![]()
![]()
![]()
In developing criteria, indicators, and verifiers, we emphasize that the conservation of evolutionary processes is a necessary biological criterion by which the sustainability of forests can be judged and we use genetic indicators of those processes to judge the sustainability of those evolutionary processes. To conserve these evolutionary processes, we define four genetic indicators that are necessary for sustainability. For each indicator, we list several ways to measure threats to the processes. These options conform with the “tool kit approach”, described above, which offers a forest manager a choice of verifying devices. Two types of verifiers are described, those which can be obtained by field assessment of demographic or morphological parameters and those which require laboratory analysis of genetic parameters. Both types contain several verifiers, and these vary in their precision and ease of use.
The proposed criterion for the conservation of genetic variation is:
Different types of forest use are likely to affect several of the processes that maintain genetic variation as indicated in Table 1. The processes of genetic change have effects that can be measured in the level and structure of genetic variation and in the processes themselves. These measures can serve as indicators of whether genetic variation and the processes of evolution are being maintained. We propose 4 indicators of the genetic processes:
1. Levels of genetic variation
2. Directional change in gene or genotypic frequencies
3. Gene migration between populations
4. Reproductive processes/mating system
As shown in Table 2, one or more indicators need to be assessed for each of the genetic processes.
TABLE 2. Relationship between genetic processes and indicators
|
Processes
|
Indicators |
|||
|
Levels of genetic variation are maintained |
There is no directional change in genic/genotypic frequencies |
There are no changes in gene flow/migration |
There are no changes in the mating system |
|
|
Drift |
X |
|
|
|
|
Direct selection |
X |
X |
|
|
|
Indirect selection |
X |
X |
|
|
|
Migration |
|
|
X |
X |
|
Mating |
|
|
|
X |
To be most useful to forest managers, we can combine the effects of forest level events on genetic processes and the indicators of those processes, as given in Tables 1 and 2, to provide managers with tools to warn of possible changes in genetic processes which arise from particular management practices or by accident. This is shown in Table 3, where indicators are directly linked to forest practices and other forest level events. Table 3 allows forest managers to determine which genetic indicators need to be assessed based on their knowledge of forest practices in the FMU. Thus, the need for genetic expertise is minimized in the assessment of sustainability of genetic variation.
For example, if harvesting of wood is taking place, the following indicators need to be assessed:
Overall levels of genetic variation
Possible directional change in genic/genotypic frequencies
Possible changes in reproductive processes/mating system.
TABLE 3. Indicators relevant to some forest operations
|
Modes of harvesting or use |
Indicators |
|||
|
Levels of genetic variation are maintained |
There is no directional change in genic/genotypic frequencies |
There are no changes in gene flow/migration |
There are no changes in the reproductive processes/mating system |
|
|
Logging - commercial species |
X |
X |
|
X |
|
Logging/wood harvesting of non-commercial species |
X |
X |
|
X |
|
Grazing |
X |
|
|
|
|
Fire |
X |
|
X |
X |
|
NTFP - reproductive |
X |
X |
X |
X |
|
NTFP - non-reproductive |
X |
X |
|
|
|
NTFP - whole individual |
X |
X |
|
|
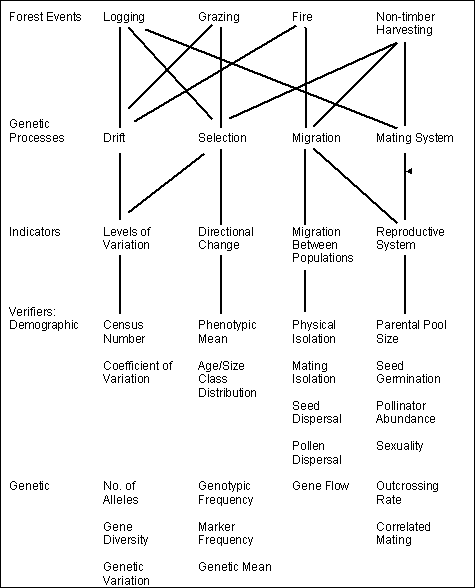
The reason for this, as shown in Figure 2, is that selective harvesting of wood primarily affects the genetic processes of drift and mating, and the indicators listed above are indicators of those processes. Using Table 3, the forest manager does not necessarily need to understand the genetic theory behind this.
For each of the indicators, the verifiers may be parameters of the genetic system that directly reflect genetic processes or other factors of the reproductive system. For example, genetic parameters such as gene frequencies can serve as measures of the level of genetic variation, and differences in frequencies can be used to estimate population differentiation. These data can also be analyzed to estimate parameters of the mating system, such as the outcrossing rate, selection effects, and migration rates. Alternatively, verifiers may be estimates of demographic statistics from which we can infer genetic processes. For example, changes in population size and age class structure may reflect changes in the levels and distribution of genetic variation as well as the mating system. The distinction between genetic and demographic verifiers reflect the point made by Brown et al. (1997) that the choice of verifier will always be a trade-off between high information content (in our case, generally genetic verifiers) and ease of measurement (in our case, generally demographic verifiers).
Thus the different verifiers that can be measured and used for any one indicator differ widely in the precision with which they estimate the genetic parameters, and in the costs and facilities required to sample and measure them. We list several verifiers for each indicator, divided into two groups. The first group of verifiers uses demographic information and is intended to serve as the first step in assessing risk of local extinction - many of the demographic verifiers can be estimated from standard forest inventory information. If additional precision is desirable, then at least one of the second set can be used. Within each group, the manager is expected to use the most informative verifier that can be feasibly estimated. The more costly and precise measures would generally be reserved for use only on the most critically threatened processes where decisions require high precision. For each indicator, verifiers within each group (demographic and genetic) are listed in the approximate order of precision with the least precise but easiest verifiers to measure, last.
Figure 2. Relationship between Forest Events and Genetic Processes and Their Indicators
The risk we seek to assess is that of local species extinction as a response to previous or proposed forest practices, and critical parameter levels are given that are expected to provide a reasonable margin of safety against that risk. The parameter levels for the two types of verifiers are expressed as deviations from some reference populations. Reference populations will ideally be areas of forest ecologically similar to the test site, where there have been no management interventions. Such conditions will often be difficult to find, so in practice the reference populations will often be forest areas which have been subject to interventions, but sufficiently long ago and/or of sufficiently benign influence that the genetic processes of the reference population can be considered to be uncompromised.
Since variation exists at many gene loci and these loci are partially independent in their evolutionary behaviour, the use of multiple verifiers can provide more complete information about the genetic processes than can single verifiers. For example mating system parameters are best measured by verifiers that are not subject to selection on vegetative vigor such as isozymes or RFLP’s, whereas parameters of adaptability to environmental conditions are best measured by verifiers of traits such as growth or phenology that may be heritable. Thus, multiple genetic verifiers are preferred over single verifiers for complex events. Physiological traits would be particularly useful as estimators of adaptability to general environmental conditions as well as to stress conditions. However, to demonstrate their heritability, existing field trials must be evaluated; and, in the absence of such trials, they must be established. This is expensive and time consuming.
The development of accurate, short-cut procedures would be very valuable. Many random DNA markers based on PCR techniques such as RAPD’s, AFLP’s, and SSR’s (see Annex 4 for definitions of these and other technical terms) can provide a multitude of data on non-selective processes since they mostly have no physiological effect. Some markers may be associated with physiological effects but finding these markers requires special experimental methods such as used in QTL analyses. Random markers are not likely to show associations in large, predominantly out-crossed species and may be useful only if very large numbers of markers are well dispersed throughout the genome (Strauss et al. 1992, FAO 2001). Some markers, such as isozymes and those derived from c-DNA probes have functional meaning. However, currently available molecular methods are either too difficult to use in most field conditions, or require such large initial development time, that they would not often be useful in the present context. More research is needed to develop procedures, facilities, and training, before the molecular methods can be useful in all but the most critical and valuable cases. Ideally, a broad mixture of phenotypic, field measures, and multiple locus molecular markers that would sample the whole genome would be available to provide direct genetic process information.
The above indicators consider the genetic processes as if they were independent of the ecosystem on which they have an impact and which impacts them. The effects of genetic management on important target species and the ecosystems in which they occur must be considered. Forest species have evolved different natural histories and are not equally easy or valuable to sustain in the future. Indicators of the capacity of species for genetic management are therefore needed to judge the likelihood that recommended actions could successfully sustain the genetic resource. A genetic “triage” (Koshy et al. 2001) can be useful in this context to assess the potential for restoration and future sustainable management. The genetic “triage” builds on long-established principles of medical triage, through which species are assigned to one of three possible groups - those for which conservation is secure, those for which local extinction is inevitable, and those for which modifications to management interventions can ensure conservation where otherwise local extinction would occur. It is only this third group which is of immediate concern to the manager.
Levels of variation within a population result from a balance among all the genetic processes that have previously been outlined (Table 1). Effective population size (Ne) is generally recognized as an indicator of the changes that alter the level of variation within a population. However Ne is extremely difficult to assess in aged structured and heterogeneous populations, whereas measurement of population size is more realistic and its estimate, in addition to the assessment of actual level of variation, may have equal prediction for the maintenance of genetic variation. Indicator 1 therefore combines two different concepts, population size and levels of variation, for which demographic and genetic verifiers are proposed. Comparisons of the verifiers between the target population and the reference population should be made on comparable and non-impacted cohorts, distributed on areas of similar sizes.
Demographic verifiers
1.D.1 Census number of sexually mature individuals
This number accounts for all individuals expected to be sexually mature at present time, according to their age or size. It also includes those individuals that will reach maturity before the next human intervention in the forest.
1.D.2 Census number of reproducing individuals
If fecundity and fertility data can be obtained (based on inventory data in surrogate populations or in the study population), then this number corresponds to the subset of 1D1 that is expected to contribute to the next generation. This verifier is easier to apply to plant than to animal species.
1.D.3 Coefficient of phenotypic variation
Assessments of in situ phenotypic variation of important adaptive traits will provide some crude indication of genetic variation of the same traits. Adaptive traits include such traits as for example, timing of flowering, sex ratio in dioecious species, colour of resin (which reflects chemical variation in the resin). An assessment of the variability in such traits within a population can provide information of genetic variation. For example, assuming low heritability for these traits (h2 =0.04), as may be the case in extremely heterogeneous environments, a coefficient of phenotypic variation of 12% would indicate that the genetic coefficient of variation is about 2.4%.
Genetic verifiers
The genetic verifiers that are proposed correspond to genetic parameters commonly used in population or quantitative genetics, and for which confidence intervals have been developed. They are ranked according to the resources needed for their assessments. Their applicability depends on the molecular or biochemical techniques available for the target species.
1.G.1 Number of alleles
The structural and functional unit of inheritance being transmitted during the reproductive process is the gene. Genes influence hereditary traits among the offspring. These genes can exist in different forms or states, known as alleles. Allelic richness is strongly affected by changes in population sizes, and could be assessed on a reduced number of loci that are polymorphic in the reference population. Rapid estimates of the number of alleles could be obtained by bulking enzyme or DNA extracts of several samples for multi-allelic and co-dominant markers (isozymes, microsatellites). We would recommend that the target population comprises all alleles present in the reference population at a frequency higher than 0.05. The sample size necessary to detect, with a probability of 95%, at least one copy of an allele present with a frequency of 0.05, is 120 alleles (60 genotypes). Assessments should be made on at least 5 loci which have been shown to be polymorphic in the reference population.
1.G.2 Gene diversity
If multi-allelic and co-dominant markers are not available, then levels of variation could be assessed at anonymous and randomly distributed markers (RAPDs or AFLPs) by using measures that account for allele frequencies (Nei’s gene diversity or expected heterozygosity, H). Phenotypic frequencies (corresponding to the presence/absence of a fragment for dominant markers) could be used for calculating these parameters. To be realistic, we propose to assess H with the same sampled size as allelic richness (60 genotypes). This will represent a 95% confidence interval, varying between 0.07 and 0.10 (depending if the species is preferentially outcrossing or not). Comparisons between the reference and target populations should be based on at least 10 loci.
1.G.3 Genetic variation
Additive variance of important adaptive traits could be estimated in common garden (field) experiments. Narrow sense heritability values or coefficient of genotypic or additive variance, could be used as verifiers for levels of genetic variation. We would recommend a sampling strategy that permits comparison of genetic variation between the target and the reference populations for traits exhibiting heritability values even as low as 0.04. This could be achieved by collecting 50 open pollinated progenies, each represented by 50 seedlings.
Critical levels
If census numbers in the target population are above critical absolute values (1.D.1 > 50 or 1.D.2 > 30) and the coefficient of phenotypic variation in the target population is higher or not significantly different from the reference population, then Indicator (1) can be considered as indicating sustainability. Decisions based on demographic verifiers rely therefore on two parameters: the census number and the level of phenotypic variation. However, if either one of these is not known, additional assessments with genetic verifiers have to be made before reaching a conclusion.
If demographic verifiers fail to indicate sustainability for Indicator (1), genetic verifiers need to be assessed. The genetic verifiers proposed are largely complementary. This means that it is not necessary to assess all such verifiers - it is sufficient to select one that fits the resources available and techniques mastered, though assessment of more than one verifier may provide additional useful data. If the selected verifier in the target population is no more than 25%, or 1 standard error, lower than the corresponding value in the reference population, then this indicator is acceptable.
Natural or human induced selection is the main force that will change gene and genotypic frequencies in a directed fashion. They operate on phenotypic traits in individuals, and directionally affect genotypic frequencies to the extent that those traits are heritable. For example, selective removal of trees with good stem form may leave a residual population with changed genotypic frequencies, leading to a form of genetic high-grading. In systems using selective harvesting or involving breeding, conscious selection may be assumed, but in many cases, indirect selection effects may also change gene and genotypic frequencies. These may be due to the complex processes such as logging which may selectively remove understorey individuals of a particular size or form, or they may be due to complexities of the genetic system where genetic changes in a given trait, such as size, may lead to changes in a correlated trait, such as age of reproduction. Directional change may also be due to non-selective causes: a history of divergent selection may result in differences of adaptive significance.
Demographic verifiers
2.D.1 Phenotypic Shifts.
If phenotypic mean differences between a reference population and the affected population are evident, then a genotypic change may have occurred that may cause a change in the economic or ecological sustainability of the population. These differences would be expected to be most strongly evident in traits with high heritability and those under direct selection pressure. Other traits that may be indirectly affected but which at the same time are of potential significance for future adaptations, may be less obviously affected, but should also be observed and monitored. They - can best be measured in common garden (field) experiments, but inferences can be drawn also on the basis of paired stands or cohorts. Only very large differences would be expected to significantly affect allele frequencies and hence differences of greater than 2 standard errors from the reference population mean would be of any concern.
2.D.2. Age/Size Class Shifts.
If differences occur in the age or size class distribution exist between a reference population and the affected population, genic level changes may be caused by age related selection. Under conditions of minimal disturbance or in steady state environments, an age class distribution which is in equilibrium may be attained and its disturbance is equivalent to an environmental disturbance. Differences from a reference cohort, or the absence of some age classes, may portend directional selection. In non-equilibrium populations, differences in age classes are expected and may not imply threats, but any large change is still worth noting and monitoring. As in the case of verifier 2.D.1, only very large differences would affect allele frequencies, and hence only statistically highly significant differences in mean values should be considered, unless simultaneous changes in other parameters, such as population size, are noted.
2.D.3. Environmental Shifts.
If differences in the environment that affect either viability or reproduction are caused by forest level events, then selection may be so strong as to affect population viability. For example, shade requiring species may be exposed to full sunlight or species requiring full light may be exposed only to shade, and neither may survive without strong frequency shifts. If survival or reproduction is less than 50% of that estimated for a reference population, then selective differences can be significant.
Genetic verifiers
2.G.1. Genotypic Frequency Shifts.
Differences in genotypic frequency distributions among cohorts may be measured as changes in heterozygosity, for example in cases where inbred individuals are selectively eliminated. However, generational changes can seldom be estimated in the management framework and hence, changes would have to be observed indirectly, among cohort populations. A simple assessment can be made on the basis of heritability estimates made on surrogate or reference populations, coupled with estimates of the selection differential resulting from the forest intervention.
2.G.2 Marker Frequency Shifts.
In some cases, selection will result in directional changes in genotypic frequencies at some marker loci among generations or from a reference population. This is the case when there is close linkage and stochastic association between the marker and the selected loci. Such effects may be estimated using as few as a dozen loci with more than 30 sample individuals per population. However, the lack of detectable changes in gene frequencies does not prove an absence of effects of selection. These changes are the most technically challenging ones to detect, and they provide only weak tests of selection; however, their value lies in the fact that they are direct measures of directional genetic change.
2.G.3 Genetic Mean Shifts.
The most direct assessment, which at the same time is the most costly one in terms of time and effort, is the establishment of a genetic estimation experiment aimed at analyzing differences in traits of adaptive significance. The establishment of such trials would generally however require more time than is available before management decisions have to be made.
Critical levels
Differences in the genetic verifiers between the reference or other comparison population and the affected population must be fairly marked for concern to be raised. If the difference is less than a 25%, or if observed changes are less than 1.5 standard errors, it is usually considered that there is little cause for concern regarding sustainability.
Migration is a strong force that reduces genetic differences among populations, but which may increase variation within populations. Migration among populations which may be adapted to different environments may cause a reduction in fitness in the receiving populations. However, if local populations have earlier been greatly reduced in size, then migration, especially through seed, can increase the mating pool and lead to increased fitness. Within any one zone in which the environment is similar, the exchange of migrants among meta-populations will generally not reduce the fitness of receiving populations. The demographic verifiers listed below are of significance to sustainability in those cases in which isolation barriers are broken between populations with different adaptations. The impact will be greatest when the adaptive differential is large and the resident population sizes are small.
Demographic verifiers
3.D.1. Physical Isolation.
Reductions in degree of physical isolation by loss of neighbors or by easing of physical or biotic barriers to pollen or seed migration.
3.D.2. Mating Isolation.
Breaking mating isolation by changes in age class distribution and by loss of neighbors that may increase the frequency of mating among distant parents.
3.D.3. Seed Dispersal.
Changes in seed dispersal distances due to physical or biotic factors affecting seed dispersal vectors. (This verifier can only be used when the seed vector is known).
3.D.4. Pollen Dispersal.
For plant species, the effective distance of pollen dispersal may be increased by factors which lead to increased pollen vector influences, caused either by changes in the species of pollen vector, or in variation in their inter-individual distances. (This verifier can only be used when the pollen vector is known).
Genetic verifiers
3.G.1 Gene Flow.
Gene flow can best be estimated by tracing neutral marker loci and estimating statistics[2] such as Fst, Gst, d, Dj, and theta statistics. The use of cytoplasmic gene markers can discriminate between male and female migrants. Low levels of differentiation suggest a large gene flow among populations. The allocation of samples within and among populations should include at least 30 individuals within populations and should emphasize between population sampling when estimation of population differences require precise estimation. The reference populations to which such statistics refer would also have to represent a range of populations.
Critical levels
Changes in the verifiers of less than 50% in the demographic verifiers, or of less than 10% in the genetic parameters, would indicate sustainability in regard to this indicator.
The reproductive processes is the bridge between generations and hence mediates evolution. The mating system involves the levels of fecundity and fertility, propensities for inbreeding versus outbreeding, and unequal or non-random mating associations. It is the primary determinant of how genetic variation is partitioned among individuals (levels of heterozygosity) and hence, of how much inbreeding depression will be expressed in the next generation. The mating process is affected by sex ratio, phenology, and specific compatibility relationships between potential mates; and is subject to selection. Measures of pollen flight and numbers of reproductive organs have proven to be difficult to obtain in the field and have failed to accurately predict the genotypic structure of the resulting seedling pool.
Demographic verifiers
4.D.1. Parental pool size
Modification of the density of potential parents indicates possible changes in future mating success. The threshold for this indicator will depend on the distance of pollen dispersal. The density will have to be assessed on sexually mature individuals. Modification of flowering phenology within the population may also portend changes in the potential parental pool.
4.D.2. Seed Germination
Modification of the percentage of seed germination (or of filled seed count) relative to the reference population, can reflect changes in mating success. It does not give information on the number nor the distribution of the mates. The percentage of germination could be assessed for 10 individuals (4 replicates of 50 seeds per tree; seeds should not be taken from a single branch on any tree) in standard conditions; and the necessary pre-treatment used if it is known. Low percentages may be signs of increasing selfing, leading to inbreeding depression. Large variations between individuals will characterise unbalanced contributions to reproduction from the different maternal parents.
4.D.3. Pollinator Abundance
Modification of pollinator abundance or composition during the mating season indicates potential changes in the male gamete pool. The procedure for estimating this factor will be different according to the types of pollinators. If the tree species is bird or bat pollinated, an animal population census should be carried out; if this is not possible then environmental modifications that can affect population size of the pollinator should be evaluated. If the species is insect pollinated, flower visitors should be captured and the pollen they carry should be identify to include only true pollinators in the estimate (pollinators should be captured from 5 individuals; if the composition is very variable, more individuals will have to be included). Comparison will have to be carried out relative to a reference population.
4.D.4. Ratio between Male and Female Flowers
Modification of the ratio of male and female functions portends changes in mating success. Particular attention should be paid to dioecous species, for which sex ratio modifications will reduce the potential mating pool. The sex ratio should be assessed on at least 30 sexually mature individuals and compared with that of a reference population.
Genetic verifiers
4.G.1. Outcrossing rate
Changes in the outcrossing rate will point to changes in potential inbreeding, and possible increases in inbreeding depression. The multilocus outcrossing rate can be estimated from open-pollinated families; seeds should be collected on 20 individuals in the population, 10 progeny per tree have to be assessed for at least 4 polymorphic loci (isozymes could be used or microsatellites). The value obtained for the outcrossing rate will have to be compared with estimations from “undisturbed” reference populations to estimate how significantly it may affect inbreeding.
4.G.2. Correlated mating
Even if the outcrossing rate is not changed, modification of gene flow within a given population can strongly affect the genetic makeup of the next generation. Comparison of multi-locus outcrossing rates with the average of single locus estimates will give some indication of the frequency of mating among relatives. Furthermore, based on this, one can estimate how many fathers have sired a given progeny. This should be sampled on at least 30 parents and for at least 6 loci with microsatellites or with 20 loci for isozymes.
Critical levels
Changes in these verifiers which are greater than 50% for the demographic verifiers, and greater than 10% for the genetic verifiers, would be considered to be critical.
|
[2] See annex 4 for glossary of
terms. |
![]()
![]()
![]()