![]()
![]()
![]()
The majority of the sites surveyed were based on the need for a semi-quantitative assessment of the resource in the proximity to Tongatapu I., the principal area of current commercial interest. Additional assessment was made of a known area in the Ha'apai Group and peripheral areas of likely occurrence. The Vava'u Group was also surveyed.
A understanding of the ecological requirements of Cladosiphon sp. is important in understanding the basis for the survey. Colonisation of habitat by the algae require three circumstances. The first is a substrate suitable for settlement by the zoospore phase. Secondly, the progressive development of the filaments by the sporophyte phase require sufficient support to maintain their position in the presence of the environmental conditions conducive to growth. Thirdly, the dispersion via current by Cladosiphon filaments require suitable areas for entanglement. A semi-protected environment with slight wave action and current are characteristic of the preferred habitat.
Settlement by spores may occur on a variety of substrates such as shell, coral surfaces or, importantly for maricultural purposes, nets. Field observation has revealed an recurring relationship between Halimeda cylindracea, H. macroloba (Fig. 7b, j) and Cladosiphon where the dead portions of the Halimeda provide substrate for settlement.
Additional to the spore settlement, is the dispersion by fragmentation of the filament-like fronds or thalli (Fig. 7f). This occurs as the fronds grow from their attached or entwined position and break off to float in the current to be ensnared by benthic material which will allow a secured basis for further growth (Fig. 7a). Though free-floating material has been observed to have fouled a variety of objects such as reef rock or other algaes, it is the meadows of Syringodium isoetifolium which provide the main habitat in which an abundance of the algae can develop. Entwining and matting of the algae occurs within as its biomass increases. It is in these areas that substantial quantities of Cladosiphon exist (Figs. d-i). The stoloniferous and upright mode of growth of Halimeda spp. provide good support for the free floating filaments.
Based of this observation, the inspection of likely locations of occurrence were confined to the presence of the seagrass beds. This relationship is complicated by the presence of two genera of seagrasses, Thalassia hemprichii which is present as monospecific beds or mixed grass meadows with Syringodium isoetifolium. Thalassia due to the flaccid nature or shape of its leaves is generally unsuitable for Cladosiphon development as it is unable to retain the algae or leaf edge abrade it. Due to the unsuitable nature of the genus, inspection of the beds is required to determine the type of seagrass and whether it has been colonised and to what degree. Thalassia was colonised by Cladosiphon in the deeper water, south of Maria Bay.
The requiremented characteristics of the grass beds are known (sandy substrate, shallow depth) and appear on aerial photographs clearly.
Areas which negate the successful colonisation of the algae are coral reef or open sandy areas. High current or wave action is also not conducive to proliferation, nor are the stagnant waters of the enclosed lagoons where circulation and temperature and salinity may vary widely.
The areas visited during the survey of Tonga are described in Table 1. The table details the site name and position, the community type, substrate, depth, current velocity and direction, and the presence or absence of Cladosiphon.
The survey of the Tongatapu area was divided into general four areas. 1) These are the fringing reef areas adjacent to Manuka and Navutoka, east of Fanga uta Bay, extending west to Talafo'ou and south to Nukuleka. 2) The embayed area south of Maria Bay to include Atata and Toke Islands and west to Tongatapu's western peninsula. 3) The offshore coral reef areas north of the eastern end of Tongatapu to include the adjacent islands of Nuku, Oneata, Manima, Pangai Motu, and Makaha'a islands. 4) Offshore the reef area inspected encompassed Monuafe, Okotau and Onevai, and Onevao.
Figure 2a, b details the area of initial harvest along the Navutoka coast as well as the island sites of harvest to the east of Nuku'alofa. Figure 3a, b details the embayed sand flats and reef west of Nuku'alofa and south of Maria Bay which includes Atata I. and Toke I. Figure 6a, b shows the offshore barrier reef areas north of Navutoka.
The presence of Cladosiphon is confined to Felemea Bay on 'Uiha I. and to a small area on Lifuka I. at Muikuku. The remainder of the area surveyed comprises fringing reef and sandy substrates which are not conducive to Cladosiphon development.
Figure 4a,b shows the area surveyed in the western Ha'apai Group. Also, the offshore reef of Nukupule (Fig. 6c), where Cladosiphon reportedly occurs, is shown to illustrate the general nature of the fringing reefs and the very confined area of the seagrass beds.
No Cladosiphon was found. All known sites of seagrass occurrence were visited. Figure 5, 5a shows the areas surveyed in the Vava'u Group.
The following figures (2a,b; 3a,b; 4a,b; 5, 5a; 6a-c, 6d) are aerial photographic views of known areas of Cladosiphon sp. occurrence. From the field survey, the general nature of the habitats are known sufficiently well to be able to estimate the areal dimension of the seagrasses. From this, some idea of the potential area of Cladosiphon can be made. The figures are presented in pairs with the latter being from the New Zealand Admiralty chart providing locational information.
Assessment of the aerial photographs has been made on the basis of limited ground truthing, much of which was retrospective from field notes. There is good confidence in the accuracy of the assessment but further ground truthing would add to its accuracy. This would be particularly valuable at the beginning of the season prior to harvest.
Assessment of the seagrasses from aerial photographs is made by correlating the areas of occurrence with the image on the photograph. Generally, the dense seagrass appear as dark areas. A good example of this is along the Navutoka coast. Equally clear are the areas in Felemea Bay, 'Uiha. Without ground truthing, confusion exists with respect to the genus of seagrass though habitat requirements usually confine Syringodium sp. to the outer portion of the reef edge. Monospecific or mixed algal occurrence also appear as dark patches and needs to be clarified. This is particularly the case in the western embayment on Tongatapu I. General fringing reef with the rubble areas are clearly discernible.
A comparison of Cladosiphon habitats is made in terms of areal estimates so that the relative extent and potential productivity of these areas can be discussed. The estimates are very general and reflect an approximation of seagrass cover. The figure does not take into account of the patchiness of the seagrass, which in some areas is substantial and may be in excess of 50%. The areas adjacent to the coast are subject to greater tidal exposure and as a consequence there is a predominance of the seagrass Thalassia, in shore. Though Cladosiphon does colonise Thalassia seagrass, it co-occurs almost exclusively with Syringodium sp. Distinguishing between the two seagrasses is not possible from the aerial photographs which introduces substantial uncertainty as to the proportion of the seagrass which might be suitable.
Table 1: Summary of information on the sites surveyed
| Sites | Position: GPS or charted track | Community type: Seagrass, coral reef or other | Substra | Depth | Curr. cm/sec | Cladosiphon sp. presence: nature and abundance |
|---|---|---|---|---|---|---|
| Tongatapu Island: General | The areas considered represent three habitat types. Those of a narrow fringing reef, detached offshore reef, and the seaward margin of a largely intertidal bay. All areas of occurrence were dependent on the presence of the seagrass Syringodium isoetifolium. This species intergrades with the seagrass Thalassia sp. but is rarely host to substantial growths of the algae as is the Syringodium. | |||||
| Navutoka:(3 sites) Fig. 2a,b | 21° 07' 53.1"S 175° 06' 18.5"W | Seagrasses extending seaward with Thalassia dominate inshore in shallower water. At the seaward fringe of the flat the seagrasses are mixed with the prevalence of Syringodium becoming dominant. Sand margin seaward with patches of coral reef. Midway through the seagrass proliferation of Caulerpa racemosa. Filamentous algae colonising patches. Abundant Dictyota sp. Epiphytic fouling of grasses | Coarse sand with rubble patches. Seagrass and algae | 1m at low water Inshore grasses exposed | 10 | This is the area of initial harvest. Residual population present as both older matted material and that which appears as regrowth with filaments extending 15–20 cm. within the seagrass |
| 21° 07' 40"S 175° 05'71.5"W | " | " | Material collected for the growth study as a rope and tank culture. | |||
| 21° 07' 60"S 175° 06' 30"W | " | " | Largely older material, matted to a thickness f 10 cm. .55–.95 kg/m2 | |||
| Nukuleka | 21° 09' 28.3"S 175° 05'69.2"W | Mixed algal band, sand margins with reef patches | 30 | No Cladosiphon | ||
| Eastern embayment south of Maria Bay: General Fig. 3a,b | Within area of 21° 4'–30'S 175° 15'– 20'W | Fringing reef which margins a sand expanse which shallows into a the bay. | Sandy with extensive seagrass beds and mixed algae. Expanse of sand on the lagoon floor. | Lagoon margin 4–6m Max 10m. Flats 1–2m mlw | 5–10 | Principal Cladosiphon locality in terms of abundance and extent. .91 – 3.75kg/m2 in densest area. |
| Atata I. and Tufaka I Fig. 3a,b | 21° 02' 80.2"S 175° 15'29.8"W to 21° 03' 78.1"S 175° 16'20.1"W to 21° 04' 75.3"S 175° 15' 50.2"W | Mixed seagrasses inshore with Syringodium dominant on outer reef flat | course sand with less rubble in the lagoon | 1–3m | - | Present in the area west of Atata on outer flat. .85kg/m2 (n=3) Sparse occurrence to Tafaka due to patchiness of seagrass. |
| Shallow margin South of Maria Bay | 21° 04' 75.3"S 175° 15'50.2"W to 21° 05' 08.8"S 175° 16'27.5"W to 21° 04' 62.8"S 175° 15'35.5"W | Broken margin of fringing reef around bay | Patches of reef rising 2–3 m | - | ||
| South of Toke Toke I. | 21° 03' 95"S 175° 17'60"W | Areas of seagrass with dense algae. Mixed algal areas. | Sand and reef on either side of peninsula of seagrass | 2m | 5–10 | Southern portion of penn. inspected with dense quantities of Cladosiphon. North to Toke I. reported to have abundance. |
| Eastern side of embayment | 21° 03' 95"S 175° 19' 50"W | Seagrasses on hummocky sand | " | 1–2m | - | |
| Islands and reef west margin of Nuku'alofa Fig. 3a,b | 21° 03' 95"S 175° 19' 62"W | Seagrasses dominant with expanses of mixed and monospecific algae | " | 1–2m | - | |
| Nukunuku; Oneata; Manima; Pangai Motu Fig. 2a,b | 21° 03' 95"S 175° 07' 75"W | " | 1–2m | - | Thalassia meadows occur to the west. Eastern side requires further inspection. | |
| Detached reef area north of Tongatapu | A platform reef with mixed algal and seagrasses inside the seaward reef margin. Cladosiphon was found between the islands of 'Onevai and Onevao in 2m depth at mid tide. The remainder of the area is either reef outcrops, Thalassia beds or sandy bottom. The occurrence is linked to the presence of Syringodium meadows and to the algae H. cylindracea. | |||||
| Makaha'a; Monuafe and Velitoa | 21° 07' 12"S 1750 07' 99"W to 21° 05' 06"S 175° 09' 23"W | Mixed algal patches w/sparse Thalassia, coral outcrops and sand | Sand and reef patches with large areas of rubble. | 1–2m | A single area to the SE was seen. Large expanses of grasses and dense mixed algae. Locality needs further study. | |
| Mokotau; 'Onevai; Onevao Fig. 6a,b | 21° 05' 22"S 175° 06' 90"W to 21° 05' 70"S 175° 05' 65"W | Mixed algal expanses to the north and seagrass and algae to the southeast(Fig. 6a) | Sand | 1–2m | ||
| Cladosiph site | 21° 05' 47"S 175° 05' 25"W | Seagrass on sand and rubble | 2m | |||
| Ha'apai Group: General descrip | Seven barrier islands extend from north to south creating the semi-protected area conducive to the occurrence of seagrasses and Cladosiphon. Seagrasses only occur extensively in Felemea Bay, 'Uiha I. A smaller expanse occurs off Muikuku on Lifuka I. Elsewhere the protected shores are largely sand beaches with fringing and patch reefs off shore. | |||||
| Nukunamu I. | 19° 43' 80.1"S 174° 16' 95.5"W | Channel and back reef area. | Sandy areas withshall reef patches. | - | Fringing reef encirclesislet. Prominent sand bar to the west. No seagrass. Shallow reef patches in channel between Foa I. | |
| Foa I. | 19° 43' 00"S174° 16'80"W to 19° 45' 60"S 174° 25' 00"W | Projections of fringing reef with vertical face and patches (Fig. 7n). | Coral reef with deeper sandy areas offshore. | No seagrasses | ||
| Lifuka I. | 190 45' 90"S 174° 20' 30"W to 19° 50' 00"S 174° 23' 00"W | Fringing reef broken by sandy areas and new port. | Sand rubble and reef patches | 1–2 | 5 | Two small areas of seagrass. Cladosiphon at Muikuku. |
| Muikuku, Lifuka I. | 19° 46' 40"S 174° 21' 15"W | Reef flat extending to the light off Muikuku Pt. Seagrasses on the flat and inshore. | Sandy inshore and on flat. Reef patches offshore | 1–2 | 5 | Cladosiphon present (.55kg/m2) Area limited |
| Uoleva I. | 19° 50' 10"S 174° 23' 60"W to 190 51' 40"S 1740 25' 90"W | General sandy island margin. Extensive area of Enteromorpha. No seagrass. | Sand | 1–7 | 5 | Cladosiphon absent |
| Tatafa I. | 190 52' 20"S 1740 25' 70"W to 190 52' 90"S 1740 25' 00"W | Seagrass inshore, largely Thalassia but Syringodium present. Rubble and reef patches offshore. | Sand and rubble | 1–6 | 15 | Cladosiphon not present perhaps due to the exceptionally warm channel water flowing off reef flat north of 'Uiha. |
| 'Uiha I .Fig. 4a,b | 190 53' 00"S 1740 23' 80"W to 190 55' 35"S 1740 25' 60"W | Large areas of Syringodium seagrass | Sand patches within the beds and seaward | 2 | 5 | Abundance of Cladosiphon in Felemea Bay. |
| Felemea Bay, 'Uiha I. Fig. 4a,b | Site 1: 190 53' 30"S 1740 25' 10"W Site 2: 190 54' 20"S 1740 25' 19"W | Seagrasses in large patches (Table 2; Figs. 4a, 7h) | - | West wind removed or bundled up Cladosiphon See appendix 1 for weight measure/400m2. | ||
| Vava'u Group: General descript Fig. 5, 5a | No Cladosiphon was seen in the Vava'u Group, though there were reports of its existence earlier in the year. The presence of seagrass beds are limited in extent by comparison with Tongatapu I. Survey of known areas of their occurrence revealed beds of limited extent. In only one area was Syringodium present. Behind the barrier or southern islands visited the occurrence of the seagrasses were limited and largely composed of Cladosiphon sp. By and large the limitation of the seagrasses and the potential for Cladosiphon colonisation is limited by both exposure and the proximity of greater depths which reduce the area of likely seagrass occurrence and encourage the growth of fringing reefs though their extent is limited by the depth. Tidal exposure of much of the area which has suitable sediments makes for an inhospitable intertidal environment. | |||||
| Kiato and Okoa Islands | 18° 39' 00"S 173° 57' 10"W to 18° 39' 65"S 173° 57' 60"W | Thalassia and Syringodium meadows. Large areas of Sargassum with substantial epiphytic growth. | Sand and rubble | 1–2 | - | No Cladosiphon observed. |
| Kaloa Island: embayment to the west and north | 18° 38' 13"S 173° 55' 82"W to 18° 38' 51"S 173° 57' 90"W | Thalassia beds and algal expanses. Largely intertidal except in the channel areas. Mangrove areas. | Sand and muds | Largely inter-tidal | - | " |
| Kaloa Island: the south and east | 18° 38' 51"S 173° 57' 90"W to 18° 39' 29"S 173° 55' 90"W | Thalassia beds on the sand flat and slope along the channel which drains the embayment (Fig. 7o). | Sand | 1–4 | - | No Cladosiphon present |
| Umuna I. | 18° 41' 30"S 173° 55' 60"W | Sandy back reef hosting a variety of algaes. Patchy of varying density. | " | |||
| Kenutu I. | 18° 42' 00"S 173° 55' 70"W | As above but with Thalassia bed at the southern end. | Sand | 1–2 | " | |
| Oloua, Mafana Is. and Leka Islets, Ofu I | 18° 40' 00"S to 18° 42' 60"S 173° 57' 45"W | Limited Thalassia beds on NW Ofa I. The remainder of the islands fringing reef. | Sand | 1–4 | " | |
| Tapana I. | 18° 42' 80"S 173° 59' 20"W | Fringing reef environment | Sand and rubble | 1–4 | " | |
| Lautala I. | 18° 43' 30"S 173° 59' 80"W | " | " | " | " | |
| Tauta I. | 18° 44' 60"S 174° 00' 20"W | " | " | " | " | |
| Taunga, Pau and Ngau Islands | 18° 44' 80"S 174° 01' 00"W | Seagrass beds (Halodule and Thalassia sp.) on the western reef flat. Large areas Enteromorpha at 3–5 m on the eastern margin. | Sandy margin with rubble with general fringing reef | 1–4 | - | No Cladosiphon observed. Seagrass beds too sparse with a higher component of Halodule. |
| Sand on the reef flat | Reef flat inter-tidal | |||||
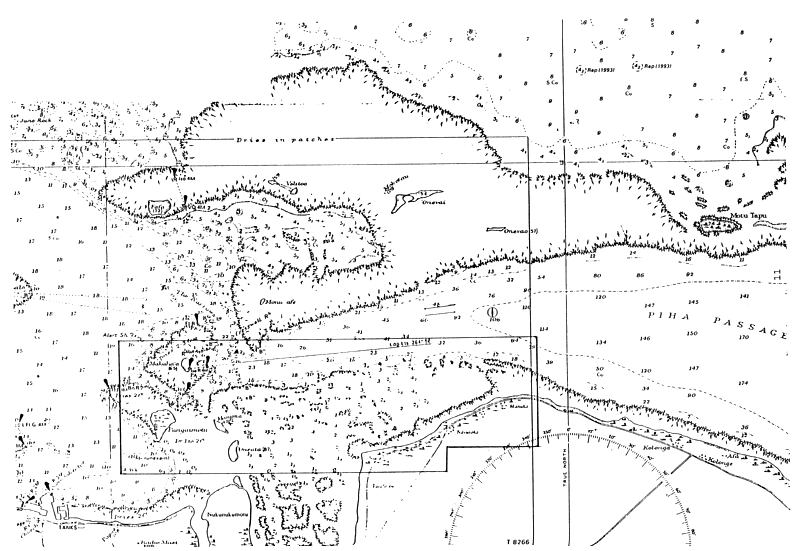
 | Figure 2a: The area of initial harvest on Tongatapu off Navutoka (located to the right in the photo). Another site of harvest is to the right of Manima and Oneata Is. See Fig. 2b for the charted area |
| Figure 2a (overleaf): Areas outlined are potential Cladosiphon habitats. In the Navutoka area, the greatest abundance is along the seaward margin of the zones defined. The inshore margin is not colonised due to intertidal exposure and the exclusive presence of the seagrass Thalassia sp. |
Figure 2b: Charted area of Fig. 2a shown in the bold lined section.
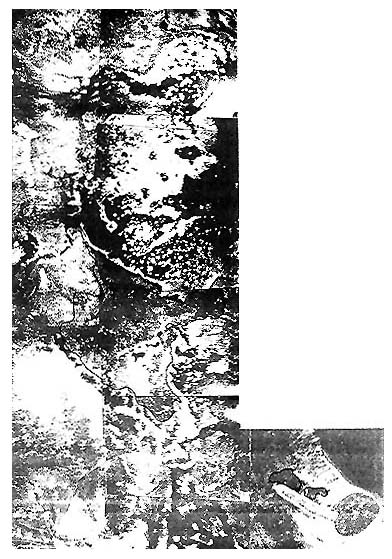 | Figure 3a: Tonga's most extensive Cladosiphon habited in the bay south of Atata and Toke Is. See Fig. 3b for detail of the charted area |
| Figure 3a (overleaf): Areas outlined are potential Cladosiphon habitats. Some of the densest concentrations were found adjacent to the southern margin of Maria Bay. Further survey will provide a more complete understanding of the areal and quantitative extent of colonisation. |
Figure 3b: Charted area of Fig. 3a shown in the bold lined section.
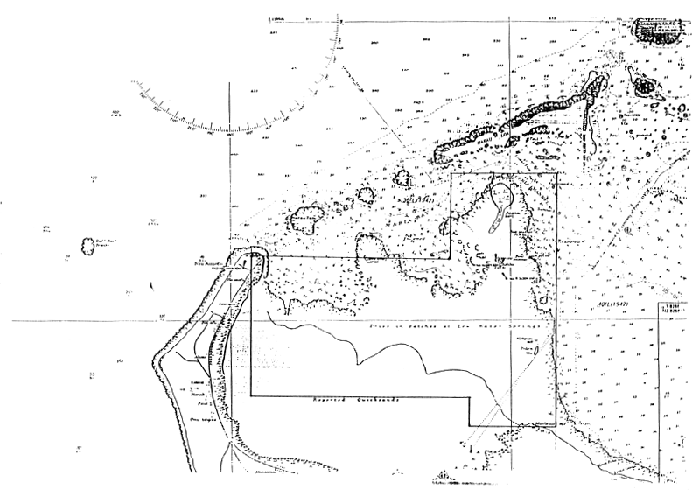
 | Figure 4a: The seagrass beds adjacent to Uiha I., Ha'apai which potentially host commercial quantities of Cladosiphon Sp The charted area of Uiha and adjacent islands are shown in Fig. 4b |
Figure 4b: Charted area of Fig. 4a shown in the bold lined box in the center of the page.
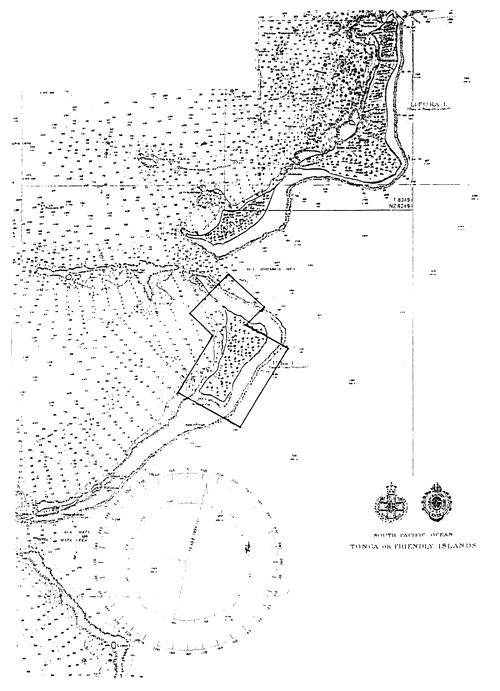
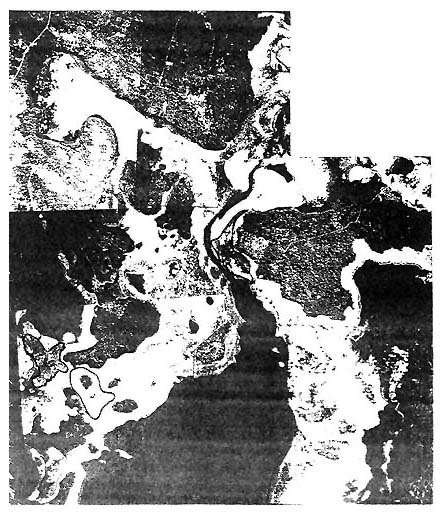 | Fig. 5: Aerial view of reported Cladosiphon habitats in the seagrass beds west of Koloa I. and north of Oloua I. See Fig. 5a for the charted details of the area. |
Figure 5 (overleaf): Areas outlined are potential Cladosiphon habitats. These areas were inspected as the result of anecdotal information on the algaes occurrence.
Figure 5b: Charted area of Fig. 5 shown in the bold lined box in the center of the page.
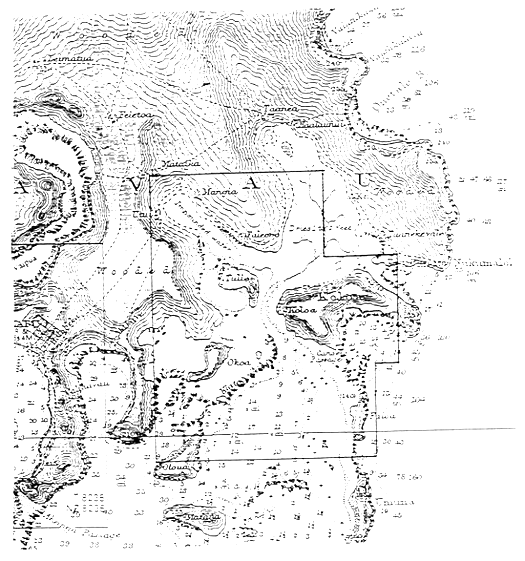
Fig. 6(a-c): Reported Cladosiphon habitats from off-shore reef areas:

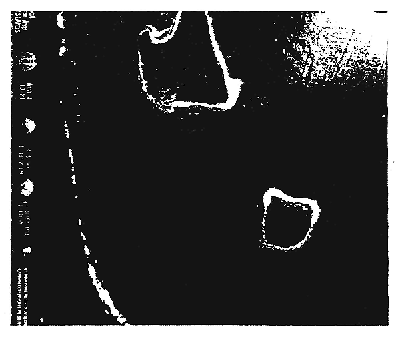
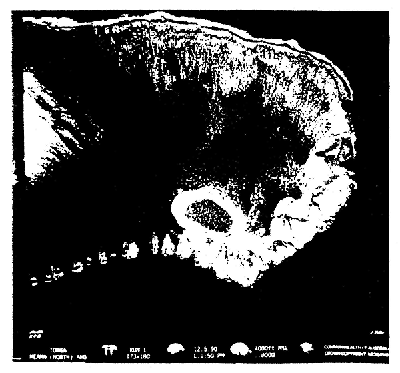
Figure 6d: Areas shown in Fig. 6 a,b on the barrier reef north of Tongatapu I.
Seagrass meadows can generally be considered a precursor to the occurrence of Cladosiphon colonisation and growth. The reason for this is that they have similar ecological requirements which include current washed reef flats where illumination is high. The reason why the Syringodium sp. are almost exclusively colonised is that they are stiffer than the Thalassia sp. and offer firmer support for the Cladosiphon colonies in what are semi-protected environments subject to minor wave action. As has been mentioned, the shape of the leaf may be important in terms of cutting or abrasion.
Syringodium sp. generally occurs on the outer portion of the reef flat which is generally subtidal. Thalassia sp. does can co-occur with Syringodium subtidally but is exclusively found in the intertidal area particularly near shore.
Due to the mode of dispersal which involves fragmentation of the Cladosiphon filaments and the transport by current, other benthos may act as a focus for the entanglement of the filament and subsequent peripheral colonisation. Examples are pieces of coral or upright Halimeda sp. or other stiff, erect algaes. The reason why these are not common sites of Cladosiphon proliferation is that colonisation requires a degree of support that is evident in the seagrass. Here the algae is able to progressively develop into a thick mat of material during its annual cycle. Areas of extensive seagrass meadows may have extensive Cladosiphon growth. With less support, the filaments fragment as they grow and are carried away by the current.
Table 2 is compiled to provide some degree of quantification and background for a general discussion of the probable extent of the resource. It is based on data limited by time and logistical constraints. Despite this, it represents a first step in sampling the standing stock to obtain some idea of quantity and attempt comparison of habitats on the basis of Cladosiphon abundance.
Other qualifications that influence the estimate include the lateness in the cycle of vegetative growth in which there is an expected seasonal reduction in the weight per metre. There is uncertainty as to the nature of the material measured as to whether it has been reduced by previous harvest, represent regrowth or is natural in abundance. Biomass will vary with season and be affected by weather both occasionally and seasonally. Sampling should be complete enough to account for patchiness in the distribution of the algae.
Additionally, the seagrass beds can, with reasonable reliability, be determined from the aerial photos. The accuracy of the information on the presence and type of the seagrass beds needs to be improved by ground truthing but serves, at this point, in providing a semi-quantitative comparison of habitats.
Knowing how much Cladosiphon is in the field will allow better management of the resource. Adequate employment and preparing the facilities for processing is part of maximising the resource. The resource could be managed in a more efficient manner with goals more realistically set and a better appreciation of the time table required.
About Table 2: Estimates of the potential for Cladosiphon harvest from this table must remain speculative pending further field assessment of the degree of Cladosiphon colonisation. The material presented in this table represents a summary of the information to date. Following are clarifications of the table headings.
Locations: Unlike Table 1, the location refers to areas defined on the aerial photographs and associated charts. They contain the sample sites more precisely detailed in Table 1.
Estimated area of seagrass habitat (ha): These areas were determined by inspection of the aerial photographs and comparison with the field notes. Unfortunately the photographs were obtained at the end of the field work so that confirmation was not possible. Good confidence exits in delimiting most areas. Areas in question are noted in the comments.
Table 2: Comparison of Cladosiphon Habitats
| Location | Estimate of seagrass habitat (ha) | Colonised habitat (ha) | Harvest to Dec. 1996 (tons) | *Field sample (kg/m2) | Comments: |
| Navutoka Coast Manuka to Niutao Pt. | 41.39 | 10.34 (25%) | 10 | .28–.47 (.55–.95) | Area of intense harvest earlier in the year. Abundant material which may be residual or regrowth. |
| Niutoka Pt. to Nukuleka | 3.82 | - | 0 | - | This area was reported to host Cladosiphon (Billings, 1996). None was observed in the brief survey though there was a dense algal assemblage. |
| Manimal. Oneata I. and Nukunuku Motu I. | 6.91–11.4 | 1.73–285 | 440 kg | - | Manta tow survey revealed only Thalassia seagrasses. As material was harvested from this area it is presumed the Cladosiphon occurs on the eastern side. Likely areas are evident in the aerial photos. |
| Onevai and Onevaols. | 54.82 | 13.70 | 0 | - | Dense mixed algal assemblages on the north side though no Cladosiphon seen. Area of occurrence between the two islands and most probably extends to Onevai I. |
| Large western bay area - Tongatapu L 14 est. | |||||
| 1) Southwest of Atatal. | 11.12 | 2.78 | - | .43 (.85) | Good seagrass development with Cladosiphon occurring in the outer seagrass area. |
| 2) West of Poloa and Alakipeau Is to Muifonua Pt. | 332.69 | 33.37 (10%) | - | - | Cladosiphon occurrence was reported to have occurred (Billings, 1996). Survey of the area in the northwest found mainly Thalassia beds and mixed algae predominantly Padina and Dictyota but no Cladosiphon.. |
| 3) Southeast of Toke I. | 278.11 | 69.53 | - | 1.25 (2.50) | |
| 4) Southwest of Toke Toke I. | 93.57 | 9.36 (10%) | - | - | No Cladosiphon seen. Thalassia beds and intermittent sand and coral reef areas. |
| 5) Far western bay adjacent to mainland. | 38.99 | - | - | - | Hillocky sandy terrain with Syringodium patches on the mounds |
| Felemea Bay, 'Uiha I., Ha'apai | 1) 7.53 2) 46.78 3) 727 4) 8.26 Sum 69.84 | 17.45 | 0 | 18.33kg/400m2 | Westeny winds had removed the Cladosiphon from the seagrass. |
| Muikuku, Lifuka I. Ha'apai | <2 | - | 0 | .25 (.5) | |
| Kiato I., Vava'u | 1.72 | - | 0 | 0 | |
| Koloa I., Vava'u | 6.50 | - | 0 | 0 | |
| Kenutu I. | <1 | - | 0 | 0 | |
| Taunga I. | <1 | - | 0 | 0 | |
The figures are estimates maximum potential habitat. Extrapolation of an accurate figure of actual colonisation could only be realised by further sampling.
Potential Cladosiphon colonisation (ha): This figure is arbitrarily set at 25% of the area estimated for the seagrass meadows. From general inspection during the field work phase, this seems like a reasonable approximation. To determine the variability in this figure, additional sampling would be required. Its usefulness is in obtaining some idea of the biomass that is likely to occur in a particular area. An estimate of 20% colonisation of available seagrass habitat may be more realistic (pers. comm. Fa'anunu).
A accurate figure would have to account for the patchiness in both the seagrass beds and the algae within the estimated area. Some areas in the beds are unsuitable for colonisation due to intertidal exposure or land effects such as fluctuations in temperature and sedimentation.
Quantity of harvested to December 1996 (tons): This is the material that was harvested and continued to be through the 1996 season.
Field sample (kg/m2): One square metre plots were sampled and weighed as a measure of the algae present in the field. In the case of Felemea Bay. 'Uiha the material was estimated by assessing the clumped algae over 100m manta tow observing the material within a 4m wide band.
Figure 7(a-o): Cladosiphon lifecycle, habitat and experimental design.
(a,b) Dispersal of Cladosiphon involves two modes:
Intertidal seagrasses adjacent to Koloa I, Vava'u Group. In deeper water, this habitat is reported to host the Cladosiphon algae.
In the field situation, the samples in both the field and experimental sample exhibited similar characteristics. They both showed a diminished vitality. The wild material had been observed 10 days earlier as entangled in the seagrass with some of the specimens showing good colour and smooth surface texture. Some matting was evident. The rope culture sample were selected on the basis of vitality. Recent inspection revealed the wild and rope samples exhibiting a decline, having similar characteristics in appearance. They appeared old with the fronds less vital in appearance than those observed in earlier periods. The algal mass was coated to a greater degree with sediment. The colour had changed from a golden brown to a dull brown with the mucilaginous sheath less evident or absent. Similarly in the tray culture, the same features were evident as a degraded appearance.
Figure 7 (a-o): Cladosiphon lifecycle. habitat and experimental design.
| a) |  | e) |  |
| b) |  | f) |  |
| c) |  | g) |  |
| d) |  | h) |  |
| i) |  | m) |  |
| j) |  | n) |  |
| k) |  | o) |  |
| l) |  |
![]()
![]()
![]()