![]()
![]()
![]()
Helen Newton Turner
INTRODUCTION
This paper deals with the role of selection in increasing the efficiency of production from small ruminants. Increasing the efficiency of output from the animals, as discussed, will fail unless attention is also paid to problems of marketing and distribution of the product. Australia is renowned for its wool production per head, but over the last 10 years the wool marketing system has been revised to increase the efficiency of the whole system. And currently, Australia is in trouble over milk marketing.
In developing countries also there may be no encouragement for owners of small flocks and herds to increase production of wool, meat or milk because they will receive no benefit without a revision of the ways in which their product is presented for sale, marketed and distributed. Further, it may not be possible to implement production improvements without access to credit.
Genetic improvement is a simple, relatively cheap and permanent way of increasing animal production, but disease control and better nutrition also make major contributions. And improvement relating to the animal is only part of a whole system. Remembering this, discussion of genetic techniques for animal improvement becomes profitable, and selection is a major tool among those techniques.
This paper makes no Claim to be a complete literature review - examples are chosen to illustrate points.
Selection may involve the choice of:
Individuals within a flock.
The small ruminants under discussion are sheep and goats, while increased efficiency is defined as an increased ratio of output (lifetime production) to input (labour, feed, management costs). The main products for sheep are meat, wool and milk, and for goats, meat and milk, plus fibre in the case of some special breeds. Basic to efficiency of production for either species is a high reproduction rate, which means more surplus animals for sale as meat, and a higher selection differential, leading to a faster response to selection.
There has been considerable discussion as to whether selection should be based on production per head, or per unit of input. A general conclusion has been (Turner and Young 1969) that production per head is preferable for grazing animals because:
Feed intake for grazing animals is costly to measure.
CSIRO, P.O. Box 184, North Ryde, Sydney, Australia.
If milking animals are artificially fed and production recorded, or if meat animals are lot-fed, then selection on output per unit of feed may be justified, while production per hectare may be needed in selecting species or breeds.
In this paper, increased production per head is taken as the criterion of increased efficiency for selection of individuals within a flock or for choosing between cross-breeding and selection, while the need for considering other factors is discussed in relation to choosing species, breed or strain.
CHOICE OF SPECIES
Influencing Factors
Discussion in this section will merely draw attention to the need for considering the best species for the environment, without detailed analysis. Some of the important factors are:
Management costs.
Adaptation
Some questions of adaptation have clear-cut answers, while others require further research. Heavily-woolled sheep, for example, do not thrive in warm, humid environments, where the choice must lie between hair sheep, sheep with coarser, lighter fleeces, goats (other than fibre-producing) or cattle.
The browsing habits of goats makes them useful in shrubby or partly-degraded semi-arid areas compared with grazing sheep (Harrington 1982, Wilson 1984), but the comparative digestive efficiency of the species for various feedstuffs may vary with breeds and circumstances. (Devendra 1978 and 1983, Brown and Johnson, 1984, el Hag et al. 1984).
Impact on the Environment
Goats or cattle may damage collection cups if run under rubber trees, whereas sheep are less likely to do so. Goats are alleged to cause erosion on steep, treeless hillside sbecause they form playgrounds; would sheep be any better ? Goats can be used to control shrubs, but will they cause erosion when the shrubs are gone ?
Markets
Markets dictate the product required. If this is clothing, the sheep, with its wool, may be chosen. If meat is the product, does the consumer prefer mutton, beef or goat meat ? Is there a high incidence of children who can tolerate goat milk but not cow's milk? When markets are distant, as in inland temperate Australia, sheep are favoured because wool can readily be stored and transported, whereas meat and milk require special facilities.
Management Costs
Woolled sheep require shearing, and in some areas regular inspection for flystrike. Goats and cattle require less oversight; cattle may also be less susceptible to disease.
CHOICE OF BREED OR STRAIN
Many genetically different breeds exist, and often genetically different strains within a breed. The problem in each case is to choose between groups, as distinct from selecting individuals within a group, so the two will be considered together.
Influencing Factors
Most factors apply as for choosing species, though details may differ.
Adaptation : Adaptation is again of primary importance. The many breeds of sheep and goats vary in physical appearance and production levels, and are grown over a wide range of environments. These environments differ in:
Exposure to disease (including internal and external parasites), and to predators.
Sheep and goat breeds differ in their products; woolled sheep give either apparel wool or coarse, carpet-type wool, as well as meat, skins and sometimes hair. Angora and cashmere goats give fibre and meat, while other goats give meat and sometimes milk, as well as hair and skins. There are some broad adaptations among these classifications; hair sheep and many goats are adapted to hot, humid climates, while apparel-woolled sheep and Angora goats are not.
The FAO Production Yearbook divides countries into three main categories-centrally planned economies, developed and developing market economies. In general the developed countries have a better environment for sheep and goats than the developing; many of the latter lie in the tropics, with the animals exposed to harsh climate, poor nutrition and a range of diseases. Production per head in the developed countries is higher than in the developing (e.g. 4–5 kg. of wool for sheep in Australia, compared with 1–2 kg, in India).
An often occurring mistake in the past has been to assume that animals can be transferred from temperate areas to raise production in harsh environments. Local breeds are adapted to their harsh conditions; exotic imports often fail to survive or even reproduce.
In recent years the importance of adaptation has been recognised; FAO is now in the process of establishing data banks of production records for livestock breeds in various countries, together with details of their environments. These should assist choice of breed (FAO 1985).
Impact on the Environment
This has received less attention in breed than species comparisons; its possible importance is stressed by Brooke and Ryder (1978).
Markets
These are important for choice of breed, strain or individual animal. There are broadly three types - world market, local market, and home use. The world market for wool pays higher prices for white apparel wool of fine diameter, with a low percentage of medullated fibre, or white carpet-type wool with some medullation. A local market may consume carpet-type wool, tolerate a higher level of medullation, and even seek some pigmented wool. If the wool is for home use in handcraft, coarser wools may be preferred for hand processing and pigmented wools may be sought. Similarly a home market may not pay attention to distribution of meat over the body as all is cut into small pieces for cooking, whereas a world (or developed country) market may give different prices for different cuts.
The whole marketing infrastructure, including distance from market, may also influence choice of product and breed.
Management Costs
These are related to adaptation. Inputs for animals in a harsh environment are likely to be lower for adapted local breeds than for imports.
Performance Data
Need
Performance data are essential for a final choice of breed. There have been movements to collect such data for decades, but the traits recorded in various places are not always the same. FAO is currently involved in a major project aimed at bringing available information together in data bases, standardizing the traits recorded, and describing the environment as well. One deficiency at present is that many of the records are made on experiment farms and not under field conditions, where the environment may be different.
Examples of summaries of available data for sheep and goat breeds will be found in Mason (1967, 1980), SABRAO (1980 and 1982), PARC (1982), Wahid (1982), Acharya (1982), Yalcin (1979).
These summaries are nearly all in what SABRAO has termed the documentation stage of recording, where animals have been run in different places. Critical breed selection should be based on evaluation, when breeds (or strains) under comparison have been run together, or in different places with a reference breed included. Collection of such information is long-term and expensive but a few experiments can be quoted. For example, Dunlop (1962, 1963) and Dun et al. (1966) compared Australian Merino strains, Atkins (1980) Australian sheep breeds, and Farid et al. (1977) Iranian sheep breeds.
Because of the paucity of such evaluations, breed (or strain) choices will often have to be made without them. Available data must then be used, seeking information from as many sources as possible, and on adequate numbers. Breeds from areas with similar environments should be sought.
Important Criteria
Characteristics to be considered depend on species and main product, and should describe also the ability of the animals to survive and reproduce. They are:
| For all: | Number of offspring weaned/year/female exposed to the male, |
| Annual death rates (each sex), | |
| Length of productive life (each sex), | |
| Incidence of major defects. | |
| For meat production: | Total weight of offspring weaned/year/female exposed to the male, |
| 12–15 month body weight (each sex). | |
| For Fibre Production: | Annual clean fleece weight (sheep and Angora goats), |
| Annual down weight (cashmere goats), | |
| Average fibre diameter, | |
| Percent medullated fibres (sheep), | |
| Percent kemp (sheep and Angoras), | |
| Length, | |
| Strength. | |
| For milk production: | Production per lactation, |
| Lactation length, | |
| Fat percent. | |
| Protein percent. |
Lifetime production is important, and can be obtained only approximately from the above figures, but actual data are rare.
Many performance records include birth weight and body measurements. These may be useful in research programmes, but are less useful in estimating the productive value of a breed.
Products not mentioned above include skins (for both species) and hair (goats and hair sheep), which are not usually objectives for selection. Fur skins from certain sheep breeds, obtained from day-old lambs, are a veryspecialized product; much research has gone into their production, but they are not discussed here.
Possible Gains
Sheep
Indian data can be used as an example to examine possible benefits of breed selection. Acharya (1982) noted four environmental regions-Northwestern arid and semi-arid, Southern Peninsular, Eastern and Northern temperate. Taking the Northwestern region as an example, he listed 11 sheep breeds, of which 5 are used for milk and 6 are not. Among those 6, Chokla and Nali have greasy fleece weights well above any others (1.37and 1.46kg. for 6 months, compared with 0.8 – 1.09) finer fleeces (28 and 35 μm, compared with 32–39 μm) lower percent medullation (24 and 31 percent, compared with 48–65), but lighter 12-month body weights (18kg. compared with 19–28) and comparable mortality rates. Figures for reproduction rates are very variable.
These figures indicate that replacement of other breeds by Chokla or Nali in the N.W. Region would lead to increased wool production, and in fact Nali rams have been used to a limited extent for wool improvement in two of the breeds normally milked (Muzzarfarnagri and Jalauni, with fleece weights of only 0.65 and 0.90 kg. respectively).
Goats
Acharya (1982) also listed 11 goat breeds in the N.W. region, the Beetal and Jamnapari being outstanding for body weight (12-month weight 22 and 30 kg, compared with 15–21 kg.) and milkproduction (201 and 177kg/ lactation, compared with 122or less) and are being used for upgrading other regional breeds.
Hair is harvested from some goat breeds, though production is not mentioned for Beetal or Jamnapari, and quantity of hair does not seem to merit consideration in selection.
CHOICE OF SELECTION OR CROSSING
When data on breeds and environments have been assembled, a choice has to be made between selection or cross-breeding. The latter may be used to exploit hybrid vigour, or to change the genotype by complete replacement or development of an intermediate. If change of genotype is involved, selection will be combined with crossing.
Exploiting hybrid vigour means maintaining pure breeds so that crosses can be continually made. Gains with small ruminants are slight except for reproduction rate; the Australian prime lamb industry is based on first cross Merino × Border Leicester or Merino × Dorset ewes. The technique has a place in developed economies, but is more difficult to exploit in developing ones, where most flocks and herds are small.
Too often in the past crossing to change the genotype has been automatically chosen for developing economies; the low performance of local breeds in their usually harsh environments has been compared with the far superior record of exotic breeds in temperate environments, and the exotics have been imported in the hope of improving the local breeds. All too often the result has been disastrous. No large-scale imports should be considered without pilot trials evaluating the proposed imports (or their crosses with local breeds) against the local breeds when run in the local environment. Usually the environment will have to be modified for the imports (or their crosses) to perform; the effect of such modification on the local breed should be recorded, as well as the inputs required for the modification.
Not all introductions of exotic breeds have failed; temperate sheep breeds such as the Corriedale have been introduced to the Peruvian Andes and to the lowlands of Patagonia as purebreds, and to the Himalayas in Kashmir for crossing with local breeds. The Australian Merino has been developed from the first imports of coarse-woolled and hair sheep from Bengal and the Cape of Good Hope, crossed with a variety of European breeds (Turner 1982, Garran and White, 1985). The failures have come from taking temperate sheep breeds into very harsh environments, with environmental stress from heat, humidity, poor nutrition and unaccumstomed diseases.
Crossing to change the genotype, using breeds from similar environments, is being practised in a number of places, particularly in relation to raising specific characteristics such as reproduction rate. A few sheep breeds in the world have outstanding lambing percentages, and are being used in this way; the Finn has been taken into a number of countries, the Romanov is used in France, and the Booroola Merino (whose high fecundity is due to a single gene) in Australia (Bindon et al., 1985).
Unfortunately failed imports, or failed crosses, have often not been recorded; it is hoped the planned FAO data bases will record both successes and failures for future guidance.
SELECTION OF INDIVIDUALS WITHIN A FLOCK
Background
Selection of individuals improves production in the current flock for the lifetime of the selected animals, these usually being females and castrates, with males in smaller numbers. Of more importance are genetic gains in the next generation, part of which come from the selected females and part from the selected males; these gains are permanent and cumulative.
The classic formulae related to selection are:
| ΔC | = | iRσ | (1) |
| ΔG | = | īh2σ | (2) |
where
| ΔC | = | gain in current flock |
| ΔG | = | genetic gain per generation |
| i | = | selection differential of selected animals, in standard units |
| ī | = | average selection differential of parents, in standard units |
| R | = | repeatability |
| h2 | = | heritability |
| σ | = | phenotypic standard deviation |
The controlling elements are permanent environmental variation (Rσ) in (1),genetic variation (h2σ) in (2) and in both selection differential, dependent on proportion saved. A high reproduction rate raises the number of animals available for selection, and in a flock of fixed size lowers the proportion saved, increasing the selection differential. Artificial insemination increases the male selection differential still further (Table 1). So far frozen semen is not as successful with sheep and goats as with cattle, and use of fresh semen sometimes causes problems with small, scattered flocks or herds. Improvements in the techniques for using frozen semen will be a great advantage; laparoscopic insemination is giving better results than previous techniques, but may not always be feasible.
A distinction must be drawn between characteristics under selection which can be measured in both sexes (e.g. fibre weight and liveweight) and those which are sex-limited (e.g. reproduction rate, milk production). For the latter, a male's female relatives must be measured; attempts to find measurable indicators in males have so far failed.
Genetic gain per generation can be converted to annual genetic gain ( ΔGa) through dividing by the average age of the parents when the offspring are Born.
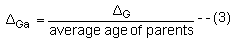
Here the average age of the parents becomes important in choosing which female relatives should be measured for sex-limited characters.
Formulae (1 and 2) apply to single characteristics, or to indices combining a number of traits, using economic weights, heritabilities and genetic correlations. Many such indices have been developed, and computers have simplified their calculation and use. If computers are not readily available or if measurement costs are high, there is still merit in independent culling levels, which in some circumstances are no less efficient, in terms of genetic gain, than the indices.
Table 2 gives available estimates of heritability for the most important performance criteria.
Selection should be made as early as possible in an animal's life. First matings for sheep are usually at 1–1/2 years, though with some breeds earlier matings are possible. Selection can be based on measurements at 12–18 months for characters not sex-limited. Heritabilities for these are sufficiently high for selection to be based on an animal's own performance unless artificial insemination is being used; it would be wise to progeny test sires before wide use with AI. The advantages of earlier matings are discussed below.
For reproduction rate and milk production, no measurements are available till after a female's first parturition. Lifetime performance of the dam is then a useful selection criterion for both sexes. Again, if AI is to be used, progeny-testing of the sire is desirable.
If full records on the dam are not available, then for the female production of multiple births at her first parturition, or her first lactation record, may be used. The advantage of using dams' records is that the female can be selected before she enters the breeding flock.
Vital Statistics
These include reproduction rate (number of offspring born and weaned per year per female exposed), death rates, length of productive life, and defects.
The length of the breeding season varies for both sheep and goats; near the equator, all-year-round breeding can occur, though there may be seasonal differences in ovulation rate. Obst et al. (1980) in Indonesia found an average ovulation rate of 1.2in Priangan sheep in September – October, compared with 1.7 – 1.9in other months. Lawson and Shelton (1982) in Texas found no Spanish does ovulating in March and April, with rates ranging from 1.17in May to 1.86in November. In higher latitudes, the breeding season is more restricted.
Increasing the frequency of parturitions is a technique for higher reproduction rates which has received more attention in recent years, three parturitions in two years being a common aim. The number of offspring born (or weaned) per female per year then is:
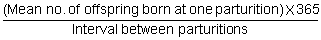
Reproduction rate - One parturition per year
The components of reproduction for yearly parturitions are:
Survival rate of offspring to weaning.
Simple arithmetic indicates that increased litter size will give more rapid progress in raising the number of offspring born than reducing the proportion of barren females. The upper limit through the latter route is 100% ; if all females produced twins, the limit would be 200%, and so on. Survival rates are usually lower for offspring from multiple births, but only on rare occasions are they sufficiently lower to offset the increased number born (Turner and Young 1969, Hanrahan 1982b). For twin births, the survival rate must be less than half that of singles, for triplets, less than one-third, for quadruplets less than one-quarter, and so on. Decreases in survival rate of this order have seldom been reported.
Although advantages in numbers from multiple births are not eliminated, higher death rates and slower growth rates lower productivity from their potential, and French workers have developed systems of early removal and artificial feeding of lambs which increase productivity, though with some financial cost (Thimonier et al. 1975). The risk of high mortalities has led some workers (e.g. Peacock 1982) to recommend against selection for twins in sheep in severe environments; she nevertheless recommended such selection in goats.
Sheep : Successful selection for increased average litter size at birth is evident in the few highly prolific sheep breeds of the world, such as the Finn (Finland), the Chios (Greece), the Dahman (Morocco) and Hu (China), the Barbados Blackbelly (Caribbean) and the Booroola Merino (Australia). All have been developed originally in small, carefully managed flocks. The length of time required to reach the current high average litter size is not known, except for the Booroola Merino (Turner 1978, 1982). When CSIRO acquired its sample in 1958, the Seears brothers had been selecting their flock (on the ewe side only) for 10–15 years, and had reached an average litter size of around 1.9;by 1984, following selection in both sexes, the average litter size in the CSIRO flock was 2.5, with an average ovulation rate of 4.2 (Piper et al. 1985). Selection has been based on lifetime performance of the dam, adjusted for age, for rams, and this plus the ewe's own performance for ewes.
Losses among multiple-born lambs increase with litter size but, as stated before, the increases in death rate are seldom enough to eliminate the raised numbers. Ewe progeny of Booroola rams by two other strains of Merino ewes weaned 16% (Piper and Bindon 1982) and 28% (McGuirk et al. 1984) more lambs than the progeny from rams of the respective strains.
Since lambs from multiple births are smaller than those from single births, economic returns from weight of lamb weaned have also been studied. A comparison of crossbred dams from Booroola and random Merinos by two meat breeds has shown that the Booroola ewe crosses produced annually 27% more lambs and returned 24% more dollars per head than the control crosses (Bindon et al. 1985).
The fast rate of selection response with the Booroola has been explained by the presence of a single gene (Piper and Bindon, 1981; Piper et al. 1985), and this discovery has stimulated a search for single-gene control of litter size in other prolific sheep breeds. Selection response has also been obtained where no single gene has been suspected. Turner (1978) reported an initial gain of 35 in the percentage of peppin Merino ewes with at least one multiple birth in their first 3 lambings through selection of twice-twinning base ewes, and an annual increase of 2.4over the next 14 years. The annual gain in lambs born per 100 ewes exposed was 2. Rzepecki (1979) reported that lambing percentages increased from 114 to 160 over 15 years (= 3 lambs/100 ewes/year) in a closed flock of Polish Merinos. Other successful responses have been reported by Wallace (1958) and Clarke (1975) with the N.Z. Romney, and Owen et al. (1980) with British breeds.
In most of these reports selection was on litter size at birth, in the case of the Australian experiments using lifetime records of the dam. Hanrahan (1982a) reported selection on the ewe's own ovulation rate and body weight in Galway ewes, with increases over unselected ewes of 0.35 in ovulation rate, 0.37 in litter size and 2.4kg. in body weight. The aim is earlier selection of the ewe on her own performance.
A fitting conclusion to this section is Mason's statement (1980, p.97) after reviewing prolific sheep breeds: “It is therefore tentatively concluded that almost any breed of sheep could be selected for prolificacy and the principal requirement is that the owner should conceive the idea”.
Goats: Many authors have reported high average litter sizes for some goat breeds e.g., Hoist and Pym (1977, Australia), Acharya (1982, India) Devendra (1983, Malaysia), Wilson (1984, Africa), Cheng (1985, China). There seem to be no reports, however, of results of selection to increase reproduction rate.
Reproduction Rate - More than one parturition per year
The components are the same as for one parturition per year, plus the interval between parturitions, which mainly depends on the length of postpartum anoestrus.
Sheep : Although many successes have been reported for more frequent lambings, there have also been failures, and I have been unable to find any references to the results of selection to increase lambing frequency. Hunter (1968) gave a very comprehensive review of information up to that time, while the possibility of selecting ewes more capable of sustaining frequent lambings has been discussed by Bernard and Fahmy (1974) and Lindahl and Terrill (1975).
For sheep which can come into oestrus all the year round, matings could occur twice a year if postpartum anoestrus lasted no longer than approximately 30 days. Mason (1980, Table 24) gives lambing intervals with ranges from 6 to 8–9 months for the hair sheep of the Americas, 6 to 7–9 months for the Asian and African and 6–7 to 10–12 for the European prolific breeds. These ranges would indicate the possibility of selecting towards twice-a-year lambing, but Mason reports only Spurlock (personal communication) as using this selection, with the Barbados Blackbelly, and he settled on 3 lambings in two years. Management requirements and the desired product dictate the appropriate frequency; if milk is a main product, more frequent lambing will interfere with lactation.
Goode et al. (1980) tried 4 lambings in 26 months with 3 crosses (Barbados × Borset, Finn × Dorset, Finn × Rambouillet) and pure Suffolk; only the Barbados cross produced any lambs at all at the fourth lambing.
Systems of lambing 3 times in 2 years have been more often successful, though not universally so. Lax et al. (1979) suggested it was not feasible for breeds in Wisconsin. Notter and Copenhaver (1980) used Finn × Rambouillet and Suffolk × Rambouillet crosses in an 8-monthly lambing programme over 5 years. The half-Finn ewes of different ages bore from 10–19 lambs over the period, compared with 7–10 for the quarter-Finn and 7 for the Suffolk cross, but both conception rate and litter size declined with successive Iambings.
Fogarty, Dickerson and Young (1984, 1985) presented an extensive analysis of an accelerated lambing trial (3 in 2 years) with 5 breeds and 2 crosses over 4 years. They concluded that “the major limitation for accelerated lambing was fertility and litter size born. These limitations might be overcome by genetic changes in length and level of breeding season fertility, and in lambing rate”.
The possibility of selecting for ability to lamb every 8 months, however, remains unanswered, more work on long term selection is needed, including an assessment of the effect on lifetime performance.
Goats : Not as much work on “accelerated” parturitions has been done with goats as with sheep, possibly because bucks often run continuously with does. Wilson (1984) quoted mean kidding intervals of 200–310 days (depending on parity) for African breeds, Devendra (1983), 259–360 days for Malaysian breeds and Acharya (1982), 229–376 days for Indian breeds. There is therefore scope for 3 kiddings in 2 years with controlled matings, but assessment of the effect on lifetime performance is desirable.
Annual death rates and length of productive life
Though these are important aspects of efficiency of production, there is no work on genetic aspects for either species.
Major defects
Nicholas (1985) has prepared a bibliography of papers reporting genetically controlled defects in domestic animals.
Meat Production
Number of surplus animals is the main component of meat production; the others are the quantity and quality of meat per animal, the latter depending mainly on the absence of excessive fat. Liveweight has been taken as the best single measurement in selection for increased lean meat production in sheep (Tallis, Turner and Brown, 1964, Bradford and Spurlock, 1972), but with consciousness of the association of fat with health problems there is now an increased demand for fat estimation in selection for meat production. Since goats produce leaner meat, the need with them is not so great.
Weaning Weight
Sheep: Weaning weight has a lower heritability than later liveweights (Table 2), but selection responses have been obtained, with gains of approximately 2% per annum over a random control (Pattie 1965) for a period of 10 years.
Goats : No selection reports have been found, but a heritability estimate of 0.35 for 5-month weight in Australian feral goats indicates that there should be response to selection.
Total weight of offspring weaned
This is a useful selection measure for ewes, which has been adopted in the New Zealand Sheep Performance Recording Scheme, Sheeplan. It combines number of lambs born, lambs' survival rate and lambs' weaning weight.
Other body weights
Responses to selection for yearling or 15-month weight in sheep have been obtained, e.g. in the Australian Merino (Turner et al. 1970). Initial selection of base animals gave a gain of 10% over a random control: during the next 14 years the gain increased by 1.5% per annum.
Fibre Production
For both species, weight per head per annum is the criterion for quantity-clean weight for preference, but the correlation with greasy weight is high, and this can often be used. Where animals are shorn more than once a year, weights of part-time shearings are added.
Sheep
Wool can be divided broadly into two categories, apparel and carpet-type. The main quality characteristic is average fibre diameter, required to be low for apparel wool and higher for carpet-type. Apparel wool requires absence or a low percentage of medullated fibre, whereas for carpet-type a certain percentage of heterotypes (with interrupted medulla) is desirable. Kemp (shed medullated fibre) is undesirable for both categories.
Other important quality traits are length (not falling below a minimum) strength (showing no tenderness, or point of reduced diameter) and pigmentation. White wool is preferred for both categories on the world market, but for some local markets pigmented wool is desirable.
Table 2 shows that the heritability levels for wool weight and quality traits are high. (Pigment of the whole or part of the fleece is genetically controlled and can be manipulated, though the presence of individual pigmented fibres in a white fleece is causing concern, and the inheritance of these is under investigation in Australia). Long-term selection for wool weight, with and without control of quality, has been reported from several experiments in Australia (Turner 1977), with gains of 2% per annum when quality was controlled through diameter. Attempts to control quality by maintaining high crimp frequency leads to lower rates of increase in wool weight, because of the high negative genetic correlation between the two.
Goats
The quality characteristics for mohair and cashmere are similar to those for wool. Although no selection experiments have been reported, Shelton (1979) concluded that selection for fleece weight had been effective as average weight per head in Texas had increased from 1.4kg. in 1910 to 3.4kg. in 1975 (= 0.03 kg/year).
Work on increasing cashmere production is in progress in Australia (Restall, personal communication), India (Bhat in SABRAO 1980) and China (Cheng 1985).
Milk Production
Milk production is measurable only in the female, and also requires more frequent recording than other characteristics considered. For this reason genetic progress will be made mainly through distribution of males (or semen) from a central necleus in which measurement and recording are possible, and in association with which progeny - testing of males can be done.
Sheep
One of the countries best-organized for achieving genetic progress in milk production from ewes is France. Flamant (1970), Flamant and Barillet (1982) and Flamant et al. (1982) have reviewed the work; miIk-recording now involves over 80,000 ewes served by progeny - tested rams, and over 130,000 subject to a simplified recording scheme and within-flock mass selection.
Goats
Both Norway and France have reported herd recording schemes for goats. The Norwegian records show an increase in lactation milk yield from 574 kg. in 1967 to 592 kg. in 1978, the correspondingly fat percentages being 3.4% and 3.2%. The highest record is given as 1960 kg.
Steine (1976, 1980) gave estimates of genetic parameters and described the Norwegian recording and progeny testing system; he predicted annual gains of 1.0 – 1.5% in milk yield using natural mating, with the Norwegian system of “buck circles”. He considered that AI for goats was at that time not suitable for wide use.
Using genetic parameters estimated in Norway, Ronningen (1967, 1980) made theoretical predictions of possible increases under Swedish conditions of 1.6% (11 kg. milk) per annum with natural mating, or 1.9–2.8% (13–19kg.) using AI.
Bouillon and Ricordeau (1975) made estimates of direct and indirect responses to selection for milk production in goats in France. Selection for milk yield would increase yields of milk and milk protein and fat, but would decrease percentages of protein or fat. If cheese-making were the aim, they suggested selection for yields of protein or fat.
Singh and Acharya (1980) suggested that selection on first lactation yield would give large responses in lifetime performance for the Beetal goat in India.
Correlations between Characteristics
Turner (1972) reviewed estimates of genetic correlations between sheep production traits, and concluded there are no important genetic antagonisms, provided wool quality is based on average fibre diameter and not,.as formerly, on crimp frequency. A high negative genetic correlation between wool weight and crimp number/unit length of staple will hinder genetic gains in wool weight if there is simultaneous selection to maintain high crimp frequency. The genetic correlation between wool weight and fibre diameter, though positive, is low, and diameter can be kept from increasing without serious loss to gains in wool weight (Turner et al. 1968).
Some negative estimates of the genetic correlation between wool weight and reproduction rate led to concern at one time, but other estimates have been negligible, and reciprocal selections for wool weight and reproduction rate in the Australian Merino have led to no changes in the other character. (Turner et al. 1972 and Turner, unpublished)
Estimates of genetic correlations for goats are not as numerous as for sheep.
Ways of increasing selection response Establishing a Nucleus
The previous discussions have related to selection within one flock. Greater genetic progress can be made if a large population can be screened and the top animals placed in a central sire-breeding nucleus. Suppose 4 percent of rams and ewes can be drawn off in this way, taking the most superior animals from a number of flocks or herds; the average standardized selection differential of the parents is then 2.154. With a heritability of 0.4 and a standard deviation of 12%, the offspring of these parents would on average be 2.154 × 0.4 × 12 = 10% superior to the average of the population from which they came. If males from this necleus are then distributed in the original population, they will give an initial production boost due to the superiority of the nucleus. Thereafter genetic progress in the main population due to selection will be at the same rate as in the nucleus. Apart from its initial advantage of including the most superior animals, a necleus has the additional advantage of being a centre in which new techniques can be concentrated.
Increasing the selection differential
The importance of a high selection differential has already been stressed, together with two avenues of increase - raising reproduction rate and using artificial insemination. Recent techniques involving multiple ovulation and embryo transfer (MOET) make it possible to increase the selection differential for females as well as males. Such techniques require a high level of facilities and technical skills; they could be applied in a central nucleus, but costs would need to be balanced against advantages, and the dangers of inbreeding would need to be watched.
Decreasing the generation interval
Reduction of average parent age from 4 years to 2 would double ΔGa, while reduction of 4 to 1 would quadruple it.
Average parent age depends on age at first parturition for both sexes, and for females on reproduction rate as well; for example, if only 25 females reached mating age per 100 mated, 5 age-groups of dams would be required to allow for annual death rates, compared with only 3 if the reproduction rate were high enough to lead to 40 replacements per 100 females mated.
Both age at first parturition and reproduction rate depend on species, breed and environment, and both can be changed by selection. Multiple ovulation techniques can be used to increase the reproduction rate; combined with embryo transfer they can be used to ensure that replacements come from young dams, so providing another avenue for decreasing generation interval. As previously stated, the techniques are not yet likely to widely applicable.
CONCLUSIONS
Selection of species and selection of breeds or strains within species can both increase efficiency of production.
Selection of individuals within a flock has led to genetic gains of 1.5 – 2.0% per annum in the character under selection. These may at first sight seem small increases, but they are permanent and cumulative. The total genetic gain after 10 years would be 10ΔGa, and the total extra product over the 10 year period would be -
Total gain = ΔGa+ 2 ΔGa+ -- +10ΔGa
= 55 ΔGa
In other words, a flock with an initial wool weight of 4 kg. of wool, with a ΔGa of 2% (=.08 kg.) would after 10 years return an extra amount of 55 X0.08 = 4.4kg. per head.
These are minimum rates of gain; various techniques exist which can increase them.
ACKNOWLEDGEMENTS
The staff of SCIRO's Division of Molecular Biology in Sydney have graciously assisted with the use of processing equipment, and Mrs. Pat Tregenza and Mrs. Margaret Dowell expertly turned an untidy mess into a professional typescript.
REFERENCES
Acharya, R. M. 1982. Sheep and goat breeds of India. FAO Animal Production and Health Paper No. 30.
Atkins, K. D. 1980. Aust. J. Exp. Agric. Anim. Husb. 20-: 272–9, 280–7.
Australia, Standing Committee on Agriculture. 1982. Goats for meat and fibre in Australia. Report of the Expert Panel appointed by the Animal Production Committee, Canberra.
Bernard, C. S. and Fahmy, M. H. 1974. Canadian Journal of Animal Science 54: 721.
Bindon, B. M., Piper, L. R. and Ch'ang, T. S. 1985. In Reproduction in sheep, pp 243–6, Australian Academy of Science. Lindsay and Pearce Eds.
Bouillon, J. and Ricordeau, G. 1975. Journees de la Recherche Ovine et Caprine 1: 124–32
Bradford, G. E. and Spurlock, G. M. 1972. J. Anim. Sci. 34: 737–45.
Brooke, C. H. and Ryder, M. L. 1978. Declining breeds of Mediterranean sheep. FAO Animal Production and Health Paper No. 8.
Brown, Lynn Elizabeth and Johnson, W. L. 1984. Int. Goat Sheep Res. 2: 212–26.
Cheng, P. L. 1985. Livestock breeds in China. FAO Animal Production and Health Division and China Academic Publishers (In Press).
Clarke, J. N. 1975. N.Z. Ministry of Agriculture and Fisheries Annual Report, Research Division 1974/5.
Devendra, C. 1978. Anim. Prod. 4:9–22.
Devendra, C. 1983. Goats : Husbandry and potential in Malaysia. Ministry of Agriculture, Kuala Lumpur, Malaysia.
Dun, R. B., Alexander, R. and Smith, M. D. 1966. Proc. Aust. Soc. Anim. Prod. 6: 66–8.
Dunlop, A. A. 1962/1963. Aust. J. Agric. Res. 13:503–31 and 14:690–703. el Hag, M. G., el Hag, G. A. and Gaali, E. 1984. International Goat and Sheep Journal 2: 274–82.
FAO. 1985. Proceedings of Expert Consultation on Animal Genetic Resources (In Press).
Farid, A., Makarechian, M. and Sefidbakht, N. 1977. J. Anim. Sci. 44: 542–8.
Flamant, J. C. 1970. Bulletin Technical Department Genetique, Animal Institut National Recherche Agronomique No. 10: 3–7, 29–34, 35–41.
Flamant, J. C. and Barillet, F. 1982. Livest. Prod. Sci. 9: 549–59.
Flamant, J. C. and Morand-Fehr, P. 1982. In Sheep and Goat Production. Elsevier Scientific Publishing Co. (Coop ed.).
Fogarty, N. M., Dickerson, G. E. and Young, L. D. 1984, 1985. J. Anim. Sci. 58: 285–311 and 60: 41–57.
Garran, J. and White, L. 1985. Merinos, Myths and Macarthurs. Pergamon Press, Sydney.
Gjedrem, T. 1969. Acta Aqric. Scand. 19:103–115.
Goode, L., Linnerud, A. C. and Tugman, D. F. 1980. Internat. Goat and Sheep Res. 1: 216–25.
Hanrahan, J. P. 1982a. Animal Production. Report from Dunsinea, Moorepark and Western Research Centres, Dublin, Irish Republic.
Hanrahan, J. P. 1982b. Animal Production. Report from Dunsinea, Moorepark and Western Research Centres. Research Report 1981, Dublin, Irish Republic.
Harrington, G. N. 1982. In Goats for meat and fibre. Report of the Expert Panel appointed by the Animal Production Committee of the Standing Committee on Agriculture, Canberra.
Hoist, P. J. and Pym, R. A. 1977. In Proceedings of Symposium on Goats for Fibre and Meat Production, University of Sydney.
Hunter, G. L. 1968. Anim. Breed. Abstr. 36: 347–78.
Lawson, Janet and Shelton, M. 1982. Internat. Goat and Sheep Res. 2: 30–7.
Lax, J., French, L. R., Chapman, A. B., Pope, A. L. and Casida, L. E. 1979. J. Anim. Sci. 49: 939–42.
Lindhal, I. L. and Terrill, C. E. 1975. J. Anim. Sci. 41: 251 (Abstr.).
McGuirk, B. J., Killeen, I. D., Piper, L. R., Bindon, B. M., Wilson, R., Caffery, G. and Langford, C. 1984. Proc .Aust. Soc. Anim. Prod. 15:464–6.
Mason, I. L. 1967. The Sheep Breeds of the Mediterranean. FAO, Rome and Commonwealth Agricultural Bureaux, Farnham Royal, England.
Mason, I. L. 1980. Prolific tropical sheep. FAO Animal Production and Health Paper No. 17.
Nicholas, F. 1985. Mendelian inheritance in animals - Bibliography (To be published).
Notter, D. R. and Copenhaver, J. S. 1980. J. Anim. Scj. 51: 1033–42 and 1043–50.
Obst, J. M., Chaniago, T. and Boyes, T. 1980. Survey of sheep and goats slaughtered at Bogor, West Java, Indonesia. Centre for Animal Research and Development, Indonesian Centre Report No. 10.
Owen, J. B., Brown, K. and Flint, R. 1980. 31st Annual Meeting, European Association for Animal Production, No. Gl-9. 6pp.
Pakistan Agricultural Research Council (PARC - 1982) - Booklet prepared for International Sheep and Wool Seminar, Islamabad.
Pattie, W. A. 1965. Aust.J. Exp. Agric. Anim. Husb. 5: 353–60.
Peacock, Christie 1982. Proceedings of a Seminar on Small Ruminant Productivity in Africa, ILCA, Addis Ababa, pp 77–80.
Piper, L. R. and Bindon, B. M. 1981. Proceedings of World Conference on Sheep and Cattle Breeding, N.Z.
Piper, L. R. and Bindon, B. M. 1982. In Proceedings of a Workshop on the Booroola, Armidale, N.S.W., PP 9–19.
Piper, L. R., Bindon, B. M. and Davis, G. H. 1985. In Genetics of Reproduction in Sheep. Butterworth - Land and Robinson Eds.
Rae, A. L. 1982. In Sheep and Goat Production. Elsevier Scientific Publishing Co. (Coop Ed.)
Restall, B. S. and Pattie, W. A. 1985. Paper read to 5th Conference of Australian Association of Animal Breeding and Genetics, Sydney, August 26–28.
Ronningen, K. 1967. A Study of genetic parameters for milk characteristics in goats. Report 232, University of Norway.
Ronningen, K, 1980. Internat. Goat and Sheep Res. 1: 18–40.
Rzepecki, R. 1979. Symposium on intensive sheep production, Helsinki, pp 83–5.
Shelton, M. 1979. The Angora industry in U.S.A. Proceedings of 2nd National Goatbreeders Conference, Perth, pp 23–31.
Shelton, M. and Bassett, J. M. 1970. Texas. A. and M. University Sheep and Angora Goat, Wool and Mohair Research Report, PR 2750, pp 38–41.
Singh, R. N. and Acharya, R. M. 1980. Internat. Goat Sheep Res. 1: 226–33.
Society for the Advancement of Breeding Researches in Asia and Oceania (SABRAO). Proceedings of Workshops on Animal Genetic Resources. Rokyo (1980 - Barker and Turner Eds.) and Kuala Lumpur (1982 - Mukherjee Ed.)
Steine, T. A. 1976. Meldinger fra Norges Landbrukshogskole 55 (4):19pp,
Steine, T. A. 1980. Internat. Goat Sheep Res. 1: 108–112.
Tallis, G. M., Turner, Helen Newton and Brown, G. W. 1964. Aust. J. EXP. Agric. 15:446–52.
Thimonier, J., Cognie, Y., Cornu, C, Schneberger, J. and Vernusse, G. 1975. Ann. Biol. Anim. Biochem. Biophys. 15(2): 365–7.
Turner, Helen Newton 1972. Anim. Breed. Abstr. 40: 621–34.
Turner, Helen Newton 1977. Anim. Breed. Abstr. 45: 9–31.
Turner, Helen Newton 1978. Aust. J. Agric. Res. 29: 327–50
Turner, Helen Newton 1980. In Proceedings of a Workshop on the Booroola, Armidale N.S.W., pp 1–8.
Turner, Helen Newton 1982. In Goats for meat and fibre. Report of Expert Panel appointed by the Animal Production Committee of the Standing Committee on Agriculture, Canberra.
Turner, Helen Newton, Brooker, M. G. and Dolling, C. H. S. 1970. Aust. J. Agric. Res. 21:955–84.
Turner, Helen Newton, McKay, Elaine and Guinane, Fay 1972. Aust. J. Agric. Res. 23: 131–48.
Turner, Helen Newton and Young, S. S. Y. 1969. Quantitative Genetics in Sheep Breeding. Macmillan Co. of Australia, Melbourne.
Wahid, A. 1982. Proceedings of International Seminar on Sheep and Wool, Islamabad (Pakistan Agricultural Research Council).
Wallace, L. R. 1958. N. Z. J. Agric. 100: 417–24.
Wilson, A. D. 1977. Aust. J. Agric. Res. 28: 501–8.
Wilson, R. T. 1984. Internat. Goat Sheep J. 2: 243–51.
Yalcin, B. C. 1979. The Sheep Breeds of Afghanistan, Iran and Turkey, FAO, Rome.
TABLE 1. Average selection differentials for different ratios males : females
| Male : female ratio | No. males required | Proportion saved (males) | Standardized Selection differential (males) | Average selection differential | Percent change over 1 : 50 |
| 1 : 25 | 40 | 0.08 | 1.858 | 1.104 | - 12 |
| 1 : 50 | 20 | 0.04 | 2.154 | 1.252 | 0 |
| 1 : 100 | 10 | 0.02 | 2.421 | 1.386 | + 11 |
| 1 : 500 | 2 | 0.004 | 2.962 | 1.656 | + 32 |
| 1 : 1000 | 1 | 0.002 | 3.170 | 1.760 | + 41 |
| Assumptions: | No. of females to be mated | 1,000 |
| Males available for selection | 500 | |
| Proportion females saved | 0.80 | |
| Standardized selection differential for females | 0.350 |
TABLE 2. Heritability estimates
| Characteristic | Species | Range of estimates | |
| Number of offspring/parturition: | |||
| Born (other than 1st parturition) | Sheep | 0.1 – 0.4 | |
| Goat | 0.1 – 0.2 | ||
| Weaned ( " " " ") | Sheep | 0.1 – 0.2 | |
| Kidding interval | Goat | 0.2 | |
| Weaning weight | Sheep | 0.2 – 0.3 | |
| Goat | 0.3 – 0.5 | ||
| Weight of lamb weaned/ewe exposed 12–16 month liveweight | Sheep | 0 – 0.2 | |
| Sheep | 0.4 – 0.6 | ||
| Goat | 0.5 | ||
| Annual fibre weight (clean or greasy) | Sheep | -Wool | 0.3 – 0.5 |
| Goat | -Mohair | 0.2 – 0.4 | |
| -Cashmere | 0.6 | ||
| Average fibre diameter | Sheep | -Wool | 0.4 – 0.6 |
| Goat | -Mohair | <0.2 | |
| -Cashmere | 0.5 | ||
| Percent medullation | Sheep | -Wool | 0,4 – 0.6 |
| Kemp score | Goat | -Mohair | >0.3 |
| Staple length | Sheep | -Wool | 0.3 – 0.6 |
| Goat | -Mohair | >0.3 | |
| -Cashmere | 0.7 | ||
| Milk yield per lactation | Sheep | 0.2 –- 0.6 | |
| Goat | 0.3 – 0.7 | ||
| Milk - Fat % | Sheep | 0.2 – 0,6 | |
| Goat | 0.3 – 0.5 | ||
Sources: Sheep: Gjedrem (1969), Turner (1977), Rae (1982).
Goats: Shelton and Bassett (1970), Steine (1976), Rae (1982) Restall and Pattie (1985).
![]()
![]()
![]()