
0621-B1
A.U. Mallik[1] and B.A. Roberts
Failure of conifer regeneration in the presence of thick ericaceous understory is a widespread phenomenon in Europe and North America, causing significant loss of forest productivity. Plants such as bilberry, crowberry, heather, Labrador tea and sheep laurel are known to cause growth inhibition in conifers such as Norway spruce, Scots pine, Sitka spruce, black spruce, red pine and balsam fir. The ericaceous shrubs grow vigorously following forest harvesting and interfere with conifer regeneration. Competition, allelopathy, nutrient sequestration and poor ecto-mycorrhization have been suggested as reasons for the conifer growth inhibition. Among all these ericaceous plants Kalmia exhibits the strongest growth inhibitory effect on black spruce. Traditional vegetation control techniques, such as herbicide application, scarification and plowing have been met with limited success. Practical methods must be developed to regenerate conifer in post-harvest Kalmia dominated sites.
This paper briefly reviews: a) the possible causes and effects of ericaceous dominance after forest harvesting, b) mechanisms of conifer growth inhibition by ericaceous plants, c) challenges in Kalmia control and d) presents the results of a ten year-old field trial showing mortality and growth response of black spruce, Japanese larch, jack pine and red pine planted in a Kalmia dominated site. We hypothesized that conifer response to Kalmia is species-specific and the use of alternative conifer can alleviate forest regeneration in Kalmia sites. Five years after planting, height growth of black spruce in Kalmia sites was 30% less than comparable sites without Kalmia. Height of jack pine, larch and red pine was increased by 66, 45 and 16% respectively compared to black spruce. Basal diameter of jack pine, larch and red pine was increased by 120, 80 and 70% compared to black spruce at the same time. However, larch experienced high mortality (31%). Ten years later black spruce growth remained poor compared to the other conifers. At this time best growth was found in larch, followed by red pine and jack pine but the latter suffered from European sawfly infestation. We conclude that planting alternative conifer species other than black spruce can enhance forest renewal in Kalmia dominated sites.
Significant loss of forest productivity is caused by natural regeneration failure as well as stunted growth of planted conifers in the presence of thick ericaceous understorey plants that dominate post-harvest sites in coastal oceanic and temperate forests of Europe and boreal and temperate rain forests of North America (Jaderlund et al. 1997; Pellissier 1993, 1994; Mallik 1994; Prescott et al. 1994; Fraser 1993). Conversion of multi-story forest structure to simple shrub dominated plant communities also has serious implications for plant and animal diversity (Mallik 1995; Bloom 2001). Plants such as bilberry (Vaccinium myrtillus) in sub-alpine France, crowberry (Empetrum hermaphroditum) in Finno-Scandinevia, heather (Calluna vulgaris) in the U.K. and western Europe, salal (Gaultheria shallon) in coastal British Columbia and American northwest, Labrador tea tea (Ledum groenlandicum) in central Canada and sheep laurel (Kalmia angustifolia, here after referred to as Kalmia) in eastern Canada are known to cause growth inhibition in conifers such as Norway spruce (Picea abies), (Mallik and Pellissier 2000) Scots pine (Pinus sylvestris) (Zackrisson and Nilson 1992), western hemlock (Thuja heterophylla), Sitka spruce (Picea sitchensis), western red cedar (Tsuga plicata), amabilis fir (Abies amabilis) (Prescott and Weetman 1994; de Montigny and Weetman 1990), and black spruce (Picea mariana) (Krause 1986; Mallik 1987, 1994, 1995; Inderjit and Mallik 1996a,b), red pine (Pinus resinosa) (Mallik and Roberts 1992) and balsam fir (Abies balsamea) (Thompson and Mallik 1989). The ericaceous understory plants grow vigorously mainly by vegetative methods following forest harvesting and interfere with conifer regeneration (Mallik 1993). Among all these ericaceous plants Kalmia exhibits the strongest growth inhibitory effect on black spruce. Each year many medium quality black spruce-Kalmia forests in eastern Canada particularly in Newfoundland and northern Quebec have been transforming into Kalmia dominated heaths (Figure 1).
Figure 1. Black spruce-Kalmia forest turned into Kalmia dominated heath by a forest fire 27 years ago near Terra Nova National Park, Newfoundland (photo, AU Mallik).

A systematic survey of 5888 5m2 circular plots (radius 1.26 m) in young black spruce plantations all across central Newfoundland showed 55% of these plantations contain Kalmia (English and Hackett 1994). Black spruce growing in sites with 25-36% Kalmia cover had significantly lower stem density, plant height, basal diameter and foliar nitrogen compared to that growing in contiguous plots with sparse Kalmia (< 1% cover) (Mallik 2001). Several biotic and abiotic factors have been suggested as reasons for conifer growth inhibition in the presence of ericaceous plants (Mallik 1999; Gallet 1994; Gallet et al. 1999; Mallik and Pellissier 2000; Sauto et al. 2008). Traditional vegetation control with herbicide and silvicultural techniques such as scarification and plowing has been met with limited success in controlling Kalmia (Mallik 1991). Practical method must be developed to regenerate conifers in post-harvest Kalmia dominated sites. Objectives of the present paper were to briefly review a) the possible causes and effects of ericaceous dominance after forest harvesting and fire in relation to conifer regeneration, b) challenges and opportunities in controlling Kalmia to enhance black spruce regeneration, and c) to present the results of a 10 year old field trial conducted in central Newfoundland by planting black spruce, jack pine, red pine and larch. We hypothesized that conifer response to Kalmia would be species specific and the use of conifers other than black spruce would enhance forest regeneration.
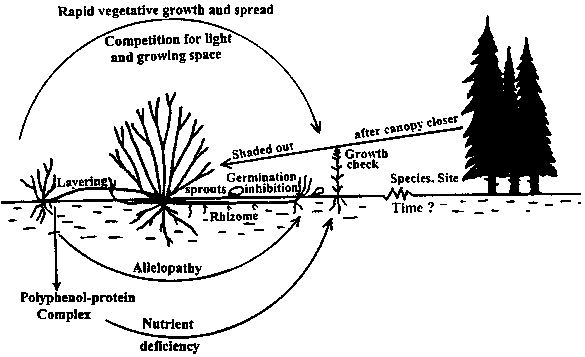
Causes and effects of ericaceous dominance in relation to conifer regeneration: Competition and allelopathy resulting from the rapid vegetative regeneration of ericaceous plants following forest canopy removal by harvesting, high rate of phenolic acid-rich litter accumulation which cause long-term soil changes through nutrient sequestration and iron pan formation are thought to be the main reasons for conifer regeneration failure in post-harvest ericaceous dominated stands (Mallik 1998). However, the degree of conifer growth inhibition is relative to specific conifer-ericaceous combination. For example Kalmia can induce black spruce growth inhibition for 40 years or more (Bloom 2001) but black spruce growth in presence of Ledum lasts for about six years (Inderjit and Mallik 1996b). In terms of mycorrhizal association conifers appear to be at a competitive disadvantage in the presence of ericaceous plants (Yamasaki et al. 1998). Bending and Reid (1996a,b) demonstrated that ericoid mycorrhizae can break down the phenol-protein complexes of ericaceous humus and thus help the ericaceous plants with their N availability but the commonly occurring conifer ectomycorrhizae can not do the same for conifers. Therefore, conifers tend to suffer from a relatively more N deficiency than the ericaceous plants in a nutritionally poor soil. Long-term occupancy of ericaceous plants in a site can reduce and often eliminate certain conifer mycorrhizae in the absence of host making it more difficult for those conifers to grow in an ericaceous-dominated heath. However, not all conifer species are equally susceptible to the phenolic acids of all ericaceous litter. The phenolic acid profiles of all ericaceous plants are not the same and they are not equally inhibitory to all conifers. Unfavorable soil microbial ecology in the presence of ericaceous plants have also been suggested as a reason for the conifer growth inhibition (Pellissier 1998; Pellissier and Sauto 1998; Mallik and Pellissier 2000; Sauto et al. 1998). Thus a combination of biotic and abiotic factors seems to be responsible for conifer growth inhibition in the presence of ericaceoius shrubs (Figure 2).
Figure 2. Conifer growth inhibition resulting from combined effects of allelopathy, competition, phenol-bound nutrient sequestration and ericoid mycorrhizae of the ericaceous plants. Some conifer regeneration can occur over time as regeneration islands amongst Kalmia heath forming a wood-savannah type vegetation (after Mallik 1998).
The success of conifer regeneration after forest harvesting in many ecosystems with ericaceous understory is predicated upon the control of ericaceous plants. Regeneration strategies of ericaceous plants after disturbance and their leaf anatomy are as such that the traditional vegetation control strategies are often of limited success. For example, the commonly used herbicides such as glyphosate (Roundup), Velpar (Hexazinon or Pronone) and Krenite (Fosamine ammonium) at the recommended doze or higher were not found to be effective in controlling Kalmia. However, Garlon (Trichlopyr) killed most of the above and bellow ground components of the plant (Mallik and Inderjit 2000). Treatments such as cutting and burning can not control the plant either but mulching showed a good promise (Mallik 1991). Although mulching treatment has some potential the high cost and difficulty in operating a mulching machine in stony soils are the obstacles against its wider use in Kalmia control. To be cost effective a new form of mulching equipment that consumes less energy and capable of working in stony soils must be developed. Plowing followed liming and nitrogen fertilization has been used in Scotland in afforestation of Calluna heathlands but multiple application of N is required to maintain normal growth of planted conifers (Gimingham 1972). Plowing and scarification in Kalmia- and salal dominated sites failed to control the ericaceous plants and remove growth-check and visible symptoms of N deficiency in conifers (Mallik 1995; Prescott and Weetman 1994). High doze of N and P application as urea (1344 and 672kgN/ha) in a jack pine stand in Quebec was reported to reduce Kalmia and blueberry (Vaccinium angustifolium) (Kumi 1984). Remeasurment of these plots 14 years later still showed significant reduction of Kalmia (Prescott et al 1995) and the authors recommended N application as an option for controlling Kalmia in conifer plantation. However, Mallik (1996) found vigorous growth of Kalmia with N applications in factorial combinations with P and K in greenhouse condition. Prescott and Weetman (1994) showed that combined treatment of salal removal by cutting or herbicide followed by dense planting of conifers and subsequent N and P fertilization can stimulate conifer growth and eradicate salal. However, at the operational level this treatment combination promises to be very expensive.
The main objective of this field trial was to determine the survival and growth performance of four conifers in the presence of Kalmia. We hypothesized that variations in survival and growth conifers in the presence of Kalmia will be species specific.
Study site: The study site was located at the Thomas Howe Demonstration Forest, Gander, Newfoundland. The site was dominated by the black spruce with Kalmia in the understory and feather moss at the ground level. Other associated tree species were balsam fir, eastern larch, and occasional white birch and red maple. The site belonged to forest capability class 4 originating from a wildfire in 1899. The site was fairly typical of the Central Newfoundland Ecoregion, (Damman 1964,1983, Meades and Moores 1989). The soils of the study area consisted of moderately well drained Orthic Humo Ferric Podzols on the medium textured tills (Meades and Roberts 1992).
Methods: The 87ha site was harvested by clearcutting in 1987-88, scarified and planted in 1992 with 2-0 container stock black spruce, jack pine, Japanese larch and red pine. Kalmia grew rapidly after clearcutting with cover ranging from 70-100%. The conifers were planted in large blocks within which ten randomly located 20mX20m plots were marked with wooden stakes. Mortality, height, root collar diameter and current years leader length of the planted seedlings in each plot was determined five years after planting. Plot wise ten-year re-measurement of the trial had to be postponed for one year due to lack of funds. However, 50 saplings of each species at random were measured for height, basal diameter and current years leader length to assess the ten-year growth performance of the conifers. Assessment of Kalmia cover in the plots was noted.
Results: Five years after planting height growth of black spruce in this site was 30% less compared to similar sites in central Newfoundland that are relatively free from Kalmia. In this site height of jack pine, larch, and red pine was increased by 66, 45 and 16% and basal diameter increased by 120, 80 and 70% compared to black spruce at the same time. Black spruce growth was consistently poor in all plots compared to the other three conifers (Table 1). Larch suffered very high mortality (31%) and jack pine had the lowest mortality five years after planting. At this time stem height and diameter growth of Jack pine was the best of all the conifers. However, growth of larch was substantially higher than all other conifers after ten years. At this time jack pine growth was poorer than red pine (Table 1). European sawfly infestation was evident in jack pine. Basal diameter and leader length of jack pine was also very poor at this time. While larch and red pine showed 202 and 192% increase in leader length between five to ten years jack pine experienced 0 increase in leader length. In over all change of growth parameters of the trees from 5 to 10 years the best performer was larch, followed by red pine, jack pine and black spruce (Table 1). Kalmia cover remained high (60 -80%) in the site. However, high growth of larch has began to produce shading effect on Kalmia causing its poor growth.
Table 1. Mortality and growth of black spruce, jack pine, red pine, and larch 5 and 10 years after planting in a post-harvest, Kalmia dominated site in Gander, central Newfoundland. Values are mean followed by standard error of mean in bracket.
| |
Mortality (%) |
Height (cm) |
Root collar diameter (cm) |
Leader length (cm) |
||||||
|
5 yrs |
5 yrs |
10 yrs |
% change |
5 yrs |
10 yrs |
% change |
5 yrs |
10 yrs |
% change |
|
|
Black spruce |
9.4 (4.3) |
56.0 (.8) |
89.7 (3.6) |
60.2 |
0.96 (0.02) |
1.95 (0.11) |
103.1 |
7.9 (0.2) |
8.3 (1.0) |
5.1 |
|
Jack pine |
3.8 (0.8) |
92.6 (1.1) |
191.7 (5.9) |
107.0 |
2.20 (0.21) |
3.56 (0.15) |
61.8 |
16.3 (0.3) |
16.3 (0.8) |
0 |
|
Red pine |
10.7 (3.2) |
65.2 (0.9) |
141.5 (4.5) |
117.0 |
1.74 (0.05) |
4.67 (0.22) |
168.4 |
10.2 (0.2) |
29.8 (1.2) |
192.2 |
|
Larch |
30.9 (4.3) |
80.8 (1.7) |
271.1 (8.3) |
235.5 |
1.78 (0.04) |
4.92 (0.16) |
176.4 |
17.3 (0.4) |
52.2 (2.2) |
201.7 |
Ericaceous understory plants of widely different geographic locations can induce growth inhibition in several conifers. Mechanism of conifer regeneration failure in the presence of ericaceous plants is complex and several biotic and abiotic factors such resource competition, chemical interference, nutrient sequestration and unfavorable rhizosphere microbial ecology were attributable to this growth inhibition phenomenon. Control of ericacious plants and release of growth check of conifers by traditional vegetation control methods are not effective. However, since the degree of conifer growth inhibition in the presence of ericaceous plants appears to be species specific there is opportunity for developing innovative methods of afforestation in ericaceous thickets. Planting alternate conifer species is one option. Our experiment shows that conifers other than black spruce perform well in Kalmia dominated site but we have yet to determine what autecological, genetic and ecophysiological properties a conifer enable it grow better than other in Kalmia dominated sites. Factors such as seedling mortality due to browsing, and plantation failure due to insect infestation provide new challenges to be overcome. In our study site high mortality of larch seedlings is attributable to heavy moose browsing since moose density in the study area has been very high (one moose/km2). We conclude that planting alternate conifer species of pines and larch provides one option in achieving forest renewal in Kalmia dominated sites. Similar approach of using alternate species can be applied in other ericaceous dominated sites of other geographic locations.
Bending, G.D. and Read, J.R. 1996a. Effects of soluble polyphenol tannic acid on the activities of ectomycorrhizal fungi. Soil Biol. Biochem. 28: 1595-1602.
Bending, G.D. and Read, J.R. 1996b. Nitrogen mobilization from protein-polyphenol complex by ericoid and ectomycorrhizal fungi. Soil Biol. Biochem., 28: 1603-1612
Bloom, R.G. 2001. Direct and indirect effects of post-fire conditions on successional pathways and ecological processes in black spruce-Kalmia forests. M.Sc. thesis, Lakehead University, Thunder Bay.
Damman, A.W.H. 1964. Some forest types of central Newfoundland and their relationship to environmental factors. Forest Science Monograph No. 8, 62p.
Damman, A.W.H. 1983. An ecological subdivision of the Island of Newfoundland. In:
G. R. South (Ed.), Biogeography and ecology of the Island of Newfoundland. Dr. W. Junk Publishers, Co. The Hague, Netherlands, pp 163-206. de Montignue, L.E. and Weetman G.F. 1990. The effects of ericaceous plants on forest productivity. pp. 83-90. In: Titus, B.D., Lavigne, M.B., Newton. P.F. and Meades, W.J., Eds. The silvics and ecology of Boreal Spruce. For. Can. Inf. Rep. N-X-271, St. John’s Newfoundland.
English, B. and Hackett, R. 1994. The impact of Kalmia on plantation performance in central Newfoundland. Newfoundland Forest ervice, Silviculture Notebook No. 2. 12p.
Fraser, L. 1993. The influence of salal on planted hemlock and cedar on northern Vancouver Island. M. Sc. Thesis, Department of Botany, University of British Columbia, Vancouver, B.C. 116pp.
Gallet, C. 1994. Allelopathic potential in bilberry-spruce forests: influence of phenolic compounds on spruce seedlings. J. Chem. Ecol. 20: 1009-1024
Gallet, C., Nilsson, M-C. and Zackrisson, O. 1999. Phenolic metabolites of allelopathic significance in Empetrun hermaphroditum leaves and associated humus. Plant Soil 210: 1-9.
Gimingham, C.H. 1972. Ecology of heathlands. Chapman & Hall, London.
Inderjit, and Mallik, A.U. 1996a. Growth and physiological responses of black spruce (Picea mariana) to sites dominated by Ledum groenlandicum. J. Chem. Ecol. 22: 575-585.
Inderjit, and Mallik, A.U. 1996b. The nature of interference potential of Kalmia angustifolia. Can. J. For. Res. 26: 18899-1904.
Jaderlund, A., Zackrisson, O., Dahlberg, A. and Nilsson, M-C. 1997. Interference of Vaccinium myrtillus on establishment, growth and nutrition of Picea abies seedlings in a northern boreal site. Can. J. For. Res. 27: 2017-2025.
Kumi, J. 1984. Effect of repeated fertilization and straw application to the organic layers under jack pine and seedling response. M.Sc. Thesis. Faculty of Forestry, University of British Columbia, Vancouver, 111p.
Krause, H. H. 1986. Ericaceous vegetation as a site factor in jack pine growth of New Brunswick plantation. Proceedings of IUFRO Workshop, 7-10 October 1985, Fredericton, N.B., Canadian Forest service, Fredericton, N.B., 182p.
Mallik, A. U. 1987. Allelopathic potential of Kalmia angustifolia to black spruce. For. Ecol. Manage. 20:43-51.
Mallik, A. U. 1991. Cutting, burning and mulching to control Kalmia: results of a greenhouse experiment. Can. J. For. Res. 67:1309-1316.
Mallik, A.U. 1993. Ecology of a forest weed of Newfoundland: Vegetative regeneration strategies of Kalmia angustifolia. Can. J. Bot. 71:161-166
Mallik, A.U. 1994. Autecological response of Kalmia angustifolia to forest types and disturbance regimes. For. Ecol. Manage. 65:231-249
Mallik, A.U. 1995. Conversion of temperate forests into heaths: role of ecosystem disturbance and ericaceous plants. Environ. Manage. 19: 675-684.
Mallik, A.U. 1996. Effect of NPK fertilization on Kalmia angustifolia. For. Ecol. Manage. 81: 135-141.
Mallik, A.U. 1998. Allelopathy and competition in coniferous forests. In: Environmental Forest Science. pp. 309-315. Sassa, K. Ed. Kluwer Academic Publishers, London.
Mallik, A.U. 2001. Black spruce growth and understory species diversity in contiguous plots with and without sheep laurel (Kalmia angustifolia). Agron. J. 93: 92-98.
Mallik, A.U. and Pellissier, F. 2000. Effects of Vaccinium myrtillus on spruce regeneration: testing the notion of coevolutionary significance of allelopathy. J. Chem Ecol. 26: 2197-2209.
Mallik, A.U. and Prescott, C. E. 2001. Growth inhibitory effects of salal (Gaultheria shallon) on western hemlock (Thuia heterophylla) and western red cedar (Thuia plicata). Agron. J. 93: 85-92.
Mallik, A.U. and Inderjit 2000. Kalmia angustifolia: Ecology and Management. Weed Tech. 15:
Mallik, A.U. and Roberts, B.A. 1994. Natural regeneration of red pine on burned and unburned sites in Newfoundland. J. Veg. Sci. 5:179-186.
Meades, W. J. and L. Moores 1989. Forest site classification manual. A field guide to the Damman forest types of Newfoundland. Forest Resources Development
Agreement, FRDA Report 003.
Meades, W. J. and B. A. Roberts 1992. A review of forest site classification activities in Newfoundland and Labrador. The Forestry Chronicle 68: 25-33.
Pellissier, F. 1993. Allelopathic inhibition of spruce germination. Acta Oecologica 14: 211-218.
Pellissier, F. 1994. Effect of phenolic compounds in humus on the natural regeneration of spruce. Phytochemistry 36: 865-867.
Pellissier, F. 1998. The role of soil community in plant population dynamics: is allelopathy a key component? Trend Ecol. Evolut. 13: 407.
Pellissier, F. and Souto, C. 1999. Allelopathy in northern temperate and boreal semi-natural woodland. Crit. Rev. Pl. Sci. 18: 637-652.
Prescott, C.E. and Weetman, G.F. 1994. Salal Cedar Hemlock Integrated research Program: A Synthesis. Faculty of Forestry, University of British Columbia, Vancouver. 85p.
Prescott, C.E., Kumi, J.W. and Weetman, G.F. 1995. Long-term effects of repeated N fertilization and straw application in a jack pine forest. 2. Changes in the ericaceous ground vegetation. Can. J. For. Res. 25:1984-190.
Roberts, B. A. and Mallik, A.U. 1994. Responses of Pinus resinosa in Newfoundland to wildfire. J. Vegetation Science 5:187-196.
Souto, X.C., Chiapusio, G. and Pellissier, F. 1998. Soil microorganisms and plant phenolics: their implication in natural forest regeneration. In: Environmental Forest Science. pp. 301-308. Sassa, K. Ed., Kluwer Academic Publishers, Dorchdrecht, The Netherlands.
Thompson, I.D. and Mallik, A.U. 1989. Moose browsing and allelopathic effects of Kalmia angustifolia on balsam fir regeneration in central Newfoundland. Can. J. For. Res. 9:524-526.
Yamasaki, S.H., Fyles, J.W., Egger. N.E. and Titus, B.D. 1998. The effect of Kalmia angustifolia on growth, nutrition, and ectomycorrhizal symbiont community of black spruce. For. Ecol. Manage. 105: 197-207.
Zackrisson, O. and Nilsson, M-C. 1992. Allelopathic effects by Empetrum hermaphroditum on seed germination of two boreal tree species. Can. J. For. Res. 22: 1310-1319.
| [1] Biology Department, Lakehead
University, Thunder Bay, Ontario, Canada P7B 5E1. Email: [email protected] |