![]()
![]()
![]()
T. T. KOZLOWSKI Y T. E. GREATHOUSE
El presente trabajo se basa en las investigaciones realizadas, en 1969, por el Profesor Kozlowski, en su calidad de consultor de fisiología forestal del proyecto de la FAO en Malasia, denominado «Plantaciones experimentales de especies forestales industriales de crecimiento rápido». Actualmente trabaja en el Department of Forestry, University of Wisconsin, Madison, Wisconsin, Greathouse trabaja en el mismo proyecto, en el Instituto de investigaciones Forestales de Kepong, Selangor (Malasia).
En los últimos años, se ha observado especialmente en relación con los muchos proyectos del PNUD/Fondo Especial ejecutados por la FAO, un interés cada vez mayor en las posibilidades de crecimiento de los pinos exóticos en el trópico como, por ejemplo, Pinus caribaea var. hondurensis. En Malasia, las principales características que han estimulado la plantación de esta especie son un alargamiento rápido y precoz, que alcanza hasta 275 cm al año, una producción de alrededor de 14 m³ por ha y por año y la ausencia de plagas importantes de hongos e insectos. En el presente trabajo, se informa sobre las características del desarrollo de las ramas de los pinos exóticos en los países tropicales y su regulación, y se señalan algunos campos de investigación en que los fisiólogos pueden hacer grandes contribuciones.
Muy poco es lo que se sabe sobre los mecanismos que regulan el crecimiento de las ramas, fuera de la semejanza que presentan los de especies tales como Pinus taeda y P. elliottii, plantadas o en la zona templada (sudeste de los Estados Unidos) o en el trópico. En ambos casos, el alargamiento se produce en presencia de más de una yema terminal por rama y por año.
Al mismo tiempo que se dilatan los entrenudos de la primera yema, al comenzar el período vegetativo, entra en actividad el meristema apical, para producir los primordios que se convertirán en las hojas y en el eje de una nueva yema terminal, la cual se forma rápidamente y, por lo general, se dilata (experimenta una exaltación vegetativa) para iniciar la segunda etapa de alargamiento de la rama, en el mismo año, que suele comenzar poco después de la formación de la yema. En un solo año, pueden formarse y dilatarse sucesivamente nuevas yemas, en cualquiera de las ramas.
El número de etapas de alargamiento del tallo, en un año, varía según el individuo, la especie y el clima. En un solo año, se ha observado el alargamiento de hasta siete yemas sucesivas, en una sola guía terminal (Wakeley y Marrero, 1958). El número de etapas de alargamiento suele estar en relación directa con el vigor del árbol. La guía de un individuo normal de P. taeda no suele presentar más de dos o tres etapas de alargamiento al año, pero hay algunos pinos que se pueden hacer crecer durante todo el año plantándolos en un medio favorable (Allen y McGregor, 1962).
Los individuos que presentan muchas etapas sucesivas de alargamiento de yemas al año pueden, o no, tener períodos de inactividad bien definidos entre una y otra. Tepper, 1963, informó que, entre una y otra etapa de alargamiento, en dos especies de pino, autóctonas de New Jersey (P. echinata y P. rigida), cesa la actividad de la yema, corroborando este hecho las observaciones hechas por Kramer (1957), en P. taeda plantado en ambiente casi óptimo, controlado. Pero se han hecho otros estudios, en los que se ha llegado a la conclusión de que algunas ramas terminales se alargan ininterrumpidamente durante gran parte del período vegetativo anual, incluso en la zona templada. Eggler (1961) informó que el crecimiento de las ramas de individuos jóvenes de P. taeda se debía al alargamiento de varias yemas terminales sucesivas. Tan pronto como la primera yema terminal del año dejaba de crecer, se formaba una segunda, que comenzaba a dilatarse rápidamente. Allen y Scarbrough (1969) observaron también un alargamiento relativamente continuo, en individuos de diez años, de P. palustris, en Misisipí. Se han observado respuestas semejantes en dos pinos del Mediterráneo (P. halepensis y P. laricio).
El predominio apical de las yemas terminales de las ramas superiores lo demuestra el número de las que se alargan sucesivamente en un año. Por ejemplo, Eggler (1961) informa que, en los Estados Unidos, las ramas superiores de los pinos del sur presentaron hasta cuatro etapas de alargamiento en un año y las inferiores, cero o una. Fielding (1965) informa de un fenómeno semejante, en P. radiata, plantado en Australia.
El crecimiento normal en el trópico de las ramas de los pinos es semejante al de la zona templada. El eje se alarga gracias a la formación de yemas sucesivas en la guía terminal del tallo principal. Después de un período de dilatación, cesa brevemente el crecimiento del tallo y se forma un nuevo racimo de yemas terminales. Poco después, las yemas recién formadas se dilatan para hacer crecer aún más la guía terminal y producir un verticilo de ramas laterales. Suelen presentarse de dos a cuatro períodos vegetativos al año, alargándose la guía terminal de 30 a 60 mm en cada uno, por término medio.
FOTO: S. HO, INSTITUTO DE INVESTIGACIÓN FORESTAL, KEPONG, MALASIA
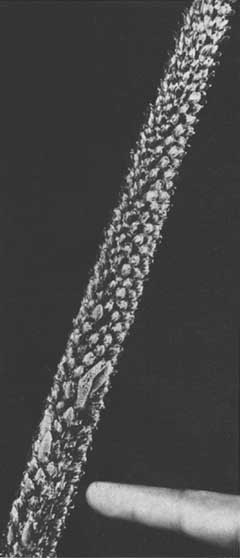
Hay, por el contrario, algunos pinos que presentan un desarrollo anormal (Figura 1) porque no producen yemas capaces de alargarse para formar las ramas laterales. Lloyd (1914) dio a este fenómeno el nombre de «cola de zorro», por la forma cónica de la parte superior de la rama que ha crecido de manera anormal (Figura 2). Constituye una forma sorprendente de predominio apical y suele presentarse en árboles que tienen hasta 6 y, en algunas ocasiones, 12 m de tallo sin ramas. Esta respuesta ambiental plantea un problema más o menos grave en todos aquellos lugares del trópico donde se plantan pinos. Entre las especies que presentan el fenómeno de la cola de zorro se encuentran: P. canariensis, P. caribaea, P. cembroides, P. echinata, P. elliottii P. kesiya (insularis), P. merkusii, P. oocarpa, P. palustris, P. radiata, P. taeda y P. tropicalis.
En la cola de zorro las agujas suelen ir disminuyendo de largo, a medida que se van acercando al extremo, cerca del cual las que no se han abierto se encuentran muy apretadas dentro de la vaina, entera aún. Más abajo, las agujas atraviesan la vaina y se alargan, dándole forma cónica a la parte superior de la rama, que se dilata ininterrumpidamente (Figura 2). Los primordios de las agujas nuevas se forman, de manera más o menos constante, en el ápice y, después, se dilatan, presentando toda una gama de tamaños y etapas de desarrollo de las agujas.
Aparentemente, en los períodos en que se forma la cola de zorro aumenta la retención de agujas, como se ha podido observar en Malasia, donde las porciones desprovistas de ramas del tallo las conservan por varios períodos vegetativos (Figura 1), para perderlas apenas la planta regresa a su curva de crecimiento normal (figuras 3 y 4). A cause del largo período de retención de las agujas, la corteza adquiere un aspecto característico, por los hoyuelos que presenta.
En grandes plantaciones en Malasia de P. caribaea var. hondurensis, de hasta 15 años de edad, se observó la cola de zorro en varios grados. Por ejemplo: (a) en algunos individuos se observó un crecimiento ininterrumpido de la yema terminal, después de plantarlos, formándose un solo tallo, sin ramas laterales (figuras 1 y 3); (b) la cola de zorro empezó a formarse desde el momento de la plantación y cesó, algunos años después, presentando las ramas una curva normal de crecimiento (Figura 4); (c) individuos que crecieron normalmente por un tiempo apareciendo enseguida, la cola de zorro (figuras 5 y 6), (d) la cola de zorro empezó a formarse desde el momento de la plantación, cesó pocos años más tarde y, después, reapareció en las guías terminales, de manera que se observaron cambios en que las ramas pasaban de una etapa de crecimiento continuo a una de repetidos períodos de exaltación vegetativa y viceversa.
A pesar de que en la mayoría de los informes acerca de la cola de zorro se presenta el fenómeno como característico de la guía terminal (Lückhoff, 1964; Lanner, 1964, 1966) en Malasia, se observaron muchos casos de formación de la misma en las ramas laterales (figuras 7 y 8) que, en forma gradual o de repente, asumieron una posición vertical. La parte apical de las ramas con cola de zorro era morfológicamente semejante a la de las guías terminales que presentaban el mismo fenómeno. El crecimiento normal junto con la cola de zorro produjeron ciertos individuos de forma grotesca (Figura 8).
En una comunicación personal, A. F. A. Lamb menciona observaciones que lo llevaron a la conclusión de que la cola de zorro tiende a ser una respuesta juvenil. En Malasia, muchos árboles de las plantaciones más viejas (15 años) todavía presentaban casos de cola de zorro.
La mayoría de los dasónomos consideran que ésta constituye una característica desventajosa, porque está en relación con las quebraduras causadas por el viento, la disminución del crecimiento diametral, la falta de formación de madera de otoño, la presencia de madera de compresión y una menor producción de semilla. Por otra parte, aunque habría que hacer investigaciones sobre las demás propiedades de la madera, es muy posible que llegue el día en que se consideren convenientes los árboles con cola de zorro, cuya madera, por lo demás, está libre de nudos.
EL DESARROLLO DE LOS ÓRGANOS REPRODUCTORES
Los individuos que tienen cola de zorro se diferencian mucho de los normales por las características del desarrollo de sus órganos reproductores. En individuos normales de P. caribaea var. hondurensis, en Malasia, los estróbilos pistilados aparecen en el cuarto superior de la parte verde de la copa pero, en ciertas ocasiones, también en las ramas inferiores, observándose, por lo general, un mayor número en aquéllas (4 a 5) que en ésta (1 a 2). En los individuos normales, la producción de polen tiende a localizarse en las ramas inferiores y en las laterales, de tercero o cuarto orden; pero, en algunas ocasiones, las ramas apicales y también las terminales de las primarias, producen polen. En los individuos normales, de hasta 15 años de edad, hubo escasez de producción de polen, en un día determinado, en relación con las necesidades de los estróbilos receptores, habiendo muchos conos que tenían solamente una o dos semillas viables.
Salvo raras excepciones, en los períodos de formación de una cola de zorro apical no se encontraron estróbilos en los individuos de hasta 15 años de edad, pero, en algunas ocasiones, la rama terminal produjo un gran número (varios cientos con frecuencia) de estróbilos estaminados que, en un momento dado, se encontraban en varias etapas de su desarrollo (Figura 9). Estos estróbilos productores de polen ocupaban el lugar que, en los individuos normales y en la mayoría de los que tienen cola de zorro, suelen ocupar los fascículos de agujas. Cuando un individuo con cola de zorro vuelve a la curva de crecimiento normal, por lo general, la de las ramas presenta etapas repetidas de exaltación vegetativa, produciéndose un verticilo de ramas laterales, donde suelen aparecer estróbilos pistilados, o estaminados, como en los individuos absolutamente normales.
La cola de zorro de los pinos tropicales parece ser un fenómeno debido, en gran parte, a la herencia, cuya expresión se ve muy modificada por los factores de la estación y del clima. En Australia, una plantación de P. caribaea var. hondurensis, procedente de genitores no seleccionados, presentó una frecuencia mucho mayor de cola de zorro que las de var. caribaea o var. bahamensis; pero los descendientes de genitores seleccionados de la var. hondurensis, una mucho menor, dependiendo su disminución del grado de conocimiento de su genealogía (Slee y Nikles, 1968). En Malasia, el porcentaje de cola de zorro en P. caribaea var. hondurensis presentó grandes diferencias, de una estación a otra, tendiendo a aumentar (hasta en un 40 por ciento), en algunas de poca altitud, donde la frecuencia de temperaturas elevadas y precipitaciones copiosas e irregulares en su distribución entre las distintas épocas del año (aproximadamente 254 cm/año) crean condiciones favorables para el desarrollo ininterrumpido de las ramas. La frecuencia de la cola de zorro es, al parecer, menor en las montañas de Camerón, en Malasia, a unos 1 520 m de altitud, que en las estaciones bajas, debido probablemente a que, en las altas, el crecimiento es más lento.
CUADRO 1. - DISTRIBUCIÓN DE LAS PRECIPITACIONES EN El NOROESTE (LANGKAWI Y KEDAH) Y EL CENTRO KUALA LUMPUR Y SELANGOR) DE MALASIA 1
|
|
Langkawi y Kedah ² |
Kuala Lumpur y Selangor ³ |
||
|
Precipitación |
Máxima en 24 horas |
Precipitación |
Máxima en 24 horas |
|
|
Milímetros |
Milímetros |
|||
|
Enero |
34 |
38 |
170 |
132 |
|
Febrero |
46 |
26 |
166 |
126 |
|
Marzo 4 |
113 |
64 |
232 |
119 |
|
Abril |
213 |
71 |
268 |
134 |
|
Mayo |
250 |
145 |
211 |
133 |
|
Junio |
282 |
143 |
122 |
100 |
|
Julio |
274 |
75 |
104 |
92 |
|
Agosto |
286 |
148 |
160 |
112 |
|
Septiembre |
389 |
89 |
183 |
194 |
|
Octubre |
401 |
86 |
281 |
221 |
|
Noviembre 5 |
208 |
64 |
257 |
122 |
|
Diciembre |
32 |
37 |
239 |
108 |
|
Promedio anual |
2 529 |
|
2 394 |
|
1 Datos de la precipitación (1879-1958) recopilados por el Drainage and Irrigation Dept., 1961, Government Press, Kuala Lumpur, Malasia. - ² Estación de Ulu Malaka, a alrededor de 3,5 km del huerto semillero. Datos de 1946-58. - ³ Estación del Hospital de Tanglin. Datos de 1879-1958. - 4 En años normales, los primeros 15 a 20 días de marzo son también secos (o sea < 30 mm). - 5 En el año normal los últimos 15 a 10 días de noviembre son también secos.
FOTO: S. Ho, INSTITUTO DE INVESTIGACIÓN FORESTAL, KEPONG, MALASIA
FIGURA 3. - Individuos normales, con ramas y otros «cola de zorro», sin ramas, de 5 años.
FOTOS: S. Ho, INSTITUTO DE INVESTIGACIÓN FORESTAL, KEPONG, MALASIA
Una estación de la Isla de Langkawi, en el noroeste de Malasia, que tiene una época de sequía bien definida (o sea una precipitación mensual inferior a 61 mm) de dos o tres meses (Cuadro 1), se eligió para hacer un huerto semillero para los ensayos de P. caribaea, donde se tomarán los datos mensualmente, por lo menos en los primeros 18 meses, para determinar si hay o no correlación entre las curvas del aumento de altura, de la cola de zorro y de la precipitación.
Datos procedentes de diversas fuentes confirman una notable influencia del clima sobre la respuesta en cola de zorro. Así, por ejemplo, en Sudáfrica, Lückhoff (1964) observó que su porcentaje disminuye en P. caribaea, a grandes alturas y latitudes, en relación con las bajas temperaturas. Por otra parte, en Zululandia (altitud media 45 a 60 m), la cola de zorro en P. caribaea var. hondurensis era, por término medio, de 43 por ciento; en Ntsubane (aproximadamente 460 m de altitud), de 26 por ciento, y, en Dargal, Natal Central (1 200 m de altitud aproximadamente), de 13 por ciento. Se encontró también una correlación entre el porcentaje de tallos torcidos y la altura sobre el nivel del mar (régimen de temperaturas). En Queensland (Australia), Slee y Nikles (1968) observaron en P. caribaea mayor frecuencia de cola de zorro en Beerwah que en Bowenia y atribuyeron la diferencia al medio más favorable de esta última estación. Hay nuevos datos que confirman la gran influencia que tiene el clima sobre la cola de zorro en P. radiata, siendo escasa su frecuencia en el Territorio de la Capital de Australia, donde el año se divide en épocas bien diferenciadas, y muy grande en Hawaii, donde sucede lo contrario (Fielding, 1960; Lanner, 1966).
También una competencia intensa puede inhibir el desarrollo de las ramas y anular el de la cola de zorro que, por ejemplo, en Honduras Británico, no se presenta en muchos rodales autóctonos de P. caribaea pero sí en algunos suelos donde la competencia de las malas hierbas es leve y desaparecen las limitaciones que se oponen al desarrollo ininterrumpido, poniéndose así de relieve el hecho de que el fenómeno no depende sólo del factor clima.
En Australia, se ha logrado hacer disminuir la frecuencia de la cola de zorro mediante una selección fenotípica intensa, seguida de otra por ensayos de descendencia (Nikles, comunicación personal). En Queensland, en el aclareo precoz de los rodales de P. caribaea var. hondurensis, de 6 a 9 m de alto, se cortaron los árboles que tenían cola de zorro sin que apareciera en los restantes.
REGULACIÓN INTERNA DEL DESARROLLO DE LAS MAMAS
En los mecanismos de regulación hereditarios y ambientales del desarrollo de las ramas de los pinos, en determinadas épocas del año, intervienen procesos fisiológicos internos (Kramer y Kozlowski, 1960; Kozlowski, 1969). Para poder comprender el carácter de la regulación interna del desarrollo de las ramas en determinadas épocas del año, hay que tener presente que su grado varía según el clima. Por ejemplo, los pinos que, en la zona templada, presentan repetidas etapas de exaltación vegetativa (v. gr. P. taeda, en el sudeste de los Estados Unidos), en el verano pasan por varios períodos sucesivos de «quiescencia» de las yemas, la cual puede considerarse como un tipo simple de inactividad reversible, en materia de desarrollo, que se presenta varias veces durante el período vegetativo y tiene por consecuencia múltiples etapas de exaltación al año. Pero, después de la última, que tiene lugar a fines de otoño, las yemas entran, poco a poco, en un estado de reposo vegetativo, muy profundo, que son incapaces de interrumpir varios de los factores ambientales que lo provocaron. En esta situación, las yemas, aunque estén predispuestas a crecer, no lo hacen, aunque se encuentren en un medio favorable, porque hay factores fisiológicos internos que inhiben su desarrollo. Este estado de reposo vegetativo se produce gradualmente. Los tejidos se encuentran en dicho estado solamente a mediados del período de reposo vegetativo invernal, considerado en su conjunto. Las bajas temperaturas, los fotoperíodos breves, o su interacción, inducen fácilmente el reposo vegetativo de las yemas propiamente dicho. En las zonas templadas, el reposo vegetativo de las yemas suele también verse interrumpido por períodos prolongados de bajas temperaturas. Para que las yemas salgan completamente del estado do reposo, tienen que verse expuestas por un número crítico de horas a una temperatura inferior a cierto límite, que varía según la especie. La duración de la exposición al frío, necesaria para interrumpir el reposo vegetativo propiamente dicho, cambia mucho según la especie, la variedad y diversas yemas, en un mismo árbol (Kramer y Kozlowski, 1960).
FOTOS: S. Ho, INSTITUTO DE INVESTIGACIÓN FORESTAL KEPONG, MALASIA
La característica externa do las fases sucesivas que llevan al reposo vegetativo de la yema es la desaceleración de la dilatación de los entrenudos y su fin, mientras que la interna es la aceleración de la actividad del meristema. A la larga, se desarrolla un grupo de primordios, en un eje corto, en la yema que se encuentra en reposo vegetativo.
Cuando un pino de la zona templada (v. gr., P. taeda) que presenta etapas de exaltación vegetativa repetidas se traslada a un clima tropical, deja de estar sometido al medio riguroso que induce un estado de reposo vegetativo muy profundo en la yema. En los trópicos, es más probable que la yema se suma en períodos de quiescencia y no de reposo vegetativo propiamente dicho.
De lo que precede, resulta claro, que, en una teoría general de los mecanismos de regulación interna del desarrollo de las ramas de los pinos, tanto del trópico como de la zona templada, hay que contemplar tres fases primarias de desarrollo de la yema, a saber: (a) evolución hacia un estado de reposo vegetativo propiamente dicho; (b) interrupción del mismo y evolución hacia un estado de actividad vegetativa y (c) comienzo, en la primavera, de la evolución hacia un estado de desarrollo ininterrumpido (Smith y Kefford, 1964). Por lo tanto, la inactividad de las yemas fluctúa entre un estado de quiescencia y otro de reposo vegetativo propiamente dicho; pero como en los pinos también se presentan etapas de transición, la inactividad de las yemas se puede considerar como un estado cuantitativo, tal como se ha hecho en relación con algunas angiospermas (Kawase, 1961).
FOTOS: S. Ho, INSTITUTO DE INVESTIGACIÓN FORESTAL, KEPONG, MALASIA
Hay muchos hechos que prueban la influencia de los factores ambientales en el reposo vegetativo do las yemas, o sea, en la síntesis, traslocación y destrucción, en cantidad variable, de las diversas hormonas reguladoras del crecimiento. Durante muchos años, se pensó que la regulación del reposo vegetativo do las yemas se producía a través de un proceso único y no a través de usa serie de procesos transitorios. Las primeras teorías sostenían que todo dependía de un solo regulador químico endógeno. Se ha insistido mucho en la regulación del reposo vegetativo en virtud de los niveles de auxina solamente. Kozlowski (1964) estudió la bibliografía de estas primeras teorías.
FOTO: S. Ho, INSTITUTO DE INVESTIGACIÓN FORESTAL, KEPONG, MALASIA
Los hechos que se conocen actualmente indican que las interacciones de las sustancias endógenas de crecimiento y de las inhibitorias son las que regulan el reposo vegetativo de las yemas. Si varía la proporción de sustancias de crecimiento, varían también los porcentajes de división y dilatación de las células y, por ende, la regulación del desarrollo y de la diferenciación. Se conocen, por lo menos, tres fases principales, y varias secundarias, de desarrollo general de las yemas, cuya regulación responde a las curvas de las hormonas reguladoras. Las de crecimiento, como, por ejemplo, las auxinas, giberellinas y citoquininas, así como las inhibitorias, según se ha observado, intervienen de varios modos. Estas últimas (v. gr., el ácido abscísico) desempeñan una función clave en la evolución del reposo vegetativo, mientras que aquéllas, sobre todo las giberellinoides, juegan un papel do gran importancia en la exaltación de las funciones vitales. Las interacciones de las diversas sustancias, que estimulan e inhiben el crecimiento, también tienen importancia en la regulación del fenómeno del reposo vegetativo, considerado en su conjunto. Los hechos fundamentales, que indican que el reposo vegetativo y su evolución lo regulan, en gran parte, las sustancias inhibitorias endógenas, pueden resumirse como se indica a continuación: (a) en presencia de un fotoperíodo breve, las hojas de muchas especies leñosas inhiben el desarrollo del extremo de las ramas; (b) cuando el fotoperíodo es breve, se encuentran cantidades mayores de sustancias inhibitorias en las hojas y en las yemas de muchas especies leñosas que cuando es largo, (c) cuando se extraen las sustancias inhibitorias de las hojas de una planta leñosa en estado de reposo vegetativo y se le inyectan a individuos de la misma especie, que no se encontraban previamente en tal estado, cesa el desarrollo de las ramas y se inicia su evolución hacia dicho estado de reposo vegetativo (Wareing, 1965).
Tanto las sustancias de crecimiento como las inhibitorias se hallan presentes en las diversas etapas de transición del reposo vegetativo; al parecer, las sustancias inhibitorias desempeñan una función primordial en relación con éste y las hormonas de crecimiento, con su interrupción, sobre todo, las giberellinoides. Es posible que haya en juego otras sustancias de crecimiento, cuyo equilibrio e interacciones desempeñen algún papel en la exaltación de las funciones vitales. La observación de que los fotoperíodos largos elevan en las yemas los niveles de hormonas de crecimiento viene a confirmar estas conclusiones. Las sustancias inhibitorias no siempre disminuyen al exaltarse las funciones vitales (Wareing, 1965). Se conocen casos de interacciones de las sustancias inhibitorias (v. gr., el ácido abscísico) y las de crecimiento. El ácido giberéllico, por ejemplo, contrarresta los efectos de las sustancias inhibitorias (Thomas, Wareing y Robinson, 1965).
Las variaciones estacionales en cuanto al tipo y cantidad de hormonas de crecimiento que se encuentran en los árboles se ajustan, en cierta medida, a la, teoría de la regulación del crecimiento de las ramas en la que entera en juego la relación entre las sustancias de crecimiento y las inhibitorias. Las yemas en estado de reposo vegetativo contienen una cantidad pequeña de hormonas de crecimiento, mientras que las que están dilatadas o abriéndose, una muy grande. En las de los pinos, se han encontrado sustancias de crecimiento y también inhibitorias (Allen, 1960; Ogasawara, 1961a, 1961b; Giertych, 1960; Giertych y Forward, 1966). Los niveles de las hormonas de crecimiento suelen ser más elevados en las yemas terminales que en las laterales. Giertych y Forward (1966) encontraron los niveles máximos de hormonas de crecimiento en la parte inferior y central de la copa de Pinus resinosa; pero también los niveles de sustancias inhibitorias eran más altos en éstas que en la superior, lo cual parece indicar un saldo favorable de hormonas de crecimiento.
En el presente trabajo, se ha insistido mucho en la regulación hormonal interna del crecimiento de las ramas, cuando las yemas se encuentran en un estado de reposo vegetativo propiamente dicho, lo cual se ha hecho para dar una idea del estado actual de los conocimientos y subrayar la importancia de las relaciones hormonales en la regulación del desarrollo de las ramas; pero, como el estado do reposo vegetativo de las yemas se puede considerar como el grado máximo de inactividad vegetativa de la rama y la quiescencia como su estado intermedio, queda mucho por conocer acerca de las mismas en los pinos tropicales, cuyas yemas, en la mayor parto del año, se hallan más bien en un estado de quiescencia que en uno de reposo vegetativo propiamente dicho. Las investigaciones fisiológicas y bioquímicas de la regulación interna del desarrollo de las ramas en los pinos tropicales han sido muy limitadas, pero los resultados de los estudios fisiológicos del desarrollo de las ramas, de algunas especies que presentan etapas repetidas do exaltación vegetativa, indican que los reguladores hormonales desempeñan una función importante. Se podría avanzar la hipótesis de que la relación entre las hormonas de crecimiento y las inhibitorias cambia, en las sucesivas etapas alternas de exaltación vegetativa y quiescencia, en las ramas de los pinos tropicales. Entre las cuestiones más importantes que aún no han encontrado respuesta, se encuentran:
1. la de las diferentes cantidades y tipos de hormonas de crecimiento e inhibitorias presentes en las sucesivas etapas de exaltación vegetativa, por lee que pasan las ramas de los pinos tropicales;2. la de las diferentes cantidades y tipos de hormonas de crecimiento e inhibitorias, que se presentan, con el tiempo, en la cola de zorro;
3. la de la diferencia que hay entre las cantidades y tipos de hormonas de crecimiento e inhibitorias en una especie que presenta etapas sucesivas de exaltación vegetativa, en la zona templada, al iniciarse y al terminar una etapa de quiescencia, comparada con una que pasa por período de reposo vegetativo propiamente dicho;
4. la de cómo cambia el ciclo anual de las hormonas que regulan el crecimiento de una especie en la zona templada y cómo lo hace cuando la misma se planta en el trópico;
5. la de las diferentes cantidades y tipos de hormonas reguladoras del crecimiento de las ramas presentes en las diferentes partes de la copa, en los pinos tropicales y en aquellos de la zona templada que presentan etapas repetidas de exaltación vegetativa;
6. la de las combinaciones de factores ambientales que inducen diversas relaciones hormonales, que influyen en las curvas previsibles de desarrollo de las ramas.
Hay excelentes oportunidades de encontrar la respuesta a las cuestiones enumeradas anteriormente y contribuir, así, a un mayor conocimiento de la regulación interna del desarrollo en los pinos tropicales. Los datos necesarios pueden ser muy útiles para obtener plantas de forma predeterminada, empleando los materiales genéticos y reguladores ambientales apropiados.
ALLEN R.M. 1960. Changes in acid growth substances in terminal buds of longleaf pine saplings during the breaking of winter dormancy. Physiologia Plantarum, 13: 555-558.
ALLEN, R. M. & MCGREGOR, W. D. H. 1962. Seedling growth of three southern pine species under long and short days. Silvae Genet., 11: 43-45.
ALLEN, R. M. & SCARBROUGH N. 1969. Development of a year's height growth in longleaf pine saplings. U.S.D.A. Forest Research Paper SO-45.
BARRETT, W. H. G. & GOLFARI L. 1962. Descripción de dos nuevas variedades del «Pino del Caribe» (Pinus caribaea Mor.). Caribbean Forester, 23: 59-71.
CHALMERS, W. S. 1962. The breeding of pine (Pinus caribaea Mor.) and teak (Tectona grandis) in Trinidad - some early observations. Caribbean Forester, 23: 100-111.
EGGLER, W. A. 1961. Stem elongation and time of cone initiation in southern pines. For. Sci., 7: 149-158.
FIELDING, J. M. 1960. Branching and flowering characteristics of Monterey pine. Canberra, Forestry and Timber Bureau. Bulletin N° 37.
FREEIZAILLAH BIN CHE YEOM. 1966. Some notes on Pinus caribaea Mor. grown in Malaya. Forestry Department of Malaya, Forest Research Institute. Research Pamphlet N° 54.
GIERTYCH, M. M. 1964. Endogenous growth regulators in trees. Bot. Rev., 292-311.
GIERTYCH, M. M. & FORWARD, D. F. 1966. Growth regulator changes in relation to growth and development of Pinus resinosa Ait. Can. J. Bot., 44: 717-738.
KAWASE, M. 1961. Growth substances related to dormancy in Betula. Proc. Am. Soc. hort. Sci., 78: 532-544.
KOZLOWSKI T. T. 1964. Shoot growth in woody plants. Bot. Rev., 30: 335-392.
KOZLOWSKI T. T. 1969. Tree physiology and forest pests. J. For., 67: 118-123.
KRAMER, P. J. 1967. Some effect of various combinations of day and night temperatures and photoperiod on the height growth of loblolly pine seedlings. Sor. Sci., 3: 45-55.
KRAMER, P. J. & KOZLOWSKI, T. T. 1960. Physiology of trees. New York, McGraw-Hill.
KUMMEROW, J. 1962. Uber Wachstumsanomalien bei Pinus radiata unter Tropischen Bedingungen. Ber. Dtsch. Bot. Gesell., 75: 37-40.
LANGE, P. W. 1966. The occurrence and development of abnormal long leaders in Pinus radiata D. Don in the Cape Midlands. Forestry in South Africa, 7: 1-10.
LANNER, R. M. 1964. Modifications in the growth habit of exotic trees in Hawaii. Proc. Soc. Am. For., 36-37.
LANNER, R. M. 1966. The phenology and growth habit of pines in Hawaii. U.S. Forest Service Paper PSW-29.
LLOYD, F. E. 1914. Morphological instability, especially in Pinus radiata. Bot. Gaz., 57: 314-319.
LÜCKHOFF, H. A. 1964. The natural distribution, growth and botanical variation of Pinus caribaea Mor. and its cultivation in South Africa. Ann. Univ. Stellenbosch, 39: 1-160.
MCWILLIAM, J. R. & RICHARDS, B. N. 1955. Caribbean pine in Queensland, some notes on its behavior and potentialities. Queensland Forest Service Note N° 4.
OGASAWARA, R. 1961a. Studies on auxins and inhibitors in Pinus thunbergii. J. Jap. for Soc., 43: 50-54.
OGASAWARA, R. 1961b. Studies on auxins and inhibitors in the buds of Pinus strobus. J. Jap. for. Soc., 43: 307-310.
ROMBERGER, J. A. 1963. Meristems, growth, and development in woody plants. U.S. Department of Agriculture, Forest Service. Technical Bulletin N° 1293.
SLEE, M. U. & NIKLES, D. G. 1968. Variability of Pinus caribaea (Mor.) in young Queensland plantations. Proc. ninth Commonwealth for Conf., 1-50.
SMITH, H. & KEFFORD, N. P. 1964. The chemical regulation of the dormancy phases of bud development. Am. J. Bot., 51: 1002-1012.
STREETS, R. J. 1962. Exotic forest trees in the British Commonwealth. Oxford, Clarendon Press.
TEPPER, H. B. 1963. Leader growth of young pitch and short leaf pines. For. Sci., 9: 344-353.
THOMAS, T. H., WAREING, P. F. & ROBINSON, P. M. 1965. Action of the sycamore «dormin» as a gibberellin antagonist. Nature, Lond., 205: 1270-1272.
WAKELEY, P. C. & MARRERO, J. 1958. Five-year intercept as site index in southern pine plantations. J. For., 56: 332-336.
WAREING, P. F. 1965. Dormancy in plants. Science Progress, 53: 529-537.
|
REVISTA CANADIENSE DE INVESTIGACIONES FORESTALES En 1971, el Consejo Nacional de Investigación de Canadá iniciará la publicación de una nueva revista internacional: Canadian Journal of Forest Research. La revista publicará artículos o notas en inglés o francés, informando sobre investigaciones científicas originales y sobre los debates por ellas suscitados. Se tocarán todos los aspectos de las ciencias forestales y se recibirán con especial agrado los estudios de carácter interdisciplinario. La publicación será trimestral y comenzará en marzo. Los originales deben enviarse al redactor jefe, Dr. J. L. Farrar, Faculty of Forestry, University of Toronto, Toronto 5, Ontario. El importe de la suscripción anual, en moneda canadiense, es: suscripción transferible, 10 dólares; suscripción personal, 5 dólares. Las suscripciones deben solicitarse a: Administration, National Research Council of Canada, Ottawa 7, Canadá, abonándolas con giro o cheque pagadero al Receiver General of Canada. |
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}