![]()
![]()
![]()
Meldy L.A. Hosang, Jelfina C. Alouw and H. Novarianto*
* Indonesian Coconut and Other Palm Research Institute, P.O. Box 1004, Manado 95001; e-mail: [email protected]
Abstract
Brontispa longissima is one of the major pests in several provinces of Indonesia. Biological control by using natural enemies such as parasitoids and entomopathogens has been proved as a promising method to control plant pests. There are three potential natural enemies for controlling B. longissima namely: pupal parasitoid (Tetrastichus brontispa), entomopathogenic fungi (Metarhizium anisopliae var. anisopliae and Beauveria bassiana). T. brontispa has an important role as pupal parasitoid both under laboratory and field conditions. Percent parasitism of pupa under laboratory and field conditions ranged from 76.7 to 87.0 percent and from 35.71 to 73.56 percent, respectively. Pathogenicity of M. anisopliae var. anisopliae and B. bassiana were examined under laboratory, and field conditions. The results showed that these fungi can infect both larvae and adults of B. longissima. The effective concentration suggested to control B. longissima in the field is 5 x 105 konidia/ml. Mortality of larval and adult B. longissima caused by M. anisopliae var. anisopliae was 100 percent and 65 percent, respectively and by B. bassiana was 100 percent and 73.75 percent, respectively. Those entomopathogenic fungi can be applied by spraying conidial suspension twice-yearly at two weeks interval. Spraying the entomopathogenic fungi M. anisopliae var. anisopliae and B. bassiana reduced the pest population at about 90.37-95.0 percent.
Key words: Biological control, Tetrastichus brontispa, Metarhizium anisopliae var. anisopliae, Beauveria bassiana, Brontispa longissima
Introduction
Brontispa longissima Gestro (Coleoptera: Chrysomelidae) is one of the important pests in several provinces in Indonesia. Both larvae and adults attacked coconut leaves, particularly unfolded leaves. Therefore, the pest can decrease coconut production. The Chrysomelid beetles attack all ages of coconut, although more damage is found in coconut plantation between four to five years old, especially in drying areas. Severe damage of this pest would kill the palms.
Various strategies have been used to control B. longissima, but most of them strongly depend on the use of insecticides. This practice substantially increases cost of production besides threat to the ecosystem. Additionally, chemical control may not be a long-term solution because of (1) the possibility that pests would develop resistance against the commonly used insecticides and (2) the increasing likelihood of outbreaks of secondary pests. Integrated pest management is one promising approach for a sustainable management of coconut plantations and could be capable to control and reduce populations of B. longissima.
Biological control by using parasitoid, predator and entomopathogenic fungus has a good chance to depress population of B. longissima in the field. Pest control by using natural enemies is not as popular as using pesticide. Biological control would decrease the use of insecticides. Therefore, it has a good impact on the environment. Additionally, this practice has a long-term impact to depress or manage the pest population on coconut plantation in low level of palm damage.
Actually, pest control has no intention to eliminate the pest totally but to maintain the natural balances by keeping the pest population below economic threshold level. Tetrastichus brontispa (pupal parasitoid) and entomopathogenic fungi Metarhizium anisopliae var. anisopliae and Beauveria bassiana are promising natural enemies of B. longissima.
Study on the use of pupal parasitoid T. brontispa has been done by some researchers (Kalshoven, 1981; Lever, 1969; Tjoa, 1952). This parasitoid also attacks Plesispa reichei (Heroetadji, 1989; Ooi et al., 1989). The use of entomopathogenic fungi to control B. longissima is still limited except for other pests. Metarhizium anisopliae isolated from Oryctes rhinoceros can also infect B. longissima under laboratory condition (Soekarjoto et al., 1994). M. anisopliae isolated from B. longissima in South Sulawesi was first reported by Hosang et al. (1996). This fungus attacks second instar larvae (L2) (100 percent) and adults (52.5 percent) under laboratroy condition. B. bassiana has intensively been used in managing several pests such as Ostrinia nubilalis (Hübner) (Bing and Lewis, 1991), Leptinotarsa decemlineata (Say), (Anderson et al., 1988), Spodoptera exigua (Hübner) (Barberchech and Kaya, 1991), Cylas formicarius (Burdeos and Villacarlos, 1989), and cotton pest, Anthonomus grandis Boheman (Wright and Chandler, 1992). This fungus is also found in the yellow rice-borer (Tryporyza incertulas (Walker), stem borer Sesamia inferens (Walker) (Kalshoven, 1981) and Nilaparvata lugens (Stal) (Domsch et al., 1980). The efficacy of B. bassiana had been evaluated on the three coconut pests namely: Tirathaba rufivena Walker, Promecotheca cumingii Baly and Plesispa reichei Chapius. The experiment results proved that these fungi were effective on the pests (Gallego and Gallego, 1988). This information demonstrated the high potential of the three entomopathogenic fungi to be developed as promising natural enemies of B. longissima. In this paper we present the pest description and biology, palm damage, current status of coconut chrisomelid beetle, and biological control by using parasitoid T. Brontispa, entomopathogenic fungi M. anisopliae var. anisopliae and B. bassiana.
Description of Brontispa longissima (Gestro)
Coconut hispid, very destructive, 9 mm long in Indonesia, Malaysia, and Pacific Islands. It also occurs in other palms. Many local varieties have been described: var. longissima with brown elytra, original described in Wolan, one of the Aru Islands, and now common in Java; var. froggatti sharp with black elytra, from new Britain and Salomon Islands; var. selebensis Gestro with a spindle-shaped, black marking on the suture of the elytra, original from South and North Sulawesi, later also found near Bogor. Numerous forms, intermediate between the last two varieties occur in Sulawesi, the Moluccas and Irian. Fertile offspring from 'longissima' and 'selebensis' crosses could be produced in the laboratory.
The color of adults varies geographically from reddish-brown in Java to almost black in the Salomon Islands and Irian (Papua). Considerable overlapping of these forms, which were for long regarded as distinct species, occurs (Kalshoven, 1981).
Biology of Brontispa longissima
Eggs The eggs are brown and flat. They are laid singly or in groups of two to four on the still-folded heart leaves (Lever, 1979). An egg measures 1.4 mm in length and 0.5 mm in width (Tjoa, 1953). The incubation period reported by several researchers ranged from three to four days (Froggatt and O'Connor, 1941; Lever, 1979); five days (O'Connor, 1940; Waterhouse and Norris, 1987); four to seven days or four days on the average (Tjoa, 1953).
Larvae The newly hatched larvae are whitish, later turn to yellowish and have an average length of 2 mm. The older larvae have an average length of 8-10 mm. Larvae avoid light and have distally U-like hooks. B. longissima undergoes four larval instars (Froggatt and O'Connor (1941) or five to six larval instars (O'Connor, 1940). The total developmental period of larvae vary about 36 days (O'Connor, 1940); 30-40 days (Froggatt and O'Connor, 1941; Waterhouse and Norris, 1987); 23-43 days (Tjoa, 1953); or 35-54 days (Lever, 1979).
Pupae The newly formed pupae are yellowish-white and have an average length of 9-10 mm and a width of 2 mm. They have distally U-shaped hooks. The pupal period is six days (O'Connor, 1940; Waterhouse and Norris, 1987); four to five days (Tjoa, 1953); or four to six days (Lever, 1979).
Adult The adult male is generally smaller than the female and measures 7.5-10 mm long and 1.5-2 mm wide. They avoid light and stay inactive inside the still - folded heart leaf during day time and active fly and attack coconut plants at night. Female lays an average of 50-100 eggs (O'Connor, 1940) until 117 eggs (Tjoa, 1953). Pre-oviposition period is 74 days (O'Connor, 1940) or one to two months (Waterhouse and Norris, 1987). The adult longevity ranges from two and a half to three months (75-90 days) Tjoa (1953).
The development from egg to adult takes five to seven weeks. The beetles then mature in other two weeks. This species is one of the thoroughly studied pest in Indonesia, with work undertaken at Bogor, as well as Bulukumba and Manado (Kalshoven, 1981).
Palm damage caused by B. longissima
During 1919-1934, B. longissima had been recognized as a pest of coconut palm in five provinces in Indonesia namely: Central Java, East Java, D.I. Yogyakarta, South Sulawesi and North Sulawesi. According to Tjoa (1953), B. longissima var. javana was found in Java, Bali, Madura, Sumba and Papua; while B. longissima var. selebensis in South Sulawesi, North Sulawesi, Flores, Seram, Aru Island and Bogor. Recently, B. longissima has spread to several provinces. The pest was also found in Sumatera and Maluku. Suprapto (1983) reported that in 1980, B. longissima caused serious damaged in area of 2 000 ha in Lampung. Madry (1993) reported losses due to the pest in nine provinces of Indonesia namely; South Sumatera, Lampung, West Kalimantan, South Sulawesi, Maluku, Irian Jaya, Bali and D.I Yogyakarta amount to Rp 298 786 000.
B. longissima start attack coconut palm aged two to three years old. The older the palm the lower the infestation. No damage is reported in coconut palm aged eight to nine years old due to the difficulties of the pest to penetrate unopened leaves to lay the eggs. In contrast, the less compact leaves are more susceptible to Brontispa attacks (Tjoa, 1953). Waterhouse and Norris (1987) concluded that the pest attacks all age stages of coconut palm with serious damage occurring in young coconut palm in the seedling and coconut palm at the age of four to five years in the field during dry season.
Light attack result in minor leaf injury, and a slight decrease in fruiting at the axils of the damaged leaves. Fruit production is significantly reduced if eight or more leaves are destroyed. Under prolonged outbreak condition, as occurred in South Sulawesi for several years, fruit-shedding takes places, newly-formed leaves remain small, the trees appear ragged, and may ultimately die (Tjoa, 1953; Kalshoven, 1981; Suprapto 1983). The newly produced leaves are favourable for the development of the pest. Population of B. longissima was higher in early infestation and was lower when severe damage takes place. This could be related to the food availability in the field.
Current status of the key coconut chrysomelid beetles
There are three chrysomelid beetles attacking coconut palm in Indonesia, namely Brontispa longissima, Plesispa reichei and Promecotheca cumingii. The distribution and losses caused by the pest are shown in Table 1, 2 and Figure 1. Attacked areas of B. longissima have decreased from about 34 289.72 ha in 1984 to 1 389 ha in 2004. It indicates that there is reduction of pest population in that area due to the action of the natural enemies and environmental factors that are not favourable for the development of the pest.
Table 1. Situation of coconut Chrysomelid beetles in Indonesia
|
No. |
Pests |
Attacked area (ha) |
Locations (province) |
|
1983/1984 |
|||
|
1. |
Brontispa longissima |
34 289.72 |
Lampung, West Java, Central Java, D.I. Yogyakarta, West Kalimantan, South Sulawesi, S.E. Sulawesi, Bali, Papua |
|
1984/1985 |
|||
|
2. |
Promecotheca cumingii |
1 812.83 |
Central Sulawesi |
|
1993 |
|||
|
1. |
Brontispa longissima |
|
South Sumatra, Lampung, West Kalimantan; South Sulawesi, Maluku, Papua, Bali, D.I. Yogyakarta |
|
September 2004 (source: Dirjen Bina Produksi Perkebunan 2004) |
|||
|
1. |
Brontispa longissima |
1 389.00 |
West Java, West Sumatera, Bangka Belitung, Papua, Nusa Tenggara Barat, East Java, Nusa Tenggara Timur, Central Java, D.I. Yogyakarta |
|
2. |
Plesispa reichei |
99.65 |
Nusa Tenggara Barat, East Java, Gorontalo |
Table 2. Coconut areas attacked by Plesispa reichei in West Kalimantan (July 2004)
|
Sub district |
Area (ha) |
Attacked area (ha) |
|
Mempawah Hilir |
4 992 |
278 |
|
Sungai Pinyuh |
2 044 |
205 |
|
Sungai Kunyit |
2 535 |
97 |
|
Siantan |
2 885 |
85 |
|
Sui Kakap |
10 188 |
55.8 |
|
Rasau Jaya |
953 |
87 |
|
Sungai Raya |
190 |
5 |
|
M.H. Utara |
450 |
375 |
|
M.H. Selatan |
1 478.5 |
4.9 |
|
Total |
25 715.5 |
1 192.7 |
Source: Extention Service West Kalimantan.
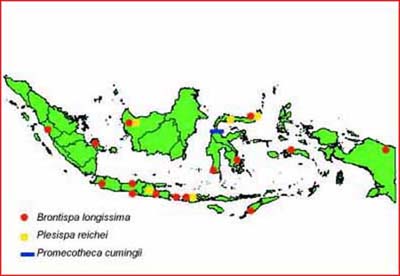
Figure 1: Distribution of Chrysomelid beetles in Indonesia
Promecotheca attacks are still limited to the areas of Central Sulawesi. The development of the pest was suppressed by the natural enemies as listed in Table 1. Generally, P. reichei attacks only young coconut palms, but it can also attack older palm as reported in West Kalimantan. About 5 percent (1 192.7 ha) of the coconut palm areas are attacked by the pest. In order to prevent the outbreaks and the spread of the pest to other coconut palm areas in Indonesia and even to other countries, the control of the pest should be done. Natural enemies of B. longissima can also be used as promising biological control agents to control P. reichei.
Biological control
Biological Control is the action of parasitoids, predators and pathogens in maintaining the pest population density at a lower average than it would occur in its absence. Biological control has recently been recognized as a promising and effective tool in the management of the most important pest on coconut palm (Sathiamma et al., 2001). Among the natural enemies used in biological control, information about predators against chrysomelid beetles is still limited. Waterhouse and Norris (1987) reported some earwigs preying on B. longissima. However, no research has been done to study the basic aspect of the predator and to develop them as an important potential biological agent of the chrysomelid beetles. This chapter presents the information regarding the parasitoids and entomopathogenic fungi of Brontispa longissima.
1. Parasitoids
The parasitoid complex of B. longissima comprises three egg parasites, Haeckeliana brontispa Ferriere, Trichogrammatoidea nana Zehntner (both Hymenoptera: Trichogrammatoidae) and a species of Ooencyrtus (Hymenoptera: Chalcidoidea), and a parasitoid of the larvae and pupae, Tetrastichus brontispa Ferriere (Hymenoptera: Eulophidae) (Lever, 1969). In Java, a complex of parasitoids occurs: (1) a strain of the trichogrammatid H. brontispa, with one wasp developing per Brontispa egg, and found on about 15 percent of Brontispa eggs in the field (Kalshoven, 1981) or 17 percent (Tjoa, 1952); (2) the encyrtid Ooencyrtus podontiae Gah. occurring on about 10 percent of the eggs (Kalshoven, 1980; Tjoa, 1952); (3) the eulophid Tetrastichus brontispa Ferr., found in 60-90 percent of the pupae and 10 percent of the larvae eggs (Kalshoven, 1980; Tjoa, 1952), developing in 18 days; about 20 specimens emerge from one Brontispa pupa. Hyperparasitoids have not been found. The same group of parasitoids could also be observed in 1940 in East Java near Kediri. Tetrastichus (= Tetrastichodes) which is a very distinctive parasitoid, is also found in other parts of Java, in Bali and Papua (Kalshoven, 1981). The Tetrastichus is the most effective parasitoid of Brontispa. Control of the beetle was achieved in Celebes by introducing this parasitoid from Java (see Lever, 1969).
1.1. Population of B. longissima and its parasitoid
Results of the pest collection done in West Java, Central Java and South Sulawesi in 1996 showed that population of B. longissima, natural enemies and palm damage in the three surveyed areas were in damaging stadium. Population of larvae and adults in West Java, Central Java and South Sulawesi were as follow: 57.42 percent and 17.98 percent; 35.86 percent and 38.34 percent; 63.63 percent and 18.93 percent, respectively (Hosang et al., 1996).
Observation results obtained in district Selayar and Jeneponto (South Sulawesi) showed that the pest in the overlapping generation or in other words egg, larval, pupal and adult stages are available in the field (Hosang et al., 1999). Total population of eggs, first instar larvae (L1), second instar larvae (L2), third instar larvae (L3), fourth instar larvae (L4), fifth instar larvae (L5), pupae and adults in seven regions of district Jeneponto were as follow 7.4 percent, 11.6 percent, 18.6 percent, 15.5 percent, 8.2 percent, 7.2 percent, 5.5 percent and 25.7 percent, respectively. So, the palm-damaging instars were 61.3 percent as larval instar (L1-L5) and 25.7 percent as adults. This information is necessary for the decision making process to control the pest when outbreaks occur.
About 40 percent of 245 pupae collected in District Selayar and 2.7 percent of 113 pupae collected in District Jeneponto were parasitized by Tetrastichus brontispa. Percent parasitism varies in every location (Table 3). Hosang et al. (1996) reported that T. brontispa parasitizing in Pakuwon (West Java), Central Java and South Sulawesi were 36.4 percent, 11.1 percent and 50.6 percent, respectively. Level of parasitization is considered lower than that reported by Kalshoven (1981) that percent parasitism was 10 percent for larvae and 60-90 percent for pupae. The differences could be caused by environmental condition in every location, insect and plant biodiversity. In addition, larvae and adults collected in District Jeneponto were infected by M. anisopliae var. anisopliae, but no infections were found in larvae and adult collected in District Selayar. It indicates that M. anisopliae var. anisopliae does not evenly distribute in all attacked areas of B. longissima in South Sulawesi. All natural enemies were tested under laboratory and field condition.
Table 3. Population of healthy and parasitized/infected B. longissima in District Selayar and Jeneponto, South Sulawesi (Hosang et al., 1996)
|
Stages |
Locations |
|
|
Selayar |
Jeneponto |
|
|
Healthy Larvae (L4 dan L5) |
725 |
271 |
|
M. anisopliae var. Anisopliae infected larvae |
0 |
31(10.3%) |
|
Healthy pupae |
147 |
110 |
|
T. brontispa parasitized pupae |
98(40%) |
3(2.7%) |
|
Healthy adults |
1 156 |
505 |
|
M. anisopliae var. Anisopliae infected adults |
0 |
18(3.4%) |
1.2. The test of T. brontispa as pupal parasitoid under laboratory and field condition
Percent parasitism of T. brontispa ranged from 76.7 to 87.0 percent under laboratory condition (Table 4). The success of parasitoids much depends on the age of pupae to be parasitized. Pupae at the age of one to two days are more susceptible than the older one.
Table 4. Percentage of parasitized pupae by T. brontispa (Hosang et al., 1996)
|
Treatments |
Percentage of parasitized pupae |
|
1 parasitized pupae |
76.7a |
|
2 parasitized pupae |
81.7a |
|
3 parasitized pupae |
83.3a |
|
4 parasitized pupae |
87.0a |
|
5 parasitized pupae |
80.0a |
The parasitization level due to the release of T. brontispa-parasitized pupae in the field was 35.71-73.56 percent. The results demonstrated the high potential of T. brontispa to be developed as a biological control agent of B. longissima.
2. Entomopathogenic fungi M. anisopliae var. anisopliae and B. bassiana
2.1. Test of different conidial concentrations of M. anisopliae var. anisopliae on different B. longissima stages
Based on the analysis of variance for mortality of third instar larvae (L3), fourth instar larvae (L4), fifth instar larvae (L5) and adults at 20 days after treatments (day) showed that there were highly significant differences among treatments (p <0.01). Results have shown that mortality of L3, L4 and L5 at concentration of 5 x 104, 5 x 105 was insignificantly different, but significantly different at the concentration of 5 x 102 and 5 x 103 conidia/ml. There was insignificant differences at the control and the concentration of 5 x 104 and 5 x 105, but significant differences were observed in adults at the concentration of 5 x 104 and 5 x 105. Significant difference was reported between concentration 5 x 104 and 5 x 105. Therefore, the lowest concentration of M. anisopliae var. anisopliae conidia that caused highest mortality of L3, L4 and L5 is 5 x 104 conidia/ml; while for adults is 5 x 105 conidia/ml (Table 5). This result was similar with Beauveria bassiana infecting B. longissima (Hosang, 1996). So, both fungi were subjected to tests in the next experiments in the screen cages.
The LC50 and LC95 values of conidial concentrations of M. anisopliae var. anisopliae on larvae and adults of B. Longissima at ten days are shown in Table 6. The result showed that for larvae, the LC50 values were 5.1 x 102 to 8.6 x 102 conidia/ml and 4.7 x 106 conidia/ml for adults.
Table 5. Mortality of L3, L4, L5 and adults at different conidial concentrations of M. anisoliae var. anisopliae at 20 days after treatment
|
Conidial concentrations (conidia/ml) |
Larval stages |
Adults |
||
|
3rd |
4th |
5th |
||
|
Control |
1.25a |
2.00a |
2.75a |
1.75a |
|
5 x 102 |
13.75b |
17.75b |
14.25b |
1.75a |
|
5 x 103 |
18.50c |
18.00b |
16.50b |
2.75a |
|
5 x 104 |
20.00d |
20.00c |
20.00c |
7.50b |
|
5 x 105 |
20.00d |
20.00c |
20.00c |
13.00c |
- Means followed by different letters within a column are significantly different at 5 percent level.
- Percentage of mortality is in parentheses.
Table 6. The LC50 and LC95 values of M. anisopliae var. anisopliae on larval and adult stages of B. longissima at 10 days after treatments
|
Stadia |
LC50 (conidia/ml) |
LC95 (conidia/ml) |
|
L3 |
5.1 x 102 |
2.0 x 104 |
|
L4 |
3.8 x 102 |
3.6 x 104 |
|
L5 |
8.6 x 102 |
6.2 x 104 |
|
Adults |
4.7 x 106 |
2.6 x 109 |
2.2. Test of different conidial concentrations of B. bassiana on different B. longissima stages
Beauveria bassiana used in this experiment was isolated from coffee pest, Hypothenemus hampei. Result of the analysis of variance on mortality of first instar larvae (L1), second instar larvae (L2), third instar larvae (L3), fourth instar larvae (L4) and adults at 20 days after treatments showed that there were highly significant differences among treatments (p <0.01). Mortality of L1 and L2 at the concentration of 5 x 103, 5 x 104 and 5 x 105 was not significantly different but the differences were observed at the control and 5 x 102 concentration. So, the lowest conidial concentration of B. Bassiana that caused highest mortality on L1 and L2 is 5 x 103 conidia/ml, 5 x 104 for L3 and L4 and 5 x 105 conidia/ml for adults (Table 7).
The LC50 and LC95 values of conidial concentrations of B. bassiana on larvae and adults of B. longissima at ten day are shown in Table 8 and the result showed that L1 was more susceptible than L2, L3, L4 and adults.
Table 7. Mortality of L1, L2, L3, L4, L5 and adults on different conidial concentrations of B. bassiana at 20 days after treatment (Hosang, 1996)
|
Conidial concentrations (conidia/ml) |
Larval stages |
Adults |
|||
|
1st |
2nd |
3rd |
4th |
||
|
Control |
3.75a |
2.50a |
2.25a |
3.25a |
2.50a |
|
5 x 102 |
11.25b |
6.25a |
4.25a |
5.25a |
2.50a |
|
5 x 103 |
20.00c |
14.50b |
5.25a |
13.25b |
7.00ab |
|
5 x 104 |
20.00c |
20.00b |
14.25b |
19.75c |
7.75b |
|
5 x 105 |
20.00c |
20.00b |
20.00c |
20.00c |
14.75c |
- Means followed by different letters within a column are significantly different at 5 percent level.
- Percentage of mortality is in parentheses.
Table 8. The LC50 and LC95 values of B. bassiana on larval and adults stages of B. longissima at 10 days after treatment
|
Stages |
LC50 (conidia/ml) |
LC95 (conidia/ml) |
|
L1 |
8.4 x 102 |
2.0 x 104 |
|
L2 |
3.1 x 103 |
6.6 x 104 |
|
L3 |
7.3 x 103 |
1.4 x 105 |
|
L4 |
5.8 x 103 |
8.4 x 104 |
|
Adults |
8.8 x 105 |
4.7 x 108 |
This is probably caused by integument of L1 being softer and thinner than that of the older larvae and adults. Therefore, B. bassiana can easily penetrate and infect the L1. The same thing is also observed by Sivasankaran et al. (1990) on Chilo infuscatellus where the second and third instar larvae of the pest were more susceptible to infection of B. bassiana than the older larvae.
Conidial concentration(s) used to control larvae of B. longissima in the field are 5 x 103 and 5 x 104 conidia/ml, while for adults is 5 x 105 conidia/ml. Therefore, effective concentration to control the pest in the field is the concentration that caused the highest mortality for both stages, 5 x 105 conidia/ml. This indicates that higher the conidial concentration, higher the pest mortality. Barson (1977) showed that mortality of S. scolytus larvae depends on the B. bassiana concentrations; lower the conidial concentrations, lower the larval mortality. The results demonstrated high potential of B. bassiana to be developed as a biological control agent of B. longissima in either partial or integrated control.
2.3. Experiment in the screen cages
The experiments adopted a completely randomized design (CRD) involving seven different application frequencies as treatments and replicated three times. The treatments were as follows: (1) spraying of B. bassiana suspension twice-yearly at two weeks interval, (2) spraying of B. bassiana suspension at two months interval, (3) spraying of B. bassiana suspension at three months interval, (4) spraying of M. anisopliae var. anisopliae suspension twice-yearly at two weeks interval, (5) spraying of M. anisopliae var. anisopliae suspension at two months interval, (6) spraying of M. anisopliae var. anisopliae suspension at three months interval and (7) control (Hosang et al., 1999).
The result showed that larval and adult population in the cages sprayed with the fungi suspensions were lower than that in the control. This result indicated that both M. anisopliae var. anisopliae and B. bassiana can be used to control B. longissima population in the field. Low population of the pest in the field is mainly caused by high rain fall. Kalshoven (1981) reported that dry season could trigger the development of population of Brontispa spp. in the field.
Based on the analysis of variance, there were significant differences among treatments at two to six weeks after treatments. Spraying of M. anisopliae var. anisopliae suspension twice at two weeks interval was different from the control but insignificant with the others. The result showed that these fungi can regulate the development of pest population in the field if field conditions are favourable for the growth and development of fungi. High rain fall not only affects the development of Brontispa but also the growth and development of M. anisopliae var. anisopliae and B. bassiana in order to control the pest in the field.
2.4. The effect of M. anisoplia var. anisopliae and B. bassiana on B. longissima in the field
Test of the effectiveness of M. anisoplia var. anisopliae and B. bassiana had been done in 1997-1998 and 1998-1999 in District Jeneponto, South Sulawesi. The experiments adopted a grouped randomized design with five treatments and replicated three times. The treatments were as follows: (a) spraying of M. anisopliae var. anisopliae suspension twice-yearly at two weeks interval and (b) spraying of M. anisopliae var. anisopliae suspension at three months interval, (c) spraying of B. bassiana suspension twice-yearly at two weeks interval, (d) spraying of B. bassiana suspension at three months interval and (e) without spraying of fungi, control.
Each plant was sprayed with fungi suspension at around ±100 ml. The unopened young leaves were subjected to spray because that part is usually attacked by Brontispa.
The preliminary observation was done by taking pest samples in the field. Result showed that all of the pest stages, eggs, larvae, pupae and adults were available in the field. The mean of B. longissima population were different at each plant. The total number of eggs, larvae, pupae and adults per plant were 0.87-4.73, 42.25-132.20, 3.77-10.93 and 13.87-47.47, respectively. The population of the pest highly reduced after treatments. The population of eggs, larvae, pupae and adults per palm per treatment four months after application (second observation) were as follows: A (0.47); B (1.66); C (12.54); D (1.77); and E (15.39). The next seven months, the pest populations were lower, except in control. Both M. anisopliae var. anisopliae and B. Bassiana are recommended to be used to control B. longissima. Introduction of these fungi reduced both pest population and plant damage. Spraying of these entomopthagenic fungi reduced the pest population at around 90.37-95.0 percent (Tumewan and Hosang, 1998). Pest population can be affected by rainfall, temperature and relative humidity. Dry season was occurred one month after treatment for four months, daily temperature was 26.35-31.15°C and relative humidity was 70-79 percent. Spraying of M. anisopliae var. anisopliae and B. bassiana suspension twice-yearly at two weeks interval or three months gave the same effect on the population of B. longissima in the field. In terms of the efficiency of spraying, the spraying of M. anisopliae var. anisopliae and B. Bassiana suspension twice-yearly at two weeks interval was suggested to regulate B. longissima in the field.
Conclusion
1.Pupal parasitoid, T. brontispa has high potential to be developed as a biological control agent of B. longissima. This parasitoid efficiently attack the pupae of B. longissima both in the laboratory and in the field. The percent parasitism under laboratory condition and in the field ranged from 76.7-87.0 percent and 35.71-73.56 percent, respectively.
2.Entomopathogenic fungi, M. anisopliae var. anisopliae and Beauveria bassiana can be used to control B. longissima.
3. M. anisopliae var. anisopliae can infect 100 percent larvae and 65 percent adults; while B. bassiana can infect 100 percent larvae and 73.75 percent adult under laboratory condition. Larval stage is more susceptible than the adult. The effective conidial concentration used to control B. longissima in the field is 5 x 105 conidia/ml. There is reduction of population in the field due to action of its natural enemies.
4.Spraying of M. anisopliae var. anisopliae and B. bassiana suspension can inhibit the development of B. longissima in the field. Spraying of M. anisopliae var. anisopliae and B. bassiana suspension twice-yearly at two weeks interval efficiently control B. longissima in the field. The level of palm damage can be reduced until about 90.37 to 95.07 percent seven months after application.
References
Anderson, T.E., D.W. Roberts and R.S. Soper. 1988. Use of Beauveria bassiana for supression of colorado potato beetle population in New York State (Coleoptera: Chrysomelidae). Environ. Entomol. 17(1):140-145.
Barbercheck, M.E. and H.K. Kaya. 1991. Competitive interaction between entomopathogenic nematodes and Beauveria bassiana (Deuteromycotina: Hyphomycetes) in soil borne larvae of Spodoptera exigua (Lepidoptera: Noctuidae). Environ. Entomol. 20(2):707-712.
Bing, L.A. and L.C. Lewis. 1991. Suppression of Ostrinia nubilalis (Hübner) (Lepidoptera: Pyralidae) by endophytic Beauveria bassiana (Balsamo) Vuillemin. Environ. Entomol. 20(4):409-432.
Burdeos, A.T. and L.T. Villacarlos. 1989. Comparative pathogenicity of Beauveria bassiana and Paecilomyces lilacinus to adult sweet potato weevil, Cylas formicarius (F.) (Coleoptera: Curculionidae). Philipp. Ent. 7(6):561-571.
Domsch, K.H., W. Gams and T.H. Anderson. 1980. Beauveria bassiana Vuill. Compendium of soil fungi. Acad. Press. 1-2:136-140.
Dunn, P.H. and B.J. Mechalas. 1963. The potential of Beauveria bassiana (Balsamo) Vuillemin as a microbial insecticide. J. Insect Pathol. 5:451-459.
Froggatt, J.L. dan B.A. O. Connor. 1941. Insect associated with the coconut palm. Pt II. New Guinea Agric. Gaz. 7:125-130.
Gallego, V.C. and C.E. Gallego. 1988. Efficacy of Beauveria bassiana Vuil. and Metarhizium anisopliae Mets. Sor. against tree coconut pest, Tirathaba rufivena Walk., Promecotheca cumingii Baly and Plesispa reichei Chapius. Annual Report. Agric. Res. PCA. 38-50.
Hoesni Heroetadji. 1989. Parasites of Plesispa reichei and Brontispa longissima in East Java. In: UNDP/FAO Integrated Coconut Pest Control Project. Annual Report 1989. Coconut Research Institute, Manado, North Sulawesi, Indonesia. 92-96.
Hosang, M.L.A. 1996. Patogenisitas Cendawan Beauveria bassiana (Balsamo) Vuillemin terhadap Brontispa longissima Gestro (Coleoptera: Hispidae). (Part of Master degree thesis). Jurnal Litri. 2(1):8-20.
Hosang, M.L.A., S. Sabbatoellah, F. Tumewan and J.C. Alouw. 1996. Musuh alami hama Brontispa longissima Gestro. Prosiding Seminar Regional Hasil-hasil Penelitian Tanaman Kelapa dan Palma Lain, Manado 19-20 Maret 1996. Buku I, 30-38.
Hosang, M.L.A., F. Tumewan and J.C. Alouw. 1999. Frekuensi dan interval waktu penyemprotan suspensi cendawan Beauveria bassiana dan Metarhizium anisopliae var. anisopliae terhadap hama Brontispa longissima. Prosiding Simposium Hasil Penelitian Tanaman Kelapa dan Palma Lain, Manado 10 Maret 1999. 28-40.
Kalshoven, L.G.E. 1981. The Pests of Crops in Indonesia. PT. Ichtiar Baru Van Hoeve, Jakarta. 701 pp.
Lever, R.J.A.W. 1969. Pests of the Coconut Palm. No. 18. FAO. Rome, Italy. 190 pp.
O'Connor, B.A. 1940. Notes of the coconut leaf hispid, Brontispa froggatti Sharp and its parasites. New Guenea Agric. Gaz. 6:36-40.
Ooi, P.A. and M.L.A. Hosang, 1989. Promecotheca cumingii Outbreaks in Central Sulawesi. In: FAO/UNDP Integrated Coconut Pest Control Project, Annual report, Coconut Research Institute, Manado, Indonesia, 97-103.
Ooi, P.A.C., Soekarjoto, S. Sabbatoellah, F. Tumewan, and R. Hoesni Heroetadji. 1989. Plesispa reichei and its parasitoid in Indonesia. In: UNDP/FAO Integrated Coconut Pest Control Project. Annual Report 1989. Coconut Research Institute, Manado, North Sulawesi, Indonesia. 85-91.
Sathiamma, B; C. Mohan, and M. Gopal. 2001. Biocontrol Potential and its Exploitation in Coconut Pest Management in Biocontrol potential and its exploitation in sustainable agriculture Vol. 2: Insect Pests edited by R.K. Upadhyay, K.G. Mukerji and B.P. Chamola. Kluwer Academic press. New York.
Soekarjoto, J.C. Alouw and J. Mawikere. 1994. Uji patogenisitas Metarhizium anisopliae terhadap hama Brontispa longissima Gestro. Buletin Balitka No. 22.
Tjoa, Tjien Mo. 1953. Memberantas hama-hama kelapa dan kopra. Noorhoff. Jakarta. 270 p.
Tumewan and M.L.A. Hosang. 1998. Pemanfaatan parasitoid dan patogen pada hama Brontispa longissima Gestro. Prosiding Konperensi Kelapa Nasional Kelapa IV. Bandar Lampung, 21-23 April 1998. 631-637.
Tumewan, F., J. Mawikere and M.L.A. Hosang. 1999. Pemanfaatan patogen serangga dalam pengendalian Brontispa longissima. Warta Penelitian dan Pengembangan Tanaman Industri. 12-14.
Waterhouse, D.F. and K.R. Norris. 1987. Biological Control Pasific Prospects. ACIAR. Inkata Press. Melbourne. 134-141; 211-218.
Wright, J.E. and L.D. Chadler. 1992. Development of a biorational mycoinsecticide: Beauveria bassiana conidial formulation and its aplication against boll weevil populations (Coleoptera: Curculionidae). J. Econ. Entomol. 85(4):113-1135.
Zelazny, B. 1989. Biological control of Oryctes rhinoceros with Metarhizium anisopliae. CRI. 7 pp.
Sathiamma, B; C. Mohan, and M. Gopal. 2001. Biocontrol potential and its exploitation in coconut pest management. In Biocontrol potential and its exploitation in sustainable agriculture Vol. 2: Insect Pests edited by R.K. Upadhyay, K.G. Mukerji and B.P. Chamola. Kluwer Academic press. New York.
Vilaysouk Khennavong*
* Director of Plant Protection Centre, Department of Agriculture, Ministry of Agriculture and Forestry, Vientiane, Lao PDR; e-mail: [email protected]
1. Country background
Lao PDR is located in the heat of the Indochina Peninsular, in Southeast Asia, latitude 14 to 23 degree north and latitude 100 to degree east.
Lao is landlocked country. It shares 505 km border with China to the north, 435 km with Cambodia to the south, 2 069 km with Viet Nam to the east 1 835 km with Thailand to the west, and 236 km with Myanmar to the north east. Lao PDR covers a total of 236~850 square kilometres three-quarters of which is mountain and plateau. The country has three distinct regions.
Coconut has been planted in Lao since ancient times until today. Even though, coconut cultivation is still not economic crop yet, it is a source of an additional income for farmers. Coconut is traditionally planted in villages and rural area around houses and big plantation for coconut is rare.
According to the weather condition and the experience of farmers, Lao has high potential to grow coconut in accordance with agro-processing industry. Therefore, for the last two years the Government has a policy focused on promoting coconut cultivation as economic crop for consumption and raw material for local agro-processing industry. In 2003-2004, the Government of Lao PDR has officially imported some coconut seed to grow in Champasack and Khammoun provinces.
2. Appearance and cause of Brontispa longissima outbreak on coconuts in Lao PDR
The coconut leaf beetle, Brontispa is native to Indonesia and it had been reported in other locations in the Asia-Pacific region. It is believed that this pest was introduced into southern Viet Nam a few years ago in shipments of ornamental palms. The beetle advanced rapidly into central and northern region of Viet Nam.
Since this report from Viet Nam, Brontispa has been reported in Hainan province in PR China, and Cambodia reported that the pest was found in late 2001 attacking coconut palms in provinces bordering Viet Nam. As in case of Lao PDR, the pest was found in the district bordering Viet Nam about 30 km and spread to other districts nearby.
After the coconut beetle outbreaks, Ministry of Agriculture and Forestry sent Lao and FAD technical team for field assessment. The survey showed that six villages in two provinces are facing the coconut beetles (Brontispa longissima) outbreak; these included Namthon village in Pakkading district of Bolikamxay province and villages Novilay, Vangboangtai, Vangboangnua, Fouang and Nabo in Sepon district of Savannaket province (Figure 1).
The losses due to coconut beetle outbreak in Lao PDR have not been estimated so far because of traditionally scattered nature of cultivation. However, it appears that coconut palms in Savannaket province have serious infestation. Therefore, close- by areas in Salavan, Champasack, Khammoun provinces and Vientiane capital are at high risk (Figure 2).

Figure 1: Sites of Brontispa longissima outbreak in Lao PDR
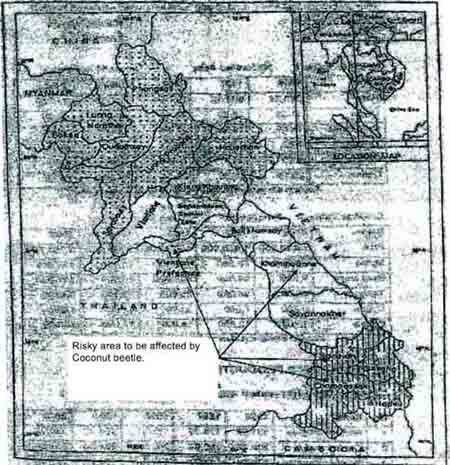
Figure 2: Areas threatened by spread of the coconut beetle
3. The main cause of Brontispa longissima outbreak in Lao PDR
The cause of beetle coconut outbreak (Brontispa longissima) in Lao is not clear. It is believed that some palm and coconut trees are imported illegally and these may have some infestation. The plant quarantine along the border is not so strict, which is primarily due to poor knowledge of the technical staff responsible for inspection of diseases and pests as well as the lack of equipment.
4. Some measures undertaken to overcome the coconut beetle outbreak in Lao PDR
Minister of Ministry of Agriculture and Forestry (MAF) announced to director of provincial, capital and special zone of agriculture and forestry to take following measures to combat the spread of coconut beetle outbreaks:
Pay attention to implement plant quarantine at every import and export check points around country as well as improve the capacity and knowledge of plant quarantine officials to inspect systematically.
Designate Bolikamxay and Savannaket province to be coconut beetle outbreak zone. No movement of plant material, especially palms or products of coconut and other palm from this area.
Facilitate plant protection technical implementation and inspect individual or concern sectors that are involved in coconut seeds, coconut palms or any plant related to palms passing Bolikamxay and Savannaket provinces must have certified paper from Provincial Agriculture and Forestry Department to certify origin or sources of product.
Province, Vientiane capital and special zone where no outbreak of coconut beetle has been noted, the Division of Agriculture must coordinate with local authority to monitor and conduct survey of coconut cultivation areas regularly and if any unexpected pests are detected these must be reported to MAF.
In order to overcome this pest, Ministry of Agriculture and Forestry has proposed and submitted letter to government regarding import of Asecodes hispinarum parasitoid. According to letter number three of International Phytosanitary Standard Measures (IPSM) of International Plant Protection Convention (IPPC), the Prime Minister office has allowed official notification to import, test, and introduce the natural enemy Asecodes hispinarum parasitoid from Viet Nam to rear and propagate in Plant Protection Centre, then release and inoculate to Pakkading district Khammoun province and Sepon district Savannaket province. Later, the Government will set an establishment of Asecodes hispinarum parasitoid in the two infested provinces. Moreover, MAF has requested emergency technical assistance from Viet Nam and international organizations in order to stop the coconut beetle outbreak in infested provinces and prevent further spread to other provinces.
Mat Hassan Othman*
* Plant Protection and Plant Quarantine Division, Malaysian Department of Agriculture, Kuala Lumpur; e-mail: [email protected]
1. Background
The coconut industry ranks fifth after oil palm, rubber, paddy, and fruits in terms of hectarage. However, it occupies only 2.4 percent or 151 044.7 of 6 269 909.9 hectares of the total cultivated land under the main crops in Malaysia. The coconut area is continually declining. It dropped from 213 14.0 hectares in 1985 to 151 044.7 hectares in 2001. The principal reason for the decline in hectarage being the low profitability. Many smallholders have changed over from coconut to oil palm mainly because of the poor returns.
Among the coconut holdings, about 69 percent are in Peninsular Malaysia, while the rest are in Sarawak (17 percent) and Sabah (14 percent). In Peninsular Malaysia, 70 percent coconut areas are located in the coastal region, cultivated mainly on alluvial soils of the west coast in Johore, Perak and Selangor and on bris (sandy) soils of the east coast.
About 92 percent or nearly 139 306 ha of coconut area are operated by smallholders and the average size of holding is about 1.1 ha. Due to the small size of holdings coupled with low yield and poor farm management, income received by the coconut smallholder has been very low. The net income being about RM 607 per year. In Malaysia, planting coconut as a mono-crop regardless of variety and management practices is not profitable enterprise.
2. Brief overview of coconut pests
The coconut palm is susceptible to attack of a large number of insects. In Malaysia, about 164 insects species have been associated with coconut palm (Ahmad Yunus and Ho Thian Hua, 1980). However, only a few of them are capable of causing considerable damage to the palm resulting in reduced growth and yield (Table 1). These pests include the Rhinoceros beetle (Oryctes rhinoceros Linnaeus), leaf-eating beetle (Promecotheca cumingii Baly and Plesispa reichei Chapuis), Artona catoxantha, nettle caterpillars (Setora nitens Walker and Thosea sinensis Walker), coconut spike moth (Tirathaba rufivena Walker), coconut skipper (Hidari irama), the Red Palm Weevil (Rhyncophorus schach Olivier), Parasa lepida Cramer, and coconut bagworms (Cremastopsyche pendula Joannis, Metisa plana Wlk, Mahasena corbetti Tams). The Rhinoceros beetle (Oryctes rhinoceros Linnaeus), leaf-eating beetle (Promecotheca cumingii Baly and Plesispa reichei Chapuis), Artona catoxantha are the major pests of concern. Widespread outbreaks are rare, but frequent localized and sporadic outbreaks of these pests have been reported (Tables 2, 3 and 4). During the outbreaks, serious damage has been observed on individual palm and small groups.
In general no control or minimum control measures were implemented when localized outbreaks occurred on mature palms. Surprisingly these pests disappeared after a time. Weather is one of the regulating factors. It was generally observed that most severe infestations become obvious in the dry season, and end with the onset of a wet season. Besides that, numerous natural enemies were recorded on various coconut pests and were assumed to play a major role in regulating coconut pest populations.
Table 1. List of common pests of coconut in Malaysia
|
No. |
Pest |
Pest Status |
Distribution |
||||||||||
|
Major |
Minor |
Wide spread |
Limited |
||||||||||
|
PM |
SR |
SA |
PM |
SR |
SA |
PM |
SR |
SA |
PM |
SR |
SA |
||
|
1 |
Rhinoceros beetle (Oryctes rhinoceros L) |
Ö |
Ö |
Ö |
|
|
|
Ö |
Ö |
Ö |
|
|
|
|
2 |
Two color coconut beetle, (Plesispa reichei Chapuis) |
Ö |
Ö |
Ö |
|
|
|
Ö |
Ö |
Ö |
|
|
|
|
3 |
Coconut leaf moth (Artona catoxantha) |
Ö |
|
|
|
Ö |
Ö |
Ö |
|
|
Ö |
Ö |
Ö |
|
4 |
Coconut leaf beetle Promecotheca cumingii |
Ö |
Ö |
|
|
|
Ö |
|
|
|
Ö |
Ö |
Ö |
|
5 |
Red stripe weevil, (Rhynchophorus schach Oliv.) |
|
|
|
Ö |
Ö |
Ö |
|
|
|
Ö |
Ö |
Ö |
|
6 |
Nettle caterpillar, (Setora nitens, Thosea sp.) |
|
|
|
Ö |
Ö |
|
|
|
|
Ö |
Ö |
Ö |
|
7 |
Coconut skipper, Hidari irava |
|
|
|
Ö |
Ö |
Ö |
|
|
|
Ö |
Ö |
Ö |
|
8 |
Coconut spike moth, Tirathaba rufivena |
|
|
|
Ö |
Ö |
Ö |
|
|
|
Ö |
Ö |
Ö |
|
9 |
Bagworm, Cremastopsyche pendula, Metisa plana Wlk, Mahasena corbetti Tams |
|
|
|
Ö |
Ö |
Ö |
|
|
|
Ö |
Ö |
Ö |
|
10 |
Coconut scale insect, Aspidiotus destructor |
|
|
|
Ö |
Ö |
Ö |
|
|
|
Ö |
Ö |
Ö |
Note: PM - Peninsular Malaysia; SR - Sarawak; SA - Sabah
3. Leaf-eating beetles
Currently two species of leaf-eating beetles, coconut leaf beetle (Plesispa reichi Chapuis) and Philippine leaf-miner (Promecotheca cumingii Baly) which belong to the sub-family hispinae of the family Chrysomelidae are the major pests of palms in Malaysia.
(a) Philippine leaf-miner Promecotheca cumingii Baly
Occurence and outbreaks
The Philippine leaf-miner was not recorded in Malaysia until the first outbreak of this pest on coconut palms occurred in Malacca in 1917 (Lever, R.A.W. 1951). The palms were badly attacked over considerable area but the beetle disappeared after a time (Gater, B.A.R. 1925). Since then no mention has been made of its presence until May 1972 when a major outbreak of the beetle on about 10 000 acres of coconut palms took place in province Wellesley and the southernmost part of Kedah (Ding, S.M. 1975). The infested palms were exclusively found on small land holdings. In this outbreak, it had been observed that the beetle besides infesting coconut palms also attacked nipah palm, oil palm, royal palm. The infestation on nipah was the most severe.
Table 2. Record of outbreaks/infestations of Rhinoceros beetle 1999-2003
|
Pest |
Year or date |
Host (Age of host) |
Status of infestation |
Location |
|
|
Planted area (Ha) |
Number Infested host |
||||
|
1. Oryctes rhinoceros L |
11/6/1999 |
Coconut |
15 ha |
15 palms |
Hilir perak |
|
10/4/2000 |
Oil Palm |
0.8 ha |
0.08 ha |
Perak Tengah |
|
|
24/4/2000 |
Coconut |
0.8 ha |
50 palms |
Dungun |
|
|
7/2/2001 |
coconut |
0.5 ha |
10 palms |
Kuala Muda |
|
|
2/4/2001 |
Coconut |
0.5 ha |
10 palms |
Kula Muda |
|
|
3/4/2001 |
Coconut |
0.75 ha |
NA |
Alor Gajah, Melaka |
|
|
18/6/2001 |
Coconut |
1 ha |
40 palms |
Kulim, Kedah |
|
|
24/9/2001 |
Coconut |
2 ha |
20 palms |
Temerloh, Pahang |
|
|
3/5/2002 |
coconut |
13 ha |
NA |
Raub |
|
|
5/2/2002 |
Coconut |
2.4 ha |
NA |
Perak Tengah |
|
|
16/12/2002 |
Coconut |
0.4 ha |
0.3 ha |
Perak Hilir |
|
|
8/1/2003 |
Coconut |
2 palms |
2 palms |
Kulim |
|
|
20/3/2003 |
Coconut |
5.8 ha |
3 palms |
Kota Bharu |
|
|
31/3/2003 |
Oil palm |
76 ha |
38 ha |
Seberang Perai |
|
|
31/3/2003 |
Coconut |
20 ha |
NA |
Kinta |
|
|
22/7/2003 |
Coconut |
4 ha |
NA |
Kerian |
|
|
30/12/2003 |
Coconut |
60 palms |
40 palms |
Rembau |
|
Note: NA - Not available.
Table 3. Record of outbreaks/infestations of leaf-eating beetle
|
Pest |
Year or date |
Host |
Status of infestation |
Location of infestation |
|
|
Planted area (Ha) |
Number Infested host |
||||
|
2. Plesispa reichei Chapuis |
4/3/2000 |
Coconut |
30 ha |
20 palms |
Telipot Kota Bharu, Kelantan |
|
7/4/2000 |
Coconut |
15 ha |
15 palms |
Pekan, Pahang |
|
|
22/6/2000 |
Coconut |
0.4 ha |
30 palms |
Pekan, Pahang |
|
|
12/6/2000 |
Coconut |
0.6 ha |
30 palms |
Kg. Kelulut, Marang, Terengganu |
|
|
26/7/2000 |
Coconut |
0.8 ha |
10 palms |
Marang, Terengganu |
|
|
2000 |
Coconut |
NA |
NA |
Tuaran, Sabah |
|
|
15/1/2001 |
Coconut |
0.2 ha |
20 palms |
Pasir Mas Kelantan |
|
|
16/2/2001 |
Coconut |
0.2 ha |
6 palms |
Rompin Pahang |
|
|
11/7/2001 |
Coconut |
5 ha |
5 palms |
Pekan, Pahang |
|
|
2001 |
Coconut |
NA |
NA |
Likas, Sabah |
|
|
22/1/2002 |
Coconut |
50 ha |
NA |
Raub, Pahang |
|
|
29/1/2002 |
Coconut |
1 ha |
NA |
Kluang Johor |
|
|
3/5/2002 |
Coconut |
13 ha |
NA |
Raub, Pahang |
|
|
17/10/2002 |
Coconut |
3 ha |
52 palms |
Pasir Mas, Kelantan |
|
|
23/1/2002 |
Coconut |
2 ha |
NA |
Temerloh, Pahang |
|
|
23/1/2002 |
Coconut |
200 ha |
2 000 palms |
Sabak Bernam, Selangor |
|
|
18/4/2002 |
Coconut |
34.4 ha |
1 800 palms |
Sabak Bernam, Selangor |
|
|
24/1/2002 |
Coconut |
200 ha |
25 ha |
Labuan, Sabah |
|
|
30/10/2003 |
Coconut |
0.5 ha |
100 palms |
Bera Pahang |
|
|
23/11/2003 |
Coconut |
7 palms |
6 palms |
Kota Star, Kedah |
|
|
29/11/2003 |
Coconut |
40 palms |
15 palms |
Kuala Lumpur |
|
|
29/11/2003 |
Manila palm |
35 palms |
10 palms |
Serdang, Selangor |
|
|
29/11/2003 |
Manila palm |
30 palms |
20 palms |
Bukit Jalil, Selangor |
|
|
29/11/2003 |
Manila palm |
15 palms |
5 palms |
Bandar Tun Razak, Kuala Lumpur |
|
|
3. Promecotheca cumingii |
1917 |
Coconut |
NA |
NA |
Malacca |
|
1972 |
Coconut |
NA |
10 000 acs |
Pulau Pinag |
|
|
1995/1996 |
Sago & Coconut |
13 115 |
Serious |
Sarawak |
|
Note: NA - Not available.
Table 4. Record of outbreaks/infestations of Artona catoxantha (2000-2003)
|
Pest |
Year or date |
Host |
Status of infestation |
Location of infestation |
|
|
Planted area (Ha) |
Number Infested host |
||||
|
4. Artona catoxantha |
23/5/2000 |
Coconut |
0.4 ha |
10 palms |
Pekan, Pahang |
|
1/6/2000 |
Coconut |
50 ha |
50 palms |
Kota Bharu, Kelantan |
|
|
14/6/2000 |
Coconut |
0.8 ha |
40 palms |
Rompin, Pahang |
|
|
7/7/2000 |
Coconut |
20 ha |
NA |
Mersing, Johor |
|
|
19/7/2000 |
Coconut |
16 ha |
NA |
Mersing, Johor |
|
|
6/2/2001 |
Coconut |
100 ha |
70 palms |
Manjung, Perak |
|
|
1/8/2001 |
Coconut |
NA |
NA |
Kuala Selangor, Selangor |
|
|
31/7/2002 |
Coconut |
1.5 ha |
1.5 ha |
Kuala Selangor, Selangor |
|
|
5/3/2003 |
Coconut |
20 ha |
20 ha |
Kluang, Johor |
|
|
8/10/2003 |
Coconut |
40 ha |
0.4 ha |
Muar, Johor |
|
|
3/11/2003 |
Coconut |
1.8 ha |
1.8 ha |
Pontian, Johor |
|
Note: NA - Not available.
Another major outbreak of Promecotheca cumingii occurred in Kuching and Samarahan districts in Sarawak in 1996 (Gumbek, M.,1999). About 13 000 hectares of coconut palms and 10 000 hectares of nipah palms were affected. The affected palms were located in the coastal areas and along Samarahan river. The initial pest infestation occurred on nipah palm (Nipa fruticans), but later it had spread to coconut palms (Coccos nuciferae), oil palms (Elaeis guineensis), sago palms (Metroxylon sagu), ornamental palms and other jungle palms.
Damage
The damage on the coconut palm is resulted from the feeding activity of both adult beetle and the larva on the leaflets. The adult beetles feed on the spongy tissue of the coconut leaflet by chewing a series of fine grooves on the lower surface, sparing a thin layer of the upper epidermis. Normally unattacked tissues in between the grooves die. In severe infestation, leaftips generally turned brown, shriveled and curled downward giving the palms a scorching appearance. All fronds are attacked but the older ones are more susceptible. The larva feeds and mines inside the leaf. It feeds on the parenchymatous leaving the upper and lower epidermis intact. In severe cases the coalescence of the larval mines and the adult feeding scars together with the natural drying up of unattacked tissue resulted in the death of extensive area of the leaflets. After a few generation of attack, the whole leaflet may be killed. In very severe infestation, except for a few young fronds, the whole palm assumed a burning appearance. Damaged leaflets consequently weaken the palm and reduce the yield. Premature fruit fall also occurred.
Table 5. Record of outbreaks/infestations of Promecotheca cumingii in Malaysia
|
No. |
Pest |
Year |
Host(Age of host) |
Status of infestation |
Location |
|
|
Planted area (Ha) |
Number Infested host |
|||||
|
1 |
Promecotheca cumingii Baly |
1917 |
Coconut |
NA |
NA |
Malacca |
|
1972 |
Coconut |
NA |
10 000 acs |
Pulau Pinag |
||
|
1995/1996 |
Sago & Coconut |
13 115 |
Serious |
Kuching/ Semarahan Sarawak |
||
Note: NA - Not available.
Cause of the outbreak
The Promecotheca cumingii is found in Malaysia since 1917. Except for three outbreaks reported in 1917, 1975 and 1996 respectively, the beetle has been otherwise under good natural control. It was suggested that outbreak could arise through several factors which are believed to upset the host-parasitoid equilibrium. These factors include favourable environmental conditions for beetle to multiply and low level or absence of natural enemies. When this occurs, rapid increase of beetle population takes place, resulting in outbreak.
Control measures
(i) Natural control
Outbreak of Promecotheca cumingii occurred occasionally and between outbreaks it would be difficult to determine the presence of this beetle. It is believed that natural control, particularly natural enemies seem to play the primary role in suppressing or terminating outbreaks and kept the pest in check. For example, in 1917 and 1975 outbreaks, the beetle died down or disappeared after a time without any chemical control (Gater, B.A.R., 1925; Ding, S.M., 1975).
Numerous natural enemies had been found to attack P. cumingii during the outbreak in province Wellesly in Peninsular and in Sarawak (Table 6). Of these natural enemies, P. parvulus and S. javanicus were the most common parasitoids. Of the two, S. javanica is normally present in numbers far exceeding that of P. parvulus and is most effective (Ding, K.M., 1975). It has attacked all the larval instars of the host while P. parvulus attack was limited mainly to the second and third instar larvae and pupae. During the outbreak it probably played an important role in suppressing the leaf-eating beetle.
During the outbreak in province Wellesley, it was found that unknown factors had caused a high larval mortality than the parasitoids (Ding, S.M., 1975). Adverse environmental conditions and overcrowding were attributed to the premature death of larvae.
(ii) Other control measures
Apart from natural control, the following measures have been employed for the control of the coconut leaf beetle:
Removal of attacked hosts/fronds
The destruction of heavily infested hosts/fronds (cut and burn) would reduce the population of the next generation.
Spraying and fogging
Table 6. Parasitoids of P. cumingii in Malaysia
|
No. |
Parasitoid Family |
Species |
Remarks |
Distribution |
|
|
PM |
SR |
||||
|
1 |
Eulophidae |
Sympiesis javanica |
Ectoparasitoid for larval stage, very common and most effective |
Ö |
Ö |
|
Pediobius parvulus |
Endoparasitoid for larval and pupal stages, very common |
Ö |
Ö |
||
|
Achrysocharis promecothecae |
Egg parasitoid (Peninsular) |
Ö |
- |
||
|
Pediobius anomalus |
Egg parasitoid, less common (Sarawak) |
- |
Ö |
||
|
Closterocerus sp. |
Egg parasitoid, less common (Sarawak) |
- |
Ö |
||
|
2 |
Brachonidae |
Adeshasp. |
Larval parasitoid |
Ö |
Ö |
|
3 |
Cleridae |
Callimerus arcutifer |
Predator. Attack egg, larva, pupa |
Ö |
- |
|
4 |
Hyphomycetes |
Beauveria bassiana |
Entomogenous fungus |
Ö |
Ö |
Note: PM - Peninsular Malaysia; SR - Sarawak
Ground spraying was not carried out during the outbreaks as it was difficult and not practical as most infested palms were very tall. In Sarawak, most of the infested areas, especially nipah palms could not be accessed through ground roads. Only fogging using propuxur was carried out in seriously infested coconut holding and the periphery of nipah areas.
Trunk injection
In Sarawak, trunk injections with 10 ml of methamidophos were carried out in areas with low infestation and the buffer zones.
Aerial spraying
Due to extensive areas infested, aerial spraying with Dipterex 95 Sp was carried out to control the outbreak in Sarawak. The spraying operation covered an area of 15 670 hectares. It was found to be effective in containing the pest infestation (Gumbek et al., 1996).
(b) Two coloured coconut leaf beetle (Plesispa reichei Chap.)
Occurrence and outbreak
The earliest record of P. reichei occurrence on coconut in Malaysia was in Johore in 1912 (Corbett, G.H., 1923). This species was found in abundance in Johore and elsewhere in Malaya. Both adults and larvae feed on the surface of the unopened leaf of coconut. Their attack is confined between the folds of tender leaves whilst they are still partially folded up.
Since it was first found, the P. reichei remained an insignificant pest as it was never reported to cause economic damage to coconut palms. But it has emerged as a major pest of coconut after the year 2000. The presence of this pest was reported throughout the country and sporadic outbreaks have occurred in various places on coconut and other ornamental palms (Table 3). In areas where an outbreak occurred, a serious damage has been observed on individual palm and small groups.
Damage
Both adults and larvae live and feed on partially unfolded coconut leaflets. The damage on the coconut palm is the result from the feeding activity of both adult beetle and the larva on the leaflets. They remove strips of tissue from both sides of the leaves. They feed in a straight line and parallel to each other. Later, these feeding lines mingle with each other so that the remaining tissue dries and rots. In severe infestation, leaflets generally turn brown giving the palms a scorching appearance. When the attack is severe and of long duration, the palms may die.
Cause of the outbreak
P. reichei is found in Malaysia since 1917. Only recently, this pest has emerged as serious pest of coconut palms. Ornamental palms, particularly coconut and Manila palms that are planted for landscape in cities and golf courses were severely attacked. The emergence of this pest could arise through several factors which are believed to upset the host-parasitoid equilibrium. These factors include favourable environmental conditions for beetle to increase and low level or absence of natural enemies. When this occurs, rapid increase of beetle population takes place, resulting in outbreak.
Control measures
(i) Natural control
It is believed that natural control, particularly natural enemies is playing the primary role in suppressing outbreaks and keeping the pest in check. In many outbreaks, it has been observed that the infested plants recovered and the beetle died down or disappeared after a time without any control action. So far there is no information available on the natural enemies attacking this pest in Malaysia.
(ii) Cultural practices
Cutting and burning of heavily infested fronds may reduce pest population.
(iii) Chemical application
During severe infestation, insecticide application is required to help quick knockdown of pest. Following are the commonly used control methods:
Insecticide spraying
Sprays were generally effective for young palms (shorter than 2.5 metre). Foliar spraying with dimethoate or mixture of chloropyrifos and cypermethrin were found to be effective against leaf beetle (Choo-Toh,1999; Sivapragasam et al., 2004).
Soil drenching
For tall palms soil drenching or trunk injection with systemic insecticides were recommended to control the pest. Drenching around the palm base with imidacloprid at 10 ml in 2000 ml water/palm was found to be the most effective. The palms begin to produce healthy new fronds after 30 days of treatment.
Trunk injection
10-15 ml of systemic insecticides such as monocrotophos or methamidophos is injected undiluted into a hole (10-15 cm deep and 1.5 cm wide) bored in the palms trunk half a metre above the ground. After the injection, the hole is sealed up with clay or bitumin.
4. Proposal
Coconut leaf-eating beetles particularly P. reichei have become important pests in Malaysia. Frequent and sporadic infestations of P. reichei beetle have been reported on the coconut and ornamental palms throughout the country. This beetle is becoming a serious pest in other countries in this region particularly in Singapore. As this pest was relatively unknown previously, very little is known about biology and ecology and effective control measures. More collaborative efforts, among affected countries, are required to develop effective short and long terms control measures of this pest.
5. References
Ahmad Yunus and Ho Thian Hua. 1980. List of Economic Pests, Host Plants, Parasites and Predator in West Malaysia, Bulletin No. 153. Ministry of Agriculture Malaysia.
Bayer Agrochem. 2/2001. The green fever.
Choo-Toh, G.T. 1999. An outbreak of Plesispa reichei Chapuis on palms in Singapore. In Sivapragasam et al. (eds.) Proceedings of the 5th International Conference on Plant Protection in the Tropics, 15 to 18 March 1999, Kuala Lumpur, Malaysia, pp. 390-393.
Corbett, G.H. 1923. Preliminary note on the presence the two-colored coconut leaf beetle (Plesispa reichei Chap.), Malay. Agric. Journal 11: pp. 64-69.
Department of Agriculture Malaysia. 2001. Crop Hectareage Statistic.
Ding, S.M. 1975. Outbreak of Promecotheca cumingii Baly on coconut palms in Province Wellessley, Malaysia. Malaysian Agric. Journal, Vol. 50: pp. 200-220.
Gater, B.A.R. 1925. Malayan Agric. Journal, Vol. 13 pp. 160-161.
Gumbek, M. 1999. Outbreak of coconut leaf miner, Promecotheca nuciferae in Sarawak. In Sivapragasam et al. (eds.) Proceedings of the 5th International Conference on Plant Protection in the Tropics, 15 to 18 March 1999, Kuala Lumpur, Malaysia, pp: 390-393.
Gumbek, M. and A.Y.A. Hassan. 1999. Aerial spraying of coconut leaf miner, Promecotheca nucifera in Sarawak., The Sarawak Meseum Journal, pp. 85-108.
Lever, R.A.W. 1951. Malay. Agricultural Journal, Vol. 34:79-82.
Sivapragasam A. and W.H. Loke. 2004. Plesispa reichei (Chapuis) (Fam.: Hispidae): a sporadic but important pest of coconut palms in Malaysia (Unpublished).
Yunus, A. and A. Balasubramaniam. 1981. Major crop pests in Peninsular Malaysia.
Bulletin No. 138, Agriculture Division, Ministry of Agriculture.
![]()
![]()
![]()