![]()
![]()
![]()
M. Cresta, J. Périssé, M. Autret
and
E. Lombardo
In a previous paper we dealt with the different protein values of various types of human diets (1). A very careful determination of a protein quality based on the amino acid content is particularly important when it is not possible to apply methods of biological measurement systematically. This quality factor should be taken into account both in assessing protein requirements and in shaping food policy.
The table "Amino Acids Content of Foods and Biological Data on Proteins" (2), compiled by the Nutrition Division of FAO, gives a number of biological data derived from tests carried out on rats fed with experimental diets, from which the essential amino acid content can be calculated a posteriori. We came to the conclusion that a study of the correlation of the biological with the chemical measurements, as well as an attempt to assess the role of the various reference proteins which have been used up to now, would prove of value in establishing a protein quality index. An IBM 1130 computer was used for all statistical calculations.
In 1946 Mitchell and Block proposed a method for assessing the quality of nutritional proteins by means of an index. (3) This method, which is still widely used, consists in calculating by the use of tables the quantity of each essential amino acid contained in the protein mixture. The values so deduced are then expressed as percentage of the amino acid content of a similar quantity of egg protein. The amino acid which shows the lowest percentage is called a limiting factor, and the percentage so obtained is the protein score 1. To prove the merits of this method, the authors showed the results of a correlation study between the chemical score (CS) established for 23 different foods and their biological value (BV) obtained from nitrogen balances.
The amino acid content of the foods and the egg protein used as reference had been determined previously by various investigators using chemical and micro-biological methods. The data referring to biological value were extracted mainly from the findings of the University of Illinois on young growing rats. By comparing these two sets of dats, Mitchell and Block ascertained that the representative values of the chemical score and of the biological value showed an extremely satisfactory coefficient of correlation (r =-0.86) 1, which (in their opinion) confirmed both the validity of adopting the chemical score for grading the quality of proteins and the hypothesis whereby the protein synthesis is limited, in relation to the requirements of the organism, by those amino acids which are present to a minor degree.
On the other hand, in the scatter diagram, the regression line representing the set of points cuts the ordinate at such a value that some proteins, although showing a zero chemical score (through the absence of one or more essential amino acids), could nevertheless have a definite biological value. The authors then put forward the hypothesis that "proteins inadequate for growth, but possibly adequate for maintenance, would possess a biological value of 39".
Mitchell and Block followed up this line of research and, in 1946-47, published a further study on the correlation of the chemical score with the protein efficiency ratio (PER), based on 23 different foods (4). The PER values were obtained by various investigators who fed rats for periods of four to eight weeks on diets containing from 8 to 12 percent of proteins. The coefficient of correlation established between the two series of data was high (r = -0.89). It was comparable to the coefficient obtained previously by correlating chemical score with biological values; in this latter case, however, results showed a degree of dispersion (8 percent of standard error in the regression equation) which was less than that obtained with the protein efficiency ratio (16 percent).
The interrelationship existing between chemical and biological measurements of protein quality also attracted the attention of the FAO Committee set up in 1955 to define protein requirements (5). The Committee examined the concordance between the two variables on 30 different foods. The biological values were established on adult men and young growing rats, while the chemical score was calculated with reference to the "provisional amino acid pattern" proposed by the Committee itself (FAO reference protein). A proper mathematical correlation study was not carried out, but nevertheless the scatter diagram showed a high degree of dispersion among the data. The Committee came to the conclusion that "complete concordance between protein scores and biological values is not to be expected in view of the incompleteness of present knowledge of the amino acid requirements of man, and of the amino acid values of foods".
The problem was re-examined in 1963 by the Joint FAO/WHO Expert Group which was entrusted with the revision of protein requirements. The Group investigated 26 foods, comparing the chemical scores with the net protein utilization (NPU), as determined on rats (6). The results showed numerous discrepancies, which the Group ascribed to the fact that biological tests and chemical analyses had been carried out on different samples and that, as a result, the measurements were liable to error. The chemical score does not, in fact, show any possible imbalance between amino acids, and there is the risk of overestimating the values of certain amino acids which are not available.
As we shall see, another (and not unimportant) reason for these inconsistencies lay in the choice of reference protein (FAO pattern, egg, milk) used to calculate the chemical score.
From the data contained in the table "Amino Acids Content of Foods and Biological Data on Protein", 339 diets of different composition were selected. These diets were used by various investigators to determine protein quality in rats. 87 of the diets were used in connexion with the biological value (BV), 187 with the protein efficiency ratio (PER) and 65 with net protein utilization (NPU). The amino acids content of each diet was determined using the above-mentioned table, taking into account the composition of the foods contained in the various diets. The source of nitrogen for the diets was either one food only, or several foods (in both cases with or without the addition of amino acids).
Only diets with a protein level of between 8 and 12 percent were considered. Of these, only diets with a protein content of 10 percent were kept for study of the NPU. In view of the standard composition of the experimental diets, this corresponds to a protein calorie ratio of around 10 percent.
Diets supplemented with high amounts of amino acids (more than 0.4 percent of lysine or more than 0.1 percent of methionine) were discarded, since at these levels certain side effects (linked to amino acid imbalance and to possible toxic effects) can possibly interfere and become a cause of heterogeneity. The number of experiments considered in the two sets of results is markedly greater than that considered in previous studies, thus allowing a significant statistical analysis.
The chemical score of each diet was calculated by the Mitchell and Block method, using various reference proteins whose composition is given in Table 1.
Essential amino acids content of various reference proteins
expressed in mg per g of nitrogen
| Egg 1946 (1) | Egg 1963 (2) | Egg 1969 (3) | Human milk (4) | Cow milk (5) | FAO Pattern (6) | |
| Isoleucine | 500 | 415 | 393 | 254 | 295 | 270 |
| Leucine | 575 | 553 | 551 | 548 | 596 | 306 |
| Lysine | 450 | 403 | 436 | 428 | 487 | 270 |
| Sulphur * A/A | 406 | 346 | 362 | 185 | 208 | 270 |
| Aromatic A/A ** | 675 | 627 | 618 | 421 | 633 | 360 |
| Threonine | 306 | 317 | 320 | 280 | 278 | 180 |
| Tryptophan | 94 | 100 | 93 | 105 | 88 | 90 |
| Valine | 456 | 454 | 428 | 284 | 362 | 270 |
| * Methionine + cystine ** Phenylalanine + tyrosine | ||||||
(1) Mitchell, see reference (3), page 2.
(2) Report of Joint FAO/WHO Expert Group (1963), Table 6B.
(3) Amino acids content of foods and biological data on proteins. Average data by column chromatography.
(4) Report of Joint FAO/WHO Expert Group (1963), Table 6B.
(5) Report of Joint FAO/WHO Expert Group (1963), Table 6B.
(6) Report of the FAO Committee on Protein Requirements (1955).
The chemical score was also calculated in terms of egg protein (1963), using, however, the method of calculation proposed in the report of the Joint FAO/WHO Expert Group (appendix 2); this method takes into consideration the total content of essential amino acids (A/E method).
Table 1 calls for some comments. Compared to the 1963 data, the egg protein (1946) proposed by Mitchell 1 gives 10 or 15 percent higher values for isoleucine, lysine and sulphur amino acids. The levels of isoleucine and sulphur amino acids are perceptibly lower in cow milk and in human milk proteins than they are in egg. The level of amino acids in the FAO provisional pattern is, generally speaking, lower than in either milk or egg. In view of these variations, it might be expected that both the type of limiting amino acid and the value of the chemical score would vary according to the choice of reference protein. In other words, if egg protein is adopted as reference, its high content of sulphur amino acids will tend to show methionine-cystine as the limiting factor in diets, and the chemical score will be lower than when milk is used as reference protein. Conversely, if cow milk protein is adopted as reference, its high content of lycine will increase the number of diets for which this amino acid is the limiting factor and the deficit will be more pronounced than if egg or human milk were used.
The reference protein pattern was thus devised in order to minimize these disadvantages. The low level fixed for most of the amino acids forming this reference protein does, in fact, diminish the deficits and penalize diets to a lesser extent. We shall have the opportunity later on to consider whether this reference protein does in fact reflect actual biological reality more closely.
Table 2 shows, for the three biological measurements considered (biological value, protein efficiency ratio, net protein utilization) the frequency of occurrence of limiting amino acids according to the nature of the reference protein and the coefficient of correlation between the biological measurements and the chemical score.
As might have been expected, nearly half of the diets are limited by methioninecystine when the various egg proteins are used as references, while the same amino acids are rarely limiting when cow milk and human milk are adopted as references. Conversely, lysine is a limiting factor in half the diets when milk is adopted as reference and only in a quarter of the diets if egg is used as the reference.
Likewise isoleucine is sometimes a limiting factor when egg is used and virtually never when milk is used. The converse phenomenon is observed for leucine.
The type and frequency of occurrence of the limiting amino acids with the FAO pattern are rather similar to the results obtained with egg protein, but the chemical scores are higher than when other reference proteins are used.
The last column in Table 2 shows the coefficients obtained by correlating biological data (BV, PER, NPU) with the protein scores on the basis of the different reference proteins.
The chemical scores obtained on the basis of the FAO pattern show a very low correlation with the biological value (r = 0.38). In the same way, the A/E method of calculating (in terms of egg proteins) the chemical score, recommended by the joint FAO/WHO Expert Group, gives an even lower coefficient of correlation (r = 0.29). The highest coefficients of correlation are obtained with human milk protein and, above all, with cow milk protein (r = 0.67). The data contained in this table do not, however, confirm the very good coefficient of correlation established by Mitchell and Block on 23 diets with egg as reference protein; as we have seen, the investigators carried out far fewer experiments and, moreover, used an amino acid pattern which was either incomplete (the value of cystine not having been taken into consideration) or based on methods some of which are technically obsolete (chemical methods).
The coefficients of correlation of the chemical score with the PER are higher than those for the biological value. The coefficient is more than 0.60 which ever reference protein is adopted, but the highest value is obtained when egg protein is used as reference protein.
The coefficients of correlation between the chemical score and the NPU are higher than those obtained with the previous biological measurements, whatever reference protein is used. However, among the various reference proteins used, egg produces outstanding correlations (r > 0.80), greater than those obtained by using milk or the FAO pattern as a reference.
This study is an attempt to ascertain whether a mathematical formula instead of the chemical score would not give a protein quality index closer to biological reality, and whether the respective importance of each essential amino acid with respect to the different biological measurements (BV, PER, NPU) can be measured by a suitable test.
Table 2 - COEFFICIENTS OF CORRELATION BETWEEN CHEMICAL SCORE AND BIOLOGICAL VALUE AND FREQUENCY OF OCCURENCE OF LIMITING AMINO ACIDS ACCORDING TO VARIOUS REFERENCE PROTEINS
| BIOLOGICAL VALUE (87 experiments) | ||||||||||
| ISOL. | LEUC. | LYS. | SULPHUR A/A | AROMATIC A/A | THREO | TRYPT. | VAL. | C.S.100 | COEFFICIENT OF CORRELATION | |
| Egg 1946 | 3 | — | 24 | 53 | — | 1 | 3 | 3 | — | 0.383 |
| Egg 1963 | 15 | — | 21 | 41 | — | 4 | 6 | — | — | 0.501 |
| Egg 1969 | 5 | — | 25 | 50 | — | 2 | 5 | — | — | 0.524 |
| Human milk | — | 3 | 44 | 2 | — | 22 | 16 | — | — | 0.619 |
| Cow milk | — | 10 | 51 | 10 | — | 12 | 3 | — | 1 | 0.673 |
| FAO Pattern | 2 | 1 | 18 | 40 | — | 21 | 3 | — | 2 | 0.378 |
| PROTEIN EFFICIENCY RATIO (187 experiments) | ||||||||||
| ISOL. | LEUC. | LYS. | SULPHUR A/A | AROMATIC A/A | THREO | TRYPT. | VAL. | C.S.100 | COEFFICIENT OF CORRELATION | |
| Egg 1946 | 73 | — | 35 | 79 | — | — | — | — | — | 0.710 |
| Egg 1963 | 44 | — | 44 | 86 | — | 4 | 3 | 6 | — | 0.726 |
| Egg 1969 | 20 | — | 52 | 106 | — | 4 | 2 | 3 | — | 0.678 |
| Human milk | — | 2 | 93 | 3 | — | 14 | 75 | — | — | 0.647 |
| Cow milk | — | 9 | 149 | 7 | — | 3 | 4 | 15 | — | 0.656 |
| NET PROTEIN UTILIZATION (65 experiments) | ||||||||||
| ISOL. | LEUC. | LYS. | SULPHUR A/A | AROMATIC A/A | THREO | TRYPT. | VAL. | C.S.100 | COEFFICIENT OF CORRELATION | |
| Egg 1946 | 22 | — | 13 | 28 | — | — | 2 | — | — | 0.827 |
| Egg 1963 | 14 | — | 15 | 27 | — | — | 6 | 3 | — | 0.826 |
| Egg 1969 | 9 | — | 18 | 33 | — | — | 3 | 2 | — | 0.820 |
| Human milk | — | 3 | 29 | 1 | — | — | 32 | — | — | 0.732 |
| Cow milk | 1 | 8 | 39 | 9 | 2 | — | 5 | 1 | — | 0.721 |
| FAO Pattern | 6 | — | 11 | 31 | — | — | 11 | — | 6 | 0.756 |
The analysis relates to the diets previously investigated. Tables 3, 4 and 5 show the results of the study of multiple regression as between the essential amino acids in the diets and the biological data.
The coefficients of correlation resulting from this multiple regression study are given in Table 6 and are compared with the maximum values previously obtained by simple correlation. In both cases, the values of "r" improve as we pass from the BV to the PER and to the NPU. For the biological value, a multiple correlation coefficient (r = 0.71) is obtained, greater than that produced by simple correlation (r = 0.67), while for the PER and NPU, the multiple regression formulae show a good estimated value which is virtually comparable to that obtained by the classical method of Mitchell and Block, provided that for the latter the chemical score is calculated by reference to 1963 egg protein.
Tables 3, 4 and 5 (column 7) show the values of Student's variable t for the different essential amino acids as these emerge from the studies of multiple correlation with the biological value, protein efficiency ratio and net protein utilization.
From the statistical standpoint, Student's variable t is defined as the ratio between the multiple regression coefficient of each amino acid and the standard deviation of this coefficient. This test measures in probability terms the importance assumed by a given independent variable (amino acid) in the multiple regression; it therefore enables the relative importance of each essential amino acid in the determination of biological measurements to be assessed.
Lysine appears to play an important part (t = 3.5) with respect to the biological value. Then come, in order of importance as given by the values of Student's variable t, tryptophan and leucine. The preponderance of lysine (t = 5.2) appears still more evident with respect to the PER. There follow the sulphur amino acids which are also of relative importance (t = 3.5), followed by valine. Lysine has an important role (t = 2.30) in the determination of the NPU, but here the sulphur amino acids take the leading place (t = 3.57) and have a preponderant importance. Tryptophan and the aromatic amino acids also have high values.
The role ascribed to certain amino acids by Student's variable t would appear to explain why some reference proteins give chemical score values which are better correlated than others with the results of the BV, the PER and the NPU.
In a given series of observations, some amino acids can show, insofar as they are limiting factors, a high frequency of occurrence although they are not of relative importance in the study of multiple regression. The role of these amino acids can thus be erroneously over-estimated, and the diet under examination is consequently penalized by the fact that the reference protein is not of adequate composition. These limiting amino acids (or rather, amino acids which are identified as such) can thus become misleading factors in the prediction of biological quality of protein from chemical score.
For this reason, only a poor correlation can be obtained between the two measurements. For example, when simple correlation between biological value and the chemical score calculated from egg, is studied, sulphur amino acids are frequently limiting (see Table 2) since egg is rich in methionine-cystine. Student's variable t shows that sulphur amino acids play a secondary role with regard to biological value (Table 4). The chemical score of these amino acids will thus interfere with the simple correlation, with the result that the correlation coefficient will drop.
On the other hand, sulphur amino acids show a reduced frequency of occurrence when milk is used as a reference protein, since milk is less rich in sulphur amino acids. The role of the sulphur amino acids, therefore, diminishes in importance and falls more in line with the results of Student's test t. As a consequence, it is not surprising that milk as a reference protein should give a better correlation with the biological value than egg.
Multiple regression between Biological Value (BV) essential amino acids and the
total of non-essential amino acids
(87 diets)
| Average mg/g N | Standard deviation | Coefficient of variation | Correlation coefficient for each A/A with BV | Coefficient of regression | Standard deviation of regression coefficient | Student's t for each amino acid | |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Isoleucine | 257 | 34 | 0.131 | 0.45352 | - 0.07897 | 0.05086 | - 1.55257 |
| Leucine | 462 | 75 | 0.163 | 0.28688 | 0.03628 | 0.01680 | 2.15943* |
| Lysine | 319 | 124 | 0.389 | 0.62374 | 0.06267 | 0.01780 | 3.52109** |
| Sulphur A/A | 211 | 54 | 0.255 | 0.10462 | 0.02171 | 0.02393 | 0.90708 |
| Aromatic A/A | 491 | 62 | 0.126 | 0.15902 | - 0.01694 | 0.02312 | - 0.73268 |
| Threonine | 223 | 40 | 0.180 | 0.62105 | 0.02649 | 0.05506 | 0.48118 |
| Tryptophan | 71 | 14 | 0.202 | 0.17842 | 0.20699 | 0.08984 | 2.30395* |
| Valine | 301 | 45 | 0.149 | 0.40615 | 0.01048 | 0.02891 | 0.36282 |
| Non-essential A/A | 3664 | 312 | 0.085 | - 0.39687 | - 0.00079 | 0.00468 | - 0.17007 |
* p = 0.05
** p = 0.01
Multiple regression between Protein Efficiency Ratio (PER)
and essential amino acids
(187 diets)
| Average mg/g N | Standard deviation | Coefficient of variation | Correlation coefficient for each A/A with PER | Coefficient of regression | Standard deviation of regression coefficient | Student's t for each amino acid | |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Isoleucine | 256 | 38 | 0.147 | 0.48144 | 0.00237 | 0.00126 | 1.88383 |
| Leucine | 496 | 89 | 0.180 | 0.08852 | - 0.00046 | 0.00048 | - 0.96459 |
| Lysine | 305 | 101 | 0.333 | 0.50204 | 0.00214 | 0.00041 | 5.19566** |
| Sulphur A/A | 217 | 59 | 0.273 | 0.39206 | 0.00249 | 0.00071 | 3.48082** |
| Aromatic A/A | 516 | 40 | 0.078 | 0.32615 | 0.00018 | 0.00102 | 0.18024 |
| Threonine | 224 | 57 | 0.256 | 0.27132 | 0.00033 | 0.00065 | 0.51645 |
| Tryptophan | 74 | 14 | 0.184 | 0.49798 | 0.00587 | 0.00379 | 1.54838 |
| Valine | 303 | 34 | 0.113 | 0.58927 | 0.00462 | 0.00168 | 2.74326** |
** p = 0.01
Multiple regression between Net Protein Utilization (NPU) and
essential amino acids
(65 diets)
| Average mg/g N | Standard deviation | Coefficient of variation | Correlation coefficient for each A/A with NPU | Coefficient of regression | Standard deviation of regression coefficient | Student's t for each amino acid | |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Isoleucine | 268 | 47 | 0.175 | 0.56147 | 0.04847 | 0.05876 | 0.82495 |
| Leucine | 476 | 86 | 0.181 | 0.32519 | 0.00639 | 0.02268 | 0.28199 |
| Lysine | 350 | 137 | 0.393 | 0.41470 | 0.03032 | 0.01316 | 2.30333* |
| Sulphur A/A | 220 | 63 | 0.288 | 0.53862 | 0.09035 | 0.02528 | 3.57323** |
| Aromatic A/A | 525 | 64 | 0.121 | 0.46113 | 0.05715 | 0.02734 | 2.09034* |
| Threonine | 234 | 39 | 0.167 | 0.54339 | - 0.04125 | 0.07572 | -0.54481 |
| Tryptophan | 75 | 17 | 0.234 | 0.58149 | 0.22008 | 0.10043 | 2.19120* |
| Valine | 327 | 78 | 0.237 | 0.37008 | 0.01256 | 0.01893 | 0.66345 |
* p = 0.05
** p = 0.01
Table 6 - MULTIPLE REGRESSION EQUATIONS AND COMPARISON BETWEEN MULTIPLE AND SIMPLE CORRELATION COEFFICIENTS
(AMINO ACIDS IN MG/G OF NITROGEN)
| BIOLOGICAL VALUE = 32.832 Isol. 0.07897 + Leuc 0.03628 + Lys 0.06267 ? Sulphur A/A 0.02171 - Aromatic A/A 0.01694 + Threon. 0.02649 + Trypt. 0.20699 + Val. 0.01048 + Non-essential A/A 0.00079 | ||
| PROTEIN EFFICIENCY RATIO = - 1.2689 Isol. 0.00237 - Leuc. 0.00046 + Lys. 0.00214 + Sulphur A/A 0.00249 + Aromatic A/A 0.00018 + Threon. 0.00033 + Trypt. 0.00587 + Val. 0.00462 | ||
| NET PROTEIN UTILIZATION = - 24.55673 + Isol. 0.04847 + Leuc. 0.00639 + Lys. 0.03032 + Sulphur A/A 0.09035 + Aromatic A/A 0.05715 - Threon. 0.04125 + Trypt. 0.22008 + Val. 0.01256 | ||
| MULTIPLE CORRELATION COEFFICIENT | SIMPLE CORRELATION COEFFICIENT (maximum value) | |
| BIOLOGICAL VALUE | 0.707 | 0.673 |
| PROTEIN EFFICIENCY RATIO | 0.724 | 0.726 |
| NET PROTEIN UTILIZATION | 0.787 | 0.827 |
The same line of reasoning can be used to explain the quality of simple correlations between the chemical score of the various proteins and the PER or NPU. Student's test shows that sulphur amino acids are here of relative importance. The use of egg as a reference protein in preference to milk will give rise to a high frequency of occurrence of sulphur amino acids as a limiting factor. The choice of this protein will give to sulphur amino acids an importance which is more in line with the data obtained from Student's test t. As a result, the correlations between the chemical score and the PER or NPU will be better with egg than with milk. The same general argument holds good for all other amino acids. The effect that they may have on the correlation will depend on their level in the reference protein, which will thus determine the frequency of occurrence of the limiting factor. The role of these amino acids will also depend on their relative importance as indicated by Student's variable t.
Calculations support this line of reasoning. We have seen that the coefficient of correlation between the biological value and the chemical score is low (r = 0.50) when egg (1963) is used as reference. Let us now take it that the sulphur amino acids are eliminated as a limiting factor and that the biological value is correlated with the chemical score of the secondary limiting amino acid. In this way the influence of the sulphur amino acids is minimized, in full accordance with the minor role indicated for them by Student's variable t in relation to biological value. The relevant calculations show that, in this case, the coefficient of correlation does in actual fact improve and that its value increases from 0.50 to 0.60.
In the case of the PER and NPU, however, Student's test t shows that sulphur amino acids are of considerable importance. If we eliminate them as before and only take into account the chemical score of the secondary limiting amino acid, it follows logically that the correlation between the two variables will decrease. This, in fact, is what is shown by the calculations. The coefficient of correlation which, for the PER, was 0.73 changes to 0.62 and for the NPU from 0.83 to 0.68.
Here, therefore, is the proof that some amino acids play different parts according to whether one refers to biological value, PER or NPU. Their respective importance can be measured by means of a multiple regression analysis using Student's test t.
Whenever the chemical score is used to estimate the ability of a food protein to maintain the nitrogen balance (BV), the reference proteins which will give the best correlation will be those with a high level of lysine, tryptophan and leucine.
According to these data, the pattern of milk protein is more acceptable than egg protein. However, neither cow milk or human milk seems to have an optimum amino acid composition with respect to biological value. Cow milk is richer than human milk in leucine and lysine (positive aspect) but is poorer in tryptophan (negative aspect). Neither completely meets the requirements indicated by the multiple correlation. In this respect, the multiple regression equation (Table 3) appears to give an estimate of the BV which comes closer to biological reality.
When the chemical score is used for estimating the capacity of food proteins to meet growth requirements, the reference protein which will lead to the best correlation with measurements of changes in body weight (PER) should be selected from among those with high levels of lysine, sulphur amino acids and valine. Likewise, if it is desired to obtain a protein score which shows good correlation with nitrogen retention measurements (NPU), the reference protein should have a high level of sulphur amino acids, lysine, tryptophan and aromatic amino acids. The calculation has shown that egg protein is the reference protein which best meets these requirements.
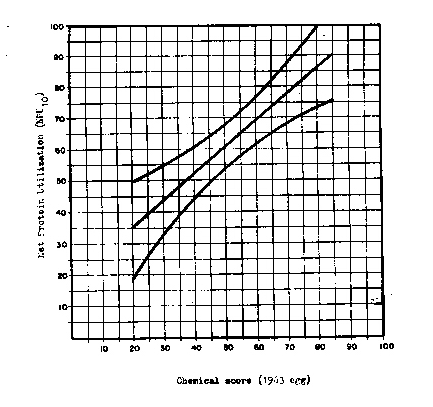
On a practical level, it should be noted that, of the three biological measurements considered, the chemical score shows the highest degree of correlation with the NPU. This should not be interpreted as a criterion of NPU quality and does not necessarily mean that this method comes closer to biological reality than the other two. Given the high correlation, it is simply possible to predict, with a smaller margin of error than for the other biological measurements, the value of the biological test on the basis of the chemical score calculated by means of the egg taken as reference. Figure 1 shows the regression line and the standard deviation obtained by correlating the values of the NPU and of the chemical score calculated from egg (1963). That part of the regression line is drawn which covers the values of the chemical scores falling within a distance of two standard deviations from the average score. The standard deviation of the regression line is represented by the two curves on either side of the regression line, which delimit a confidence interval of ± 1 standard deviation and includes the degree of variation around the regression line. As can be seen the standard deviation is small around the average value and increases as the further one deviates from it.
Instead of basing oneself on the regression line in order to determine the NPU from the chemical score, it appears desirable to utilize the NPU values given by the lower limit of the confidence interval (- 1 standard deviation). The estimate thus presents a safety margin of about 85 percent, for in probability terms there will now be a chance of only about 15 percent that the value found will be less than the NPU determined experimentally.
According to Miller and Payne (1951) (7) the standard NPU is obtained on diets having a protein calorie ration of 4 percent, that is, having a minimum level capable of meeting maintenance requirements. They showed that, beyond this value, protein utilization drops more or less linearly as the protein calorie/ratio increases. In other words, the organism's ability to use the proteins diminishes as the protein calorie ratio of the diet rises. They thus arrive at the concept of the operative NPU. The authors propose a formula and table (Miller and Payne 1961) (8) which, taking into account calories of protein origin, enable the operative NPU to be calculated from the standard NPU or from the chemical score calculated on the basis of the FAO pattern.
We have shown that the CS calculated from egg gives the highest correlation with the NPU. Since our regression calculations apply to NPU10, in order to render them "operative" we have established, from the formula of Miller and Payne, a new equation:
NPU op = NPU10 (1,225 - 2.250 P)
in which the NPU10 of a diet is obtained from the data in Figure 1 and P is the percentage of protein calories in the diet.
Finally, we may wonder, with Hegsted (1969) (9) if the results of the NPU established on young growing rats can be directly transposed to growing children and adult men. Rakowska and Coll (1967) (10), studying 46 diets, obtained different operative NPUs depending on whether they were established in children or in rats. In the case of the latter, significantly higher values (child NPU/rat NPU = 0.82) are obtained. It therefore seems that, contrary to current practice, it is necessary to apply here again a correction factor to establish the operative NPU of diets intended for human feeding.
FIGURE 1 - REGRESSION LINE AND CONFIDENCE INTERVALS (± SD) RESULTING FROM THE CORRELATION BETWEEN NPU10 AND CHEMICAL SCORE
Reference protein: 1963 egg
The practical use of the results of the study of correlation between biological and chemical measurements raises a number of problems, as shown by the discussion of the data on NPU.
First of all, it would be necessary for laboratories to carry out a set of biological determinations (BV, PER and NPU) on a sufficiently high number of foods and diets and for the amino acid composition to be determined on the same samples by column chromatography. Statistical treatment of the results of the type which we have used would then be performed. At the same time it would be desirable to accumulate data on the modification of protein utilization according to the various protein calorie ratios of the diets and to study how far the data obtained from animals can be extrapolated to man. This would be a vast project involving substantial, financial and technical resources. It would seem that some laboratories are already embarking on this course. (11) This cannot be encouraged too highly, for a revision of the report on protein requirements will only constitute a genuine improvement of our existing knowledge when more precise data are available on the relationships between the amino acid composition of foods, their protein quality and their ability to satisfy growth and maintenance requirements in man.
(1) Autret, M., Périssé, J., Sizaret, F., Cresta, M., Protein Value of Different Types of Diet in the World: Their Appropriate Supplementation. Nutrition Newsletter, FAO, Vol.6, No. 4, 1 - 29 (1968).
(2) FAO. Amino Acids Content of Foods and Biological Data on Proteins (1969, in press).
(3) Mitchell, HH., Block, E.J. Some Relationships between the Amino Acid Contents of Proteins and their Nutritive Value for the Rat. J. Biol. Chem. 163 (1946), 599 - 620.
1 In fact, from the first stages of their research, the authors adopted as a representative measurement of protein quality not a percentage but the difference between its value and 100. This does not in any way alter the order of classification but it does cause the coefficients of correlation to assume a negative value.
(4) Block, R.J., Mitchell, H.H. The Correlation of the Amino Acid Composition of Proteins with their Nutritive Value. Nutr. Abst. Rev. 16 (1946-47), 249 - 278.
(5) FAO. Protein Requirements. FAO Nutritional Studies No. 16 (1958).
(6) FAO. Protein Requirements. FAO Nutrition Meetings Report Series No. 37 (1965).
(7) D.S. Miller, P.R. Payne. Problems in the prediction of protein values of diets. The influence of protein concentration. Brit. J. Nutr., 1961, 15, 11-19.
(8) D.S. Miller, P.R. Payne. Problems in the prediction of protein values of diets: the use of food composition tables. J. Nutr., 1961, 74, 413-418.
(9) D.M. Hegsted. Nutritional value of cereal proteins in relation to human needs. From AACC - Protein-enriched cereal foods for world needs. Ed. Max Milner, 1969.
(10) M. Rakowska, W. Szkittadz, A. Szczygiel. Comparative determination of NPU in whole day diet, on children and on rats. Proc. VII Intern. Cong. Nutr. Hamburg, 1966.
(11) E. Kofranyi, F. Jekat. The minimum N-requirement of man. Proc. VII Intern. Congr. Nutr. Hamburg, 1966.
![]()
![]()
![]()