![]()
![]()
![]()
by
D.A. HUGHES
Institute of Marine Sciences
University of Miami
Miami, Florida 33149, U.S.A.
Abstract
The activity of pink shrimp shows a clear phase relationship with the day-night cycle. This study examines the mechanisms whereby the synchrony of emergence from the sand at the time of sunset is maintained between all members of the population.
The results indicate that emergence is under the influence of a circadian rhythm in phase with the light-dark cycle. A periodically changing responsiveness to a light-dark transition indicated the importance of the latter as a ‘Zeitgeber’, maintaining synchrony between the circadian rhythm and the light-dark cycle.
A 24 h feeding rhythm was indicated by experiments in which shrimp, induced to emerge to feed during the day, would again emerge and carry out food searching movements at approximately the same time the following day when no food was present.
The adaptive significance of these rhythms and their effect on the synchrony of emergence is discussed.
FACTEURS DETERMINANT LA SORTIE DE LA CREVETTE Penaeus duorarum
Résumé
L'activité de Penaeus duorarum se révèle nettement liée au rythme nycthéméral. L'auteur étudie les mécanismes par l'action desquels tous les individus de la population sortent du sable au moment où le soleil se couche.
D'après les résultats, la sortie de l'animal est sous l'influence d'un rythme quasi journalier correspondant à la variation nycthémérale. La modification périodique de la réponse au passage de la lumière a l'obscurité met en relief l'importance de ce dernier phénomène, qui joue le rôle de “Zeitgeber”, assurant le synchronisme entre le rythme quasi journalier et le cycle nycthéméral.
Un cycle alimentaire de 24 heures a été mis en évidence par des expériences durant lesquelles les crevettes, qui avaient été incitées à sortir pour se nourrir durant la journée, réapparaissaient à peu près à la même heure le jour suivant et se mettaient en quête de nourriture même en l'absence de celle-ci.
L'auteur examine l'importance que présentent ces rythmes du point de vue de l'adaptation et leur incidence sur le synchronisme de la sortie.
FACTORES QUE REGULAN LA APARICION DEL CAMARON ROSADO Penaeus duorarum
Extracto
La actividad del camarón rosado muestra una clara relación de fase con el ciclo día-noche. En este estudio se examinan los mecanismos por los que se mantiene entre todos los miembros de la población la sincronía de la salida desde la arena en el momento del crepúsculo.
Los resultados indican que tal salida se halla bajo la influencia de un ritmo cuasi-diario en la fase con el ciclo luz-oscuridad. Una respuesta periódicamente cambiante a la transición luz-oscuridad indicó la importancia de la última como “Zeitgeber”, manteniendo la sincronía entre el ritmo cuasi-diario y el ciclo luz-oscuridad.
Los experimentos realizados con camarones, a los que se inducía a salir para alimentarse durante el día, indicaron que el ritmo de alimentación era de 24 horas; éstos volvían a salir y se desplazaban en busca de alimentos a la misma hora aproximadamente del día siguiente, cuando carecían de ellos.
Se examina la importancia de la adaptación de estos ritmos y sus consecuencias sobre la sincronía de la salida.
Diurnal patterns of activity occur in most species of shrimp. Most species are recorded as active above sand at night and buried by day, and in this category are Metapenaeus bennettae Racek and Dall (as M. mastersii (Haswell)), M. monoceros (Fabricius Penaeus aztecus Ives, P. japonicus Bate, P. monodon Fabricius and P. semisulcatus de Haan (Idyll, 1950; Dall, 1958; Egusa and Yamamoto, 1961). On the other hand, P. setiferus (Linnaeus) is said to be active by day and is thought to burrow at night (Idyll, 1950). Racek (1959) records that the Australian shrimp P. plebejus Hess and Metapenaeus macleayi (Haswell) are only active just before and just after sunrise. It was found that P. indicus H. Milne Edwards never burrowed by day or night (personal observations).
The nocturnal activity of P. duorarum Burkenroad, has been recorded by Idyll (1950), Iversen and Idyll (1959), Williams (1955, 1958), Viosca (1957), Eldred (1958), Eldred et al. (1961), Fuss (1964), Fuss and Ogren (1966) and Wickham (1966). Most of these records are, however, the results of incidental observations, and little is known of the nature of the control of this periodicity in behavior.
Fishermen have long been aware that overcast weather or murky waters can increase their catches during the day. Conversely, an increase of light intensity at night may cause reburrowing of shrimp which are already above sand (Eldred et al., 1961). Fuss and Ogren (1966) suggested that “the daily activity patterns of pink shrimp seem to be primarily controlled by photoperiod and other modifying environmental factors but there is some evidence of an endogenous component”. The presence of the latter was clearly shown by Wickham (1966) who found that a rhythm controlling the times of activity would persist for longer than a week, under apparently uniform external conditions.
This study concentrates on one aspect of the diurnal activity pattern, the initial emergence from the sand at dusk. This activity is markedly synchronised in all members of the population. An attempt is made here to determine the factors eliciting emergence and maintaining its synchrony in all individuals.
The shrimp used in the experiments were all juveniles (subadults) and were housed in twelve 60 1 aquarium tanks with a sand substrate 6 to 8 cm deep. The water, maintained at a depth of approximately 20 cm was in constant circulation and aerated through subsand filters.
The tanks were enclosed in a light-proof enclosure within a laboratory. The shrimp were exposed either to natural day-night cycles, by uncovering the window against which the tanks were placed, or artificial cycles of light and dark, by the use of a time switch connected to a 150 W flood lamp. The flood light was diffused by deflecting it off the white roof of the enclosure.
The shrimp were netted from a canal in the Everglades of southern Florida. In each tank 6 to 10 shrimp (3 to 12 cm total length) were housed and fed on small pieces of fish (mullet). Virtually no mortality occurred and, unless the water became polluted or the circulation and aeration mechanisms failed, the shrimp appeared healthy.
3.1 Diurnal pattern of burrowing behavior
The activity of this species indicates a clear phase relationship with the day-night cycle. Fig. la shows the typical pattern of burrowing behavior of 50 shrimp recorded on three consecutive days when exposed to a natural day-night cycle. The shrimp are active above the substrate at night but are inactive and buried during the day. The close association between sunset and the onset of activity (in this case emergence) confirms the records of Fuss and Ogren (1966) and Wickham (1966). These workers also reported a two peaked pattern of occurrence above sand at night, similar to that shown in the figure.
The persistence and circadian periodicity of this pattern under conditions of low light intensity (Wickham, 1966) is confirmed by the results expressed in Fig. 1b. The pattern is shown here for only three consecutive days, but, as shown by Wickham (1966), the evening “peak” of emergence was still clearly noticeable after one week and the free running period throughout this time was clearly longer than 24 h.
These results suggest that a circadian rhythm in phase with the light-dark cycle is responsible for the observed pattern of occurrence above sand (Fig. 1a).
When the light-dark transition was advanced or retarded by 1 h, the time of emergence closely followed the transition, suggesting that the latter is the factor which maintains synchrony between the circadian rhythm and the light-dark cycle (Zeitgeber). If this is the case, a “periodically changing sensitivity of the organism to the stimuli of the Zeitgeber” (Aschoff, 1965) should be apparent. To determine this, emergence was recorded after imposing light-dark transitions at 09:00, 12:00, 15:00 and 18:00 h. Prior to each experiment, all shrimp (n = 75–100) were maintained under a 12 h - 12 h light-dark cycle for at least 10 days. This cycle coincided closely with the natural cycle. The time of the light-dark transition was 18:00 h and was abruptly imposed.
Patterns of emergence, compared here and in later experiments, were markedly consistent for different groups of shrimp maintained under identical light-dark cycles and temperatures, and similarly fed. These patterns can therefore be used as reliable indices of the influence of all stimuli effecting emergence under a particular set of conditions.
Emergence, as a response to a light-dark transition imposed at 18:00 h was relatively rapid (Fig. 2). At a temperature of 25°C, 90 percent or more emerged within the first 30 min of the light-dark transition. Following emergence, the shrimp carried out swimming and food searching movements; they seldom reburrowed until several hours later, at the time of the quiescent period between the peaks of activity.
If the light-dark transition was imposed at a time other than at 18:00 (i.e. at 09:00, 12:00 or 15:00 h) emergence occurred more slowly. Seldom more than 50 percent emerged within 90 min. Those that did were seldom active and a large proportion would reburrow within an hour.
This is clear evidence for a changing responsiveness to the light-dark transitions and implies that the light-dark transition is the Zeitgeber, or an important component of it, which maintains the synchrony between the internal oscillation of the rhythm and the external light-dark cycle.
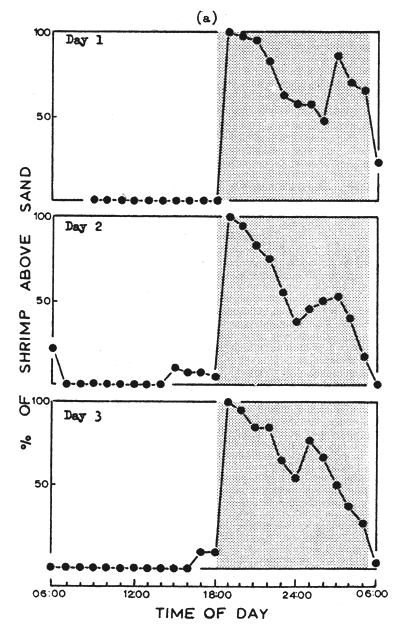 | 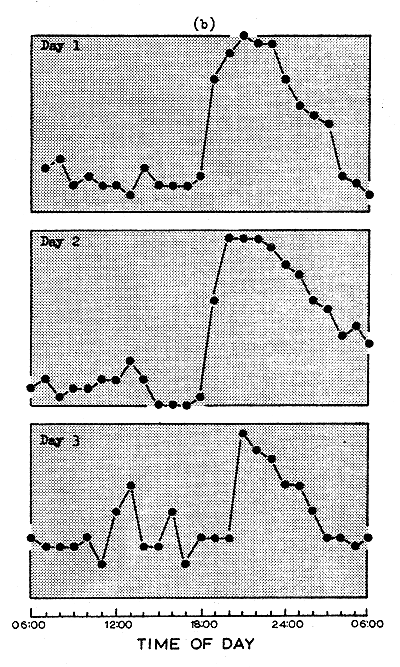 |
Fig. 1 The burrowing of shrimp during three consecutive days when exposed to (a) a natural day-night cycle and (b) to constant low intensity illumination. Ten shrimp were housed in each of five tanks: counts of the number of shrimp above sand were taken at hourly intervals.
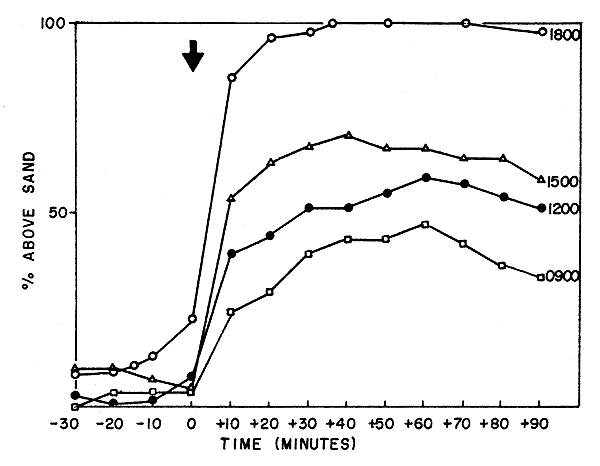
Fig. 2 The pattern of emergence of shrimp when exposed to light-dark transitions imposed at 09:00, 12:00, 15:00 and 18:00 h. The shrimp had been entrained to a LD 12:12 cycle in which the light-dark transition occurred at 18:00 h.
3.2 Phase-shifting of the light-dark cycle
To elucidate further the nature of the factors responsible for the phase synchronization of the rhythm with the environment, experiments involving shifts of the phase of the light-dark cycle were conducted.
The phase of the rhythm should be readily resynchronized to a shifted Zeitgeber, and the response of the rhythm to such a shift has been commonly employed to indicate the phase dependence of the rhythm on a particular environmental factor. (For a review of the literature on entrainment see Aschoff, 1963). Aschoff (1960) stated that “if the Zeitgeber is fully effective the organism should follow the shift within a few periods and should regain the original phase relation to the Zeitgeber”.
In the following experiments, the phase of the light-dark cycle was altered through an angle of 120°, so that the light-dark transition occurred at 10:00 h instead of 18:00 h. A 12 h - 12 h light-dark cycle was again imposed after the phase-shift, and the pattern of emergence at the time of the light-dark transition was recorded for several days under the altered regime. The behavior of two groups of 40 shrimp, all of similar size, was compared. Group A were fed each day after the records of emergence were completed (at approximately 11:30 h). Group B were starved after the cyle had been shifted. Daily records were continued until emergence was once more maximal (as at 18:00 h prior to the phase-shift) and until subsequent activity occurred without reburrowing. At this time, the circadian rhythm was considered to have returned to its prior phase relationship with the shifted Zeitgeber.
Emergence of group A after exposure to only one cycle of the altered light regime was almost maximal. The equivalent pattern of emergence was only achieved by group B, following a series of transient stages, after four days exposure to the altered cycle. It appeared, however, that group A did not achieve complete resynchronization with the shifted light-dark cycle after only one day because, despite the almost maximal emergence, there still remained the tendency of some shrimp to reburrow after emergence. (This is presupposing that emergence and subsequent activity are both responses to the same oscillating system). By the third day, however, the pattern of emergence and subsequent activity of group A could not be distinguished from that of shrimp long exposed to a particular light-dark cycle. Group B, by comparison, required six days to reach an equivalent pattern.
3.3 The effects of feeding and food deprivation on emergence
Experiments were conducted to determine whether the slower resynchronization of the rhythm with the light-dark cycle which occurred in group B (unfed) compared with group A (fed) was due to starvation causing a consequent depression of activity.
The pattern of emergence of 40 shrimp which had not been fed for four days was compared with that of an equivalent group, maintained under identical conditions but fed each day after the onset of the dark period. For this experiment, emergence was again recorded when a light-dark transition was imposed at 10:00 h on shrimp that had been entrained to emerge at 18:00 h.
In two out of three repetitions, the rate and degree of emergence of the unfed shrimp was greater than that of the fed shrimp. On the third occasion, there was no noticeable difference between the two groups. These results suggest that the difference in the time taken to resynchronize the rhythms of the fed and the unfed shrimp, following phase shifting of the external light cycle, was not a consequence of starvation.
3.4 Evidence for a feeding rhythm
To separate emergence which could be the result of a feeding rhythm from normal nocturnal emergence and activity, the shrimp were fed at midday, and their emergence recorded the following day. (Under normal circumstances the shrimp do not emerge during the day, however, when not satiated, they can be induced to emerge if food is placed in the tanks).
Two groups (A and B), each comprising 25 shrimp, were used in this experiment. They were both held under identical conditions (LD 13½:10½) but were fed at different times.
On day 1, group A was fed at 12:30 h; group B was fed at 19:45 h.
On day 2, records of emergence were taken. Group A emerged in large numbers at approximately midday (24 h after they had been fed). Very few of group B emerged during the same period.
Group B was then fed at 13:40 h (and emerged to feed).
Group A was not fed until 19:45 h that evening.
On day 3, the results were similar to those of day 2 except that in this case, group B emerged at approximately 13:40 h while very few of group A emerged at that time.
The time of activity of this species indicates a clear phase-relationship with the day-night cycle. The most marked manifestation of this association is the close relation existing between the time of emergence from the sand and the time of the light-dark transition of the day-night cycle. Emergence of almost the entire population takes place within a period of 20 to 30 min at about the time of sunset, and, should the time of this light-dark transition be shifted in either direction by an hour, it is always followed by a shift in the mean time of emergence. The findings of this study elucidate the mechanisms whereby this synchrony of emergence between all members of the population is maintained.
Two rhythms are involved: (i) a circadian rhythm in phase with the light-dark cycle and (ii) a 24 h feeding rhythm. Experiments were designed to distinguish the influence of each and they are discussed separately below.
(i) Circadian rhythm of emergence from the sand
The shrimp showed a periodically changing responsiveness to light-dark transitions (Fig. 2). Although they would always, to a greater or lesser extent, emerge when a light-dark transition was imposed, their rate and degree of emergence was far greater at the time of the light-dark transition of a light-regime to which they had become entrained. The phase dependence of the rhythm on the light-dark cycle was shown by experiments in which the latter was shifted through 120°. Six days of entrainment to the altered cycle was required to resynchronize the rhythm in shrimp which were not fed following the shift of cycle, indicating that the basis of this rhythm is independent of the 24 h feeding rhythm. This is in accordance with Aschoff's (1963) findings which showed that in general, many organisms need three to six days for a 6 h phase shift, i.e. 1 to 2 h shift per day.
An indication of the influence of the rhythm is further shown by the fact that, after phase synchrony had been achieved in the above mentioned experiment, emergence of about 40 percent of the population occurred at the time of the light-dark transition even when no transition was imposed.
(ii) 24 h feeding rhythm
The time taken to resynchronize the phase of the rhythm with a phase shift in the light-dark cycle was considerably shorter in shrimp fed after the light-dark transition than in those unfed during the period of reentrainment. A feeding rhythm with an approximate 24 h periodicity, was probably responsible for this difference. This rhythm was indicated by experiments in which shrimp, induced to emerge to feed during the day, would again emerge and carry out food searching movements at approximately the same time the following day when no food was present.
A characteristic of this rhythm is the readiness with which it can be re-set, regardless of the extent of the shift involved. The strength of the response is suggested by the fact that emergence took place despite the inhibitive factor of the high light intensity.
The results are evidence for the existence of an endogenous “time sense” similar to that first described in bees by Forel (1910) and reviewed more recently by Renner (1960).
4.1 The possible adaptive significance of synchrony of emergence
The nocturnal activity of shrimp and the close association of its onset with the light-dark transition has a number of possible advantages. The observations of Fuss (1966) have indicated that shrimp active above the substrate during the day readily fall prey to many species of fish. A great measure of protection from predators is acquired, therefore, by the limitation of the time of activity to the time of darkness.
In inshore areas where tidal influence is strong, shrimp are passively displaced by either ebb or flood tide. By being passively transported solely by the flood tide, postlarvae are carried into the nursery areas while the juveniles, by being transported only on the ebb tide, are carried out of them (Hughes, 1969). Due to this mechanism of transportation, shrimp, emerging at very different times in the evening, would often become separated by great distances if no synchrony of emergence existed between all individuals.
In addition, it is probable that all activities occurring at night are related temporally to the time of onset of activity at dusk. Consequently, this initial synchrony will considerably influence the synchrony of subsequent activities.
The evidence for this derives from experiments carried out in the current-chamber described by Hughes (1969). Changes in direction or intensity of swimming within the current occurred at similar times in all individuals (manuscript in preparation). It is therefore possible that aggregations of shrimp are largely maintained by the rhythmic control over most activities, and their consequent synchrony in all individuals. Maintenance of these aggregations may have survival value with regard to predators and would be clearly advantageous at times of mating and spawning, producing groups at a similar developmental stage at suitable spawning sites.
The apparent advantage of the circadian rhythm controlling emergence and possibly subsequent activity is clear, but that played by the 24 h feeding rhythm is less clear. In regions where tidal influence is not strong, displacements associated with the ebb and flood tide may not occur. Under these conditions, their first activity following emergence will probably be feeding. Thus, the two rhythms will be virtually in phase and will reinforce each other in such a way that emergence of all individuals should become more closely synshronised. However, when regular displacements by tide occur, the feeding rhythm may interact with responses to ebb and flood tides, in such a way that feeding occurs only when it will not conflict with the time during which displacement by the tide occurs.
Aschoff, J., 1960 Exogenous and endogenous components in circadian rhythms. Cold Spring Harb.Symp.quant.Biol., 25:11–28
Aschoff, J., 1963 Comparative physiology: diurnal rhythms. A.Rev.Physiol., 25:581–600
Aschoff, J., 1965 Response curves in circadian periodicity. In Circadian clocks, edited by J. Aschoff, Amsterdam, North-Holland Publishing Co.
Dall, W., 1958 Observations on the biology of the greentail prawn, Metapenaeus mastersii (Haswell) (Crustacea Decapoda:Penaeidae). Aust.J.mar.Freshwat.Res., 9(1) :111–34
Egusa, S. and T. Yamamoto, 1961 Studies on the respiration of the “Kuruma” prawn Penaeus japonicus Bate. - 1. Burrowing behavior with special reference to its relation to environmental oxygen concentration. Bull.Jap.Soc.scient.Fish. 27:22–6
Eldred, B., 1958 Observations on the structural development of the genitalia and the impregnation of the pink shrimp, Penaeus duorarum Burkenroad. Tech.Ser.Fla St.Bd.Conserv., (23):26 p.
Eldred, B. et al., 1961 Biological observations on the commercial shrimp, Penaeus duorarum Burkenroad, in Florida waters. Prof.Pap.Ser.mar.Lab.Fla., (3):1–139
Forel, A., 1910 Das Sinnesleben der Insekten. München, Reinhardt
Fuss, C.M., 1964 Observations on burrowing behaviour in the pink shrimp, Penaeus duorarum Burkenroad. Bull.mar.Sci.Gulf Caribb., 14(1):62–73
Fuss, C.M. and L.H. Ogren, 1966 Factors affecting activity and burrowing habits of the pink shrimp, Penaeus duorarum Burkenroad. Biol.Bull.mar.biol.Lab.,Woods Hole, 130(2):170–91
Hughes, D.A., 1969 On the mechanisms underlying tide associated movements of Penaeus duorarum. FAO Fish.Rep., (57) Vol.3:867–74
Idyll, C.P., 1950 The commercial shrimp industry of Florida. Educ.Ser.Fla St.Bd Conserv., (6):1–31 (Re-issued, August 1957)
Iversen, E.S. and C.P. Idyll, 1959 The Tortugas shrimp fishery: the fishing fleet and its method of operation. Tech.Ser.Fla St.Bd Conserv., (29):1–35
Racek, A.A., 1959 Prawn investigations in eastern Australia. Res.Bull.St.Fish.N.S.W., (6):57 p.
Renner, M., 1960 The contribution of the honey bee to the study of time-sense and astronomical orientation. Cold Spring Harb.Symp.quant.Biol., 25:361–67
Viosca, P., 1957 The Louisiana shrimp story. Bienn.Rep.La.Dep.Wildl., 7:102–5
Wickham, D.A., 1966 Observations on the patterns of persistent activity in juvenile pink shrimp, Penaeus duorarum Burkenroad. University of Miami (MS Thesis)
Williams, A.B., 1955 A contribution to the life histories of commercial shrimp (Penaeidae) in North Carolina. Bull.mar.Sci.Gulf Caribb., 5(2):116–46
Williams, A.B., 1958 Substrates as a factor in shrimp distribution. Limnol.Oceanogr., 3(3):283–90
![]()
![]()
![]()