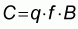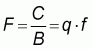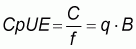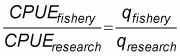![]()
![]()
![]()
An understanding of fishing effort is fundamental for assessing and managing fish stocks. Most management principles involve deciding directly or indirectly upon the amount of fishing effort (f) that should be applied to the stock to obtain a certain amount of catch (C) that is sustainable over time (Rothchild, 1977). Furthermore, the most commonly used contemporary method of estimating the relative abundance of an exploited fish stock is by using the catch per unit effort (C/f) as an index of abundance.
The basic assumption in fisheries theory is that catch (C) and stock abundance, or standing biomass (B) are related by
|
|
(1) |
where f is a measurement of the nominal fishing effort or intensity, and q is the so-called catchability coefficient (defined below). The nominal fishing effort is expressed in for example the number of fishermen, the number of boat-days, the number of meters of gillnet set, the number of hooks set, the number of pulls or shots made, etc. For fisheries data, however, it is generally difficult to measure the nominal effort precisely, and in particular to standardize it in terms of relative fishing power. Unfortunately, due to changes in the catchability coefficient q, there is no necessarily fundamental relation between the magnitude of the nominal effort and the magnitude of the catch. Therefore, for stock assessment purposes, there is a need for a measure of fishing effort that has a constant effect upon the fish population. This measure, commonly used in the population dynamics literature, is the so-called fishing mortality.
The fishing mortality (F) is simply defined as the fraction of the average population taken by fishing. In other words, F can be considered as an invariant measure of effort (Rothchild, 1977). F is also called the instantaneous rate of fishing mortality, i.e. the rate at which fish are dying due to fishing, and therefore expressed per time unit, usually per year. F can be measured without reference to the nominal effort, the configuration of the fishing gear, or the manner in which the gear is employed. F can be defined as
|
|
(2) |
Although F is defined as the fraction of the average population abundance taken by fishing, and therefore one would expect it to take values less than 1, it can in practice have a value of more than 1 on an annual basis for stocks with a high biological regeneration rate. This is because the annual productivity for such stocks, and therefore the cumulated annual catches, can be much higher than the average standing abundance (mean biomass). These are stocks - often smaller-sized tropical fish species - with a so-called high biological turnover, or high production to biomass ratio (P/B). These production concepts are further defined in Appendix 1.
The catch rate (C/f) or Catch per Unit of Effort (CpUE) is the catch per unit of effort over a time interval and defined as
|
|
(3) |
For scientific research surveys, or experimental fishing, effort is standardized and fishing gears kept constant in order to keep a simple relationship between catch rates and population abundance (B), i.e. to minimize the inherent measurement errors and/or variations in f and q. However, this so-called fishery-independent monitoring of stocks through scientific surveys is expensive and surveys often cannot generate the amount of data needed for the evaluation of states or changes in fish stocks or mortality rates (fishing pattern), especially not in the highly diverse tropical freshwater systems discussed in this report.
In many parts of the world, the main supply of information on fishing effort, catch and catch rate is through monitoring of fisheries input (fishing effort) and output (catch), i.e. through fishery-dependent monitoring. Long-term monitoring of fish stocks therefore is almost by necessity dependent on information obtained through the fisheries exploiting them and with that on the official fisheries statistical system in use. Fishery-dependent monitoring entails at least the collection of two essential parameters in fisheries statistics: catch (C) and fishing effort (f) and from these the derivation of catch rate (C/f) (FAO, 1999). CEDRS maintained to address information needs for fishery management vary in their degree of administrative and statistical sophistication, but all share the collection and maintenance of these basic parameters.
Catchability (q) is defined (see equation 3) as the relationship between the catch rate (CpUE) and the true population size (B). So the unit of catchability is fish caught per fish available per effort unit and per time unit. Catchability is also called gear efficiency (Hilborn and Walters, 1992) or sometimes fishing power, and is strongly related to gear selectivity (defined below) because it is species and size dependent. Sometimes gear selection is simply defined as the relative change in q (Godø, 1990). Therefore the fishing mortality (F) as a function of the size (length) of fish, i.e. the fraction of fish caught per fish in the population, has the same shape as the gear selection curve (S), but with a different value depending on the nominal effort.
In other words, when effort (f) is equal to 1 (unit) then:
|
q = F = S |
(4) |
This means that q can conceptually be considered as the probability of any single fish being caught. Therefore q ranges between 0 and 1.
However, the probability of a fish being caught at any time depends on several factors, which are not only man-made, and can broadly be grouped as biological or technological:
Biological factors include:
Technological factors include:
As both the unit and the different notation show, the catchability coefficient (alias efficiency, or fishing power, or probability of a fish being caught), is therefore a composite and very complicated factor. Conceptually, however, ‘fish catchability’ implies primarily changes in fish behaviour (May, 1984), whereas ‘fishing efficiency’ indicates changes in fishing practices (Neis et al., 1999) or in relative fishing power. As information on the possible causes of variation in q is normally lacking, the biological and technological factors are for practical purposes normally assumed invariant of abundance, time, species, size/age, and the individual skills of the fishermen. However, only under this very rigid assumption can the catch rate (CpUE) be considered directly proportional to the stock abundance (Hilborn and Walters, 1992) and be used as an index of the stock size.
Consequently, the catchability (q) cannot be quantified directly if catch rates are used to estimate stock sizes. The standard solution to evaluate changes in efficiency (fishing power) in a fishery over time, and with that the catchability, is therefore to compare catch rates from commercial and research fishing where the catchability of the research fishing is holding constant from year to year (Neis et al.,1999):
|
|
(5) |
This method requires several years of data in order to detect relative changes in the efficiency of the commercial fishery. This lag in time, before eventual changes are discovered, will lead to overestimation of stock size if the commercial fishing efficiency or fishing power is rising (Pope, 1977).
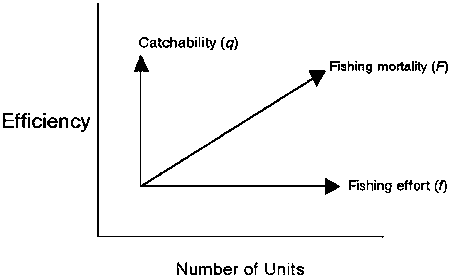
The variability and elusiveness of the parameter q, and the difficulties in quantifying it, is a very important reason for the difficulties in analysing the relationship between the magnitude of nominal effort (which is also difficult to measure) and the direct effect on the regenerative capacity of the stocks. Changes in q, which are mostly unaccounted for, induce additional uncertainty in the parameter catch rate as an index of stock abundance, if q simply varies through time. If there is a unidirectional change in q, as a result of, for instance, increased fishing power of a unit of effort over time, catch rates - and with these stock abundance - will be systematically overestimated. This is also an important reason why, to a large extent, fisheries science is operating with the parameter ‘fishing mortality (F)’ instead of the parameter ‘fishing effort (f)’. Unfortunately, fishing mortality is notorious for its incomprehensibility outside the fisheries scientific community. However, returning to equation (2), the elusiveness of F, and its relationship with nominal effort and efficiency in a biological sense (i.e. the efficiency with which a fish is caught) may become somewhat less obscure when illustrated graphically (Figure 2.1).
A generally important technical measure for fishing gears is the size selectivity which is defined as the probability of fish being retained in a fishing gear as a function of the length of the fish (Misund et al., 2001). These probabilities are often expressed as various mathematical models. A selection curve (i.e. the probability of capture plotted against the size of the fish) for trawl gears is mostly sigmoid or S-shaped, whereas bell-shaped curves are normally the case for gillnets and hooking gears. Important selectivity measures are L50 - defined as the length of the fish where the fish have a 50% probability of being retained by the gear on encounter - and the selection factor - defined as L50 divided by mesh size in cm. In addition to the selection range which is defined as L75 - L25 (L75 is the length of the fish where 75% of the fish is retained, and L25 is the length where 25% of the fish is retained), these parameters describe the size selection characteristics of fishing gears.
Figure 2.1 Fishing mortality (F) as a resultant of nominal fishing effort (f) and catchability (q). The number of units expresses nominal fishing effort, while catchability can be expressed by the efficiency of one unit of fishing effort
All fishing gears are species and size selective: this is also true in multispecies fisheries where one type of gear may catch a set of species, while another gear or the same gear used in a different way or different area may catch another set. This means that all fishing gears are only able to catch a certain portion of the total (multispecies) fish community present. The use of the catch rate as an index of abundance of a fish stock is therefore further complicated by the selectivity of a fishing gear. Catch rates only reflect the abundance of the fishable stock or, in other words, that portion of a fish population or fish community that can be caught by a specific gear. Catch rates can be used as an index of abundance for the total stock, under the assumption that all specimens within a (multispecies) stock at some stage during their life become part of the fishable stock.
The area of operation of a gear, the inconstant behaviour of the fish relative to the gear, and the size of the fish determine the part of a stock that can be caught by a gear. As discussed, these factors are all included in the parameter catchability. Selection may therefore differ in different areas of operation simply because of the species and sizes present on a fishing ground. For example, seines with small mesh sizes operated in shallow areas near shore that act as nursery grounds will have a higher probability of catching juvenile specimens than if they are operated further offshore where these fish are not present. Species behaviour may change seasonally as a function of several factors, such as migrations, spawning or temperature, and with this the probability of catching it with a certain gear will also change. Furthermore, species may develop avoidance behaviour towards gear, which will result in a lower catchability. Some species are notorious for their ability to avoid certain gear, for example the ubiquitous redbreast Tilapia is very difficult to catch with gillnets.
Fishing gears are intrinsically associated with selectivity, and selectivity, or the impact of fishing on an ecosystem, is an essential component of a management programme. The importance of selectivity is therefore a key point for most researchers and managers, and any non-selective capture method automatically carries the connotation of being harmful or destructive, or at least leading to growth-overfishing seen from the traditional single-species perspective. Mesh size and gear restrictions are therefore among the most easily applied and widely used management regulations. Consequently most nations have imposed legislation that bans certain gears and mesh-sizes with the aim of protecting the resource (Gulland, 1982). Although many of these regulations originate from problems associated with the large scale-fisheries (Misund et al., 2001), they are often uniformly applied in all sectors. However, selectivity seems much more of a problem for industrialized fisheries that dump on average about 45% of their catch, while small-scale artisanal discards average only 5% (Bernacsek, 1989) despite the fact that they mostly operate in more multispecies environments. Although numerous authors have already pointed to the problems of defining the “right” mesh-size in a multispecies fishery, the notion of regulations on selectivity still persists. In addition, small-scale fisheries, such as those studied in this report, often use a variety of gears, both traditional and modern. Many of these gears and particularly the traditional ones, such as seines, small mesh-sizes, drive- or beat fishing, barriers and weirs, are often classified as illegal under the pretext of being non-selective with assumed negative impacts on the fish populations.
The preceding section is based on fisheries theory and definitions. It is open to question, however, whether the concepts of ‘nominal fishing effort (f)’ and ‘catchability (q)’ - in particular where it deals with technological factors and their biological impact on the fish stocks - can be readily applied within social and economic treatments of fishing effort. In fisheries economics, for example, fishing is analysed in terms of investments and revenue, and ‘effort’ is therefore conceptually associated with cost. According to Hannesson (1993), it is generally impossible to develop any general valid proportion between the nominal effort in terms of units and fishing effort in the economic sense due to technological differences and/or changes and different inputs of fuel, equipment, manpower, etc. Therefore in fisheries economics, fishing effort (investment) is sometimes defined as the mortality generated on a fish stock because that will more accurately predict the catch (or revenue). This makes effort strictly equivalent with the fishing mortality (F) used in population dynamics:
|
C = F · B =? · f · B |
(6) |
Comparing with equation (2), fisheries economics then appear to have similar problems with the relationship between the nominal effort and the catch as in fisheries biology with the elusive catchability coefficient q.
As for fisheries economics, Brox (1990) does not distinguish between nominal effort and fishing mortality. However, as a social scientist, Brox is concerned with identifying how different ways of changing effort may lead to very different social results. He therefore introduces the distinction between ‘horizontal’ and ‘vertical’ changes in fishing effort (or fishing mortality). Horizontal growth is related to the growth in the number of fishermen: “Fishermen’s children grow up and establish households that base their economy on participating in fisheries.... Fishing communities also absorb people born outside fishing districts, especially through marriages, and young migrants initially looking for temporary employment”(Brox, 1990:233). It is important to note that horizontal growth is different from demographic growth. While demographic growth in the traditional meaning of the concept is understood as a function of birth- and mortality rates, horizontal growth of effort also includes migration and changes in people’s occupations.
Vertical growth is related to growth in capital use and technological level. “The aggregate effort may also increase because each operating unit acquires more technical equipment. This happens when some of the formerly stagnant units begin to accumulate capital, but also as investors from other branches of the economy, other regions or even other countries somehow decide to participate in harvesting the resources.” (Ibid.)
These concepts, which for the sake of clarity we have renamed population-driven and investment-driven growth in fishing effort, seem to come very close to the above distinction between nominal changes of effort and changes in efficiency. From a biological point of view, however, it is the interaction between the catching process (gear) and the fish that is important. Thus, with regard to ‘numerical growth of fishing effort’, it is the total number of units of gear and their employment that matters. Who invests and at what level is irrelevant for the fishing mortality: 10 fishermen owning 100 nets each is the same as 100 fishermen owning 10 nets each. Investment-driven change in effort is therefore not directly equal to the biological term ‘catchability’, but more related to ‘efficiency’. The two concepts will only coincide if the unit of fishing effort is understood to be production units and if the biological variables (e.g. behaviour) included in catchability are excluded.
The importance of distinguishing between socio-economic processes leading to simply numerical changes in nominal fishing effort or increased efficiency from capital investments, lies in the different effect each of these processes has on the sustainability of exploitation. The hypothesis is that when investment-driven growth of effort accelerates, it represents a potentially much bigger growth in absolute fishing mortality than that expected to be caused by demographic processes. This expectation is based on the presumption that investment-driven growth lead to technological changes towards more effective gear, whereas population-driven growth, just leads to ‘more of the same’. It is important not to consider population-driven changes in effort as a mere reflection of the demographic changes in a community. As mentioned above, demographic change is generally defined as the difference between birth rates and death rates in a given population, but population-driven change in fishing effort - as indicated by Brox - also includes changes due to migration or to changes in occupations. As will be shown, these variables are sometimes more important than the demographic changes.
Institutions constitute a central element in analysis of effort development. In fisheries management literature, institutions are often seen as government’s way of organizing their fisheries administration. We see institutions as much more than this. In recent years the ‘neo-institutionalist’ approaches in economy and political science (e.g. Elinor Ostrom, Douglas North, Oliver Williamson and Jean-Philippe Platteau) has dominated the understanding of social institutions. A commonly used definition is: “Institutions are rights and rules that provide a set of incentives and disincentives for individuals geared to minimize transaction costs.”
Several scholars have pointed out that the focus on transaction costs and market imperfection is too narrow and neglects the historical dimension in how rules have been constructed. From a sociological perspective, Scott (1995) brings in the cognitive aspects and argues that institutions are cultural, social structures and routines. From social anthropology, Berry tends to view institutions “...not as the rules themselves, but as regularised patterns of behaviour that emerge from underlying structures or sets of ‘rules in use’. ... Rather than existing as a fixed framework, ‘rules’ are constantly made and remade through people’s practices” (Leach et al., 1999: 237). The works of, for instance, Berry (1989, 1993), Bierschenk and Olivier de Sardan (1998) and Peters (2000), convincingly argue that local institutions crucial in the regulation of people’s access to vital resources are often unclear and even lack coherence. Their research shows that so-called ‘local management systems’ have often emerged as a result of negotiation and accommodation in long-term power struggles. Various actors, including a variety of external powers operating at different periods of time, have based their claims and legitimacy on different ‘logics’ and values. The lack of hegemonic power at the local level has facilitated their continued co-existence and has entailed an institutional landscape where rules are characterized by ambiguity and sometimes even by contradiction.
Our task here is not to support either the neo-institutional or the more constructivist approach to institutional analysis, but simply to point out the differences and alternatives that they represent in the analysis of regulation and access to natural resources. In the same way that the ‘management’ and the co-management approaches represent alternative ways for understanding existing management regimes, studies of African fisheries have shown how both approaches have important contributions to provide (Jul-Larsen, 1999). While the neo-institutional approach may shed light on the fact that artisanal fisheries sometimes seem to out-compete more industrialized fisheries, the constructivist approach may help us understand important relationships between the local institutional landscape and prospects of economic development.
We will also emphasize that institutions can be either formal or informal and may be created or evolve over time. The nature of institutions can be explained by the use of an iceberg analogy. The top, visible part of the iceberg can be taken as the formal and written institutions, whereas the lower part of the iceberg, which is not visible but exists, can be taken as the informal and unwritten institutions. Both formal and informal codes may be violated and therefore punishments are enacted. The essential part of the functioning of institutions is thus determined by whether their codes can be enforced, the cost of enforcement and the severity of the punishment.
![]()
![]()
![]()