
0296-B1
E.H. (Ted) Hogg[1], James P. Brandt and B. Kochtubajda
Global models predict that the earth’s climate will become warmer over the next century, as atmospheric concentrations of carbon dioxide (CO2) and other greenhouse gases continue to increase. The rate of warming is expected to be especially rapid in northern continental areas, where temperatures have already increased by up to 2°C over the past century. There is conflicting evidence, however, as how northern forests may be affected by these changes. Future forest growth may be stimulated by rising CO2 levels and longer growing seasons, but there is also a serious risk of negative impacts, notably increases in drought, fire, and damage by insects and diseases. In the western Canadian interior, forests dominated by aspen (Populus tremuloides Michx.) are especially vulnerable to future climate change, because they occur in a climatically dry zone along the northern edge of the Great Plains. Dieback and reduced growth of aspen forests has been noted in some areas, especially following periods of drought and large-scale insect outbreaks. This poses concerns for the future of these aspen forests under climate change projections for a warmer and drier climate. To address these concerns, we established a regional study (CIPHA) that includes annual monitoring of 72 aspen stands in climatically-sensitive areas across western Canada. Tree-ring analysis of these stands has shown that during 1950-2000, regional aspen growth has undergone several cycles of collapse and recovery. These cycles were mainly driven by drought and insect defoliation, which caused reduced growth. Since 1998, the region has been affected by one of the worst droughts on record. To date (2002), relatively modest levels of aspen dieback have been recorded in these stands, but continued monitoring should provide an early indication of any regional-scale aspen decline that may occur in the near future.
Recent global model simulations suggest that the earth=s climate will warm by an average of 1.4° to 5.8°C during the 21st century, as a result of human-induced increases in carbon dioxide (CO2) and other greenhouse gases (IPCC 2001a). The projected rate of warming is expected to be greatest in northern continental regions, which include the boreal forests of North America and northern Asia. Recent analyses of the climate record indicate that significant warming has already occurred in some areas, notably in the western Canadian boreal forest, where temperatures increased by 1.7° - 2.0°C between 1948 and 2001 (Environment Canada 2002).
From a climate change perspective, much of the recent international focus on forests has been on their role in taking up carbon from the atmosphere (e.g., Kurz and Apps 1999, IPCC 2000). However, the question of climate change impacts on the world=s forests and their management is also emerging as a critical global issue for the 21st century (IPCC2001b).
One of the fundamental questions for future forest management and conservation is how tree growth over large areas is likely to be affected by the anticipated global warming. Despite technological advances in simulation modelling and remote sensing, great uncertainty remains even in predicting the direction of change in the potential productivity of the world=s northern forests. Part of this uncertainty arises from the difficulty in determining how the effects of positive and negative factors will balance in a given geographic area, as illustrated diagrammatically in Figure 1.
Figure 1. Diagram summarizing major factors likely to affect productivity, biomass and carbon uptake of northern forests under global climate change.

On the one hand, climatic warming may be expected to benefit the productivity of forests, especially in cold climatic regions where the length of the growing season is limited by low temperatures. Further benefits may be expected owing to the role of CO2 in the photosynthesis of trees and other plants: as CO2 levels increase, forest growth will tend to increase due to ACO2 fertilization@, if other factors are not limiting. It has also been suggested that large-scale forest productivity has been significantly enhanced by human-induced increases in nitrogen (N) deposition (Chen et al. 2000). The inclusion of these positive effects in global models has led to projections of increasing forest productivity and carbon sequestration over the next century, especially in northern forests (e.g., Cao and Woodward 1998; White et al. 1999). In a survey of 11 leading ecologists, most believed that standing biomass of northern forests is likely to increase under future climate change with a doubling of CO2 levels, although soil carbon was generally expected to decrease (Morgan et al. 2001). A recent global analysis of remote sensing measurements from 1981 to 1999 suggests that the recent warming trend has led to an overall Agreening@ of most northern areas, especially in Eurasia (Zhou et al. 2001).
On the other hand, however, there are a number of potentially negative factors, whose impacts are often difficult to predict. For example, recent results from an outdoor field experiment in the northern U.S. showed that the expected benefits of elevated CO2 on aspen growth were negated when ground-level ozone was raised to the levels occurring in more polluted areas (Isebrands et al. 2001). Under global climate change, future increases in the severity of extreme climatic events may be expected to impact forests in a wide variety of ways (IPCC 2001b). In some areas, the projected warming poses a risk for much drier conditions in northern continental regions, potentially leading to large-scale impacts on forests either directly (Hogg and Hurdle 1995), or in combination with drought-induced increases in fire (Flannigan et al. 1998). There is evidence that increased drought stress associated with recent climatic warming has already led to reduced tree growth in some high-latitude areas such as Alaska (Barber et al. 2000). Potentially, climate change may also lead to losses of boreal forest cover following disturbance, in areas that become too dry for regeneration to occur (Hogg and Schwarz 1997). Furthermore, climate change poses a risk of increasing forest damage from outbreaks of pests (Volney and Fleming 2000), including both native and exotic species (Krcmar-Nozic et al. 2000). The concerns for large-scale impacts also extend to the tropics, where large areas of rain forests have been recently devastated by the combined impacts of drought, selective logging, and fire (Siegert et al. 2001). In the northern hardwood forests of the eastern U.S. and Canada, thaw-freeze events in winter and early spring have also been implicated as a major cause of forest dieback episodes during the past century (Auclair et al. 1996), but such events are rarely considered in large-scale models.
In the western Canadian interior, forests dominated by trembling aspen (Populus tremuloides Michx) are expected to be especially vulnerable to the impacts of a warmer and drier future climate (Hogg and Hurdle 1995). Trembling aspen is the most abundant deciduous tree species in the North American boreal forest, where it has become an important resource for the forest industry since the 1980s (Peterson and Peterson 1992). Aspen is also the predominant tree in the drought-prone aspen parkland zone along the northern edge of the Canadian prairies, where it forms stunted patches of forest interspersed with cropland and grassland. Since the early 1990s, dieback and reduced growth of aspen forests has beennoted in several areas of Canada. Previous studies (e.g., Hogg and Schwarz 1999, Hogg et al. 2002) indicate that the aspen decline in these areas was caused by a combination of climatic factors (drought and early spring thaw-freeze events) and multiple-year defoliation by the forest tent caterpillar (Malacosoma disstria Hbn.). This has posed concerns about the current status of western Canadian aspen forests, and how the productivity and health of these forests might be affected in the near future under climate change.
To address these concerns, we established a large-scale study entitled AClimate Change Impacts on Productivity and Health of Aspen@ (CIPHA). CIPHA consists of a network of long-term research plots in 72 aspen stands across western Canada, where annual monitoring of forest health and dieback was initiated in the year 2000. One of the first steps in this study has been to conduct analyses of tree rings to examine past variation in aspen growth at the regional scale (1800 km x 500 km area) of this study. Specifically, we postulated that over the past 50 years, regional aspen growth has been more strongly affected by cycles of drought and insect defoliation, rather than by temperature variation.
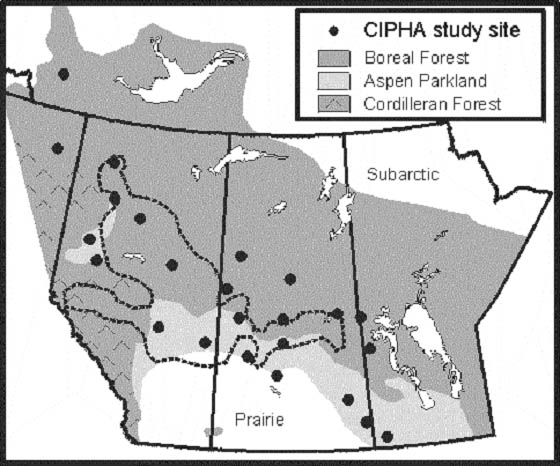
The study area includes 24 study sites in the southern boreal forest and adjacent aspen parklands of the western Canadian interior (Figure 2). At each site, three pure, undisturbed aspen stands (40-80 years old) were selected within a distance of 25 km (total of 72 stands). During the year 2000, two long-term research plots were established 50-100 m apart within each stand. The plots were rectangular, with a width of 10 m and sufficient length (typically 15-35 m) to include at least 25 living aspen trees. Heights and diameters (1.3-m height) of all aspen in the plots were measured and annual assessments of forest health were initiated. Factors including dieback and mortality, incidence of damage by insects, fungal pathogens, extreme weather were also monitored. Live basal area (m2 ha-1) of aspen in each plot was calculated from the tally of stem diameters and plot area. In early autumn 2000, 3 aspen trees located near each plot were felled for tree-ring analysis (total of 432 trees) and disks were collected from several heights; this analysis focuses on disks from the 1.3-m height. Disks were dried (50o C), sanded and polished, and ring widths along two radii per disk were measured manually under magnification. Ring widths were used to calculate tree growth as annual increment in stem cross-sectional area (Hogg et al. 2002). Annual stand growth (m2 ha-1 yr-1) was estimated by using stand basal area to Ascale up@ the tree-based growth increments. The history of severe insect defoliation was determined from the presence of abnormally pale-coloured Awhite@ tree-rings (Hogg and Schwarz 1999) and from insect survey records (see Simpson and Coy 1999). Past climate variation for the CIPHA study region was assessed from the historical records of temperature and precipitation at Environment Canada climate stations adjacent to each of the 24 study sites. Average total precipitation and temperature were calculated for the main period of aspen stem growth (1 May to 31 July) and for each 12-month period ending on 31 July. A preliminary analysis of factors affecting growth variation was conducted using multiple regression with precipitation, temperature and insect defoliation (percentage incidence of white rings) as independent variables, and differenced, log-transformed aspen growth as the dependent variable (Hogg et al. 2002).
Figure 2. Map showing location of CIPHA study areas in western Canada where aspen growth and health are being monitored. The area enclosed by the dashed line was affected by severe drought (lowest precipitation on record) during 1 September 2001 to 31 July 2002 (from Agriculture Canada and Agri-food Canada 2002).
The tree-ring analysis shows that aspen forests in western Canada have undergone several cycles of collapses in growth followed by recovery since 1950 (Figure 3). Preliminary regression analysis indicated that annual precipitation and insect defoliation were the most significant factors governing interannual growth variation, whereas the influence of temperature (either annual or growing season) was not significant. Aspen growth was greatly reduced during 1961-1964, 1979-1984, and 1988-1995, corresponding to periods with regional drought and large-scale outbreaks by forest tent caterpillar. The greatest oscillation in regional-scale growth included a 50% growth reduction between 1976 and 1981, followed by a doubling of growth between 1981 and 1986. The last peak in aspen growth was in 1997, following a cool, moist period with little defoliation. Regional aspen growth started to decline during 1998, which was the warmest year on record in the region (Environment Canada 2002), and had decreased by a total of 30% by 2000.
Figure 3. Year-to-year variation in average aspen growth and precipitation for the 24 CIPHA study areas of western Canada. Growth was determined from tree-ring analysis of 432 aspen stems, and is expressed as annual increment in stand basal area for trees that were living in the year 2000. Major years of drought and insect defoliation are also shown.
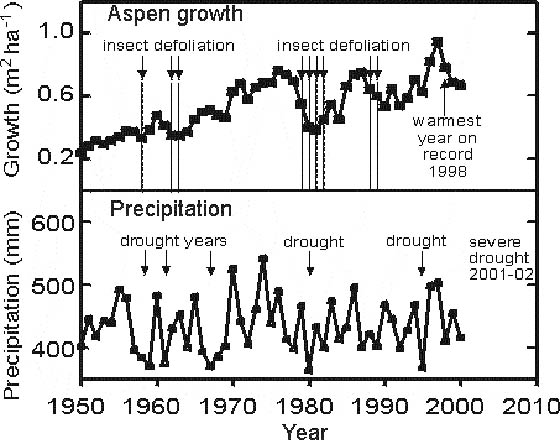
The results show that during the period 1950-2000, western Canadian aspen forests have undergone major oscillations in growth, even at the regional scale encompassed by this study. Drought and insect defoliation were the most important factors driving interannual variation in aspen growth, as noted in two previous tree-ring studies of aspen in western Saskatchewan (Hogg and Schwarz 1999) and northwestern Alberta (Hogg et al. 2002). Repeated defoliation by forest tent caterpillar has also been implicated as a factor causing recent dieback and mortality affecting more than 300,000 ha of aspen forest in the Kapuskasing area of northern Ontario. (G. Howse, Canadian Forest Service, pers. comm., 2002). However, several other factors may impact aspen growth and ultimately lead to crown dieback and decline of aspen forests in these regions. These include the influence of thaw-freeze events in late winter or spring, as well as damage by fungal pathogens and wood-boring insects, which tend to increase in aspen stands following drought or defoliation events (Hogg et al. 2002).
Following the period covered by this study, large portions of the CIPHA study region in western Canada experienced the worst drought on record in 2001-2002 (Figure 2), and in some of these areas, the drought has lasted over 5 years (1998-2002). Based on the results of our retrospective tree-ring analysis, it may be anticipated that this severe drought will lead to major reductions in aspen growth in this region. Stand growth may recover rapidly after the end of this drought, but there is also a risk of extensive aspen dieback in future, especially if there is a major outbreak of insect defoliators such as forest tent caterpillar. To date (2002), only modest levels of dieback have been noted within the CIPHA plots, which are situated in relatively large, undisturbed stands. However, the drought has led to extensive, severe dieback of aspen and other trees in municipalities, farm shelterbelts, and along the edges of forests in the affected area (E.H. Hogg, pers. obs., August 2002). Continued monitoring and a future update of tree-ring analysis within the CIPHA study will determine the longer-term impact of this drought on the aspen forests of this region. Future directions include the "scaling up" of tree-ring analyses for annual estimates of net primary production and the validation of models for projecting the potential impacts of climate change over the next few decades.
There is considerable uncertainty about how the health and productivity of northern forests is being affected by human-induced changes in the earth = s climate and atmosphere. Preliminary results from our regional study of aspen forests in western Canada indicate that drought and insect defoliation have been the most important factors governing year-to-year variation in growth over the past 50 years. Despite recent advances in remote sensing and modelling technologies, there is an ongoing need for large-scale, ground-based monitoring of forest productivity and health to enable Aearly warning@ detection of any changes that may be already occurring in response to global change. Such early detection could be critical for future decision-making in a wide variety of forest-related sectors, including the forest industry, management of wildlife, parks, and protected areas, and is also highly relevant to the current international focus on the potential role of forests in slowing the rate of global climate change through carbon sequestration.
This work was conducted through funding support from the Canadian Forest Service, the Government of Canada's Climate Change Action Fund, the Program of Energy Research and Development, the Meteorological Service of Canada, and Mistik Management Ltd. We thank R. Brett, M. Michaelian, and T. Hook for technical leadership and support, and J. Hammond, B. Tomm, J. Weber, A. Engel, R. Raypold, D. Sherling, B. Vroom, C. Charmin, P. Christensen, A. Durand, A. Gossell, A. Hamelink and T. Young for field and laboratory assistance.
Agriculture and Agri-food Canada, 2002. Drought watch, 31 July 2002, web site at http://www.agr.gc.ca/pfra/drought/index_e.htm
Auclair, A.N.D., J.T. Lill and C. Revenga, 1996. The role of climate variability and global warming in the dieback of northern hardwoods. Water, Air, and Soil Pollution 91 (3-4): 163-186
Barber,V.A., G.P. Juday and B.P. Finney, 2000. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 405 (6787): 668-673.
Cao, M. and F.I. Woodward, 1998. Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their responses to climate change. Global Change Biology 4(2): 185-198
Chen, W., J. Chen and J. Cihlar, 2000. An integrated terrestrial carbon-budget model based on changes in disturbance, climate, and atmospheric chemistry. Ecological Modelling 135: 55-79.
Environment Canada, 2002. Climate trends and variation bulletin for Canada, Annual 2001 temperature and precipitation in historical perspective, web site at http://www.msc-smc.ec.gc.ca/crm/bulletin/annual01/index.html
Flannigan, M.D., Y. Bergeron, O. Engelmark and B.M. Wotton, 1998. Future wildfire in circumboreal forests in relation to global warming. Journal of Vegetation Science 9: 469-476.
Hogg, E.H. and P.A. Hurdle, 1995. The aspen parkland in western Canada: A dry-climate analogue for the future boreal forest? Water, Air, and Soil Pollution 82: 391-400.
Hogg, E.H. and A.G. Schwarz, 1997. Regeneration of planted conifers across climatic moisture gradients on the Canadian prairies: implications for distribution and climate change. Journal of. Biogeography 24: 527-534.
Hogg, E.H. and A.G. Schwarz, 1999. Tree-ring analysis of declining aspen stands in west-central Saskatchewan. Canadian Forest Service, Northern Forestry Centre, Information Report NOR-X-359.
Hogg, E.H., J.P. Brandt and B. Kochtubajda, 2002. Growth and dieback of aspen forests in northwestern Alberta, Canada, in relation to climate and insects. Canadian Journal of Forest Research. 32: 823-832.
Intergovernmental Panel on Climate Change (IPCC), 2000. Land use, land-use change, and forestry. Edited by R.T. Watson, I.R. Novel, N.H. Bolin, N.H. Ravindranath, D.J. Veraro and D.J. Dokken. Cambridge University Press, New York.
Intergovernmental Panel on Climate Change (IPCC), 2001a. Climate change 2001: The scientific basis. Edited by J.T. Houghton, Y. Ding, D.J. Griggs, M. Noguer, P.J. van der Linden and D. Xiaosu. Cambridge University Press, UK.
Intergovernmental Panel on Climate Change (IPCC), 2001b. Climate change 2001: Impacts, adaptation and vulnerability. Edited by J.J. McCarthy, O.F. Canziani, N.A. Leary, D.J. Dokken and K.S. White. Cambridge University Press, UK.
Isebrands, J.G., E.P. McDonald, E. Kruger, G. Hendrey, K. Percy, K. Pregitzer, J. Sober and D.F. Karnosky, 2001. Growth responses of Populus tremuloides clones to interacting elevated carbon dioxide and tropospheric ozone. Environmental Pollution 115 (3): 359-371
Krcmar-Nozic, E., B. Wilson and L. Arthur. 2000. The potential impacts of exotic forest pests in North America: a synthesis of research. Canadian Forest Service, Pacific Forestry Centre, Information Report BC-X-387, ix + 33 p.
Kurz, W.A. and M.J. Apps, 1999. A 70-year retrospective analysis of carbon fluxes in the Canadian forest sector. Ecological Applications 9: 526-547.
Morgan, M.G., L.F. Pitelka and E. Sheviakova. 2001. Elicitation of expert judgments of climate change impacts on forest ecosystems. Climatic Change 49 (3): 279-307
Peterson, E.B. and N.M. Peterson, 1992. Ecology, management, and use of aspen and balsam poplar in the Prairie Provinces, Canada. Forestry Canada, Northern Forestry Centre, Special Report 1. 252 p.
Siegert, F., G. Ruecker, A. Hinrichs and A.A. Hoffmann, 2001. Increased damage from fires in logged forests during droughts caused by El Nino. Nature 414 (6862): 437-440.
Simpson, R. and D. Coy, 1999. An ecological atlas of forest insect defoliation in Canada 1980-1996. Canadian Forest Service, Atlantic Forestry Centre, Information Report M-X-206E, 15 p.
Volney,W.J.A. and R.A. Fleming, 2000. Climate change and impacts of boreal forest insects. Agriculture, Ecosystems and Environment 82 (1/3): 283-294.
White, A., M.G.R. Canell, A.D. Friend, M. Parry and M. Livermore, 1999. Climate change impacts on ecosystems and the terrestrial carbon sink: a new assessment. Global Environmental Change 9: S21-S30.
Zhou, L., C.J. Tucker, R.K. Kaufmann, D. Slayback, N.V. Shabanov and R.B. Myneni, 2001. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. Journal of Geophysical Research 106 (D17): 20069-20083.
| [1] Canadian Forest Service,
Natural Resources Canada, Northern Forestry Centre, 5320-122 St., Edmonton,
Alberta, T6H 3S5. Tel: (780)-435-7225 (EHH), (780)-435-7326 (JPB); Fax:
(780)-435-7359; Email: [email protected] |