1974. Serie de Informes Especiales Inst.Mar Peru-Callao. IM-164.
Instituto del Mar del Perú. 1974e. Operación Eureka XXXI, 3-4 set 1974. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-168.
Instituto del Mar del Perú. 1976a. Operación "Búsqueda de Peces Reclutas", 14-15 nov 1974. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-171.
Instituto del Mar del Perú. 1976b. Operación Eureka XXXII, 14-15 febero 1975. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-177.
Instituto del Mar del Perú. 1976c. Operación Eureka XXXIII, 6-7 agosto 1975. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-178.
Instituto del Mar del Perú. 1977a. Operación Eureka XXXVI, 11-13 agosto 1976. Serie de Informes Especiales Inst.Mar Perú-Callao. IM-185.
Instituto del Mar del Perú. 1977b. El recurso anchoveta, el medio ambiente, la regulación de su pesquería desde 1960 a 1977 y sus perspectivas. Informe al Despacho Ministerial.
Instituto del Mar del Perú. 1981. Informe sobre la situación de los recursos pelágicos a principios de 1981 y las proyecciones de pesca. Informe al Despacho Ministerial.
Instituto del Mar del Perú. 1982. Informe preliminar de la Op. Eureka XLVII, del 27 febrero - 2 mar 1982. Informe al Despacho Ministerial.
Johannesson, K.A. y A. N.Robles. 1977. Echo surveys of Peruvian anchoveta. Rapp.P-v.Réun.Cons. int.Explor.Mer. 170:237-244.
Johannesson, K.A. y R. Vílchez. 1981. Note on hydroacoustic observations of changes in distribution and abundance of some common pelagic fish species in the coastal waters of Peru, with special emphasis on anchoveta. pp. 287-323. In Workshop on the effects of environmental variation on the survival of larval pelagic fishes (G.D. Sharp, ed.). IOC Workshop Report No.28. IOC/UNESCO, Paris.
Jordán, R., J. Csirke y I. Tsukayama. 1978. Situación de los recursos anchoveta, sardina, jurel y caballa a junio de 1978. Informe Inst.Mar Perú-Callao. No 56.
MacCall, A.D. 1981. The consequence of cannibalism in the stock-recruitment relationship of planktivorous pelagic fishes such as Engraulis. pp. 201-220. In Workshop on the effects of environmental variation on the survival of larval pelagic fishes (G.D. Sharp, ed.). IOC Workshop Report No.28. IOC/UNESCO, Paris.
Mendiola, B.R. de. 1978. El alimento de la anchoveta Engraulis ringens J. en una área de afloramiento (San Juan). Informe Inst.Mar Perú-Callao. 49:1-15.
Mugica, R. 1972. Oceanografía de mar peruano. Historia Marítima del Perú 1:216-474.
O'Brien, J.J. y H.E. Furlburt. 1972. A numerical model of coastal upwelling. J.Phys.Oceanog. 2:14- 26.
Paul, C.A., S.Zuta y D.V. Hansen. (En prensa). Drift buoy measurements in the eastern tropical Pacific during 1979-1980.
Quinn, W., T.V. Neal y S.K. Esbensen. 1982. Southern oscilation, El Niño and Indonesian droughts. Nature. 4 March 1982:296.
Rasmusson, E.M. y T.H. Carpenter. 1982. Variations in tropical sea surface temperature and surface wind fields associated with the southern oscilation/El Niño. Mon.Wea.Rev. 110(5):109-384.
Santander, H. 1981. Fluctuaciones del desove de anchoveta y algunos factores relacionados. pp. 255-274. In Workshop on the effects of environmental variation on the survival of larval pelagic fishes (G.D. Sharp, ed.). IOC Workshop Report No.28. IOC/UNESCO, Paris.
Santander, H. y O.S. de Castillo. 1977. Variaciones en la intensidad del desove de sardina, Sardinops sagax sagax J. en la costa peruana en los años 1966-1973. Bol.Inst.Mar Perú-Callao. 3(3):73-94.
Sharp, G.D. 1981. Report on the workshop on effects of environmental variation on survival of larval pelagic fishes. pp. 15-59. In Workshop on the effects of environmental variation on the survival of larval pelagic fishes (G.D. Sharp, ed.). IOC Workshop Report No.28. IOC/UNESCO, Paris.
Stommel, H. 1948. The westward intensification of wind-driven ocean currents. Trans.Amer. Geophys.Un. 29:202-206.
Tomczak, M., Jr. 1981. Coastal upwelling systems and Eastern Boundary Currents: a review of terminology. Geoforum. 12(2):179-191.
Tsukayama, I. 1983. (En prensa). Recursos pelágicos y sus pesquerías "Seminario Regional sobre Recursos Pelágicos", COCIC X, Comisión Permanente del Pacífico Sur (CPPS). Guayaquil- Ecuador, 25-29 octubre 1982.
Tsukayama, I. y M.A. Alvarez. 1981. Fluctuaciones en el stock de anchovetas desovantes durante los ciclos reproductivos de primavera 1964-1978. pp. 233-240. InWorkshop on the effects of environmental variation on the survival of larval pelagic fishes (G.D. Sharp, ed.). IOC Workshop Report No.28. IOC/UNESCO, Paris.
Ware, D. y I. Tsukayama. 1981. A possible recruitment model for the Peruvian anchovy. Bol.Inst. Mar Perú-Callao. Vol.Extraordinario:55-61.
Wyrtki, K. 1963. The horizontal and vertical field of motion in the Peru Current. Bull.Scripps Inst.Oceanogr.8(4):313-345.
Wyrtki, K. 1964. The thermal structure of the eastern Pacific Ocean. Deutsches Hydrogr.Inst., Erganzungsheft. A(6):84.
Wyrtki, K. 1965. Surface currents of the eastern tropical Pacific Ocean. Bull.Inter-Amer.Trop. Tuna Comm. 9(5):270-304.
Wyrtki, K. 1982. The southern oscilation, ocean-atmosphere interaction and El Niño. Mar. Tech.Soc.J. 16(1):3-10.
Zuta, S., D.Enfield, J. Valdivia, P. Lagos y C. Blandin. 1976. Aspectos físicos del fenómeno El Niño 1972-73. FAO, Informes de Pesca. (185):3-61.
Zuta, S. y O. Guillén. 1970. Oceanografía de las aguas costeras del Perú. Bol.Inst.Mar Perú-Callao. 2(5):157-324.
Zuta, S. y H. Santander. 1974. El ambiente oceánico y su relación con los organismos marinos. Primer Sem.Nac.de Sist.Ecolog.Recurs.Nat. y Medio Ambiente, junio 1974, Lima.
Zuta, S. y W. Urquizo. 1972. Temperatura promedio de la superficie del mar frente a la costa peruana, periodo 1928-69. Bol.Inst.Mar Peru-Callao. 2(8):462-519
![]()
![]()
![]()