![]()
![]()
G.F.FRISONI, CEMAGREF, Section Aménagements littoraux et Aquaculture,
O. GUELORGET, Chercheur CNRS, Laboratoire d'Hydrobiologie Marine, U.S.T.L.
INTRODUCTION
Les milieux lagunaires littoraux apparaissent depuis la plus haute antiquité comme des sites d'une grande importance économique. Cette réputation s'appuie sur l'exploitation des ressources aussi bien minérales que biologiques.
Déjà les Phéniciens utilisaient certains sites lagunaires pour la fabrication du sel (Salines d'Ibiza -Baléares-…) marchandise de grande valeur commerciale dans l'antiquité.
On se souvient également, de la notoriété de l'étang de Diana en Corse ; port principal de cette dépendance romaine, il était en outre un centre réputé de production d'huîtres comme en témoigne l'amoncellement de coquilles formant aujourd'hui une île dans la lagune actuelle. L'exploitation des mollusques lagunaires alimentaient aussi une industrie florissante dans l'antiquité, celle de la pourpre issue du murex. La plus belle trace de cette industrie apparaît sur la rive sud de la Bahiret el Biban (Tunisie) sous la forme d'une colline uniquement constituée de tests vides de ce gastéropode.
Il faut souligner enfin la place essentielle du domaine lagunaire dans la pêche artisanale côtière ; pratiquée depuis es temps les plus anciens, cette activité a permis l'émergence de techniques adaptées et spécifiques aux lagunes, comme en témoigne les pêcheries fixes (bordigues) intallées sur le pourtour de la Méditerranée.
L'importance économique des lagunes côtières n'a fait que s'accroître au cours du temps. La diversification des activités, la mise en exploitation de sites de plus en plus nombreux, l'intérêt commercial pour de nouvelles espèces, l'amélioration des techniques assurant notamment le passage de la cueillette à l'élevage font de ces milieux des gisements encore riches de ressources exploitables à l'heure actuelle.
Les bassins évaporitiques fossiles ou actuels, fournissent en abondance des minéraux et des éléments indispensables aux activités humaines (Halite ; Gypse ; sels potassiques ; éléments extraits des saumures - Magnésium, Brome, Potassium…). En 1971 la production de sel (Halite), provenant en quasi totalité du domaine paralique1, atteignait 143 millions de tonnes. A titre d'exemple la production du Salin de Giraud (Bouches-du-Rhône) dont la surface évaporante est de 10500 ha, avoisine le million de tonnes de halite commercialisable par an. (Le prix moyen du sel alimentaire ou industrie) est compris entre 10 et 15 dollars la tonne).
Selon NIXON (1981) la production halieutique des milieux paraliques, à l'échelle mondiale est extrêmement variable et se situe entre quelques kilogrammes et une tonne par hectare et par an. En Méditerranée, selon AMANIEU et LASSERRE (1981) les pêcheries lagunaires contribuent pour 8 à 10% au moins de l'ensemble des captures (Mer Noire exclue). Ces auteurs soulignent “l'extrême disparité des rendements qui s' étagent entre 6 kg/ha/an (lagune de Nador au Maroc) et 149 kg/ha/an (lagune de Venise en Italie), voire 172 kg/ha/an (estuaire de l'Ebre en Espagne)”. Dans ces conditions, il apparaît difficile de proposer un rendement moyen en domaine paralique. On peut toutefois se convaincre de l'importance économique des pêches lagunaires en constatant avec AMANIEU (1973) que pour l'année 1971, la seule production de la pêche aux petits métiers dans le étangs de Thau et de Berre était supérieure en valeurs (33000 KF) à la production des chalutiers (30000 KF) du littoral français.
La production conchylicole (élevage de mollusques filtreurs-huitres et moules-) annuelle des étangs méditerranéens oscille aux alentours de 15 T par hectare. A titre d'exemple les étangs du Languedoc-Roussillon ont produit en 1980 environ 9000 T de moules et 5000 T d'huitres représentant respectivement 15% et 5% de la production nationale. En comparaison et d'après les statistiques FAO/UNDP (1975, 1976), les productions mondiales d'huitres et de moules atteignent respectivement 591 386 T et 328 517 T. Cette activité assure le maintien dans la région Languedoc-Roussillon de 2000 emplois directs et 6000 emplois indirects. Dans le années 70, elle représentait pour le seul étang de Thau un revenu de 15 000 à 20 000 KF selon les estimations (AMANIEU 1973, DELBOS et al., 1973).
Outre la conchyliculture et la pêche lagunaire, le domaine paralique accueille des élevages de poissons et de crustacés. Si certains types d'élevages s'appuient sur des techniques déjà maitrisées (valliculture en Italie) d'autres élevages relèvent encore à l'heure actuelle de l'expérimentation et de la recherche. Exceptée l'Italie (5850 T en 1978). La production des élevages de poissons est encore limitée sur le pourtour méditerranéen (12 372 T en 1978).
A titre de comparaison l'aquaculture japonaise a produit en 1978, 133 448 tonnes de poisson (QUERELLOU, 1980).
Compte tenu de l'extention et de la mise au point de ces élevages, la production méditerranéenne attendue pour 1990 (Anonyme, 1979 in QUERELLOU, 1980) est de 61 000 tonnes de poissons et 3350 tonnes de crevettes.
Ormis les productions d sels la fonction essentielle du domaine paralique est donc d'ordre biologique, notamment en ce qui concerne les fortes productions végétales et animales générées. Cependant, il a un rôle écophysiologique important pour de nombreuses espèces des domaines marin et continental limitrophes. Ainsi le domaine paralique est un lieu privilégié, voire indispensable du cycle biologique de nombreuses espèces (migrations génésiques et trophiques, nurseries, maturation…). De ce fait il contribue largement à l'enrichissement des écosystèmes adjacents. AMANIEU et LASSERRE (1981) ont montré par exemple, qu'il existe une relation directe entre la production halieutique démersale et la production lagunaire le long des côtes de Méditerranée.
Toutefois, ces caractéristiques communes à l'ensemble des milieux paraliques ne doivent par masquer la grande variété morphologique et la réelle diversité géochimique de ce domaine.
La variété morphologique est à l'image de la nomenclature mouvante et inhomogène qui s'applique aux différents bassins paraliques: étangs, lagunes, marais, rias, abers, estuaires, lagons, mangroves, sebkhas, salins, marais salants, bahiras…
Ces termes décrivent des zones d'origine géologique fort différentes (sédimentaire, fluviatile, éolienne, tectonique…) donc aux formes, à la bathymétrie, et aux faciès sédimentaires, très variés.
La diversité des situations morphologiques, hydrologiques et climatiques conduisent à une diversité (et une variabilité) extrême des gammes de salinité et des gradients géochimiques des milieux paraliques. La caractéristique commune à ces champs de salinité est toutefois leur organisation longitudinale depuis la communication avec la mer vers les marges continentales.
Cette grande variabilité géochimique a conduit de nombreux auteurs à proposer des classifications basées sur la salinité (PETIT, 1953 ; d'ANCONA, 1954 ; AGUESSE, 1957 ; PETIT et SCHACHTER, 1959, SEGESTRALE, 1959 ; MARS, 1966 ; AMANIEU, 1967 ; MARAZANOF, 1972).
A cet égard, la salinité a généralement été considérée comme le paramètre écologique fondamental de la distribution des peuplements dans les milieux paraliques.
Cependant GUELORGET et PERTHUISOT (1982) ont montré que contrairement aux idées communément admises, la salinité n'est pas le facteur écologique directeur des milieux paraliques. Cette découverte résulte de l'observation d'une vingtaine de bassins paraliques méditerranéens et tropicaux par l'équipe pluridisciplinaire du GREDOPAR (Groupe d'Etude du Domaine Paralique) qui a abordé l'étude simultanée des peuplements phytoplanctoniques (G.F.FRISONI) et benthiques (O. GUELORGET) et de la sédimentologie (J.P. PERTHUISOT) (FRISONI, 1984; GUELORGET et PERTHUISOT, 1983; GUELORGET, 1985.).
Il en ressort que la localisation des espèces et les gradients quantitatifs (biomasse, densité…) des peuplements “lagunaires” (sensuslato) dépendent essentiellement d'un paramètre original que l'on peut exprimer, en chaque point considéré, comme le temps de renouvellement des éléments d'origine marine et qui est appelé le “confinement”. Ce terme, utilisé de manière empirique et intuitive dans diverses disciplines des Sciences de la nature est ambigû : en l'employant nous l'entendons “par rapport à la mer”. En un point donné d'un écosystème paralique, le “confinement” correspondrait alors à la déficience ou raréfaction en ce point des éléments vitaux “d'origine marine” (PERTHUISOT et GUELORGET, 1982.).
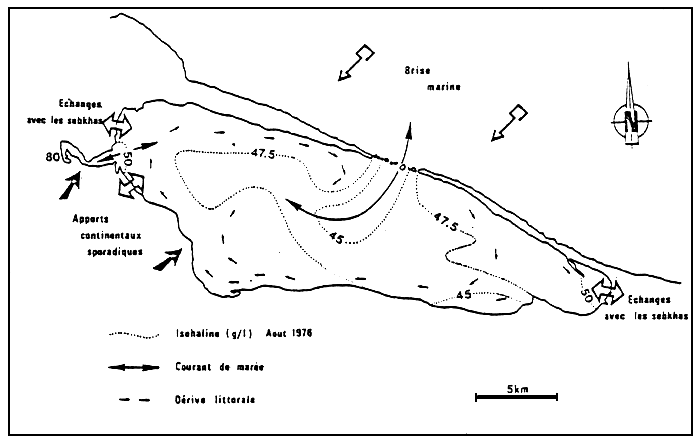
Ainsi par exemple, la petite monocotylédone, Ruppia maritima, est abondante dans les polders hollandais où DEN HARTOG (1971) lui assigne une “tolérance de salinité” comprise entre 1,5 et 23 %. On trouve cependant cette phanérogame dans l'étang d'Urbino (Corse) à 35 % à la Bahiret el Biban (Tunisie) à des salinités voisines de 70 % , (GUELORGET et al., 1982), ou encore dans le salin de Giraud (Camargue) à 80% (PERTHUISOT, 1983).
Par ailleurs, Pirenella conica, présente à la Bahiret el Biban à 70 %, se trouve aussi dans les lagunes de la baie d'Amvrakia (Grèce) à des salinités très faibles (5–10%) ou encore à des salinités voisines de celle de la mer dans des lagunes du golfe de Suez (GUELORGET, 1985).
Les tapis algaires (ou cyanobactériens) eux mêmes, sont capables de coloniser les milieux paraliques aussi bien sous-salés que sursalés.
On pourrait multiplier les exemples en s'adressant à des populations autres que benthiques. Inversement, des espèces à juste titre considérées comme marines, peuvent vivre et prospérer dans des zones nettement sursalées : madréporaires du golfe de Salwa (BASSON et al. 1977), ou sous-salées : oursins de la baie de Vonitsa (Grèce) (GUELORGET, 1985.). Mais ceci ne semble pouvoir se produire que lorsque le milieu est largement ouvert sur le domaine marine et constamment renouvelé.
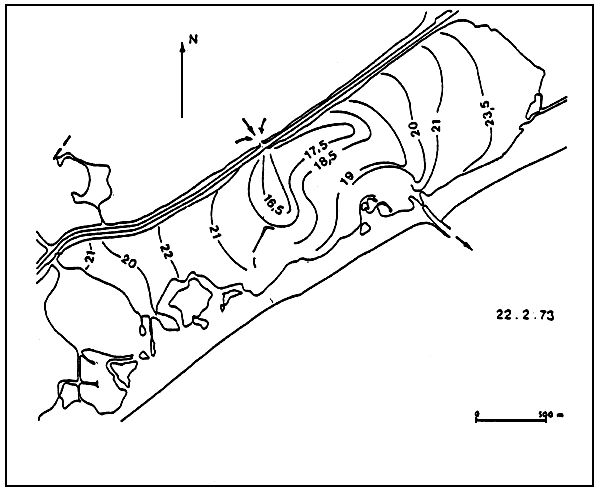
A l'image de ces espèces, les peuplements végétaux t animaux s'organisent indépendemment de la salinité, suivant un gradient de confinement. Ainsi l'organisation biologique des peuplements est semblable que l'on s'adresse aux milieux sursalés ou sousalés comme le montre l'étude comparée de la Bahiret el Biban (Tunisie) et l'étang du Prévost (Languedoc) (Fig 1 et 2).
Sur la base de ces constatations nous définirons tout d'abord l'organisation zonale de l'écosystème paralique, celle-ci induit l'existence d'une zonation “auquacole”, superposée à la zonation biologique naturelle).
I - LA ZONATION BIOLOGIQUE
La zonation biologique s'organise schématiquement selon la distance à la mer ouverte, il en va de même pour les gradients géochimiques (quel que soit leur signe). Dans le détail, on remarque que plus l'échange d'eau avec la mer est faible, plus les peuplements deviennent “typiquement paraliques” c'est-à-dire composés d'espèces eurybiontes, peu nombreuses, à fore densité. A l'inverse, plus on se rapproche des zones où les mouvements d'eau permettent un bon renouvellement du milieu, plus les peuplements sont riches en espèces, plus la densité diminue. (GUELORGET et al., 1981, 1982 a et b ; PERTHUISOT, 1983).
Le terme de “confinement” choisi pour exprimer cet éloignement à la mer recouvre encore des notions complexes et abstraites et constitue une grandeur non mesurable en l'état actuel de nos connaissances. A défaut d'une unité, on peut tenter d'établir une échelle qualitative. Celle-ci ne peut s'appuyer ni sur la géochimie, ni sur la sédimentologie. Il est donc nécessaire d'utiliser les points singuliers de la zonation biologique et à cet égard, el benthos est commode : outre que sa récolte est en général relativement aisée, ses espèces autochtones et son observation directe assez facile, il intègre les variations mineures et à court terme du milieu, et reflète les conditions globales de celui-ci. En particulier, il a été montré l'existence d'espèces strictement paraliques (comme Ruppia Spiralis et Pirenella conica c'est-à-dire inféodées à ce type de milieu, inconnues en mer ouverte et indifférente à la salinité.
Nos travaux permettent ainsi de définir 6 degrés de confinement à partir des points singuliers de la répartition des espèces benthiques telles que nous les avons recensées dans les lagunes étudiées, tout au moins dans la zone la plus proche de la mer (Proche paralique) (PERTHUISOT et GUELORGET, 1982). (Cf. annexe et fig. 3).
Cette échelle de confinement a été établie de façon primordiale à partir des peuplements benthiques. Le phytoplancton, en tenant compte de son caractère instantanée et de ses fluctuations rapides en fonction des mouvements de masse d'eau, s'intègre parfaitement au schéma proposé (FRISONI, 1984).
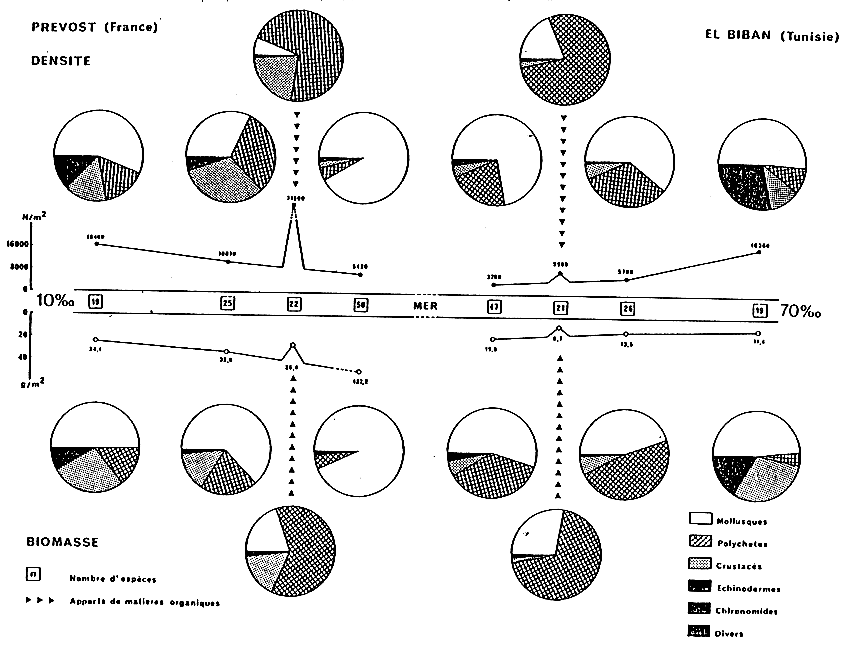
Toutefois, les variations quantitatives des peuplements phytoplanctoniques et benthiques (macrofaune invertébrée) sont différentes en fonction de l'éloigenement du domaine marin (Fig. 4).
En effet, la biomasse chlorophyllienne (exprimée en mg de chlorophylle a par m3 - dosage fluorimétrique) augmente régulièrement depuis les eaūx marines (valeurs proches de l'unité) jusqu'à la zone V. Ainsi les étangs situés en zone II et III présentent une biomasse comprise entre 1 et 4. Dans les étangs de zones IV et V dominantes la biomasse atteint des valeurs de l'ordre de 20 ; ces mēmes zones soumises par ailleurs à des pollutions d'origines urbaines et agricoles offrent des valeurs de biomasse de l'ordre de 1000.
Au-delà de cette zone V, l'allure de la courbe dépend du pôle vers lequel on se dirige (dulçaquicole ou évaporitique). En milieu ; dulçaquicole on assiste, soit au maintien d'une biomasse élevée (milieu eutrophe), soit à une chute de cette biomasse (milieu oligotrophe).
En milieu sub-évaporitique, on observe dans tous les cas une baisse brutale de la biomasse phytoplanctonique. Dans cette zone extrême le maillon primaire n'est plus guère représenté que par le microphytobenthos (tapis algaire).
La biomasse de la macrofaune benthique (exprimée en g de poids sec après décalcification, par m2) présente une courbe similaire. Cependant, on observe un décalage de celle-ci par rapport à la précédente : la position du maximum est reportée en zone III.
Toutefois, comme pour le maillon primaire, ce schéma peut présenter des fluctuations verticales (valeurs du maximum) en fonction des variations saisonnières, interannuelles, de l'enrichissement du milieu et des pollutions organiques.
On retiendra les valeurs suivantes :
- de l'ordre du gramme en zone I, de l'ordre de la dizaine de gramme en zone II, de l'ordre de 50 grammes en zone III peu soumise aux rapports organiques (El Biban) allant jusqu'à 500 grammes dans les zones très enrichies (Prévost).
Au-delà de zone III, la courbe traduit une diminution des valeurs jusqu'à la zone VI où, comme dans le cas du maillon primaire, deux possibilités se présentent :
- augmentation de biomasse par contamination de la faune dulçaquicole lorsque l'on reste en milieu aquatique dessalé ;
- disparition de la macrofaune benthique lorsqu'on se dirige vers le pôle évaporitique, terme de passage avec le domaine terrestre.
Ainsi, au niveau de la zone IV, la baisse de la biomasse benthique et l'augmentation de la biomasse phytoplanctonique se matérialisent par un croisement des deux courbes.
Dans la mesure où la majorité des étangs peri-méditerranéens se situe, à cheval sur les zones III, IV et V, ce phénomène se traduit dans le milieu par un antagonisme quantitatif entre macrofaune benthique et phytoplancton.
Cette diversité dans l'organisation biologique du domaine paralique entraîne l'existence d'une organisation fonctionnelle superposée.
Le confinement provoque l'apparition de gradients de fonction qui se traduit au niveau de chaque zone par une modification des relations trophiques.
Les gradients des biomasses phytoplanctoniques et benthiques depuis le domaine marin vers les confins continentaux (fig. 4) illustrent cette diversité fonctionnelle : en effet, l'augmentation progressive des biomasses de la macrofaune benthique jusqu'en zone III est liée à l'augmentation concomitante de la biomasse chlorophyllienne, Au delà, en domaine paralique propre (zone IV, V), la biomasse benthique chute (diminution de la richesse spécifique, nanisme des espèces lagunaires, conditions de milieu rigoureuses…), tandis que la biomasse chlorophyllienne continue d'augmenter, parfois de façon exagérée. Il en résulte une diminution de la consommation du maillon primaire par le maillon secondaire, ce qui entraîne une forte accumulation de matière organique. Enfin le long du gradient de confinement les espèces benthiques se relaient et l'on rencontre successivement des espèces thalassiques, des espèces mixtes (essentiellement suspensivores), puis des espèces strictement paraliques (essentiellement détritivores).
A une organisation biologique structurale se superpose une zonation fonctionnelle basée sur des espèces et des peuplements précis. Dés lors il est aisé de concevoir que pour une exploitation optimale des ressources biologiques du domaine paralique, par le biais de l'aquaculture en particulier, on ne pourra pas faire n'importe quoi, n'importe où.
II - APPLICATION DE LA NOTION DE CONFINEMENT A L'AQUACULTURE :
LA ZONATION AQUACOLE.
L'aquaculture désigne l'ensemble des activités humaines relatives aux problèmes d'élevage d'animaux (poissons, mollusques, crustacés) et plus rarement de culture de végétaux (algoculture) aquatique (AMANIEU 1974). Différents auteurs en ont proposé une typologie (FERLIN, 1982 ; QUERELLOU, 1980 ; RAVAGNAN, 1980). En ce qui nous concerne (FRISONI et GUELORGET, 1981), nous retiendrons la classification suivante :
| - 1) Elevage extensif | - valliculture, |
| - tapiliculture, | |
| - 2) Elevage semi-intensif en éclos ou lagunes, | |
| - 3) Elevage intensif | - grossissement en cages flottantes, ou en bassins, |
| - prégrossissement, | |
| - écloserie, | |
| - 4) Conchyliculture suspendue. | |
Pour les trois premiers types, cette classification tient compte d'une intensification progressive de la technologie appropriée et des besoins en énergie.
Par aquaculture extensive on considère l'exploitation de l'ichtyofaune lagunaire aux moyens de pêcheries fixes, sans modification notable du milieu naturel. L'exemple le plus typique de cette technique culturale étant la valliculture italienne, décrite par RAVAGNAN (1980) qui la décompose en plusieurs étapes d'aménagement depuis le type “primordial” (utilisation du milieu lagunaire en l'état intial) jusqu'au type “structuré” ou “équipé” nécessitant quelques aménagements comme l'endiguement ou le recreusement (Fig 5).
On inclut dans l'élevage extensif, la tapiliculture ou élavage des Veneridae (palourdes par exemple) en milieu ouvert et sur substrat naturel.
L'aquaculture semi-intensive comprend actuellement l'élevage en enclos ou lagune aménagée. Elle intéresse les mêmes espèces que le type précédent (muges, anguilles…) et pourrait connaître un essor dans l'élevage de certains crustacés comme les pénéides. La différence fondamentale avec l'élevage extensif réside dans l'apport de nourriture complémentaire (naturelle ou artificielle), qui est ici nécessaire.
L'aquaculture intensive utilise des structures artificielles soit à terre (bassins), soit en lagunes (cages flottantes). Elle repose intégralement sur un apport de nourriture extérieure. Elle fait appel à l'utilisation de technologies sophistiquées nécessitant une forte consommation d'énergie. La qualité de l'eau utilisée reste un des soucis constant de l'éleveur. Ce type d'élevage inclut toutes les étapes allant de l'écloserie au grossissement en passant par le prégrossissement. Il intéresse essentiellement les poissons (loups et sparidés), les crustacés (penceïdes), et les mollusques au stade de l'écloserie.
La conchyliculture suspendue concerne l'élevage des huitres (ostréïculture) et des moules (mytiliculture). Quelle que soit la technologie utilisée (suspension, à plat, bouchots, claire…) le type d'élevage repose sur l'utilisation de la productivité naturelle du milieu.
Ainsi cette classification arbitraire est également basée sur une diversité des exigences vis-à-vis des conditions de milieux notamment du point de vue trophique. Les gradients fonctionnels mis en évidence dans le domaine paralique auront donc une incidence sur la répartition des potentialités aquacoles de ce domaine.
Par ailleurs, l'existence de peuplements spécifiques au sein de chaque zone (espèces thalassiques, mixtes ou paraliques) implique l'exploitation en un site donné d'espèces inféodées à la zone d'accueil.
On doit alors admettre que la mise en valeur aquacole d'un milieu paralique doit impérativement tenir compte de son organisation biologique, c'est-à-dire de la nature des espèces et du degré de confinement.
A cet égard, tout développement en aquaculture nécessite au préalable une caractérisation écologique du milieu.
Cette caractérisation doit permettre de définir la zonation biologique du milieu selon son gradient de confinement. En fonction de cette dernière on peut alors proposer une zonation aquacole du domaine paralique.
Dans les zones les plus proches du domaine marin (zones I et II), on envisagera uniquement une aquaculture de type intensif. La faible teneur en matière organique des eaux et du sédiment, la permanence des courants d'origine marine, la relative stabilité des paramètres physicochimiques de l'eau confèrent à cette portion du domaine paralique une vocation pour l'élevage intensif. Actuellement celui-ci est essentiellement représenté par des écloseries, des élevages en bassin à terre, ou en cages flottantes et concerne des espèces thalassiques ou mixtes (le loup -Dicentrarchus labrax, la daurade -Sparus auratus, les pénéïdes -Penaeus japonicus et P.keraturus…).
Ces élevages qui concentrent un nombre élevé d'animaux dans un volume d'eau restreint exigent en effet une bonne qualité constante de l'eau.
La faible biomasse du compartiment primaire (phytoplanctonique en particulier) n'autorise pas les élevages extensifs qui, par définition, sont basés sur une alimentation d'origine naturelle.
Au delà de ces deux premières zones on pénètre en zone III caractérisée par de fortes biomasses phytoplanctoniques et benthiques. On peut envisager alors l'élevage d'espèces mixtes choisies parmi les pélécypodes filtreurs qui dominent largement les peuplements de cette zone tant en substrat meuble (palourde -Ruditapes decussata) qu'en substrat dur (moule -Mytilus galloprovincialis, huitres -Ostrea edulis, Crassostrea gigas).
La richesse trophique de la zone III permet également l'élevage semi-intensif d'espèces “mixtes” de poissons et crustacés. Les conditions de milieu caractéristiques de la zone III (bathymétrie, nature du substrat…) autorisent souvent) l'utilisation d'enclos ou de parcs.
Dans cette zone, il faut signaler le développement d'épibiontes filtreurs (Ascidies, balanes, bryozoaires…) qui participent à la salissure des structures d'élevage et entrent en compétition avec les animaux élevées.
Cependant, le développement inconsidéré de la conchyliculture entraînant la multiplication des structures d'élevages en espace restreint, risque à moyen terme de déséquilibrer le milieu d'accueil. En effet, la concentration des structures perturbe le régime hydrodynamique en ralentissant les courants et en modifiant leur trajet, et favorise l'accumulation de matière organique essentiellement sous forme de biofécès.
La barrière physique ainsi constituée peut alors décaler le milieu dans l'échelle de confinement et diminuer les potentialités conchylicoles de la zone considérée.
La circulation des eaux, la dispersion et la biodégradation des rejets organiques peut être améliorées par la mise en place de structures flottantes et mobiles couvrant à charge égale une plus large superficie.
Dans les zones de confinement avancé (zone IV et V) se rencontrent uniquement des espèces paralique généralement de très petite taille (cf.infra) dont on ne connaît pas de représentant susceptible de faire l'objet d'une expoitation rentable.
Bien qu'en quantité abondante le phytoplancton est ici représenté par les flagellés (Dinoflagellés - Gymnodinium…; Chlorophycées - Nanochloris… par exemple) parfois toxiques pour les filtreurs conférant alors aux eaux une qualité nutritive médiocre et souvent incompatible.
Les zones IV et V sont généralement constituées par des tranches d'eau faibles, forte amplitude thermique et haline… et un engraissement organique important. Toutes ces conditions environnementales favorisent l'apparition de crises dystrophiques remettant ainsi en cause l'implantation d'élevage en raison des mortalités estivale qu'elles entraînent.
Dans ces zones confinées on est limité à l'élevage extensif de poissons comme les muges, les anguilles, espèces paraliques dont l'éthologie est adaptée aux conditions de milieu. Cependant aménagements hydrauliques (valiculture par exemple) sont nécessaires pour limiter les effets néfastes du confinement (dystrophie…).
Dans le paralique lointain, au-delà des tapis algaires, et en domaine évaporitique on peut envisager l'exploitation (voir l'élevage…) de population monospécifique d'Artémia salina qui présente un intérêt certain comme source de nourriture en aquaculture intensive (écloserie par exemple). Ce crustacé phyllopode est particulièrement abondant dans les partements saliniers les plus concentrés, associant à la potentialité minérale des salins une vocation également de production biologique.
Cette organisation zonale des milieux littoraux appliquée à l'élevage d'animaux aquatiques tant vertébrés, qu'invertébrés peut être étendue au règne végétal ouvrant la voie à l'algoculture depuis les algues unicellulaires planctoniques aux monocotyledones.
CONCLUSION
L'organisation biologique du domaine paralique dépend étroitement des relations avec le domaine marin. L'éloignement (hydrologique) de la mer au fur et à mesure que l'on pénètre dans le domaine paralique et l'épuisement progressif d'éléments vitaux d'origine marine, que l'on peut traduire en terme de confinement, modifient la nature des peuplements en place et les relations trophiques existant entre les différents composants biotiques de l'écosystème. Des peuplements différents se succèdent salon une zonation marquée depuis les communications avec la mer jusqu'aux zones les plus confinées.
Au cours de ce cheminement se rencontrent alors des espèces à l'éthologie très différentes -filtreurs et migrateurs dans les zones marinisées-limivores, brouteurs, détritivores fouisseurs dans les zones les plus reculées.
Il apparaît ainsi que l'implantation d'espèces aquacoles dans un milieu littoral n'est possible qu'autant que l'éthologie de cette espèce corresponde aux potentialités biologiques offertes par la zone dont la mise en valeur est envisagée.
Il faut alors insister sur la prise en compte préalable de l'organisation biologique du milieu avant tout projet d'implantation de structure et d'espèce aquacole. L'investigation de paramètres biotiques et abiotiques doit permettre une caractérisation écologique du milieu. Ainsi pourront être définies les potentialités aquacoles de l'écosystème dont nous avons montré que chaque zone est caractéristique de certaines filières aquacoles. En effet, un lagune n'est pas fortement utilisable dans son intégrité et n'importe quel développement technologique ne peut être envisage n'importe où.
Ainsi une lagune confinée, située dans sa majorité en zone IV, peut être décalée vers la zone III par un aménagement hydraulique approprié (ouverture ou élargissement de passe, meilleure orientation des flux). Le déconfinement qui en résulte augmente ainsi les potentialités conchylicoles du milieu.
Outre, une meilleure adéquation entre projet de développement et capacités naturelles du milieu, une parfaite connaissance de l'écosystème nous laisse envisager une optimisation de ses ressources naturelles.
Sur la base d'une connaissance écologique du milieu suffisante peut donc être envisagé un aménagement léger respectant l'organisation initiale de l'écosystème tout en optimisant ses ressources naturelles.
Nous avons cependant montré que la mise en place de structures d'élevages favorisaient parfois le décalage du milieu dans le sens d'un confinement excessif (ralentissement des courant - apport de matière organique…).
Une parfaite connaissance de son organisation biologique et surtout une observation suivie de cette organisation doit permettre la mise en place d'aménagements appropriés dans le but de conserver au milieu ses capacités de production et de mieux les gérer.
Enfin, la prise en compte de cette organisation propre au domaine paralique nous laisse espérer la mise en place de techniques reconstituant les chaînes trophiques rencontrées dans ces milieux. C'est ainsi qu'l n'est pas illusoire d'envisager la mise en place d'élevage “stratifié” permettant la cohabitation de filteurs (coquillages) et de détritivores (crustacés): l'installation des premiers dans la zone appropriée favorisant l'installation à la périphérie (ou en profondeur) d'une zone plus confinée favorable aux seconds.
La diagnose écologique du milieu apparaît ainsi en domaine paralique comme un atout pour une meilleurs préservation du milieu et pour une optimisation de son exploitation.
EL BIBAN
LE PREVOST
Champ de Salinité
Fig 1
Repatition de peuplement
Fig 2
Fig 3
La Zonation Biologique
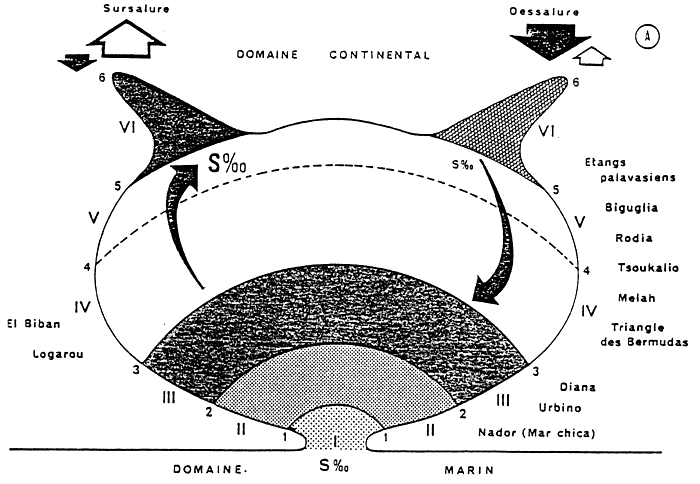
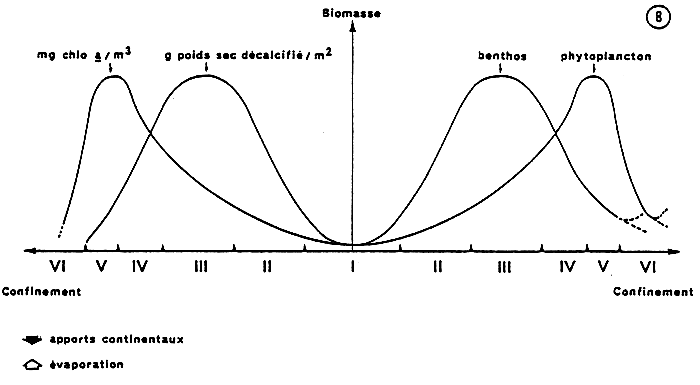
Fig 4
La gradient quantitatif
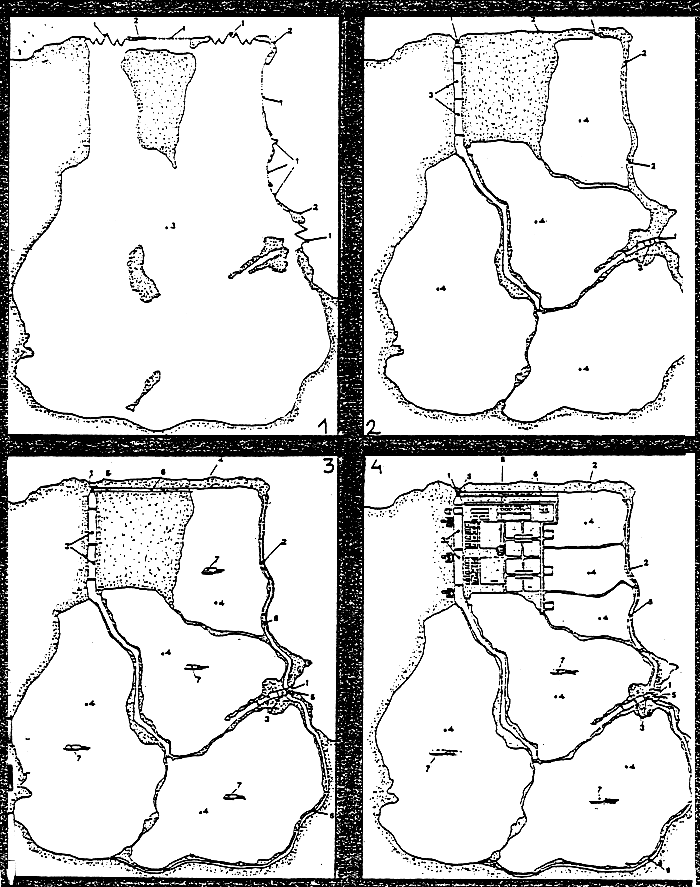
| 1. | Dispositifs de communication contrôlée avec le milieu extérieur | |
| 2. | Digues | Dela Pisciculture “pimordiale |
| 3. | Bassins de récolte | [d'aprés G. RAVAGNAN] |
| 4. | Bassins de pâturage | |
| 5. | Station de pompage | |
| 6. | Canaux collecteurs | |
| 7. | Fig. 5 | |
ANNEXE
«Zone I - Zone comprise entre les degrés 0 et 1: dans cette zone située à proximité immédiate des communications avec la mer et qui n'est qu'une continuation du domaine marin, la macrofaune est composée de nombreuses espèces strictement thalassiques appartenant généralement au cortège faunistique de la biocénose des Sables Fins Bien Calibrés (S.F.B.C.) (Pérès et Picard, 1964).
Zone II - Zone comprise entre les degrés 1 et 2: à partir du degré l, on peut considérer que la pénétration dans le domaine paralique est effective. La macrofaune benthique est caractérisée par la perte des espèces sensibles strictement marines, les plus «sténohalines». On y rencontre encore des Mollusques: Mactra corallina M. glauca. Tellina tenuis, Donax semistriatus, D. trunculus, Acanthocardia echinata, Dosinia exoleta, les Polychètes: Audouinia tentaculata, Magelona papillocomis, Owenia fusiformis, Phyllodoce mucosa, Pectinaria koreni, les Crustacé: Portumnus latipes, et les Echinodermes: Asterina gibbosa, Holoturia polii, Paracentrotus lividus, et enfin notons la présence de Branchiostoma lanceolatum dans les faciès sableux exempts de «pollution organique».
La macroflore phanérogamique est marquée par la perte de Posidonia oceanica et la dominance de petites Phanérogames telles que Cymodocea nodosa, Zostera noltii et de Caulerpa prolifera
La limite de cette zone (degré 2) est marquée par la disparition totale des Echinodermes.
Zone III - Zone comprise entre les degrés 2 et 3: cette zone est dominé par «espèces mixtes» (1) (Venerupis decussara. V. aurea, Scrobicularia plana. Carbula gibba. Loripes lacteus. Gastrana fragilis, Akera bullata, Nephthys hombergii, Armandia cirrosa, Glycera convoluta, Upogebia littoralis…) en ce qui concerne la macrofaune benthique.
Zone IV - Zone comprise entre les degrés 3 et 4: le degré 3 est défini par la perte de toute faune thalassique et les espèces présentes sont strictement paraliques: (Abra ovata, Cerastoderma glaucum, Hydrobia acuta. Nereis diversisolor. Gammarus insensibilis. G. aequicauda, Corophium insidiosum…)
La macroflore est caractérisée par l'apparition de Ruppia spiralis.
Zone V - Zone comprise entre les degrés 4 et 5: dans cette zone, la production phytoplanctonique (Diatomées pennées, Cyanophycées) est maximale: la surface du sédiment est en voie de colonisation par les Cyanophycées. Ainsi la sédimentation est essentiellement organique et le milieu interstitiel hautement réducteur.
Le peuplement est composé essentiellement d'une faune vagile comprenant des Crustacés détritivores (Sphaeroma hookeri, S. rugicauda, Corophium insidiosum, Idotea balthica), des Gastropodes brouteurs (Hydrobia acuta, Pirenella conica), des Polychètes comme Nereis diversicolor, et des larves de Chironomidae dans la tranche tout à fait supérieure du sédiment (quelques millimètres). Dans cette zone, et tout particulièrement dans les milieux très sous-salés, apparaissent les éléments de la faune dulçaquicole (Tricoptères, Oligochètes, Odonates). Par ailleurs, dans les milieux sursalés, on peut éventuellement rencontrer des éléments de la faune évaporitique (Artemia salina).
Les herbiers, lorsqu'ils existent, sont dominés soit par les characées, avec évantuellement des herbiers mixtes.
Zone VI - Zone comprise entre les degrés 5 et 6: cette zone représente le passage au paralique lointain, soit dulçaquicole, soit évaporitique et se caractérise habituellement par la colonisation quasi-totale du substrat par les cyanobactéries formant des tapis algaires ou des édifices stromatolithiques. La passage à l'eau douce est marqué par l'apparition d'espèces dulçaquicoles strictes et par la persistance de quelques espèces paraliques vagiles (Sphaeroma hookeri, Microdeutopus gryllotalpa, Gammarus insensibilis). Les milieux sub-évaporitiques se caractérisent par l'absence de macrofaune benthique, à l'exception de brouteurs qui fréquentent momentanément cette zone pour des raisons trophiques (broutage des tapis algaires).
Aux confins de cette zone s'arrête le proche paralique avec la disparition des Foraminifères qui coincide, vers le pôle évaporitique, sensiblement avec la disparition des édifices stromarolithiques bien constitués.
Au-delà, dans le paralique lointain, le rôle du confinement semble s'atténuer, du moins dans l'état actuel de nos connaissances.»
![]()
![]()