![]()
![]()
![]()
L. Hakkari and P. Selin
Hydrobiological Research Centre, University of Jyvaskyla
Jyvaskyla, Finland
K. Westman
Finnish Game and Fisheries Research Institute
Helsinki, Finland
and
M. Mielonen
City Office for Sports and Outdoor Recreation
Helsinki, Finland
ABSTRACT
In 1973, one-summer old whitefish fingerlings were stocked in two small forest lakes: Lake Valkea-Mustajarvi, an oligohumous lake with a surface area of 13.9 ha and a mean depth of 3.3 m; and Lake Majajarvi, a polyhumous lake with a surface area of 3.4 ha and a mean depth of 4.0 m. The stocking density was 35 fingerlings of C. muksun (Pallas) and 35 fingerlings of C. peled (Gmelin) per hectare in both lakes.
The zooplankton biomass (wet weight) was 40.4 kg/ha in Lake Valkea-Mustajarvi and 26.1 kg/ha in Lake Majajarvi in June–September 1974. The amount of the large, preferred plankton species, Heterocope appendiculata and Holopedium gibberum, was less in Lake Majajarvi than in Lake Valkea-Mustajarvi; Daphnia galeata and Leptodora kindti were lacking entirely in Lake Majajarvi. Respectively, small zooplankters, rotifers, Ceriodaphnia quadrangula and Bosmina longirostris were more abundant in Lake Majajarvi.
According to the stomach analyses, cladocerans (Daphnia, Bosmina) and Holopedium usually dominated in the summer and autumn, and copepods (especially Eudiaptomus sometimes Cyclops) in the winter and spring. Bottom fauna (e.g., chironomids, culicids, trichopterids) constituted more than 50 percent of the stomach content in only two of the 29 fish samples.
The similarity index was low (45±4.9 percent) in Lake Valkea-Mustajarvi but quite high (78.3±9.1 percent) in Lake Majajarvi. Effective predation in the latter lake diminished the occurrence of important food species, with the result that Bosmina coregoni and Daphnia cristata became objects of predation.
Thus, both similarity of food items and trophic competition were high in Lake Majajarvi.
The difference between the intensity of competition in the two lakes was observed in the stomach fullness and condition indices as well as in the growth rate. The growth rate of C. muksun was high in Lake Valkea-Mustajarvi: the total length of four-year old C. muksun individuals was 39.4±0.4 cm.
The zooplankton production utilizable by the next trophic level was estimated at circa 400 kg/ha in Lake Valkea-Mustajarvi but only 130 kg/ha in Lake Majajarvi in June–September 1974. Whitefish production in Lake Valkea-Mustajarvi was estimated as accounting for over 60 percent of the theoretical total fish production calculated on the basis of utilizable zooplankton. According to the zooplankton species composition, utilization of zooplankton was higher in Lake Majajarvi than in Lake Valkea-Mustajarvi in 1974. Whitefish numbers in Lake Majajarvi may have decreased during the winter of 1974–75 due to starvation and possibly a low oxygen concentration. Thus, a stocking rate of 70 whitefish fingerlings per hectare was suitable for Lake Valkea-Mustajarvi but too great for Lake Majajarvi.
RESUME
En septembre 1973, on a repeuplé deux petits lacs forestiers du sud de la Finlande (Valkea-Mustajarvi et Majajarvi) avec des juvéniles (un été) appartenant à deux espèces de poisson blanc: Coregonus muksun (espèce indigène planctophage) et Coregonus peled (poisson blanc sibérien, espèce introduite). On a mis 487 unités de chaque espèce dans le premier lac, 119 dans le second. Les deux lacs sont oligotrophes; le premier est oligohumique, le second polyhumique. Leur superificie est respectivement de 13,9 et 3,4 ha et leur profondeur moyenne de 4,0 et 3,3 m.
On a analysé le contenu de l'estomac de ces poissons et on a constaté que leur nourriture se composait essentiellement de cladocères (Daphnia, Bosmina et Holopedium) en été et en automne et de copépodes surtout Eudiaptomus spp. et, occasionnellement, Cyclops spp.) en hiver et au printemps. Seuls deux des 31 estomacs analysés contenaient plus de 50 pour cent (en poids) de faune benthique (par exemple, chironomides, culicides, trichoptérides, hydracariens).
L'indice de similarité alimentaire (cf. Shorygin, 1939) entre les deux espèces de poisson blanc était plus élevé dans le lac de Majajarvi (72 pour cent) que dans celui de Valkea-Mustajarvi (48 pour cent) en 1974. Le pourcentage de faune benthique dans le premier de ces lacs était faible. La biomasse du zooplancton en juin–septembre 1974 (poids humide) était de 26 kg/ha dans le lac de Majajarvi et de 40 kg/ha dans celui de Valkea-Mustajarvi. La production de zooplancton utilisable au stade trophique suivant était estimée à 400 kg/ha environ (en juin/septembre) dans le lac de Valkea-Mustajarvi et à 130 kg/ha seulement dans le lac de Majajarvi. En 1974, le pourcentage de remplissage des estomacs (cf. Windell, 1971) montrait que les poissons étaient insuffisamment nourris. Dans le lac de Valkea-Mustajarvi, ce pourcentage était de 47,5/1 000 pour Coregonus muksun et de 34, 2/1 000 pour Coregonus peled; dans le lac de Majajarvi, il était respectivement de 25,4/1 000 et 30,1/1 000. En outre, le taux de croissance du poisson blanc montrait clairement qu'il y avait une forte concurrence alimentaire dans le lac de Majajarvi. En octobre 1974, le poids moyen de C. muksun dans le lac de Valkea-Mustajarvi était de 108 g, contre 33 g seulement dans le lac de Majajarvi; pour C. peled, il était respectivement de 183 et 66 g.
In recent years more attention has been paid in efforts to improve the economic utilization of Finland's numerous small lakes by stocking of whitefish which feed on zooplankton. Such whitefish are generally not found in the natural communities of small Finnish lakes. In 1973 the Evo Inland Fisheries and Aquaculture Station began a comparative study of the suitability of the domestic Coregonus muksun (Pallas) and Coregonus peled (Gmelin), imported from the U.S.S.R. in 1965 for fish cultivation in small waters (Pruuki, Pursiainen and Westman, 1983).
The aim of the present investigation was to study the food and trophic competition between these two whitefish species stocked in 1973 in two small forest lakes in southern Finland, both the property of the Evo Inland Fisheries and Aquaculture Research Station.
Lakes Valkea-Mustajarvi and Majajarvi are situated in Lammi, Evo (25°5'E; 61°12'N). The water in these lakes is quite different (Table 1). Lake Valkea-Mustajarvi is oligohumic and Lake Majajarvi polyhumic. Oxygen loss was observed in March–April in the hypolimnion of both lakes, and in some years also in the epilimnion of Lake Majajarvi.
The growth and production of the whitefish stocked in Lake Valkea-Mustajarvi has been presented elsewhere (Pruuki, Pursiainen and Westman, 1983).
Zooplankton samples were collected with a Sormunen tube sampler and filtered through a 50 um mesh (Hakkari, 1978). Altogether, 64 zooplankton samples were collected between October 1973 and October 1974. The zooplankton biomass was estimated as wet weight using standard volumes (Hakkari, 1978). The production of the most important zooplankton taxa was estimated according to the graphical method of Winberg (1971).
In October 1973, one-summer old whitefish fingerlings were stocked in the lakes, 35 C. muksun and 35 C. peled per hectare. The fish material collected with gill nets and fyke nets in 1974–76 consisted of 300 individuals in 29 samples (of. Table 3). Species, total length (RT-measure), weight and the weight and species composition of the stomach contents were analysed. Samples were preserved frozen. The zooplankton and fish samples were taken by the author Matti Mielonen and the analyses were carried out by the author Pirkko Selin.
Eudiaptomus spp. (E. gracilis and graciloides), Daphnia cristata and Holopedium gibberum dominated the zooplankton in Lake Valkea-Mustajarvi; and the mean total biomass in this lake was 40.4 kg/ha during the growing season 1974 (Table 2). The mean biomass in Lake Majajarvi was only 65 percent, i.e., 26.1 kg/ha that of Lake Valkea-Mustajarvi. The dominant zooplankton taxa in Lake Majajarvi were Eudiaptomus spp., Cyclops spp., and Daphnia cristata; D. galeata and Leptodora kindti were entirely lacking. On the other hand, the amounts of rotifers, Bosmina longirostris and Ceriodaphnia quadrangula were higher than in Lake Valkea-Mustajarvi.
If the ingestion rate of predatory zooplankton is taken as four times their own production (Winberg et al., 1972) then the useful zooplankton production left for fish and invertebrate predators was at least 400 kg/ha in Lake Valkea-Mustajarvi, and 130 kg/ha in Lake Majajarvi in June–September 1974.
Zooplankton was the main food of both whitefish species (Table 3). Only in two of the 29 fish samples did bottom and surface fauna (e.g., chironomids, culicids, trichopterids, hydracarins) constitute more than 50 percent of the stomach contents. Chironomids and culicids were partly as pupae in the spring. According to Grimas (1961) they are important food for fish at that time.
The stomach contents were clearly simpler in Lake Majajarvi. Coregonus peled and C. muksun fed on almost the same food items. The size of food items changed only slightly in the food contents of one and four-year old whitefish. Asellus aquaticus was only observed in the diet of C. muksun with a length of more than 34 cm.
The similarity index that may have values between 0 and 100 percent (Shorygin, 1939) was low between C. muksun and C. peled (average 45 ± 4.9 percent) in Lake Valkea-Mustajarvi but quite high (average 78.3 ± 9.1 percent) in Lake Majajarvi in 1974. The differences between the lake indices were statistically significant (t = 3.24 t5 percent = 2.31; Fig. 1). In 1975 the similarity between the species in Lake Valkea-Mustajarvi seemed to be quite high (cf. Table 3).
The forage ratio (FR) (Zaret, 1980) showed that both species preferred Holopedium in Lake Valkea-Mustajarvi in the summer. In both lakes, C. muksun seemed to prefer Daphnia slightly more intensively than C. peled. Bosmina c. obtusirostris was the only strongly preferred taxon in Lake Majajarvi; however, in Lake Valkea-Mustajarvi it was only slightly preferred in the spring and autumn. The most abundant copepod taxa, Eudiaptomus and Cyclops, were avoided during the summer despite their rather high biomass (Figs. 2 and 3).
The stomach fullness index (Windell, 1971) gives information as to the sufficiency of food. The mean stomach content indices were as follows:
| Lake Valkea-Mustajarvi | Lake Majajarvi | ||
| 1974 | 1975 | 1974 | |
| C. muksun | 47.5 %oo | 32.1 %oo | 25.4 %oo |
| C. peled | 34.2 %oo | 18.8 %oo | 30.1 %oo |
The index values for C. muksun were statistically higher in Lake Valkea-Mustajarvi than in Lake Majajarvi in summer in 1974 (t = 2.32x).
The Fulton condition index of C. muksun was significantly higher in Lake Valkea-Mustajarvi than in Lake Majajarvi and slightly significantly higher by C. peled in 1974:
| Lake Valkea-Mustajarvi | Lake Majajarvi | t-test | |
| C. muksun | 0.768±0.017 | 0.678±0.013 | 4.117xx |
| C. peled | 0.838±0.027 | 0.748±0.012 | 3.091x |
The results agree well with the stomach content index values. C. muksun and C. peled were highly significantly bigger in Lake Valkea-Mustajarvi than in Lake Majajarvi in the autumn 1974, when whitefish were two years old (C. muksun: t = 16.41xxx; C. peled: t = 15.13xxx). Respectively, C. peled was highly significantly bigger than C. muksun in Lake Valkea-Mustajarvi (t = 8.07xxx; cf. Pruuki, Pursiainen and Westman, 1983) and in Lake Majajarvi (t = 7.81xxx) at that time (Table 4).
Sympatric whitefish species have an ability to segregate into different feeding niches, especially as the gill raker counts are different (Nilsson, 1960; Lindstrom and Nilsson, 1962). In this study the gill raker counts were close to each other, roughly 50. Feeding segregation appeared to take place in Lake Valkea-Mustajarvi where a varied food supply made it easy. The segregation was observed as a low similarity (percentage of overlap in feeding habits) between the species in 1974.
For example, Nilsson (1960, 1965) stated that segregation between whitefish species took place when competition was high. Also, segregation became clearer toward autumn when the food was on the decrease. This was also found by Heikinheimo-Schmid (1982) in some lakes in northern Finland.
In this study no segregation was found in Lake Majajarvi during hard competition for food between C. muksun and C. peled in 1974. In spite of the sparse material, the standard error of the mean of similarity between the food composition of C. muksun and C. peled was small, as seen in Fig. 1. Obviously, the result gave a real picture of the situation in the lake.
Large species, such as Leptodora and Heterocope appendiculata, as well as Holopedium gibberum, were preferred in Lake Valkea-Mustajarvi. Thus the size and visibility of the prey species had an effect on food selection, as pointed out earlier by many authors (e.g., Brooks and Dodson, 1965; Hrbacek and Hrbackova-Esslova, 1966; Reif and Tappa, 1966; Hakkari 1978). Eudiaptomus and Cyclops were avoided in summer, perhaps because of their agility (cf. Dodson, 1974).
The size of fish had no clear effect on the food composition. C. muksun, 30–40 cm in length, fed mainly on small items, e.g., Daphnia, Bosmina and Cyclops (cf. Table 3). Heikinheimo-Schmid (1982) supposed that the size of densely-rakered whitefish does not have as great an effect on the food composition as that of whitefish with a low gill raker count.
Many Scandinavian investigations have shown that among the zooplankters consumed by most species of Coregonus, Bosmina coregoni is by far the most important food item (Nilsson and Pejler, 1973). Its presence in the stomach content indicates a high feeding pressure on zooplankton. The number of large, preferred species is low, and selection becomes intensively concentrated on Bosmina coregoni, as was observed in Lake Majajarvi in 1974.
Fish predation has an effect on the species composition of zooplankton (e.g., Zaret, 1980). The differences in the predation pressure between L. Valkea-Mustajarvi and Majajarvi are seen in the proportions of preferred and avoided species. Rotifers are too small to be food for whitefish. They compete for food with cladocerans and benefit by the predation on cladocerans. This has been shown, e.g., in fish ponds with a natural food supply, where fingerlings of C. peled were raised (Hakkari and Dahlstrom, 1969). The increase of the small cladocerans Bosmina longirostris and Ceriodaphnia quadrangula in Lake Majajarvi agrees well with the results of Nilsson and Pejler (1973) in Swedish whitefish lakes. Respectively, the proportion of the large species Holopedium, Daphnia galeata, Leptodora and Heterocope appendiculata was smaller in Lake Majajarvi, which also indicates more effective predation in Lake Majajarvi than in Lake Valkea-Mustajarvi.
The utilizable zooplankton production for fish and predatory invertebrates was more than 400 kg/ha/year in Lake Valkea-Mustajarvi and 130 kg/ha/year in Lake Majajarvi. The abundance of invertebrate predators, e.g., Chaoborus, was low in both plankton and stomachs; and other predators, e.g., Gammarus, were not observed even in the stomach content of whitefish.
The productivity of the natural fish composition of Lake Valke-Mustajarvi has obviously also been higher than that of Lake Majajarvi, because of the greater transparency of the water. Thus, when 70 one-summer old whitefish fingerlings per hectare were introduced, the zooplankton productivity in Lake Valkea-Mustajarvi was able to bear the increased predation but zooplankton production in Lake Majajarvi could not meet the demand for food. Therefore, the final results were:
The whitefish in Lake Valkea-Mustajarvi were in good condition and had a good rate of growth while those in Lake Majajarvi starved and had a lower growth rate; and
In Lake Majajarvi both the biomass and production of preferred large zooplankton were low because of the effective predation.
The potential fish production based on the zooplankton production in 1974, calculated using a food coefficient of 8 (Phillips, 1972; Winberg et al., 1972) was about 50 kg/ha/year in Lake Valkea-Mustajarvi but only 16 kg/ha/year in Lake Majajarvi. Veijo Pruuki (manuscript) estimated that the maximum biomass of a stocking year-class was 200–300 kg/ha/1 000 whitefish fingerlings in Lake Valkea-Mustajarvi. According to Pruuki, it imparts a net production of 20–30 kg/ha, if stocking density is 100 fingerlings per hectare.
If zooplankton production is assumed to be lowered from the 1974 level because of the increased fish production resulting from the whitefish introduction (cf. Nilsson and Pejler, 1973), fish production based on zooplankton may rise to more than 60 percent of theoretical fish production. According to the zooplankton species composition, utilization of zooplankton was higher in Lake Majajarvi than in Lake Valkea-Mustajarvi in 1974.
Although several trial fishings were made, the catch was only four C. peled in Lake Majajarvi in 1975. The amount of whitefish diminished during the winter of 1974–75 as a result of starvation and, perhaps, because of the low oxygen concentration.
Brooks, J.L. and S.I. Dodson, 1965 Predation, body size and composition of plankton. Science Wash., 150:28–35
Dodson, S.I., 1974 Adaptive change in plankton morphology in response size-selective predation: a new hypothesis of cyclomorphosis. Limnol.Oceanogr., 19:721–9
Grimas, U., 1961 The bottom fauna of natural and impounded lakes in northern Sweden. Rep.Inst.Freshwat.Res.Drottningholm, (46):95–123
Hakkari, L., 1978 On the productivity and ecology of zooplankton and its role as food for fish in some lakes in Central Finland. Biol.Res.Rep.Univ.Hyvaskyla, 4:3–87
Hakkari, L. and H. Dahlstrom, 1969 Zooplankton in studies of inland waters. Limnologisymposiun, 1968:61–8 (in Finnish, English summary)
Heikinheimo-Schmid, O., 1982 Siian ravinnosta luonnontilaisessa ja saannostelyssa jarvessa. Riista-ja kalatalouden tutkimuslaitos. Kalantutkimusosasto. Monistettuja julkaisuja, 4:1–64
Hrbacek, J. and M. Hrbackova-Esslova, 1966 The taxonomy of the genus Daphnia and the problem of “biological indication”. Verh.Int.Ver.Theor.Angew.Limnol., 16:1661–7
Lindstrom, T. and N.-A. Nilsson, 1962 On the competition between whitefish species. In The exploitation of natural animal populations, edited by E.D. Le Cren and M.W. Holdgate. Oxford, Blackwell Scientific Publications, pp. 326–40
Nilsson, N.-A., 1960 Seasonal fluctuations in the food segregation of trout, char and whitefish in fourteen north Swedish lakes. Rep.Inst.Freshwat.Res.,Drottningholm, (41):185–205
Nilsson, N.-A., 1965 Food segregation between salmonoid species in north Sweden. Rep.Inst.Freshwat.Res., Drottningholm, (46):95–123
Nilsson, N.-A. and B. Pejler, 1973 On the relation between fish fauna and zooplankton composition in north Swedish lakes. Rep.Inst.Freshwat.Res., Drottningholm, (53):51–77
Phillips, A.M., 1972 Calorie and energy requirement. In Fish nutrition, edited by J.E. Halver. New York, Academic Press, pp. 1–28
Pruuki, V., M. Pursiainen and K. Westman, 1984 A comparative study of the growth and production of the native whitefish (Coregonus muksun) and the introduced whitefish (C. peled) stocked in two small forest lakes in southern Finland. EIFAC Tech.Pap., (42) Vol. 1:91–108
Reif, C. and D.W. Tappa, 1966 Selective predation: Smelt and cladocerans in Harveys Lake. Limnol.Oceanogr., 11:437–8
Shorygin, A.A., 1939 Pitanie, izbiratelnya sposobnost i pishchevye vzaimootnosheniya neketorykh Gobiidae Kaspiiskogo morya. Zool.Zh., 18(1):27–53
Winberg, G.G. (ed.), 1971 Methods for the estimation of production of aquatic animals. London, Academic Press, 175 p.
Winberg, G.G. et al., 1972 Biological productivity of different types of lakes. In Productivity problems of freshwaters, edited by Z. Kajak and A. Hillbricht-Ilkowska. Warsaw, PWN - Polish Scientific Publishers, pp. 383–404
Windell, J.T., 1971 Food analysis and rate of digestion. IBP Handb., (3):215–26
Zaret, T.M., 1980 Predation and freshwater communities. New Haven, Yale University Press, 187 p.
Table 1 Hydrographical data from Lakes Valkea-Mustajarvi and Majajarvi from 1976 to 1981
| L. Valkea-Mustajarvi | L. Majajarvi | |||
| Depth (m) | 1 | 9 | 1 | 9 |
| Oxygen concentration in March–April (mg/l) | 8.1–12.3 | 0.9–1.8 | 3.7–9.5 | 0.0–1.3 |
| pH | 5.8–7.2 | 5.8–6.5 | 5.6–5.7 | 5.6–6.3 |
| Water colour (mg Pt/l) | 10–30 | 25–50 | 140–160 | 140–300 |
| Conductivity (uS) | 18–24 | 18–30 | 23–26 | 25–30 |
| Area (ha) | 13.9 | 3.4 | ||
| Greatest depth (m) | 10.0 | 12.0 | ||
| Mean depth (m) | 3.3 | 4.0 | ||
Table 2 The mean biomass (B) and production (P) of the most important zooplankton taxa in Lakes Valkea-Mustajarvi and Majajarvi in June–September 1974
| L. Valkea-Mustajarvi | L. Majajarvi | |||
| B(kg/ha) | P(kg/ha) | B(kg/ha) | P(kg/ha) | |
| Rotifera | 2.31 | 30.98 | 3.04 | 45.72 |
| Limnosida frontosa | 0.01 | 0.96 | - | - |
| Diaphanosoma brachyurum | 0.57 | 28.50 | 0.30 | 19.92 |
| Holopedium gibberum | 7.30 | 200.00 | 0.89 | 26.16 |
| Daphnia cristata | 8.30 | 156.00 | 3.29 | 63.60 |
| D. galeata | 1.10 | 25.20 | - | - |
| Bosmina coregoni obtusirostris | 4.26 | 60.40 | 2.80 | 49.20 |
| B. longirostris | + | + | 0.30 | 3.64 |
| Leptodors kindti | 0.18 | 3.41 | - | - |
| Ceriodaphnia quadrangula | 0.01 | 0.25 | 0.22 | 4.09 |
| Heterocope appendiculata | 2.71 | 9.10 | 0.84 | 3.20 |
| Eudiaptomus spp. | 8.75 | 71.20 | 9.50 | 62.40 |
| Cyclops spp. | 4.87 | 55.40 | 4.88 | 32.20 |
| Total | 40.37 | 641.40 | 26.06 | 310.10 |
Table 3 The numbers and total lengths (cm) of the whitefish species in different seasons and the dominant food items in the stomachs (% of biomass)
| Lake Valkea-Mustajarvi | Lake Majajarvi | ||||||||
| C. muksun | C.muksun | ||||||||
| 1974 | Winter (I-III) | Spring (IV-V) | Summer (VI-IX) | Autumn (X) | 1974 | Winter (I-III) | Spring (IV-V) | Summer (VI-IX) | Autumn (X) |
| No. | 8 | 14 | 25 | 21 | No. | 8 | 3 | 12 | 9 |
| Size(cm) | 11.3–12.6 | 11.7–15.0 | 14.4–24.6 | 20.7–25.6 | Size(cm) | 10.5–14.7 | 11.6–13.3 | 11.7–18.7 | 15.5–19.5 |
| Main food | Daphnia(49%) Chironomidae (28%) | Culicidae (71%) Chironomidae (11%) | Holopedium (39%) Daphnia(44%) Leptodora (12%) | Daphnia(44%) Holopedium (20%) Bosmina(19%) | Main food | Eudiaptomus (59%) Cyclops(41%) | Cyclops(68%) Eudiaptomus (21%) | Bosmina(68%) Daphnia(27%) | Bosmina(39%) Daphnia(35%) |
| 1975 | Winter (III) | Spring (V) | Summer (VII) | Autumn (XII) | |||||
| No. | 11 | 10 | 6 | 14 | |||||
| Size(cm) | 22.8–26.7 | 24.3–26.3 | 28.2–31.3 | 31.6–34.2 | |||||
| Main food | Eudiaptomus (91%) | Daphnia(46%) Chironomidae (34%) | Daphnia(54%) Cyclops(28%) | Daphnia(61%) Asellus(16%) Cyclops(13%) | |||||
| 1976 | Winter (I-II) | Spring (V) | Summer (VI-IX) | Autumn (XII) | |||||
| No. | 9 | 4 | 14 | ||||||
| Size(cm) | 31.8–39.6 | 30.8–36.2 | 38.0–41.5 | ||||||
| Main food | Daphnia(46%) Cyclops(32%) | Daphnia(46%) Cyclops(16%) Eudiaptomus (13%) | Bosmina(71%) Asellus(14%) | ||||||
| C.peled | C.peled | ||||||||
| 1974 | Winter (I-III) | Spring (IV-V) | Summer (VI-IX) | Autumn (X) | 1974 | Winter (I-III) | Spring (IV-V) | Summer (VI-IX) | Autumn (X) |
| No. | 0 | 10 | 52 | 8 | No. | 0 | 2 | 31 | 12 |
| Size(cm) | 12.2–14.9 | 14.9–28.1 | 27.0–36.0 | Size | 11.6–12.2 | 12.4–22.0 | 17.6–22.7 | ||
| Main food | Cyclops(48%) Eudiaptomus (27%) Culicidae (16%) | Daphnia(20%) Cyclops(16%) Holopedium (13%) | Holopedium (27%) Heterocope (22%) Cyclops(14%) | Main food | Cyclops(74%) Eudiaptomus (24%) | Bosmina(80%) Daphnia(12%) | Bosmina(47%) Daphnia(44%) | ||
| 1975 | Winter (III) | Spring (V) | Summer (VII) | Autumn (XII) | 1975 | Winter (III) | Spring (V) | Summer (VII) | Autumn (XII) |
| No. | 3 | 1 | 11 | No. | 0 | 0 | 0 | 2 | |
| Size(cm) | 28.0–28.5 | 29.6 | 30.5–36.0 | Size | 27.6–30.2 | ||||
| Main food | Eudiaptomus (75%) Cyclops(21%) | Daphnia(85%) | Daphnia(54%) Cyclops(25%) | ||||||
Table 4 The mean total lengths (cm) of C. muksun and C. peled in Lakes Valkea-Mustajarvi and Majajarvi in the autumn 1973–76
| C. muksun | C. peled | |
| 1973 | ||
| Valkea-Mustajarvi | 10.0 | 9.4 |
| Majajarvi | 10.0 | 9.4 |
| 1974 | ||
| Valkea-Mustajarvi | 24.1±1.1 | 28.1±0.3 |
| Majajarvi | 17.1±0.4 | 20.6±0.4 |
| 1975 | ||
| Valkea-Mustajarvi | 34.3±0.6 | - |
| Majajarvi | - | 28.6±1.6 |
| 1976 | ||
| Valkea-Mustajarvi | 39.4±0.4 | - |
| Majajarvi | - | - |
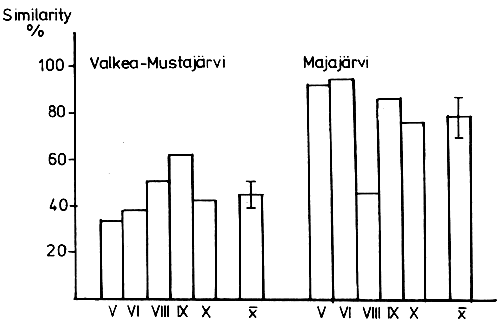
Fig. 1 The similarity index (Shorygin, 1939) between C. muksun and C. peled in L. Valkea-Mustajärvi and L. Majajärvi
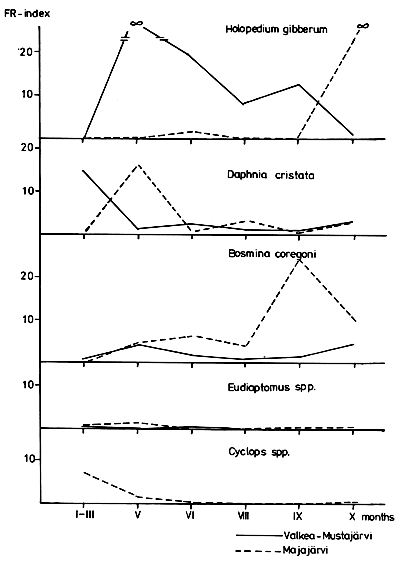
Fig. 2 The forage ratio (FR) index (Zaret, 1980) for C. muksun in L. Valkea-Mustajärvi and L. Majajärvi
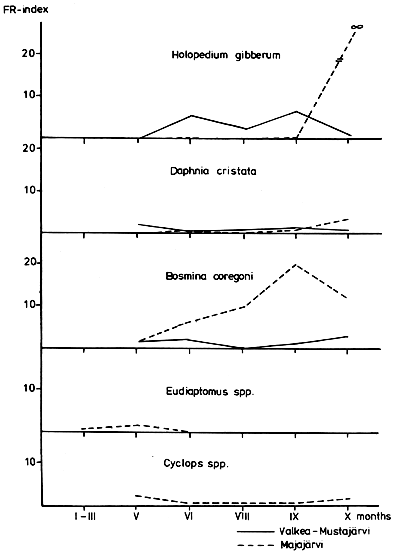
Fig. 3 The forage ratio (FR) index (Zaret, 1980) for C. peled in L. Valkea-Mustajärvi and L. Majajärvi
![]()
![]()
![]()