![]()
![]()
![]()
The genesis of integrated pest management (IPM) was a response to the emergence of problems associated with the reliance on chemical controls for insect pests by governments, extension systems and farmers. The search for solutions to these problems led to the development of a more holistic view of what constituted an agro-ecosystem and how human intervention could either enhance or disrupt one. This chapter briefly looks at the development of IPM in general, its application in tropical irrigated rice and the evolution in the understanding of tropical irrigated rice-field ecology.
With the introduction of chemical insecticides following World War II, plant protection specialists were able, almost without restriction, to make use of powerful poisons. The goal of plant protection specialists in the early 1950s was 100-percent control (eradication). Entomologists such as Ray F. Smith and A. E. Michelbacher sounded warnings against such unrestricted use and proposed an approach that tolerated low levels of pest populations to allow natural enemy populations an opportunity to suppress pest populations. They proposed an approach to crop protection that was based on the analysis of the ecology of a given agro-ecosystem and included biological controls. Their work resulted in the development of what was known as integrated pest control. They proposed that the goal of pest control strategies was to tolerate pest populations that were below specified threshold levels. (J.A. Litsinger, Peter E. Kenmore and G. Aquino 1982)
Box 6.1 contains a summary of a presentation by R.F. Smith and H.T. Reynolds to an FAO symposium in 1966 concerning the ecological approach inherent in integrated pest control.
|
Box 6.1 Principles of integrated pest control 1. The use of chemical pesticides, without regard to the complexities of the agro-ecosystems in which they are used, has been a major cause of disruption and undesirable side effects. These include: target pest resistance and/or resurgence, secondary pest outbreaks, residue problems and environmental pollution. 2. The agro-ecosystem is a unit composed of the total complex of organisms in the crop area together with the overall conditioning environment. There must be an analysis of the agroecosystem to determine population dynamics and mortality factors operating on pest populations. 3. The kinds of crops, agronomic practices, patterns of land use, weather, total complexity, and self-sufficiency of the agro-ecosystem affect its stability. As complexity increases, particularly among trophic interactions, there is usually an increase in the stability of the agro-ecosystem. Integrated pest control should seek to preserve or improve this complexity. 4. Levels or limits of tolerable damage are more important than pest population levels. Tolerable levels of damage vary widely with market conditions, stage of the crop, local conditions or grower economics, and the personal values of the people concerned. The presence of pests is not an indication of a threat of economic damage to the crop. 5. In more sophisticated programmes, individual fields are surveyed for populations of pests, parasites, predators and pathogens. On the basis of this information and consideration of the time of the year, stage of growth of the crop and weather conditions, a prediction can be made of population trends and potential damage. This type of sampling and prediction requires a solid base of fundamental biological and ecological data. 6. All but the most sterile of man-made environments have some biotic agents influencing pest populations. Appropriate consideration must be given to biotic control agents. In some fortunate situations, the biotic agents are all that is necessary to have satisfactory economic control. The failure of natural enemies to keep a given pest under control should not cause us to invoke control practices that disrupt the controlling action of natural enemies of other species in the same agroecosystem. (R.F. Smith and H.T Reynolds 1966) |
Integrated pest control eventually became integrated pest management. The stress on decision-making and integration of tactics and the allowance of tolerable thresholds were seen as a move away from control and towards management (J.A. Litsinger, P.E. Kenmore and G. Aquino 1982).
By the 1970s, according to Barfield and Swisher (1994), two schools of thought regarding IPM emerged:
1. The school of tactical IPM. For this group, IPM required the use of pesticides, but only when necessary. This school promoted the technological fix.
2. The school of strategic IPM. This group emphasized the importance of understanding the agroecosystem. They promoted the ecological approach to pest control.
The first school gave rise to crop scouting and economic thresholds as tools to decide when to spray. In essence, this school adopted a position of cooperating with the pesticide industry. They treated farmers as passive recipients of technology. The second school stressed ecological insight and the avoidance of the use of insecticides. This school embraced farmers as partners in agricultural development (ibid). By the late 1970s, the ecological foundation of IPM was overshadowed and the tactical school was ascendant. Crop protection systems were advocating multiple tactics, the timing of pesticide applications, government subsidies to lower pesticide costs for farmers, economic threshold levels, and national forecasting systems. By the early 1980s, IPM had become so closely associated with pesticide use that the term was interpreted by many to mean integrated pesticide management. (Monica Moore 1996)
Asian rice farmers were to feel the impact of the ascendancy of the tactical school of thought in IPM with the coming of the Green Revolution. In the early 1960s the first high-yielding rice varieties were released by IRRI. International researchers urged the use of insecticides to control pests. Essentially this advice was accepted because it seemed an astute approach to protecting the investment that had been made in the international research and development of these new varieties. The new varieties were assumed to be irresistible to pests and the cost of pesticide was seen as an acceptable insurance premium. Because farmers were seen as being unable to deal with the complexities of pest control, prophylactic use of insecticides was promoted. This control tactic relied on calendar-based applications of pesticides. Farmers were told to apply insecticides based on the number of days after transplanting or the stage of plant development. In Indonesia, the calendar-based approach resulted in farmers applying broad-spectrum systemic insecticides to the nursery bed and at transplanting. Farmers then made additional applications of non-systemic broad-spectrum insecticides some forty days after transplanting and at or around the milky stage of seed development. The government, apparently not content with promoting calendar applications to farmers, contracted with companies to provide large-scale aerial applications of insecticides.
Tactical IPM arrived in tropical rice production systems with the introduction of economic threshold levels (ETLs). This tactic seemed to be a rational way to reduce the amount of insecticides that were being applied under the calendar-based system. ETLs failed to take into account the role of natural enemies in suppressing pest populations. General ETLs were developed for insecticide use throughout a given country. Farmers were told to count the number of a pest, for example BPH, and spray when there were "x" many BPH per hill of rice. The ETLs were often set at artificially low levels to reduce risk. During the 1970s ETLs in different countries varied widely. In Japan ETLs for BPH were set at five per hill, in the Philippines 20 adults per hill at 40 days post-transplanting, in India the ETL was estimated at two to five nymphs or adults per hill (K. Sogawa and C. H. Cheng 1979). ETLs were developed for all insect pests and, given the steady increases of sales of insecticides across the region, appeared to encourage insecticide use rather than reduce it.
National forecasting systems were another approach taken to control pests. These systems took the onus for decision making off the farmer and allowed professionals to take charge. Forecasting systems based their control tactics on ETLs. The greater the fear of a particular pest the lower the ETL and the more pressure put on farmers to apply insecticides. Surveillance systems sometimes made recommendations to farmers regarding insecticide applications. The systems also included pest eradication brigades to handle outbreaks. Outbreak stocks of insecticides were purchased by the crop protection system and stored for distribution to locally organized pest eradication brigades when the forecasting system detected an outbreak.
Prophylactic spraying, the use of ETLs, and forecasting systems all placed limitations on the management role of rice farmers. Crop protection experts assumed that professionals could take better decisions than could farmers. It was not until 1990, when the Indonesian government instituted a farmer IPM education programme, that a truly farmer-based IPM system was promoted. The process of getting to this point required several discoveries on the part of researchers and the realization on the part of both scientists and government that it was appropriate for farmers to be the ultimate decision makers in an IPM crop protection system.
|
Box 6.2 Hunting spiders: predators of BPH
Predators are the most important natural enemies of BPH. Together with parasitoids and insect pathogens they keep populations of BPH down. An important group of predators commonly found in rice fields is the spiders. Of particular importance are the hunting spiders, especially Lycosa pseudoannulata, which is often found near the water level in the same area where BPH feed. A lycosid will eat as many as 20 BPH per day. The spider's voracious appetite makes it a very important natural enemy of BPH. |
Scientists began working out some of the basics of rice-field ecology in the late 1970s and early 1980s. Their attention was generally focused on the brown planthopper, which at the time was ravaging rice fields across Southeast Asia. They discovered that BPH outbreaks were insecticide induced (P.E. Kenmore 1980) and that breeding rice varieties resistant to BPH under continued pressure of insecticide use was futile (K.D. Gallagher 1984). Researchers soon worked out an ecologically based means to BPH control (P.A.C. Ooi 1988). Yet it was to take some time before the broader ecological approach became part of mainstream extension. An example of the slowness of the ecological approach to catch on is the Illustrated Guide to Integrated Pest Management of Rice in Tropical Asia, published by IRRI in 1986, six years after Kenmore demonstrated that BPH outbreaks were pesticide induced. This manual for extension workers and other agriculturalists used 211 pages to describe 35 specific pests, 19 pages discussed pesticides and their use and only 20 pages were devoted to pest parasites and predators.
|
Box 6.3 Cyrtorhinus lividipennis An important predator of BPH is the mirid bug, Cyrtorhinus lividipennis. It feeds on eggs and very young nymphs of BPH. Ecological studies show that this mirid is very sensitive to build-ups of BPH populations and usually increases in population in response to increasing BPH populations.
A study in Sekinchan, Malaysia (Ooi 1986), showed that a build-up of BPH was followed by a similar buildup of Cyrtorhinus. Unlike spiders, Cyrtorhinus populations react to the presence of BPH. However, this tendency is disrupted when insecticides are applied. To understand this, two fields were selected. In one field, the farmer continued with his spraying while in another field, he was persuaded not to spray and to observe what happened. Weekly samples were taken of pests and predators in both fields. The field that was not sprayed regularly was the only field in the block that did not suffer from BPH-caused "hopperburn". This was attributed to the presence of predators, especially C. lividipennis. |
Eventually most researchers came to accept that Asian rice farmers were gifted with what R.A Smith had called a "fortunate situation"; biotic agents in a rice agro-ecosystem were able to control pests within tolerable levels as long as insecticides were not allowed to disrupt that system (P.A.C. Ooi and B.M. Shepard 1994; M.J. Way and K.L. Heong 1994).
During the 1980s there was an increasing number of studies on the various predators and parasites of the main pests in tropical rice. Pests no longer occupied centre stage by themselves; they had to share it with their natural enemies. The following table, based on Ooi and Shepard (1994), presents the common predators and parasitoids that cause mortality among rice stemborers, leaffolders, brown planthoppers and green leafhoppers.
Table 6.1 Predators and parasitoids of common insect pests
|
Stage attacked |
|||
|
Yellow stemborer |
Egg |
Larva |
Pupa |
|
Parasitoids |
Tetrastichus schoenobii |
Temelucha philippinensis |
Tetrastichus ayyari |
|
Ferriere |
(Ashmead) |
Rohwer |
|
|
Trichogramma japonicum |
Stenobracon nicevillei |
|
|
|
Ashmead |
(Bingham) |
|
|
|
Telenomus rowani |
Bracon chinensis |
|
|
|
(Gahan) |
Szepligeti |
|
|
|
Predators |
Conocephalus longipennis |
Lycosa pseudoannulata << |
[also attacks adults] |
|
(de Haan) |
(Boesenberg and Strand) |
|
|
|
Micraspis spp. |
|
||
|
Ophionea spp. |
|
||
|
Ants |
|
||
|
Microvelia douglasi atronlinea |
|
||
|
Mesovelia vittigera |
|
||
|
Limnogonus fassarum (F.) |
|
||
|
Leaffolders |
Egg |
Larva |
Pupa |
|
Parasitoids |
Copidosomopsis nacoleiae |
Temelucha philippinensis |
Xanthopimpla |
|
(Eady) |
Cotesia augustibasis |
flavolineata Cameron |
|
|
Trichogramma japonicum |
(Gahan) |
Tetrastichus ayyari |
|
|
(Ashmead) |
Trichomma enaphalocrosis |
Rohwer |
|
| |
Uchida |
|
|
|
Predators |
Conocephalus longipennis |
Lycosa pseudoannulata |
|
|
M. vittaticollis |
Micraspis spp. |
|
|
|
Cyrtorhinus lividipennis |
Ophionea spp. |
|
|
|
Reuter |
Ants |
|
|
|
Micraspis crocea |
Water bugs |
|
|
|
Mulsant |
|
|
|
|
Brown planthopper |
Egg |
Nymph |
Adult |
|
Parasitoids |
Oligosita yasumatsui |
|
|
|
(Viggiani and Subba Rao) |
|
|
|
|
Anagrus spp. |
|
|
|
| |
Pseudogonatopus spp. |
Pseudogonatopus spp. |
|
|
Predators |
Cyrtorhinus lividipennis |
Cyrtorhinus lividipennis |
|
|
Lycosa. pseudoannulata |
Lycosa pseudoannulata |
||
|
Migrovelia douglasi |
Migrovelia douglasi |
||
|
atrolineata Bergroth |
atrolineata Bergroth |
||
|
Synharmoni octomaculata |
Synharmoni octomaculata |
||
|
(F.) |
(F.) |
||
|
Paederus fuscipes Curtis |
Paederus fuscipes |
||
| |
Curtis |
||
Table 6.1 provides some indication of the myriad parasitoids and predators that exist in the tropical rice agro-ecosystem. In this table, the predators and parasitoids that attack yellow stemborers, brown planthoppers and green leaffolders are presented. The predator complex that feeds on eggs and larvae of leaffolders is nearly the same as for stemborers. A major exception is the cricket, M. vittaticollis, which is a voracious feeder on the eggs of leaffolders. Leaffolder egg predation averaged about 50 percent in both direct-seeded and transplanted rice in field experiments. Kamal (1981) reported 70- percent leaffolder mortality by predators. The parasitoid complex for leaffolder larva and pupa is richer than that for eggs. Rao et al. (1969) listed 32 species of larval and eight species of pupal parasitoids for rice fields in India (1969).
|
Box 6.4 Conocephalus longipennis (de Haan) Parasitoids and parasites cause mortality among all the various species of stemborers. C. longipennis has been reported to consume up to 65 percent of the eggs of yellow stemborers (Pantua and Litsinger 1984). This grasshopper will reportedly feed on rice panicles, but its role as a predator is far more important. C. longipennis can consume more than eight yellow stemborer egg masses in three days (Rubia et al. 1990). C. longipennis feeding on stemborer eggs
|
The generalist nature of some predators is apparent in Table 6.1. The wolf spider, L. pseudoannulata, feeds on stemborer larvae and adults, leaffolder larvae and the nymphs and adults of BPH and green leafhoppers. The mirid, C. lividipennis, feeds on the eggs and nymphs of the BPH, the green leafhopper, and the eggs of leaffolders. The grasshopper, C. longipennis, feeds on the eggs of stemborers and leaffolders.
Parasitoids tend to be more specific in their tastes. The predators and parasites that appear in the table are but the tip of the iceberg. Settle et al., in four years of sampling irrigated rice fields on the island of Java, found 765 species of arthropods. Of that number, 40 percent or 306 species were predators and 24.4 percent, 187 species, were parasitoids. Herbivore species accounted for only 16.6 percent or 127 species.
Tetrastichus schoenobil, a parasite attacking stemborer eggs

(To summarize the above: early pest control specialists looked only at pests and they tried to eradicate them. Smith and others promoted a broader ecological perspective in trying to control pests. They advocated an analysis that included pests, predators, pathogens and the plant. This perspective lost out to the simplification of counting pests and spraying based on economic thresholds. Researchers and crop protection specialists in tropical Asia fell in line with this perspective until they were jolted out of it by massive outbreaks of BPH. By the early 1990s a picture emerged of the ecology of the tropical ricefield agro-ecosystem that suggested that crop protection in rice could not be successful unless its analysis included the plant, pests, predators and parasitoids. Yet the picture was not complete. Most of the components of the rice-field agro-ecosystem were understood, but no one had offered an explanation of how the system worked as a whole. Settle and his group, working as part of the FAO technical assistance team to the Indonesian national IPM programme, put the puzzle together. They were able to develop a general explanation of how the tropical irrigated rice agro-ecosystem functioned. The following is an extract from a paper presented by Settle to a conference in China in 1998, in which the functioning of the tropical irrigated rice agro-ecosystem is explained.)
Whereas the specific conditions critical for management decisions regarding inputs vary over a small spatial scale, agro-ecosystems do have a general structure and dynamics that are reasonably consistent for the entire system. In essence, it is possible for us to think in terms of a general theory for the structure and dynamics of specific agricultural ecosystems. IPM is not a theory in a strict scientific sense; rather, it is a set of practical guidelines for how to best manage a specific crop. The rice IPM FFS learning approach, however, is based on well-established scientific theories, supported by good field data.
The fact that all ecological systems, even highly complex ones like tropical irrigated rice, are structured by a very few key variables allows us to propose a general theory for any ecological system. Research over the past 20 years in applied ecology of managed systems shows that ecosystem dynamics, regardless of the system, are organized around a small number of nested cycles, each driven by a few dominant variables (Gunderson et al. 1995, Holling 1992).
"A small number of plant, animal and a-biotic processes structure biomes over scales from days and centimetres to millennia and thousands of kilometres. Individual plant and biogeochemical processes dominate at fine, fast scales; animal and a-biotic processes of mesoscale disturbance dominate at intermediate scales; and geomorphological ones dominate at coarse, slow scales. The physical architecture and the speed of variables are organized into distinct clusters, each of which is controlled by one small set of structuring processes. These processes organize behaviour as a nested hierarchy of cycles of slow production and growth alternating with fast disturbance and renewal." (Gunderson et al. 1995, p. 27)
The basic outline of how a few key variables and processes determine the dynamics of an entire ecosystem in irrigated tropical rice are as follows (see Settle et al. 1996 for details).
Key process #1: energy is stored as organic matter in the soil and brought into the system by micro-organisms and detritivorous insects (Figures 5 and 6). From the time that water first floods a farmer's field in preparation for planting, organic matter - derived from residues from the previous crop cycle, organic waste in irrigation water and algal growth - provides the energy for an array of micro-organisms. The energy flow begins with bacteria being eaten by protozoans and rotifers and continues upwards to larger zooplankton. In a parallel flow, detritivorous insects, such as the larvae of flies and beetles and especially the minute but abundant Collembola, feed directly on decaying organic matter, including material floating on the surface of the water. This process is fundamental to aquatic systems and will be found in all irrigated rice systems.
Figure 5 Trophic-level energy flow diagram for tropical irrigated rice
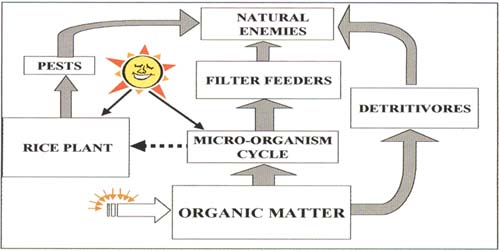
Consistently low pest populations in tropical rice result from the fact that natural enemies - especially generalist predators - are not directly dependent on pest populations. Rather, there are three separate avenues for energy flows to natural enemy populations:
1.from organic matter via micro-or ganism cycles and filter -feeding insects,
2.from organic matter via detritivorous insects and
3.from the rice plant via herbivores.
A more detailed elucidation of the energy-flow diagram for tropical irrigated rice at the functional group level is shown in Figure 6.
Figure 6 Functional-group-level energy flow diagram for tropical irrigated rice
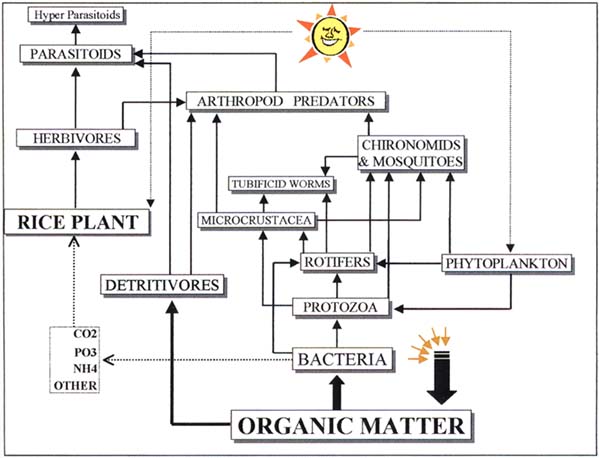
Key process #2: aquatic plankton provides food for filter-feeding insects. The small to mediumsized zooplankton and the phytoplankton found in all flooded rice fields provide food for filter feeders (midges and mosquitoes). Densities of chironomid midge larvae have been observed in irrigated rice fields in the thousands/m2 range. Again, abundant midges and mosquito populations are a general characteristic of rice systems.
Key process #3: Larvae and adult plankton feeders and detritus feeders provide a consistent and abundant source of alternative food for generalist predators from early in the season until after harvest. Figure 7 shows the results of observations in two plots in a field in West Java. One plot, "a", was not treated with insecticides; the other, "b", received early-season applications of carbofuran and azodrin. Alternative prey for predators, in the form of filter feeders and detritivores (or neutrals), show a consistent peak at about 30 days after transplanting (Figure 7a). The existence of abundant alternative prey effectively frees predator populations from a dependence on the pest populations, giving predators the opportunity to develop well in advance of the normal pest populations. As a result, mortality of pest populations due to predation is high -beginning with early-season pest migrants and carrying on throughout the season. This process minimizes the likelihood that pest populations can escape control by natural enemies and reach outbreak levels. Furthermore, high populations of detritivores found in harvested stubble fields assure a continued food source for generalist predators for some time after harvest.
Figure 7 Trophic-level population dynamics in a field in West Java, 1992
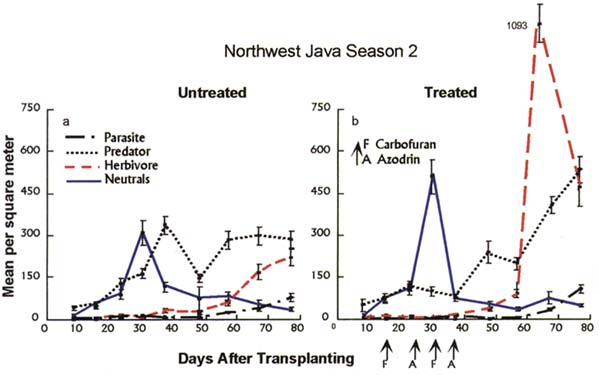
In plot "b", early-season applications of insecticides by farmers lead to a suppression of early-season predator populations and a subsequent resurgence of pest populations (mostly BPH). This is a classic example of pesticide-induced pest resurgence, the phenomenon that is responsible for the problems with BPH over the past 30 years.
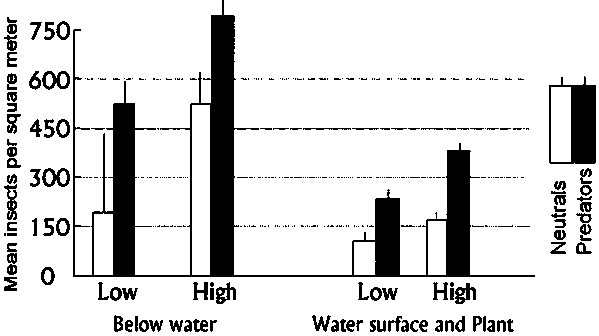
In order to test the hypothesis of whether energy flows from organic matter to predators in tropical rice systems (Figures 5 and 6), plots were set up with low and high organic matter and examined during the first 40 days. High organic matter resulted in significantly more neutrals and predators below the water, on the water surface and in the plant (Figure 8). A somewhat different experimental approach conducted in China reached the same conclusion (Wu et al. 1994). Wu et al. eliminated midge and mosquito larvae by using highly selective Bacillus sphaericus insecticide. In response, predator populations in early-season irrigated rice were diminished for lack of food and pest populations were subsequently higher compared with controls.

Figure 8 Soil organic matter experiment
Results of six Indonesian surveys show that some 19 percent of the arthropod species are detritivores or plankton feeders (Figure 9). Except for pioneering work done in China (e.g. Wu et al. 1994) previous recognition of these functional groups has been largely ignored or misunderstood. For example, chironomids were all considered to be root feeders (phytophagous) and therefore thought to be harmful, but this is not generally true (see Settle et al. 1996 for details). Recent studies in southern China using ELISA techniques have shown that chironomids represent up to 80 percent of the diet of spiders early in the season (Zhang Wenqing, Institute of Entomology and State Key Laboratory for Biological Control, Zhongshan University, Guangzhou, pers. comm. to W.H. Settle).
Figure 9 Trophic level distributions from vacuum samples over six sites in West and Central Java - A. Number of Species
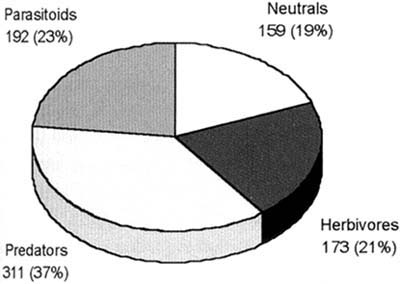
Figure 9 Trophic level distributions from vacuum samples over six sites in West and Central Java - B. Fraction of Total Abundance
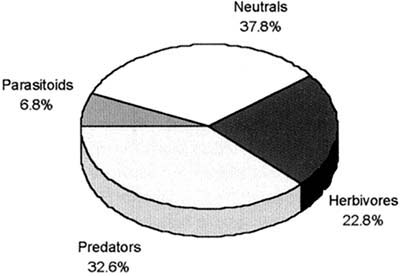
The existence of diverse populations of natural enemies, supported by abundant alternative food species, assures that populations of pests are consistently maintained at low levels. In effect, the structured biodiversity of arthropods in tropical irrigated rice functions to consistently suppress pest populations by denying them refuge in time or space. All the key variables can be found in any rice ecosystem - only when the process is disrupted do pest populations explode, causing serious damage. From this, a set of implications for rice IPM practice can be determined.
The use of insecticides disrupts and destabilizes natural enemy populations. The use of insecticides is by far the most common cause of pest outbreak, especially for pests such as the rice brown planthopper. These kinds of pest outbreaks are generally referred to as pesticide-induced resurgence (Figure 7).
Several factors combine to enable resurgence to occur.
Eggs of many pests, such as BPH, are not susceptible to chemical sprays.
Insecticides create a refuge for the development of pest populations by reducing the abundance of natural enemies.
Migratory abilities of pests are generally better and their generation is many times faster than those of natural enemies.
Certain landscape designs can cause delays in the arrival of natural enemies after long dry fallow periods. In many tropical areas, the potential exists for year-round cultivation of rice. However, some areas, either from natural constraints or by government design, are planted synchronously over thousands of hectares and have long (3-4 month) dry fallow periods. These are the areas in which natural enemy populations are weakest and where pest outbreaks are most frequent (Settle et al. 1996). Take, for example, a typical area in North West Java: in the middle of a 3 000 ha area, dry and devoid of refuge for 3-4 months, there simply is not enough food and shelter to support significant natural enemy populations and some species are absent altogether. After such a long dry spell, when the next season of rice is planted, it takes up to half a season to build up predator populations that would otherwise have been there from the beginning. Again, as pests are better re-colonizers than natural enemies, the potential for outbreak is much higher in these synchronous, large-scale areas.
Much attention has been given to the ideas of synchronous planting and breaking the pest cycle with long dry fallow periods, to the point that these ideas have become ingrained almost as fundamental principles of IPM. However, a close look at the empirical data and experiments that purport to demonstrate these principles will show that the support is very weak. Finally, our developing theory of irrigated rice ecosystems allows us to question the logical foundations of such principles.
Healthy soils, high in organic matter, are the foundation for a healthy ecosystem. Soil organic matter is not emphasized as much for irrigated rice as for dry-land cropping systems because of the lesser need for good soil texture. However, as we have seen, soil organic matter is the foundation for energy cycles that ultimately support high populations of natural enemies. We also know that high levels of soil organic matter help prevent leaching of nutrients in sandy and degraded soil conditions. Areas that entirely remove or burn their rice stubble, putting no soil organic matter back into the system, may suffer from lower natural enemy populations and more frequent pest outbreaks (we have some supporting evidence for this, but this is a hypothesis that needs to be examined more generally).
One of the biggest problems with many of the developments in IPM over the years has been the tendency to generalize and make recommendations for farmers across large and highly heterogeneous areas. This has been true for all manner of input recommendations including fertilizers, pesticides and rice varieties. This problem, ecological heterogeneity, has also severely limited the effectiveness of government monitoring and forecasting systems. All of these practical issues vary on a small spatial scale. This local specificity requires that farmers become IPM experts. The main crop protection approaches since the late 1960s, from the perspective of donor support, are presented in Table 6.2. The recommendations or decision criteria of each approach reveal a steady progression in the accommodation of ecological heterogeneity and farmer control of agro-ecosystem management.
Table 6.2 Pest control approaches in tropical irrigated rice
|
Calendar-based applications |
Surveillance systems |
ETL-based decisions by farmers |
Farmers as IPM experts |
|
Farmers, in this approach, apply insecticides based on number of days post-sowing or transplanting. Goal: prophylactic control of pest populations. Relies on broad recommendations and assumes homogeneity among planting conditions. Developed in 1960s. |
Usually an activity of agriculture departments. Based on ETLs
developed at national level to be applied in widely differing
conditions. |
The count-and-spray approach relies on use of criteria that
assumes all local agro-ecosystems. |
Farmers as decision Makers; decisions based on analysis of
agro-ecosystem. |
Governments across Asia have enacted policy in support of one or more of the four approaches presented above. Some countries have supported each of the approaches over the last four decades, often using more than one approach at the same time. Countries have often adopted new approaches without abandoning old approaches, despite glaring contradictions. Presented in roughly chronological order of emergence from left to right, these four approaches place an increasingly larger burden on the user in terms of ecological knowledge, observation and analysis. Each successive approach requires more data for decision making and the decisions made cover increasingly smaller units of area and time. This increased precision in decision making, not surprisingly, has led to better control of insect pests and reduced use of pesticides. The FFS was designed to address the problem of ecological heterogeneity and local specificity by placing the control of small-scale agro-ecosystems in the hands of the people who manage them.
![]()
![]()
![]()


