![]()
![]()
![]()
Genetics provides the basis for developing breeding programs aimed at improving productivity and fitness of yak. However, quantitative genetic characterization is largely absent for the yak because the prerequisite of recording pedigrees and performance has not been widely practised in relation to the yak population. This is due in part to the remoteness and harsh conditions of most of the yak territories, but also because, until now, yak keeping has been more of a way of life than a strictly economic activity.
The techniques of cytogenetics and, more recently, of molecular genetics, which have been used widely to characterize other farm livestock and explore the basis of their heredity, are being applied in the yak, though not, as yet, to the same extent. Much of this work on other species of cattle is, however, proving relevant to yak. Thus, the identification of quantitative trait loci (QTL) and the exploration of marker-assisted selection in cattle may also provide a basis for improving the yak.
These studies in the yak are also providing evidence of the extent of breed differentiation that, in turn, is relevant for conservation policies. Further, the cytogenetics and, more importantly, the molecular information are providing evidence of the relationship of the yak to other cattle species. In this context, the new evidence supports the view that the yak should be assigned to the subgenus Poephagus and not to the more widely adopted Bos. This chapter reviews the studies in these areas with yak.
The aim of yak breeding, as for other livestock, is to improve productivity or to affect other desirable changes. For some traits it is easy to bring about changes because the traits are relatively simply inherited and controlled by a very small number of genes; for example, most coat colours or the presence or absence of horns. Moreover, for such so-called qualitative characteristics, the inherited component is the major and usually the only component responsible for variation in the trait across a population. By contrast, the so-called quantitative traits (variation in quantity rather than quality of the trait), represented by most aspects of performance, such as milk and meat production, reproductive rate, health and so on, are also affected to a greater or lesser extent by the environment.
Thus, even if the same complement of genes for growth rate were present in two groups of animals, those that were better fed would grow better. For this reason it is difficult to differentiate between the extent of genetic and non-genetic influences, unless very precise rules are followed in making the comparisons and there are good records of both parentage and performance. The important point is that for the vast majority of traits of the yak, the individual's phenotype is not equivalent to its genotype. However, under uniform conditions for contemporary animals, the better performing will generally be genetically more desirable than less-well performing contemporary individuals. It is by this means that, over generations, livestock breeders have been able to make changes and improvements to their livestock, although as discussed elsewhere in this book, individual yak productivity is, at best, only one of the criteria considered by yak herders in maintaining their herds.
The way to effect genetic change, generally intended to produce some improvement, is to utilize identifiable genetic variation in a herd or larger grouping within a breed by means of selection, or to use differences among breeds (or, for example, between domestic and wild yak) by a process of crossbreeding and, finally, and of particular relevance for the yak, to make use of differences between the yak and other species of cattle by way of hybridization. Each of these routines has its rules and particular consequences, reviewed in relation to yak by Wiener (1994, 1997). Particularly for purposes of selection, the conditions for identifying genetic variation cannot reasonably be met for the great majority of the yak under the normally extensive management, although Goe and Stranzinger (2002) recently proposed a recording scheme they regard as practical under these conditions. Even on the few government or research farms where the potential ought to exist for accurate and sustained recording of parentage and performance, a prerequisite for accurate selection, these opportunities for selection do not appear to have been much exploited (with at least one honourable exception). In consequence, yak have been left out of the revolution in breeding practice of most of the other major livestock species - particularly those kept relatively intensively like poultry, pigs and dairy cattle. Nonetheless, a number of breed development schemes for yak, run by herders, have existed for a long time and some more recent schemes have led to the development of a new breed of yak - based on a cross of domestic and wild yak. These matters are described in Chapter 2 and referred to again in the concluding chapter.
With regard to crossbreeding and to hybridization involving yak, the nature of the genetic reasons for changes in performance, heterosis in particular, have not been quantified because the means for doing so are not usually present (see Chapter 3). Claims for the degrees of heterosis generated by crossbreeding or hybridization with yak will need verification.
New methodologies are emerging, in particular those of molecular genetics and cytogenetics. These aim, in the case of molecular genetics, to identify genetic variation directly from differences within the sequences of the DNA (deoxyribonucleic acid), which is the material of chromosomes on which the genes are located. The cytogenetic approach, which has a longer history, attempts to identify variation in chromosomal structures as seen in preparations under a microscope. The ultimate hope is that such variation, close to the gene or actually involving the gene, can be linked to the differences in fitness and performance in the population or even at an individual animal level. Because such techniques, though complex and demanding, do not depend in the first instance on large-scale herd recording, they provide a challenge for geneticists working with yak as they do for those concerned with other species. A significant start has been made in these areas with yak; the rest of this chapter will provide recent results and, where necessary, some background to the techniques.
As already noted (Chapter 2), yak with 30 chromosome pairs have the same number as other cattle, and domestic and wild yak have similar complements. Li Jiyou et al. (1994), Han Jianlin (1996), Zhong Jingcheng (1996) and Nivsarkar et al. (1997) reviewed the progress of research on chromosomes of the Chinese and Indian yak in comparison with Bos taurus cattle and their hybrids with yak. Zhong Jingcheng found that the structure and size of all 29 pairs of autosomes and the X-chromosome are very similar among five Chinese breeds but that the Y-chromosome shows differences in terms of its relative length and structure (see Table 15.1). However, these differences are small and cannot be taken as markers for distinguishing the species of the yak from either Bos taurus or Bos indicus cattle (Gupta et al., 1996) or to distinguish the yak breeds from each other (Han Jianlin, 1996).
Halnan (1989) and Zhong Jingcheng (1996) published some detailed G- and C-banded karyotypes of the yak in order to investigate genetic differentiation among the yak breeds. After comparing the C-banded patterns and Ag-NORs between the Maiwa and Jiulong yak, Zhong Jingcheng (1996, 1997) concluded that the visible differences in heterochromatin quantity and distribution between these two breeds were not significant, as similar differences occurred among the individuals within a breed. In spite of proposals for standardization of karyotyping across laboratories, no reliable data are available for use as genetic markers on the chromosome structures of different yak breeds (Han Jianlin, 1996).
Table 15.1 X- and Y-chromosome polymorphisms in yak* [Source: Zhong Jingcheng, 1996; Nivsarkar et al., 1997]
|
Population |
X-chromosome |
Y-chromosome |
||
|
Relative length |
Arm ratio |
Relative length |
Arm ratio |
|
|
Maiwa |
5.11 |
1.90 (SM) |
1.89 |
2.67 (SM) |
|
Jiulong |
4.87 |
1.77 (SM) |
1.71 |
2.60 (SM) |
|
Tibet |
4.59 |
2.08 (SM) |
2.08 |
(SM) |
|
Huanhu |
4.72 |
1.89 (SM) |
2.36 |
2.46 (SM) |
|
Zhongdian |
4.80 |
1.86 (SM) |
2.56 |
1.87 (SM) |
|
Kyrgyz |
5.78 |
2.32 (SM) |
|
(SM) |
|
Indian |
4.95 |
(SM) |
2.44 |
(M) |
|
Wild yak |
4.96 |
1.96 (SM) |
1.57 |
2.31 (SM) |
* Note: Relative length = length of individual chromosome/total length of the haploid with X-chromosome included; arm ratio = length of short arm/length of long arm, with reference to a nomenclature of chromosome, when the arm ratio is between 1 and 1.7, the chromosome is referred to as metacentric (M), between 1.7 and 3 as submetacentric (SM), between 3 and 7 as subtelocentric and beyond 7 it is called telocentric.
A few studies (cf. Chapter 7) have tried to link the differences in the sex chromosomes between the yak and Bos taurus or Bos indicus cattle to the sterility of the F1 male hybrids (Guo Aipu, 1983). However, this seemed to be an unlikely approach for resolving the problem because crosses of Bos taurus and Bos indicus produce fertile male offspring in spite of there being differences in the Y-chromosome between these two cattle species (Nivsarkar et al., 1997).
The similarities between the yak and cattle chromosome structures and the relevant linkage maps, may have one beneficial consequence for yak, if indeed it can be shown that cattle provide an adequate model for yak in respect of quantitative trait loci (QTL), which is referred to more fully in ensuing sections (for a general description, see Fries and Popescu, 1999).
One of the aims of the molecular technology, which is discussed later, is to provide genetic markers that may be related to performance traits or to health (or disease) and which might also provide information about relationships among the animals examined. The search for such markers in blood and in milk has a much longer history, and before discussing the molecular technology, it is useful to draw attention to the biochemical marker work with yak.
Zhang Yuezhou et al. (1994), Zhong Jingcheng (1996, 1997) and Nivsarkar et al. (1997) reviewed the advances in yak genetic diversity in terms of blood and milk protein and enzyme polymorphisms used to estimate the genetic differentiation and relationship among the yak breeds. Han Jianlin (1996) and Zhang Caijun (1997) noted that in these investigations there were a total of 54 genetic loci consisting of 14 protein or enzyme loci detected in plasma or serum, 35 proteins or enzymes in erythrocytes and five milk protein loci. As reported by these authors, the methods used in different laboratories for these determinations, as well as the nomenclature employed in description, were not uniform. Moreover, sampling from different breeds of yak was mostly rather limited. For these reasons, comparisons across investigations cannot readily be made and these studies have contributed less to the genetic characterization of yak populations than had been hoped. Most importantly as noted by Han Jianlin (1996) and Zhang Caijun (1997), only a few of these genetic markers showed polymorphisms. For example, nine out of the 41 loci studied, or only 24.5 percent of the proteins or enzymes in the blood of yak, were found to be polymorphic, which is a far lower proportion than that found in the ordinary cattle (Hines, 1999). Consequently, these studies have provided relatively little information by which to differentiate the genetic relationships among yak breeds.
In another comparative study conducted in one laboratory with more loci involved, Tu Zhengchao et al. (1997) reported that in the Datong, Tianzhu White, Bazhou, Zhongdian, Maiwa and Jiulong yak (5 - 56 samples), only four out of the 34 blood protein and enzyme loci were polymorphic. The percentage of polymorphic loci was 11.8 percent, and the mean individual heterozygosity across the breeds calculated to be only 0.015. This suggests a very low level of genetic diversity in the Chinese yak breeds. The average genetic distance among the breeds was estimated as 0.0006 and the average coefficient of gene differentiation across the breeds as 0.0313. This implies that at the level of blood protein polymorphisms only 3 percent of the nuclear genetic constitutions detected in these six yak breeds could be attributed to the differences between the breeds. The rest (97 percent) was most likely to be due to variation within breeds. They suggested that such a low genetic diversity in these six Chinese yak breeds may have resulted from one or more severe bottlenecks due to diseases or natural disasters in the yak population.
More recently, Tao Shixin and Han Jianlin (2002) used 269 samples from six pure yak populations in Gansu and Qinghai provinces and confirmed that there are no polymorphisms at the Hb loci. Qi Xuebin and Han Jianlin (2002) reported that in 608 yak-milk samples collected from six geographically different yak production areas in Gansu province, the average genetic differentiation between populations at the beta-lactoglobulin locus was only 8.4 percent. The authors suggested that this might have resulted from introgression of cattle genes into the yak population studied, as hybridization is practised in some of the areas from which the yak were sampled.
Rozhkov and Galimov (1990) found, by affinity electrophoresis in the saliva of yak, a new AMY-1 allelic variant (AMY-1D). The yak was found to be monomorphic for this allele while other cattle types and hybrids studied by these authors had other alleles. Thus, zebu cattle in the northern Caucasus had AMY-1B and AMY-1D alleles, Red Steppen cattle in the Caucasus and Holmogor cattle from the Moscow region had AMY-1A and AMY-1B alleles while Kazakh White Head cattle in Altai only showed the AMY-1B allele. Hybrids produced from yak bulls and Bos taurus cows had both the AMY-1B and AMY-1D alleles, while hybrids of Bos indicus and Bos taurus cattle had all three alleles (A, B and D). This suggests that this locus can be useful in investigating introgression between the yak and other cattle species.
DNA-based studies
Recent studies have focused on variation at the level of the DNA sequence. DNA has the advantage of allowing the development of relatively robust tests. It can be sampled from almost any tissue and a very large number of different markers can be developed based on DNA sequence variation. To put yak into perspective in relation to the hugely studied human genome, the latter has a genome size of 3 300 mbp, while that of cattle and yak is estimated to be 3 000 mbp in size.
Not all DNA consists of genes. There are two types of DNA in a cell: the nuclear DNA and the mitochondrial DNA (mt-DNA). Nuclear DNA is responsible for most of the physical characteristics of the yak, and a nuclear DNA profile normally points exclusively to a specific yak. The most common type of nuclear DNA consists of unique, or single-copy sequences, which account for about 57 percent of total DNA in cattle (and yak are assumed to be similar). Other DNA is called satellite DNA, which is sufficiently different in density from the major portion of DNA to enable it to form recognizably separate peaks or "satellites" when the total DNA is subjected to certain density gradient tests. Satellite DNA is not dispersed throughout the genome; instead it is located at specific sites in the genome. Cattle DNA contains eight different satellites, which together constitute about 23 percent of the nuclear DNA of cattle. The remaining 20 percent of DNA is characterized by repeat sequences and is called interspersed-repetitive DNA. Vaiman (1999) pointed out that at least 50 percent of the cattle genome consists of repetitive sequences.
These estimated proportions of the three categories of cattle nuclear DNA derive from work in the late 1980s. It is worth noting that these could be subject of drastic revision if recent human genome data provide a clue. Earlier estimates of the proportion of single-copy DNA for the human were around 60 percent - not dissimilar to the cattle values. However, recent revision suggests that this proportion in humans is only around 25 percent, or has been reduced from about 100 000 (Hwu et al., 1986) to 30 000 - 40 000 protein-coding genes (International Human Genome Sequencing Consortium, 2001).
Mitochondrial DNA (mt-DNA
Under a microscope, yak cells, like the other eukaryotic cells, appear to contain hundreds of smaller organelles within the cytoplasm of the cell. Many of these smaller structures are the mitochondria, each containing its own DNA. The mitochondrial genome is much smaller than the nuclear genome in terms of the DNA quantity - there are only 16 338 base pairs (bp) in cattle, a tiny fraction of the amount of nuclear DNA (and yak are thought to be similar) (Anderson et al., 1982). Its structure is also relatively simple. Mammalian mt-DNA is very compact and simple compared to the nuclear DNA. Mitochondria have essential functions in the energy metabolism of cells; but for purposes of this chapter it is not their function that is of primary interest but the fact that their DNA profile can be used to differentiate individual animals.
The genetics of mt-DNA differs from that of nuclear DNA in several important ways: The most important feature that distinguishes mt-DNA from nuclear DNA in the present context is its maternal inheritance. The egg harbours several hundred thousands mt-DNA from the mother, while the sperm has only a few hundred mt-DNA. Thus, the few sperm mt-DNA that enter the egg have almost no effect on the mitochondrial genotype. Therefore, a yak mitochondrial DNA profile would be identical to his/her mother's and all of his/her maternal relatives. This means that the mt-DNA profile of a yak is not unique to that individual but rather points to his/her maternal lineage.
Several molecular approaches have been applied to investigate the yak's genetic markers in both mt-DNA and nuclear DNA. These are RFLPs (restriction fragment length polymorphisms), microsatellites and VNTRs (variable number tandem repeats), SSCPs (single strand conformation polymorphisms) and SNPs (single nucleotide polymorphisms). They are used in terms of understanding relationships between breeds within the yak species and among various cattle species, as a potential aid to breeding.
RFLP is a technique in which organisms may be differentiated by analysis of patterns derived from cleavage of their DNA at a precisely defined length, such as the whole mt-DNA or a specific DNA fragment amplified through a polymerase chain reaction (PCR). If two organisms differ in the distance between sites of cleavage of a particular restriction endonuclease, the length of the fragments produced will differ when the DNA is digested with a restriction enzyme. The similarity of the patterns generated can be used to differentiate species (and even yak breeds) from one another.
Polymorphisms detected by RFLP are inherited differences found among the individuals in a population. Yak mt-DNA RFLPs have been employed as a tool to demonstrate relationships between and within breeds of the yak (Zhao Xinbo et al., 1994; Tu Zhengchao et al., 1998). In particular, Tu Zhengchao et al. (1998) using 20 endonucleases and 90 samples collected from Jiulong (20 samples), Maiwa (20), Zhongdian (21), Huanhu (20) and Tianzhu White (9) yak breeds found that six out of the 20 enzymes detected polymorphisms with five haplotypes (I to V) across a total of 56 digestion sites, of which the haplotype II, III and IV were derived from the haplotype I by changing the digestion sites and V subsequently from the II, respectively. The divergence times among these five haplotypes are from 0.045 - 1.295 million years ago, among which the haplotype I is thought to have separated from III 0.635 million years ago, I from II 0.725 million years ago and III from II 1.295 million years ago. The average genetic diversity (HT) is 0.1065 across the five breeds and the average net genetic distance between either two of the five breeds is 0.000201 (see Table 15.2). The gene differentiation coefficient is only 0.0291, or only 3 percent of the total population differentiation is attributed to the genetic variation among the populations. This indicates a very low genetic differentiation among the yak breeds studied and is similar to the conclusion drawn from protein and enzyme loci (see previous reference). Further clustering of the Jiulong and Huanhu yak into one group and of the Zhongdian and Tianzhu White yak into another group are not in agreement with the knowledge from historical origin and geographic distribution, possibly due to limited and random mt-DNA RFLPs and eventually limited genetic differentiation in yak.
Table 15.2 The nucleotide diversity (upper right) and net nucleotide divergence (left down) between the yak breeds [Source: Tu Zhengchao et al., 1998]
|
Breeds |
No. samples |
Zhongdian |
Jiulong |
Maiwa |
Tianzhu White |
Huanhu |
|
Zhongdian |
21 |
|
0.003492 |
0.003252 |
0.002460 |
0.003715 |
|
Jiulong |
20 |
0.000352 |
|
0.004008 |
0.003425 |
0.004293 |
|
Maiwa |
20 |
0.000297 |
0.000151 |
|
0.003282 |
0.004224 |
|
Tianzhu |
9 |
0.000112 |
0.000139 |
0.000181 |
|
0.003650 |
|
Huanhu |
20 |
0.000398 |
0.000038 |
0.000154 |
0.000187 |
|
Sulimova et al. (1996) using PCR-RFLP investigated the polymorphisms of the 5'-untranslated region and exon 4 of kappa-casein gene in yak and found a new variant in yak compared to other Bovini species.
mt-DNA markers for detecting the introgression of cattle into yak or vice versa. As stated in Chapter 7, the F1 male hybrids and the B1 to B3 male backcrosses from hybridization of yak mated with either Bos taurus or Bos indicus cattle are sterile. Therefore, the exploitation of heterosis thought to arise in these hybrids usually stops at the F1 generation. However, there is, in the field, a small amount of backcrossing of the fertile F1 females to yak bulls or to bulls of the other cattle species. This opens the possibility for introgression of cattle genes into the yak population (and vice versa).
To assess the level of cattle mt-DNA introgression in yak, Han Jianlin et al. (2002), using a cattle diagnostic mitochondrial DNA marker, found that cattle introgression through the female lineages is either absent or low in the five yak populations studied. Out of 239 animals, only one female in the Bhutanese population and two females in the Tianzhu White yak had the cattle mitochondrial DNA. Qi Xuebin et al. (2002a) confirmed that in a total of 963 samples from 24 yak populations there were 21 individuals from 13 yak populations carrying a cattle mt-DNA with the frequencies of cattle introgression ranging from 1.5 percent (Tianzhu White yak) to 10.6 percent (Tianzhu Black yak). In general, the introgression of cattle mt-DNA in yak is rare, and it is mostly limited to marginal and agro-pastoral areas where yak and cattle coexists. However, as these results do not take into account possible cattle introgression through the male lineage (in terms of the cattle nuclear DNA from the use of a cattle bull on a yak female at the first step in hybridizing), it is possible that they underestimated the level of cattle introgression in the yak populations.
In a separate study, Bailey et al. (2002) found that less than 1 percent cattle mt-DNA in a survey of yak mt-DNA types with samples from Nepal, Bhutan, China and Mongolia. They claimed that this is encouraging for those who are concerned about the effects of introgression of cattle into yak. It is important to note that mt-DNA can remain unperturbed even when high levels of introgression are observed at the nuclear level.
In a study of 11 native cattle breeds in South China using mt-DNA RFLPs based on ten restriction endonucleases' digestion, Yu et al. (1999) found that there was an introgression of mt-DNA, at a frequency of two out of seven samples analysed, from yak to a cattle breed (the Diqing cattle kept in northwestern Yunnan province where the Zhongdian yak are also found). They proposed that such an introgression might eventually contribute to the adaptation of the Diqing cattle to the cold weather and high altitude at 2 800 - 3 300 m.
AFLPs or its fluorescent version (fAFLP) are a DNA fingerprinting technique that detects DNA restriction fragments in the animal nuclear genome by means of PCR amplification. It requires no sequence information or probe collections prior to the generation of AFLP fingerprints. This is of particular benefit when studying organisms where very little DNA marker information is available.
Buntjer et al. (2002) included three yak in a phylogenetic study of bovine species based on AFLP fingerprinting and all samples produced relatively rich polymorphic markers (see section, Systematics).
Microsatellites (or SSRs, simple sequence repeats) are stable, polymorphic, easily analysed and occur regularly throughout an animal genome, making them especially suitable for genetic analysis. Microsatellites are co-dominant markers, so that all alleles can be scored. The availability of microsatellite markers located throughout the animal genome has also facilitated livestock genome scans for loci affecting quantitative production traits (QTL). Microsatellites are generally recognized as neutral so that selection and environmental pressure do not influence their expression directly.
To identify microsatellite markers from a eukaryotic genome is not as easy as using them. The isolation and characterization of microsatellites, their sequencing and the testing of primers can be time consuming and expensive. However, the conservative flanking sequences at every microsatellite marker allow the primers designed from the sequences to be used across closely related species. Since the late 1990s, Hishida et al. (1996), Ritz (1997), Hanotte et al. (2000), Wang Minqiang (2000) and Dorji et al. (2002) have tried the cross-species amplification of cattle autosomal microsatellite markers in yak and mostly succeeded in getting effective and polymorphic loci. Particularly, Ritz (1997) and Hanotte et al. (2000) using the same set of 31 to 39 yak samples collected from Bhutan found that 6 out of 8 and 19 out of 20 cattle microsatellite loci successfully amplified polymorphic yak microsatellite markers, respectively. Wang Minqiang (2000) tried 13 cattle microsatellites for 28 Datong yak and 20 Gannan yak and discovered both breeds showed polymorphic markers at 13 loci (see Table 15.3).
Using a set of eight cattle autosomal microsatellites, including TGLA53, TGLA122, TGLA73, AGLA293, BM2113, BM1824, CSSM066 and ETH3, Dorji et al. (2002) found that in a total of 169 yak sampled from three yak populations of western Bhutan (106), central Bhutan (32) and eastern Bhutan (31) all eight loci are polymorphic in the Bhutanese yak. They also found that there is substantial genetic variability within the populations, with average heterozygosity range of 0.644 - 0.680. A neighbour-joining tree constructed from Nei's standard genetic distances grouped western and central Bhutan yak in one clade separate from eastern Bhutan yak. The genetic distances between the yak from eastern Bhutan and from the other two regions suggest that the populations have been separated for at least 4 000 years and that they have exchanged less than two migrants per generation. Based on these results, Dorji et al. categorized the Bhutanese yak populations into two types of (1) western and central Bhutan yak and 2) eastern Bhutan yak (cf. Chapter 11, part 2).
Han Jianlin et al. (2002) also tried a set of cattle Y-chromosome-derived microsatellite loci and found that INRA124 failed in the amplification of a yak Y-chromosome locus and INRA 126 amplifies microsatellite from both sexes of yak samples.
Table 15.3 Cattle microsatellite loci used in cross-species amplification in yak and their polymorphisms
|
Ritz (1997) |
Hanotte et al. (2000) |
Wang minqiang (2000) |
|||
|
Bhutanese yak |
Bhutanese yak |
Datong yak and gannan yak |
|||
|
Loci |
Polymorphic |
Loci |
Polymorphic* |
Loci |
Polymorphic |
|
BM1824 |
5 |
BM1824 |
4 |
BM1824 |
5 |
|
ETH003 |
3 |
BM2113 |
4 |
BM2113 |
7 |
|
ETH010 |
5 |
ETH003 |
2 |
CSSM66 |
7 |
|
ETH121 |
7 |
ETH010 |
3 |
ETH152 |
7 |
|
ETH131 |
5 |
ETH225 |
4 |
ETH185 |
5.5 |
|
ETH225 |
5 |
ILST005 |
Failed |
ETH225 |
5 |
|
ILST005 |
Failed |
ILST006 |
3 |
HEL1 |
5.5 |
|
CYP21 |
11 |
SPS115 |
Monomorphic |
HEL5 |
4 |
|
MHCII |
8 |
|
|
HEL9 |
8 |
|
AGLA293 |
5 |
|
|
HEL13 |
4 |
|
MGTG7 |
7 |
|
|
INRA005 |
5 |
|
MGTG4B |
5 |
|
|
TGLA126 |
4 |
|
TGLA48 |
5 |
|
|
TGLA127 |
4 |
|
TGLA53 |
8 |
|
|
|
|
|
TGLA57 |
5 |
|
|
|
|
|
TGLA73 |
9 |
|
|
|
|
|
TGLA122 |
8 |
|
|
|
|
|
TGLA126 |
5 |
|
|
|
|
|
TGLA227 |
5 |
|
|
|
|
|
TGLA263 |
3 |
|
|
|
|
Note: Polymorphic is presented as the allele numbers detected at that loci.
Therefore, neither is suitable for the cross-species amplification yak in this domain. The two microsatellites, BM861 and INRA189, are Y-specific in both cattle and yak with the presence of Bos taurus, B. indicus and B. grunniens diagnostic alleles. This was confirmed by Qi Xuebin et al. (2002b) with 252 male yak samples. Both Han Jianlin et al. (2002) and Qi Xuebin et al. (2002b) found that INRA189 locus is a polymorphic marker in yak with three alleles, the monomorphic locus BM861 reported by Han Jianlin et al. (2002) in 83 male yak samples has been shown by Qi Xuebin et al. (2002b) to be polymorphic in 252 samples. Meanwhile, Qi Xuebin et al. (2002b) also added a third polymorphic Y-chromosome specific locus, BYM-1, in yak being detected with a cattle-derived microsatellite and discovered ten Y-specific microsatellite haplotypes based on the detected combinations among these three polymorphic loci in yak.
From these preliminary reports, it is clearly seen that the cattle microsatellites can be useful in characterizing yak genetic diversity. However, the loci involved in each experiment differ from each other, and the results therefore are not comparable. To efficiently explore the advantage of the these new technologies, the International Livestock Research Institute initiated a project on the genetic characterization of yak with microsatellite genotyping and DNA sequencing in 1999 in collaboration with scientists and institutions in Bhutan, China, India, Kyrgyzstan, Mongolia, Nepal, Pakistan and Russia. A total of about 1 000 samples from up to 30 yak breeds (or populations) have been collected from these countries.
VNTRs (minisatellites) are a short sequence of DNA repeated a number of times on some eukaryotic chromosomes. It is possible to cut out the segment of the chromosome containing the VNTRs, run the total DNA on a gel and identify the VNTRs by hybridization with a probe specific for the DNA sequence of the repeat. The number of repeated segments at a locus varies between individuals. Some VNTR sequence segments are found at only a single locus in the whole genome. Other VNTR sequence segments occur at many loci in the entire genome. These loci are dispersed among the chromosomes and images produced by multi-locus VNTR probes yield more information since one probe is simultaneously probing 10 - 30 loci. In the description above, RFLP is used to detect the variability in the number of repeats. Several PCR-based methods can also be used to detect VNTRs.
Nijman and Lenstra (2001) compared several distinct centromeric satellites, a complex of tandem repeats abundant mostly in eukaryotic genome, among bovine species with yak included. They found that the 1.711b sequence of yak resembles the bison and wisent sequence but in satellite IV yak shares several other heterogeneous nucleotide positions with gaur and banteng.
SSCP is a method for distinguishing between similar-sized DNA fragments according to the mobility of the single-stranded DNA under polyacrylamide gel electrophoresis. In combination with automated detection of fluorescently labelled PCR products, SSCP is a convenient method for screening mutations and typing DNA polymorphisms. SSCPs are codominant markers, like RFLPs.
Prinzenberg et al. (2002) found using PCR-SSCP analysis in the Gannan yak that the k-casein (CSN3) exon IV revealed a two allele polymorphism showing intermediate migration patterns compared to the cattle CSN3*A and B alleles. DNA sequences of these two alleles showed that all yak had nucleotide sequences corresponding to threonine in amino acid position 136 (identical to CSN3*A) and alanine in position 148 (identical to CSN3*B). This is in accordance with a sequence reported from wisent (Bison bonasus) (CSN3*GBison) and may represent the common ancestor of CSN3*A and B variants of cattle. Moreover, a 12 bp insertion resulting in a duplicated nucleotide and amino acid motive was found in one yak allele compared to the other. Position of the insertion could not be unequivocally assigned. The duplication is either corresponding to the codons for amino acids 147 - 150 or 148 - 151, which are repeated identically. In 18 yak typed by SSCP analysis, the long variant was found with frequencies about 70 percent and the short variant about 30 percent, implicating the longer variant being the predominant and probably the older allele in yak. The loss of the insertion may have led to the ancestral CSN3 allele from which all variants in Bos indicus and Bos taurus known today have evolved.
It is worth noting that this pilot study on milk protein polymorphism at the DNA level may shed more light on the understanding of yak genetics as they are coding genes and evolved under both natural and artificial selection over long periods. Consequently, they may also contribute to the detection of genetic markers for use in breeding programmes for genetic improvement, particularly in relation to milk quality and quantity.
SNPs are DNA sequence variations that occur when a single nucleotide in the animal mt-DNA or nuclear genome sequence is altered and detected by traditionally direct DNA sequencing protocol. For example, a SNP might change the DNA sequence AAGGCTAA to ATGGCTAA. SNPs occur at one SNP every 1.9 kilobases in the human genome. SNPs can occur in both coding (gene) and noncoding regions of the genome. Many SNPs have no effect on cell function, but it is believed that others could predispose organism to disease or influence their response to a challenge. SNPs are evolutionarily stable - not changing much from generation to generation - making them easier to follow in population studies. SNPs also have properties that make them particularly attractive for genetic studies. They are more frequent than microsatellite markers, providing markers near to or in the locus of interest, some located within the gene (cSNP), which can directly influence protein structure or expression levels, giving insights into functional mechanisms.
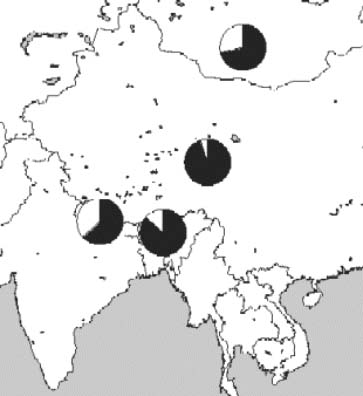
Bailey et al. (2002) using sequence data of the D-loop of mt-DNA found two divergent haplogroups, Y1 and Y2, in yak; but there does not appear to be a significant difference between the Chinese, Bhutanese, Nepalese and Mongolian yak. They supposed that a movement of the yak across the mountainous regions in Central Asia eventually reduced the amount of geographical partitioning observed as shown in Figure 15.1 after the domestication estimated to have occurred 5 000 years ago, based on a molecular clock. Nucleotide diversity values have been calculated for both the haplogroups Y1 and Y2 and for all Y (both 1 and 2) and they are very different. Therefore, the authors suggested a dual domestication of the yak. In comparison with European cattle, it is clearly seen that the variability found in yak is of the same order as that found in European cattle.
The total nucleotide variation for both yak haplogroups, is highest because it reflects the whole mt-DNA domestic pool from both centres of domestication. Lower nucleotide diversity for the Y1 and Y2 is thought to be due to both the limited number of samples included and a relatively recent domestication of the yak compared to the other cattle.
However, it would be incautious to adopt this conclusion without further study with a large number of samples collected in a large geographic area covering most of the yak-raising countries.
Figure 15.1 A distribution map of the Y1 and Y2 haplogroups in samples from China, Bhutan, Nepal and Mongolia, where the shaded area of the pie is Y1 and the nonshaded area is Y2 [Source: Bailey et al., 2002].
In contrast to the qualitative traits, most (if not all) characters of morphological, physiological, behavioural and economic importance in yak are quantitative traits that result from the complicated interaction of many genes. Therefore, it is not possible to classify yak into one or another discrete class. For instance, growth rate and the milk yield of yak vary among individuals and are dependent on many different physiological systems with possibly hundreds of genes influencing the phenotype. Such characters are described as quantitative and knowledge of the phenotype of an individual does not allow any specification of its genotype in terms of particular genes.
Many pairs of genes influence the expression of any quantitative trait. It is likely that a trait, such as milk yield in yak, must be influenced by many genes relating to hormone production and to the multitude of enzymes involved in metabolism. In reality, however, the contribution of an individual locus to the expression of a trait ranges from small to large. Should one gene play a large role in determining expression of a quantitative trait, it is termed a major gene. For example, major genes are known to affect the expression of double muscling in Belgian Blue cattle and fertility in Booroola sheep and Meishan pig.
The loci controlling the quantitative traits are called quantitative trait loci, or QTL (It is also found in some publications as ETL to represent the economic trait loci). The identification of the QTLs or closely linked genetic markers would allow genetic selection on the basis of both an individual's breeding value and on genotype using a "marker-assisted" breeding programme. It has been well demonstrated that mapping markers linked to QTLs might identify regions on the chromosomes that may contain genes involved in the expression of the quantitative trait.
Because large-scale conventional selection procedures are rare for the yak - or at best suboptimal - the use of marker-assisted selection might be particularly appropriate for yak once the techniques and the identification of QTLs have been made. Some discussion of the potential merits was provided by Wiener and Bishop (2002) who noted that, in general, marker-assisted selection is considered to be most beneficial for traits that are difficult or expensive to measure, traits that only occur in one sex (e.g. milk or reproduction) or traits that are expressed late in life (e.g. longevity). A further potential use of markers is for selection for disease resistance (Crawford et al., 2000). If a genetic marker is available that indicates resistance to a particular disease, it should be possible to reduce or even eliminate the disease without having to expose the animals to that disease.
So far, the work of most relevance for yak breeding is that which has taken place in other cattle; for example, the intensive investigation on the marker-assisted selection for milk production in dairy cattle (Sonstegard et al., 2001) or for growth and carcass composition in beef cattle (Casas et al., 2000). However, so far no effort has been made to discover specific QTLs or closely linked markers in the yak.
Molecular-genetic techniques have made it possible to identify differences between individuals at the DNA level. In particular, the development of microsatellite markers as an abundant source of polymorphic and convenient markers has boosted the generation of linkage maps in the most important livestock species. These maps provide the basis for detection and exploitation of genes segregating at the QTLs (Montaldo and Meza-Herrera, 1998). After the development of high-density genetic maps of polymorphic markers, it became possible for the first time to dissect genetically quantitative traits in many species. The analysis of QTL segregating in crosses between inbred lines of animals indicated that in some cases only a small number of genetic loci contributed to a large proportion of the variance of each trait, which confirms the presence of major genes (see above) and provides a tempting target for further molecular investigation (Flint and Mott, 2001). Major genes have been detected for the carcass characteristics in both pigs and cattle and also for parasite resistance in sheep (Montaldo and Meza-Herrera, 1998). Most dairy-related QTLs have also been identified in Holstein grandsire families (Sonstegard et al., 2001).
The Bovini species consists of both domestic and wild cattle and buffalo species, which started to diverge more than four million years ago (Lenstra and Bradley, 1999). Many are among the most important species playing a key role throughout the history of mankind and fulfilling many agricultural, economic, cultural and religious functions. According to some authorities, Bovini can be divided into genera Bovina, Bubalina and Syncerina. Bovina are then separated into subgenera of Bos and their wild relatives Bison (Felius, 1995). Paleontological evidence indicates that taxa within the Bovina diverged from one another within the past 20 million years (Savage and Russell, 1983). As referred to in Chapter 2, there have been several changes in the name and taxonomic position of the yak in the last two centuries and both the Bos and the Poephagus genera have been used by various authors but with the same species name of grunniens.
Numerous attempts have been made to clarify phylogenetic relationships among the genera of the tribe Bovini, including the yak, but the results have been ambiguous. Bohlken (1958, 1961) first grouped the yak with cattle (Bos taurus), followed by gaur (Bos gaurus) and banteng (Bos javanicus) in one clade, which then clustered with the American bison (Bison bison) and European bison (Bison bonasus). Groves (1981) concluded that the gaur and banteng represented the most primitive members of the Bos/Bison clade and that the two species of Bison were most closely related to the yak. Olsen (1990, 1991), who provided a useful review of the taxonomic evidence, also found that the yak has the same arrangement of premaxillaries, maxillaries and nasals as in Bison and are different from the structure in Bos. But Olsen also noted a difference from bison on the intervening extension of the dorsal margin of the maxilla that prevents the nostrils from reaching the premaxillae in the yak - possibly due to a different arrangement of the facial muscles that control the lips and nostrils in yak. This led Olsen to propose a re-establishment of the yak in the genus Poephagus, different from the genus Bos and Bison. In a study of phylogeny of the tribe Bovini, Geraads (1992), using a matrix of 57 (mainly cranial) characters and 32 taxa of fossil and current Bovini, considered Bos + Bibos + Bison + Poephagus + L. etruscus-callisarni + Epileptobos as a mono- or paraphyletic group and concluded that the yak is close to the bison among the current forms.
Since the late 1980s, the sequences of mitochondrial DNA of yak have been included in some of the phylogenetic studies of Bovidae. Miyamoto et al. (1989) sequenced the 12S rRNA and three tRNA genes, and a 247 bp partial hypervariant D-loop fragment of four taxa in the tribe Bovini and the results showed a similar topology of phylogeny with Groves (1981), the yak grouped with the Bison bison first, with an average divergence of 2.6 percent for the conservative rRNA/tRNA genes and 9.1 percent for the D-loop fragment and then followed by Bos taurus (Kraus et al., 1992). Ward et al. (1999) using a partial mt-DNA control region of 667 base pairs, found that the percentage nucleotide divergence of Bos indicus and Bos taurus from the yak was 24.23 percent and 29.53 percent, respectively, but that of American bison and European bison from the yak was only 12.28 percent and 16.59 percent, respectively.
Within the nuclear genome alone [on 15 endonucleases' site mapping of nuclear ribosomal DNA (rDNA)], Wall et al. (1992) confirmed the position of yak as more closely related to the bison and wisent than to Bos taurus and Bos indicus cattle as earlier suggested by Groves (1981). Hassanin and Douzery (1999) sequenced the cytochrome b and 12S rRNA of mt-DNA and aromatase cytochrome P-45 and the promoter segment of the lactoferrin-encoding genes of nuclear genomic DNA in the yak alongside other Bovidae species. With the combined data of these four mitochondrial and nuclear markers, the yak clustered with the American bison first and then grouped with Bos taurus in the phylogenetic tree.
Ritz et al. (2000) used 20 bovine microsatellite markers to determine the phylogenetic relationships in the tribe Bovini, including the yak and found that the yak emerged as a separate subgenus Poephagus within the genus Bos. Buntjer et al. (2002) claimed that the published phylogenies of this tribe Bovini, based on mitochondrial DNA, contain anomalies, while nuclear sequences show only low variation. They used AFLP fingerprinting to detect variation in loci distributed over the nuclear genomes of Bovini species. Computer-assisted scoring of electrophoretic fingerprinting patterns yielded 361 markers, which provided sufficient redundancy to suppress stochastic effects of intraspecies polymorphisms and length homoplasies (co-migration of non-homologous fragments). Tree reconstructions reveal three clusters: African buffalo with water buffalo, ox with zebu and bison with wisent. Similarity values suggest a clustering of gaur and banteng, but bifurcating clustering algorithms did not assign consistent positions to these species and yak. They proposed that because of shared polymorphisms and reticulations, tree topologies are only partially adequate to represent the phylogeny of the Bovini. Principal-coordinate analysis positioned zebu between a gaur/banteng cluster and taurine cattle. This correlates with the region of origin of these species and suggests that genomic distances between the cattle species have been influenced by genetic exchange between neighbouring ancestral populations. The yak clearly stands alone in a separate group but relatively close to the cluster of bison and wisent. In the multivariate image, regression-specific PCR and AFLP analysis (Buntjer, 1997), there was a clustering of yak with both bison and wisent that are clearly different from other non-buffalo Bovini species, in agreement with the morphological study by Geraads (1992). Buntjer (1997) explained this as due to both yak and bison/wisent being adapted to relatively cold conditions, suggesting a correspondence between genetic distance and geographical origin of the species.
In conclusion, it seems clear that the yak is different from Bos taurus and Bos indicus cattle and closer to the American bison (Bos bison) in terms of its cranial morphology and its genome including both the mt-DNA and nuclear DNA markers surveyed, even with inclusive phenotypic characteristics. Therefore, in agreement with Olsen (1991), it seems that in the choice of nomenclature for both the domestic yak and the wild yak, the subgenus, Poephagus, seems more appropriate than Bos.
References
Anderson, S. et al. (1982). Complete sequence of bovine mitochondrial DNA. Conserved features of the mammalian mitochondrial genome. Journal of Molecular Biology, 156: 683-717.
Bailey, J.F. et al. (2002). Genetic variation of mitochondrial DNA within domestic yak populations. Proceedings of the third international congress on yak, in Lhasa, China, 4-9 September 2000. International Livestock Research Institute (ILRI), Nairobi, pp. 181-189.
Bohlken, H. (1958). Vergleichende Untersuchungen an Wildrindern (Tribus Bovini Simpson, 1954). Zool. Jahrb., 68: 113-202.
Bohlken, H. (1961). Haustiere und Zoologische Systematick. Z. Tier. Zuchtungsbiol., 76: 107-113.
Buntjer, J.B. (1997). DAN repeats in the vertebrate genome as probes in phylogeny and species identification. Academic thesis, Utrecht University, Netherlands.
Buntjer, J.B. et al. (2002). Phylogeny of bovine species based on AFLP fingerprinting. Heredity, 88: 46-51.
Casas, E. et al. (2000). Quantitative trait loci affecting growth and carcass composition of cattle segregating alternate forms of myostatin. J. Anim. Sci., 78:560-569.
Crawford, A.M., Dodds, K.G. and McEwan, J.C. (2000). DNA markers, genetic maps, and the identification of QTL: general principles. In: Axford, R.F.E. et al. (ed.), Breeding for disease resistance in farm animals, 2nd edition. Wallingford, U.K., CABI Publishing, pp. 3-26.
Dorji, T. et al. (2002). Genetic diversity in Bhutanese yak (Bos grunniens) populations using microsatellite markers. Proceedings of the third international congress on yak, in Lhasa, China, 4-9 September 2000. International Livestock Research Institute (ILRI), Nairobi, pp. 197-201.
Felius, M. Cattle breeds - an encyclopaedia. Misset uitgeverij bv, postbus 4, 7000 BA Doetinchem, Netherlands, pp. 12-15.
Flint, J. and Mott, R. (2001). Finding the molecular basis of quantitative traits: successes and pitfalls. Nature Review Genetics, 2: 437-445.
Fries, R. and Popescu, P. (1999). Cytogenetics and physical chromosome maps. In: Fries, R. and Ruvinsky, A. (ed.), The Genetics of Cattle. CABI, Wallingford, UK. pp. 247-327.
Geraads, D. (1992). Phylogenetic analysis of the tribe Bovini (Mammalia: Artiodactyl). Zoological Journal of the Linnean Society, 104: 193-207.
Goe, M.R. and Stranzinger, G. (2002). Monitoring of traits for yak and yak hybrids. Proceedings of the third international congress on yak, in Lhasa, China, 4-9 September 2000. International Livestock Research Institute (ILRI), Nairobi, pp.291-299.
Groves, C.P. (1981). Systematic relationships in the Bovinii (Artiodactyla, Bovidae). Zeitschrift fuer Zoologische Systematik und Evolutionsforschung, 19: 264-278.
Guo Aipu (1983). A comparative study on chromosomes of oxen (Bos taurus), yak (Bos grunniens) and their hybrids. Acta Genetica Sinica, 2: 137-143.
Gupta, N. et al. (1996). Chromosomes of yak. Indian Journal of Animal Sciences, 66: 453-457.
Halnan, C.R.E. (1989). Karyotype and phenotype in cattle and hybrids of the genus. In: Halnan, C.R.E. (ed.), Cytogenetics of Farm Animals. Wallingford, UK, CABI, pp. 235-255.
Han Jianlin (1996). Yak genetic resources in China: evaluation of chromosome, protein and mt-DNA polymorphism. In Miller, D.G., Craig, S.R. and Rana, G.M. (ed), Proceedings of a workshop on conservation and management of yak genetic diversity, at ICIMOD, Kathmandu, 29-31 October 1996. ICIMOD (International Centre for Integrated Mountain Development), Kathmandu, pp. 175-183.
Han Jianlin et al. (2002). Low level of cattle introgression in yak populations from Bhutan and China: Evidences from Y-specific microsatellites and mitochondrial DNA markers. Proceedings of the third international congress on yak, in Lhasa, China, 4-9 September 2000. International Livestock Research Institute (ILRI), Nairobi, pp. 190-196.
Hanotte, O. et al. (2000). Cattle microsatellite markers for amplification of polymorphic loci in Asian Bovidae. In: Shrestha, J.N.B. (ed), Proceedings of the 4th global conference on conservation of domestic animal genetic resources held in Kathmandu, Nepal, 17-21 August 1998, pp. 47-49.
Hassanin A. & Douzery, E.J.P. (1999). Evolutionary affinities of the enigmatic saola (Pseudoryx nghetinhensis) in the context of the molecular phylogeny of Bovidae. Proceedings of Royal Society London, B 266: 893-900.
Hines, H.C. (1999). Blood groups and biochemical polymorphisms. In: Fries, R. and Ruvinsky, A. (ed.), The Genetics of Cattle. Wallingford, U.K., CABI, pp. 77-121.
Hishida, O. et al. (1996). Cross-species amplification and polymorphism of microsatellite loci in Asian bovidae. Proceedings of the eighth AAAP Animal Science Congress, 13-18 October 1996. Vol. 2. Japanese Society of Zootechnical Science, Tokyo.
Hwu, H.R. et al. (1986). Insertion and/or deletion of many repeated DNA sequences in human and higher ape revolution. Proceedings of the National Academy of Sciences of the U.S.A., 83: 3875-3879.
International Human Genome Sequencing Consortium (2001). Initial sequencing and analysis of the human genome. Nature, 409: 860-921.
Kraus, F., et al. (1992). Mis-pairing and compensational changes during the evolution of mitochondrial ribosomal RNA. Molecular and Biological Evolution, 9: 770-774.
Li Jiyou, et al. (1994). Advance on the research of chromosomes in yak (Bos grunniens). Proceedings of the first international congress on yak. Journal of Gansu Agricultural University (Special issue June 1994), pp. 73-76.
Lenstra, J.A. and Bradley, D.G. (1999). Systematics and phylogeny of cattle. In: Fries, R. and Ruvinsky, A. (ed.), The Genetics of Cattle. CABI, Wallingford, U.K., pp. 1-14.
Miyamoto, M.M., Tanhauser, S.M. and Laipis, P.J. (1989). Systematic relationship in the artiodactyls tribe Bovini (family Bovidae), as determined from mitochondrial DNA sequences. Systematic Zoology, 38: 342-349.
Montaldo, H.H. and Meza-Herrera, C.A. (1998). Use of molecular markers and major genes in the genetic improvement of livestock. EJB Electronic Journal of Biotechnology, 1 (2): 1-7.
Nijman, I.J. and Lenstra, J.A. (2001). Mutation and recombination in cattle satellite DNA: a feedback model for the evolution of satellite DNA repeats. Journal of Molecular Evolution, 52: 361-371.
Nivsarkar, A.N., Gupta, S.C. and Gupta, N. (1997). Yak production. Indian Council for Agricultural Research, New Delhi, 394 pp.
Olsen, S.J. (1990). Fossil ancestry of the yak, its cultural significance and domestication in Tibet. Proceedings of the Academy of Natural Sciences of Philadelphia, 142: 73-100.
Olsen, S.J. (1991). Confused yak taxonomy and evidence of domestication. Illinois State Museum Scientific Papers, Vol. 23: 387-393.
Prinzenberg, E.M., Han Jianlin and Erhardt G. (2002). Variants of CSN3 in Chinese yak (Bos grunniens). Proceedings of the 28th International conference of Animal Genetics held in Göttingen, Germany, 11-15 August 2002, p. 127 (abstract).
Qi Xuebin and Han Jianlin (2002). Introgression makes yak populations genetically different: Evidence from Beta-lactoglobulin variations. Proceedings of the third international congress on yak, in Lhasa, China, 4-9 September 2000. International Livestock Research Institute (ILRI), Nairobi, pp. 202-208.
Qi Xuebin et al. (2002a). Cattle mitochondrial DNA introgression in yak (Poephagus or Bos grunniens). Proceedings of the 28th International conference of Animal Genetics held in Göttingen, Germany, 11-15 August 2002, p.102 (abstract).
Qi Xuebin et al. (2002b). Y-chromosome specific microsatellite polymorphism in Chinese yak. Proceedings of the 7th world congress on genetics applied to livestock production held in Montpellier, France, 19-23 August 2002, 33, 509-512.
Ritz, L.R. (1997). Genetic diversity in the tribe Bovini. PhD dissertation of University of Bern, Switzerland, 79 pp.
Ritz, L.R. et al. (2000). Phylogenetic analysis of the tribe Bovini using microsatellites. Animal Genetics, 31: 178-185.
Rozhkov, YuI and Galimov, I.R. (1990). Salivary gland amylase polymorphism in pigs and cattle detected by affinity electrophoresis. Animal Genetics, 21: 277-283.
Savage, D. and Russell, D.E. (1983). Mammalian paleofaunas of the World. Reading, MA., U.S.A., Addison-Wesley.
Sonstegard, T.S., Van Tassell, C.P. and Ashwell, M.S. (2001). Dairy cattle genomics: Tools to accelerate genetic improvement. Journal of Animal Science, 79 (E. Suppl.): E307-315.
Sulimova, G.E., Badagueva, IuN and Udina, I.G. (1996). Polymorphism of the kappa-casein gene in populations of the subfamily Bovinae. Genetika, 32:1576-1582. [in Russian]
Tao Shixin and Han Jianlin (2002). Population genetic variations of haemoglobin in yaks, cattle and their hybrids. Proceedings of the third international congress on yak, in Lhasa, China, 4-9 September 2000. International Livestock Research Institute (ILRI), Nairobi, pp. 213-215.
Tu Zhengchao, Zhang Yaping and Qiu Huai (1997). Genetic diversity and divergence in Chinese yak (Bos grunniens) populations inferred from blood protein electrophoresis. Biochemical Genetics, 35: 13-16.
Tu Zhengchao, Zhang Yaping and Qiu Huai (1998). Mitochondrial DNA polymorphism and genetic diversity in Chinese yak. Acta Genetica Sinica, 25: 205-212.
Vaiman, D. (1999). The molecular genetics of cattle. In Fries, R. and Ruvinsky, A. (ed.), The Genetics of Cattle. Wallingford, U.K., CABI, pp. 123-161.
Wall, D.A., Davis, S.K. and Read, B.M. (1992). Phylogenetic relationships in the subfamily Bovinae (Mammalia: Artiodactyla) based on ribosomal DNA. Journal of Mammalogy, 73: 262-275.
Wang Minqiang (2000). Study on some germplasm characteristics of Datong yak. Doctoral dissertation of Northwest Science & Technology University of Agriculture and Forestry, Shaanxi, China, 94 pp.
Ward, T.J. et al. (1999). Identification of domestic cattle hybrids in wild cattle and bison species: a general approach using mt-DNA markers and the parametric bootstrap. Animal Conservation, 2: 51-57.
Wiener, G. (1994). Opportunities and constraints for genetic improvement with reference to the yak. Proceedings of the first international congress on yak. Journal of Gansu Agricultural University (Special issue June 1994), pp. 35-44.
Wiener, G. (1997). Yak breeds and the utilization of genetic diversity. Proceedings of the second international congress on yak, in Xining, China, 1-6 September 1997. Xining, China, Qinghai People's Publishing House, pp. 50-53.
Wiener, G. and Bishop, S.C. (2002). Opportunities for the improvement of yak production with particular reference to genetic options. Proceedings of the third international congress on yak, in Lhasa, China, 4-9 September 2000. International Livestock Research Institute (ILRI), Nairobi, pp. 225-234.
Yu, Y. et al. (1999). Mitochondrial DNA variation in cattle of South China: origin and introgression. Animal Genetics, 30: 245-250.
Zhang Caijun (1997). Advances of studies on biochemical genetics in yak. Proceedings of the second international congress on yak, in Xining, China, 1-6 September 1997. Xining, China, Qinghai People's Publishing House, pp. 55-58.
Zhang Yuezhou et al. (1994). Current status and future development of the studies on protein polymorphism on yak. Proceedings of the first international congress on yak. Journal of Gansu Agricultural University (Special issue June 1994), pp. 62-66.
Zhao Xinbo, Zhong Guanghui and Cai Li (1994). Studies on mitochondrial DNA RFLP of yaks and cattle. Proceedings of the 1st international congress on yak. Journal of Gansu Agricultural University (Special issue June 1994), pp. 96-98.
Zhong Jingcheng (1997). The genetic diversity of Chinese yak and its significance. Proceedings of the second international congress on yak, in Xining, China, 1-6 September 1997. Xining, China, Qinghai People's Publishing House, pp. 65-69.
Zhong Jingcheng (1996). Yak genetics and breeding. Sichuan Scientific and Technology Press, Chengdu, China, 271 pp.
|
[17] Han Jainlin is a
molecular geneticist at the International Livestock Research Institute, Kenya.
He is also Professor of Animal Genetics and Breeding, Gansu Agriculture
University, China and Executive Secretary of the International Yak Information
Center and China Breed Association in Lanzhou, China |
![]()
![]()
![]()