![]()
![]()
![]()
T. T. KOZLOWSKI ET T. E. GREATHOUSE
Le présent document se fonde sur les recherches effectuées en 1969, alors que le professeur Kozlowski assumait les fonctions d'expert-conseil en physiologie végétale dans le cadre d'un projet de la FAO en Malaisie visant à l'établissement de plantations pilotes d'essences à croissance rapide pour la production de bois d'industrie. M. Greathouse est affecté au projet de l'Institut de recherche forestière, à Kepong, Selangor, Malaisie.
Le potentiel de croissance des pins introduits sous les tropiques a suscite ces dernières années un intérêt accru, notamment dans le cadre de nombreux projets du Programme des Nations Unies pour le développement/Fonds spécial exécutés par la FAO. On peut citer en exemple Pinus caribaea var. hondurensis. En Malaisie, la précocité de sa croissance (jusqu'à 275 centimètres par an), sa production d'environ 14 mètres cubes par hectare et par an, ainsi que l'absence d'infestation sérieuse par les insectes et les crypto-games ont contribué à en encourager la plantation. Le présent document traite des caractéristiques et du mécanisme de la croissance des pousses de pins introduits dans les pays tropicaux et signale quelques domaines de recherche auxquels les physiologistes peuvent apporter une contribution majeure.
On ignore presque tout des mécanismes qui régissent la croissance des pousses, si ce n'est qu'ils sont analogues pour les pins comme Pinus taeda et P. elliottii que l'on trouve dans la zone tempérée (sud-est des Etats-Unis) et pour ceux qui peuplent les tropiques. Dans l'une et l'autre de ces deux zones, la croissance comporte souvent l'allongement de plusieurs bourgeons terminaux par pousse et par an.
Alors que les entre-noeuds du premier bourgeon se développent à un stade précoce, le méristème apical entre en activité et donne naissance à des «primordia» qui deviennent les feuilles et l'axe d'un nouveau bourgeon terminal. Ce dernier se forme rapidement et, d'ordinaire, se développe de manière à produire un second allongement la même année, souvent peu après la formation du bourgeon. Sur l'une quelconque des pousses, d'autres bourgeons peuvent ultérieurement se former et se développer pendant la même année.
Le nombre d'allongements de la tige par an varie selon les arbres, les essences et les fluctuations climatiques. Il est arrivé que, sur une seule pousse terminale, jusqu'à sept bourgeons successifs s'allongent en un an (Wakeley et Marrero, 1958). Le nombre d'allongements est souvent lié à la vigueur de l'arbre. Bien que la pousse terminale d'un spécimen moyen de P. taeda ne s'allonge pas plus de deux à trois fois par an, on peut amener certains pins à s'allonger tout au long de l'année en les cultivant dans un milieu favorable (Allen et McGregor, 1962).
Les arbres dont les bourgeons s'allongent plusieurs fois par an peuvent présenter ou non des périodes d'inactivité entre les allongements. Tepper (1963) signale la cessation d'activité du bourgeon entre des allongements chez deux pins originaires du New Jersey (P. echinata et P. rigida); il corrobore ainsi les observations de Kramer (1957) sur P. taeda élevé en milieu contrôlé quasi optimal. Toutefois, d'autres études ont conduit à la conclusion que certaines pousses terminales s'allongent de façon continue pendant une grande partie de la période annuelle de croissance, même, dans la zone tempérée. Selon Eggler (1961), la croissance des pousses des jeunes P. taeda découle de l'allongement successif des bourgeons terminaux. Dès que le premier bourgeon terminal de l'année cesse de s'allonger, un deuxième se forme et commence à se développer rapidement. Allen et Scarbrough (1969) relèvent également un allongement des pousses assez constant sur des P. palustris de dix ans au Mississippi, tandis que l'on observe un comportement analogue chez deux pins méditerranéens (P. halepensis et P. laricio).
La dominance apicale dans les bourgeons terminaux des branches supérieures se reflète dans le nombre d'allongements des bourgeons par an. Eggler (1961) signale qu'aux Etats-Unis les branches supérieures des pins méridionaux s'allongent jusqu'à quatre fois par an, alors que l'allongement est nul ou unique dans les branches inférieures. Fielding (1965) rend compte d'un phénomène analogue chez P. radiata en Australie.
Sous les tropiques, la croissance normale du pin se fait comme dans la zone tempérée. L'axe s'allonge par extension des bourgeons formés successivement sur la pousse terminale de la tige principale. Après une période de développement, la tige cesse brièvement de croître et un nouveau groupe de bourgeons terminaux se constitue.
Peu après, les bourgeons nouvellement formés s'étendent, allongeant encore la pousse terminale et produisant un verticille de branches latérales. D'une manière générale, ces périodes de croissance interviennent de deux à quatre fois par an, la pousse terminale s'allongeant chaque fois d'environ 30 à 60 millimètres en moyenne. Certains pins, par contre, se développent anormalement (figure 1), faute de former des bourgeons qui s'allongeraient pour donner des branches latérales. Lloyd (1914) a qualifié cette croissance de «foxtailing» en raison de la ressemblance que présente la partie supérieure anormalement allongée avec un cône ou une «queue de renard» (figure 2). Le foxtailing, forme frappante de dominance apicale, donne souvent des arbres ayant une tige dépourvue de branches sur 6 mètres, voire sur 12 mètres. Cette réaction au milieu pose un problème plus ou moins grave aux pins cultivés sous les tropiques. Sont sujets, entre autres, à ce phénomène P. canariensis, P. caribaea, P. cembroides, P. echinata, P. elliottii, P. kesiya (insularis), P. merkusii, P. oocarpa, P. palustris, P. radiata, P. taeda et P. tropicalis.
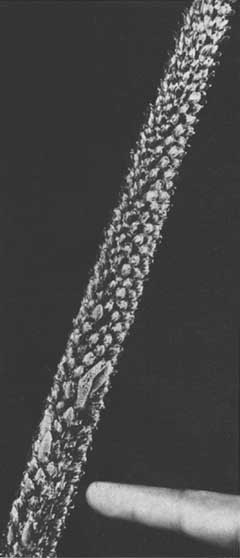
Sur un rameau en «foxtail», les aiguilles se raccourcissent généralement à mesure qu'elles se rapprochent de l'extrémité. Tout près de cette dernière, elles sont étroitement tassées et encloses dans des gaines compactes. Plus bas, les aiguilles percent leurs gaines et s'allongent, conférant à la partie supérieure du rameau en expansion constante un aspect conique (figure 2). De nouveaux primordia d'aiguilles se forment de façon plus ou moins continue au sommet, puis se développent, donnant une succession d'aiguilles de diverses dimensions à divers stades d'expansion.
Il semble que les périodes de foxtailing favorisent la persistance des aiguilles. Comme on l'a observé en Malaisie, les parties de la tige dépourvues de branches conservent leurs aiguilles pendant plusieurs périodes de croissance (figure 1), mais les perdent peu après que l'arbre soit revenu à son allure normale de croissance (figures 3 et 4). Cette persistance des aiguilles sur une longue période confère à l'écorce un aspect piqueté caractéristique.
On a relevé des phénomènes de foxtailing plus ou moins prononcés dans les grandes plantations malaisiennes de P. caribaea var. hondurensis ayant jusqu'à 15 ans d'âge. C'est ainsi que: a) sur certains arbres, la pousse terminale continuait de croître sans arrêt après la plantation en champ donnant une tige unique sans branches latérales (figures 1 et 3); b) le foxtailing se manifestait au moment de la plantation en champ, pour cesser au bout de quelques années. Ensuite la croissance de la pousse revenait à une allure normale (figure 4); c) les arbres se développaient régulièrement pendant un certain temps, puis présentaient du foxtailing (figures 5 et 6); d) le phénomène de foxtailing commençait au moment de la plantation en champ, cessait au bout de quelques années puis reprenait dans les pousses terminales. Ainsi a-t-on observé le passage de l'allongement continu à l'allure de croissance normale et vice versa.
Bien que la plupart des auteurs volent dans le foxtailing un phénomène caractéristique de la pousse terminale (Lückhoff, 1964; Lanner, 1964, 1966), on a observé en Malaisie maints exemples de foxtailing sur les pousses latérales (figures 7 et 8). Peu à peu ou brusquement celles-ci s'orientaient verticalement. Les parties supérieures excisées des branches en foxtail étaient, morphologiquement, analogues à celles des pousses terminales en foxtail. L'alternance de la croissance normale et du foxtailing donne souvent des sujets aux formes grotesques (figure 8).
Dans une communication personnelle, A.F.A. Lamb faisait état des observations qui l'avaient amené à conclure que le foxtailing tend à être une réaction juvénile. En Malaisie, nombre d'arbres dans les plus vieilles plantations (15 ans) présentent encore une croissance en foxtail.
La plupart des forestiers jugent néfaste le foxtailing qui prédispose à la rupture par le vent, limite la croissance en diamètre, empêche la formation de bois d'automne, entraîne la présence de bois de compression et restreint la production de graines. D'un autre côté, quand les autres qualités du bois auront été mieux étudiées, fil pourra être un jour souhaitable de produire des arbres à croissance en foxtail pour obtenir du bois sans noeuds.
CROISSANCE REPRODUCTIVE
Les arbres en foxtail diffèrent surtout des arbres normaux par leurs caractéristiques de croissance reproductive. En Malaisie, les P. caribaea var. hondurensis normaux ont des strobiles pistillés dans le quart supérieur du houppier. Certains sujets en portent aussi sur des branches inférieures. D'une manière générale, le nombre de strobiles pistillés par branche est plus grand sur les branches supérieures: quatre ou cinq par branche supérieure, contre un ou deux seulement par branche inférieure. La production de pollen chez les arbres normaux a tendance à se localiser sur les branches inférieures et les branches latérales du troisième ou du quatrième ordre. Parfois, cependant, les rameaux apicaux aussi bien que les pousses terminales des branches primaires produisent du pollen. Jusqu'à 15 ans chez les sujets normaux, la production de pollen est constamment insuffisante par rapport aux strobiles réceptifs. De nombreux cônes ne contiennent qu'une ou deux graines viables.
Hormis quelques rares exceptions, les arbres allant jusqu'à 15 ans ne portent pas de strobiles pendant les périodes de croissance apicale en foxtail. Cependant, la pousse terminale donne à l'occasion un grand nombre (souvent plusieurs centaines) de strobiles staminés qui, dans un moment donné, se présentent à des stades de développement divers (figure 9). Ces strobiles producteurs de pollen apparaissent à des points où, chez les sujets normaux comme chez les arbres en foxtail se forment des faisceaux d'aiguilles. Lorsqu'un arbre en foxtail revient à une croissance normale et produit un verticille de branches latérales, des strobiles pistillés et/ou des strobiles staminés apparaissent généralement sur ces branches latérales, comme sur les arbres tout à fait normaux.
Il semble que le foxtailing des pins tropicaux soit un phénomène de croissance fortement héréditaire, dont l'expression se trouve modifiée de façon considérable par la station et le climat. En Australie, on a noté que dans des peuplements non sélectionnés de P. caribaea var. hondurensis la fréquence des foxtails était beaucoup plus élevée que dans des peuplements non sélectionnés de var. caribaea ou var. bahamensis. Toutefois, les descendants d'arbres de var. hondurensis sélectionnés avaient moins tendance au foxtailing la diminution de l'incidence dépendant du degré de contrôle exercé sur la parenté (Slee et Nikles, 1968). En Malaisie, P. caribaea var. hondurensis est nettement plus sujet au foxtailing dans certaines stations que dans d'autres. L'incidence se porte (jusqu'à 40 pour cent) dans quelques stations de plaine où la constance des températures élevées et les fortes précipitations non saisonnières (à peu près 2500 millimètres par an) favorisent la croissance continue. Elle est moins forte, semble-t-il, sur les Cameron Highlands à des altitudes d'environ 1 500 mètres, sans doute parce que la croissance est plus lente en altitude.
TABLEAU 1. - VARIATIONS DANS LA RÉPARTITION DES PRÉCIPITATIONS EN MALAISIE DU NORD-OUEST (LANGKAWI, KEDAH) ET CENTRALE (KUALA LUMPUR, SELANGOR) 1
|
Mois |
Langkawi, Kedah2 |
Kuala Lumpur, Selangor3 |
||
|
Précipitations |
Maximum en 24 heures |
Précipitations |
Maximum en 24 heures |
|
|
Millimètres |
|
Millimètres |
|
|
|
Janvier |
34 |
38 |
170 |
132 |
|
Février |
46 |
26 |
166 |
126 |
|
Mars 4 |
113 |
64 |
232 |
119 |
|
Avril |
213 |
71 |
268 |
134 |
|
Mai |
250 |
145 |
211 |
133 |
|
Juin |
282 |
143 |
122 |
100 |
|
Juillet |
274 |
75 |
104 |
92 |
|
Août |
286 |
148 |
160 |
112 |
|
Septembre |
389 |
89 |
183 |
194 |
|
Octobre |
401 |
86 |
281 |
221 |
|
Novembre 5 |
208 |
64 |
257 |
122 |
|
Décembre |
32 |
37 |
239 |
108 |
|
Moyenne annuelle |
2529 |
|
2394 |
|
1 Statistiques pluviométriques (1879-58) réunies par le Département du drainage et de l'irrigation, 1961. Government Press, Kuala Lumpur, Malaisie. - 2 Station d'Ulu Malaka, sise à 3,5 kilomètres environ du verger à graines, données pour 1946-58. - 3 Station de Tanglin Hospital, données pour 1879-58. - 4 Pendant les années moyennes, de la première moitié aux deux tiers du mois de mars, la période est également «sèche» (c'est-à-dire < 30 millimètres). - 5 Dans les années moyennes, du dernier tiers à la moitié de novembre, la période est également sèche.
On a choisi dans l'île de Langkawi, en Malaisie du Nord-Ouest, une station à saison sèche caractérisée par des précipitations mensuelles inférieures à 61 millimètres pendant trois à quatre mois (tableau 1) pour établir un verger à graines de P. caribaea. Des mesures seront relevées mensuellement pendant au moins 18 mois pour déterminer s'il y a corrélation entre la croissance en hauteur, le foxtailing et le régime des précipitations. Diverses sources confirment l'influence très nette du climat sur le phénomène du foxtailing. C'est ainsi qu'en Afrique du Sud, Lückhoff (1964) a constaté la diminution de ce phénomène chez P. caribaea aux altitudes et aux latitudes qui connaissent des températures fraîches. Sur le littoral du Zoulouland (altitude moyenne de 45 à 60 mètres), par exemple, le taux de foxtailing de P. caribaea var. hondurensis est de 43 pour cent en moyenne; il est de 26 pour cent à Ntsubane (altitude d'environ 460 mètres), et de 13 pour cent à Dargal (altitude d'environ 1200 mètres) dans les Midlands du Natal. La proportion des tiges tordues est également liée à l'altitude au-dessus du niveau de la mer (régime thermique). Au Queensland (Australie), Slee et Nikles (1968) ont enregistré des cas plus fréquents de foxtail de P. caribaea à Beerwah qu'à Bowenia, et ils ont imputé cette différence au milieu plus favorable de Beerwah. Une autre preuve de l'effet marqué du climat sur le phénomène de foxtailing ressort des données montrant la rareté de ce phénomène chez P. radiata sous le climat à saisons marquées du territoire fédéral de l'Australie et sa fréquence sous le climat moins saisonnier d'Hawaii (Fielding, 1960; Lanner, 1966).
Une forte concurrence végétale peut également entraver la croissance de la pousse et empêcher le foxtailing. C'est ainsi que nombre de peuplements spontanés de P. caribaea au Honduras britannique échappent à ce phénomène. Sur certains sols toutefois, où la concurrence des herbes est faible, la croissance continue est libre de toutes entraves et les pins présentent le phénomène du foxtailing qui, par conséquent, du point de vue mésologique, ne dépend pas du seul climat.
En Australie, on est parvenu à réduire efficacement le foxtailing grâce à une sélection phénotypique intensive, suivie d'une sélection par essais de descendance visant à éliminer cette caractéristique (Nikles, communication personnelle). Les sujets à queue de renard présents dans les peuplements jeunes sont souvent éliminés lors des éclaircies lorsque les arbres atteignent 6 à 10 mètres de haut. L'éclaircie précoce de peuplements de P. caribaea var. hondurensis au Queensland a permis de supprimer les arbres en foxtail, sans amener l'apparition de ce même phénomène chez les sujets subsistants.
RÉGULATION INTERNE DE LA CROISSANCE
Les facteurs héréditaires et mésologiques de la croissance saisonnière des pins agissent par l'intermédiaire de processus physiologiques internes (Kramer et Kozlowski, 1960; Kozlowski, 1969). Pour tenter de comprendre la nature de ce mécanisme interne, il importe de reconnaître que, selon les différents climats, il agit à des degrés divers pour suspendre la croissance. C'est ainsi que chez certains pins à poussées périodiques de la zone tempérée (par exemple P. taeda du sud-est des Etats-Unis), les bourgeons passent par une série de phases de repos («quiescence») pendant l'été. Ce repos est une forme simple d'inactivation réversible qui se répète plusieurs fois pendant la période de croissance et se traduit par de multiples poussées annuelles. Toutefois, après la dernière poussée annuelle à la fin de l'automne, les bourgeons entrent progressivement dans un profond état de dormance qui ne pourra plus être interrompu par plusieurs des facteurs mésologiques auxquels il est dû. Ces bourgeons peuvent être prédisposés à la croissance mais ils n'y parviendront pas, même en milieu favorable, parce que les conditions physiologiques internes s'y opposent. Cet état de dormance véritable s'installe assez progressivement. Les tissus ne se trouvent en repos profond qu'au milieu de la période hivernale d'assoupissement. La dormance véritable du bourgeon peut être aisément amenée par les basses températures, les jours courts, ou l'interaction des deux facteurs. Dans les zones tempérées, le bourgeon est habituellement tiré de son sommeil par des périodes prolongées de basses températures. Pour éliminer totalement la dormance, il faut exposer les bourgeons à une température basse limite pendant un certain nombre d'heures. La durée d'exposition au froid nécessaire pour interrompre la dormance véritable du bourgeon varie énormément selon les espèces, les variétés ainsi que les différents bourgeons des mêmes arbres (Kramer et Kozlowski, 1960).
Extérieurement, les phases successives amenant l'état de dormance se caractérisent par le ralentissement, puis l'arrêt de l'expansion du mérithalle. Intérieurement, toutefois, la mise en dormance implique une activité intensifiée du méristème. A la longue, un groupe de primordia se développe sur un axe court du bourgeon en sommeil. Lorsqu'un pin à poussées périodiques de la zone tempérée (par exemple P. taeda) est transféré sous un climat tropical, il n'est pas soumis au milieu rigoureux qui suscite un profond état de dormance. Il est vraisemblable que, sous les tropiques, les bourgeons de cet arbre seront plus sujets à des périodes de quiescence qu'à une véritable dormance.
De ce qui précède, il semble évident que toute théorie générale sur le mécanisme de la régulation interne de l'expansion des pousses de pins, tant dans les zones tropicales que tempérées, doit tenir compte de trois phases principales du développement du bourgeon: a) mise en sommeil progressive amenant à l'état de dormance véritable; b) interruption de cette dormance et passage à l'état de non-dormance, et c) début de la croissance au printemps menant à une phase de développement régulier (Smith et Kefford, 1964). En d'autres termes, l'inactivité du bourgeon peut aller de la quiescence à la dormance véritable. Etant donné toutefois la manifestation de phases transitoires, l'inactivité du bourgeon chez les pins peut se caractériser comme un état quantitatif, ainsi qu'on l'a fait pour certaines espèces d'angiospermes (Kawase, 1961).
Il est abondamment prouvé que le milieu influe sur la dormance du bourgeon en régissant la synthèse, le transfert ou la destruction de diverses quantités et sortes d'hormones régulatrices de la croissance. On a longtemps pensé que la dormance du bourgeon relevait d'un processus unique plutôt que d'une série de processus transitoires. Selon maintes théories anciennes, le processus, dans son ensemble, serait limité par un seul régulateur chimique endogène. On insistait beaucoup sur la régulation de la dormance au moyen des seuls niveaux d'auxines. Kozlowski (1964) a passé en revue ces premières théories.
Les preuves dont on. dispose maintenant montrent que la dormance du bourgeon est régie par les interactions entre promoteurs et inhibiteurs endogènes de la croissance. Divers dosages de promoteurs de croissance peuvent amener des taux différents de division et d'expansion cellulaires, réglant ainsi la croissance et la différenciation. On reconnaît dans le développement général du bourgeon au moins trois phases majeures et plusieurs phases mineures, chacune déterminée par l'interaction des régulateurs de croissance.
Il semble qu'entrent en jeu, à des degrés divers, des promoteurs tels que les auxines, les gibberellines et les cytokinines, ainsi que des inhibiteurs. Ces derniers (par exemple l'acide abscisique) favoriseraient puissamment l'apparition de l'état de dormance alors que les promoteurs de croissance, notamment les substances analogues à la gibberelline, contribueraient largement à son interruption. Les interactions entre les divers promoteurs et inhibiteurs de croissance sont également importantes pour la régulation du phénomène général de dormance. Les arguments principaux, prouvant que la manifestation de la dormance est en grande partie régie par des inhibiteurs endogènes, peuvent se résumer comme suit: a) en période de jours courts, les feuilles de nombreuses plantes ligneuses arrêtent la croissance du sommet de la pousse; b) les feuilles et les bourgeons de nombreuses plantes ligneuses contiennent de plus grandes quantités inhibiteurs en période de jours courts qu'en période de jours longs; c) lorsque l'on extrait les inhibiteurs des feuilles d'une plante ligneuse en sommeil pour les administrer à des plantes de la même espèce, l'allongement de la pousse cesse et le processus de dormance se déclenche (Wareing, 1965).
Promoteurs et inhibiteurs sont présents pendant les diverses phases transitoires de dormance. Alors que les inhibiteurs paraissent jouer un grand rôle dans l'incitation à la dormance, il semble que l'interruption de cette dernière soit liée dans une large mesure à l'activité de promoteurs hormonaux, notamment de substances analogues à la gibberelline. Il se peut qu'interviennent d'autres promoteurs de croissance et que leurs équilibres et interactions influent sur l'interruption de la dormance. Le fait qu'en période de jours longs les niveaux des promoteurs de croissance s'accroissent dans les bourgeons, corrobore ces conclusions. La cessation de l'état de dormance n'entraîne pas toujours une diminution des inhibiteurs (Wareing, 1965). Il est prouvé que les inhibiteurs (par exemple l'acide abscisique) et les promoteurs de croissance agissent les uns sur les autres. C'est ainsi, par exemple, que l'acide gibberellique contrarie les effets des inhibiteurs (Thomas, Wareing et Robinson, 1965).
Les variations saisonnières constatées dans les types et doses de régulateurs hormonaux présents chez les arbres cadrent assez bien avec une théorie de la croissance fondée sur la modification de l'équilibre des promoteurs et des inhibiteurs. Alors que les bourgeons dormants contiennent; peu de promoteurs de croissance, les bourgeons renflés ou en éclosion en comptent de grandes quantités. Promoteurs et inhibiteurs de croissance ont été trouvés dans des bourgeons de pin (Allen, 1960; Ogasawara, 1961a, 1961b; Giertych, 1960; Giertych et Forward, 1966). Les niveaux des promoteurs de croissance sont généralement plus élevés dans les bourgeons terminaux que dans les bourgeons latéraux. C'est dans la partie inférieure ou médiane du houppier de Pinus resinosa que Giertych et Forward (1966) ont trouvé les plus fortes proportions de promoteurs de croissance. Cependant, les niveaux des inhibiteurs étaient aussi plus élevés dans la partie inférieure ou médiane que dans la partie supérieure du houppier, et il semblerait que dans cette dernière, les promoteurs de croissance l'emportent sur les inhibiteurs.
Le présent document s'est longuement étendu sur la régulation hormonale de la croissance dans des cas où il y a dormance véritable du bourgeon. On a voulu passer en revue les connaissances actuelles et souligner l'influence des modifications de l'équilibre hormonal sur la croissance. La dormance pouvant toutefois être considérée comme un cas limite d'inactivité et la quiescence comme un état intermédiaire, de plus amples renseignements sont nécessaires sur les relations hormonales chez les pins tropicaux dont les bourgeons sont en état de quiescence plutôt que de dormance véritable pendant une grande partie de l'année. Jusqu'à présent, les recherches physiologiques et biochimiques sur le mécanisme interne de la croissance chez les pins tropicaux ont été très limitées. D'après les études physiologiques effectuées sur la croissance d'un certain nombre d'espèces à poussées périodiques, il apparaît toutefois que les contrôles hormonaux jouent un rôle important. On peut supposer raisonnablement que l'équilibre entre promoteurs et inhibiteurs hormonaux se modifie pendant la séquence poussée - repos - poussée des pins tropicaux. On est tenté de penser que, dans les cas de foxtailing, il y a prédominance constante d'un ou plusieurs promoteurs de croissance. Toutefois, de plus amples recherches s'imposent sur les divers aspects des modifications internes dans les régulateurs hormonaux de la croissance des pins tropicaux. Il faut, entre autres questions importantes, répondre aux suivantes:
1. Quelles différences y a-t-il dans les quantités et les types de promoteurs et d'inhibiteurs de croissance pendant les poussées successives des pins tropicaux?2. Quelles différences y a-t-il, au cours du temps, dans les quantités et les types de promoteurs et d'inhibiteurs chez les sujets présentant du foxtailing?
3. Quelles différences y a-t-il dans les quantités et types de promoteurs et inhibiteurs d'une espèce à poussées périodiques en zone tempérée pendant les phases d'assoupissement et de réveil, d'une part, et pendant la phase de dormance véritable, d'autre part?
4. En quoi le cycle annuel des modifications dans les régulateurs de croissance de cette espèce est-il différent en zone tempérée et sous les tropiques où la même espèce est introduite?
5. Quelles différences y a-t-il dans les quantités et les types de régulateurs hormonaux de croissance en divers points du houppier chez les pins tropicaux et chez les pins à poussées périodiques de la zone tempérée?
6. Quelles sont les combinaisons de facteurs mésologiques qui engendrent divers équilibres hormonaux internes liés à des allures de croissance prévisibles?
On dispose d'excellents moyens pour répondre à quelques-unes des questions ci-dessus et éclairer ainsi le mécanisme interne de la croissance des pins tropicaux. De pareilles données aideraient sans doute à produire des pins tropicaux à forme prévisible, grâce à l'emploi de matériaux génétiques appropriés et au contrôle du milieu.
ALLEN R.M. 1960. Changes in acid growth substances in terminal buds of longleaf pine saplings during the breaking of winter dormancy. Physiologia Plantarum, 13: 555-558.
ALLEN, R. M. & MCGREGOR, W. D. H. 1962. Seedling growth of three southern pine species under long and short days. Silvae Genet., 11: 43-45.
ALLEN, R. M. & SCARBROUGH N. 1969. Development of a year's height growth in longleaf pine saplings. USDA. Forest Research Paper SO-45.
BARRETT, W. H. G. & GOEFARI L. 1962. Descripción de dos nuevas variedades del "Pino del Caribe" (Pinus caribaea Mor.). Caribbean Forester, 23: 59-71.
CHALMERS, W. S. 1962. The breeding of pine (Pinus caribaea Mor.) and teak (Tectona grandis) in Trinidad - some early observations. Caribbean Forester, 23: 100-111.
EGGLER, W. A. 1961. Stem elongation and time of cone initiation in southern pines. For. Sci., 7: 149-158.
FIELDING, J. M. 1960. Branching and flowering characteristics of Monterey pine. Canberra, Forestry and Timber Bureau. Bulletin No 37.
FREEIZAILLAH BIN CHE YEOM. 1966. Some notes on Pinus caribaea Mor. grown in Malaya. Forestry Department of Malaya, Forest Research Institute. Research Pamphlet No 54.
GIERTYCH, M. M. 1964. Endogenous growth regulators in trees. Bot. Rev., 292-311.
GIERTYCH, M. M. & FORWARD, D. F. 1966. Growth regulator changes in relation to growth and development of Pinus resinosa Ait. Can. J. Bot., 44: 717-738.
KAWASE, M. 1961. Growth substances related to dormancy in Betula. Proc. Am. Soc. hort. Sci., 78: 532-544.
KOZLOWSKI T. T. 1964. Shoot growth in woody plants. Bot. Rev., 30: 335-392.
KOZLOWSKI T. T. 1969. Tree physiology and forest pests. J. For., 67: 118-123.
KRAMER, P. J. 1967. Some effect of various combinations of day and night temperatures and photoperiod on the height growth of loblolly pine seedlings. For. Sci., 3: 45-55.
KRAMER, P. J. & KOZLOWSKI, T. T. 1960. Physiology of trees. New York, McGraw-Hill.
KUMMEROW, J. 1962. Uber Wachstumsanomalien bei Pinus radiata unter Tropischen Bedingungen. Ber. Dtsch. Bot. Gesell., 75: 37-40.
LANGE, P. W. 1966. The occurrence and development of abnormal long leaders in Pinus radiata D. Don in the Cape Midlands. Forestry in South Africa, 7: 1-10.
LANNER, R. M. 1964. Modifications in the growth habits of exotic trees in Hawaii. Proc. Soc. Am. For., 36-37.
LANNER, R. M. 1966. The phenology and growth habit of pines in Hawaii. U.S. Forest Service Paper PSW-29.
LLOYD, F. E. 1914. Morphological instability, especially in Pinus radiata. Bot. Gaz., 57: 314-319.
LÜCKHOFF, H. A. 1964. The natural distribution, growth and botanical variation of Pinus caribaea Mor. and its cultivation in South Africa. Ann. Univ. Stellenbosch, 39: 1-160.
McWILLIAM, J. R. & RICHARDS, B. N. 1955. Caribbean pine in Queensland, some notes on its behavior and potentialities. Queensland Forest Service Note No 4.
OGASAWARA, R. 1961a . Studies on auxins and inhibitors in Pinus thunbergii. J. Jap. for Soc., 43: 50-54.
OGASAWARA, R. 1961b. Studies on auxins and inhibitors in the buds of Pinus strobus. J. Jap. for Soc., 43: 307-310.
ROMBERGER, J. A. 1963. Meristems, growth, and development in woody plants. U.S. Department of Agriculture, Forest Service. Technical Bulletin N° 1293.
SLEE, M. U. & NIKLES, D. G. 1968. Variability of Pinus caribaea (Mor.) in young Queensland plantations. Proc. ninth Commonwealth for Conf., 1-50.
SMITH, H. & KEFFORD, N. P. 1964. The chemical regulation of the dormancy phases of bud development. Am. J. Bot., 51: 1002-1012.
STREETS, R. J. 1962. Exotic forest trees in the British Commonwealth. Oxford, Clarendon Press.
TEPPER, H. B. 1963. Leader growth of young pitch and short leaf pines. For. Sci., 9: 344-353.
THOMAS, T. H., WAREING, P. F. & ROBINSON, P. M. 1965. Action of the sycamore "dormin" as a gibberellin antagonist. Nature, Lond., 205: 1270-1272.
WAKELEY, P. C. & MARRERO, J. 1958. Five-year intercept as site index in southern pine plantations. J. For., 56: 332-336.
WAREING, P. F. 1965. Dormancy in plants. Science Progress, 53: 529-537.
|
CANADIAN JOURNAL OF FOREST RESEARCH En 1971, le Conseil national de la recherche du Canada commencera la publication d'une nouvelle revue internationale, le Canadian Journal of Forest Research. Elle présentera, en anglais ou en français, des rapports de travaux originaux de recherche scientifique, sous forme d'articles ou de notes, accompagnés d'observations pertinentes. Tous les aspects de la science forestière seront examinés et les communications intéressant des disciplines connexes seront particulièrement bien accueillies. (Jette revue paraîtra trimestriellement à partir du mois de mars. Les manuscrits doivent être adressés à l'éditeur, Dr. J.L. Farrar, Faculty of Forestry, University of Toronto, Toronto 5, Ontario. Abonnement annuel (dollars, canadiens): abonnement de groupe: $10,00; abonnement individuel: $5,00. Les demandes d'abonnement doivent être adressées comme suit: Administration, National Research Council of Canada, Ottawa 7, Canada, les mandats ou chèques étant libellés à l'ordre du Receveur général du Canada. |
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}