![]()
![]()
![]()
by
V.S. Kirpichnikov
Scientific Research Institute on Lake and River Fisheries
Leningrad, U.S.S.R.
Modern methods of animal selection are inseparably connected with recent achievements in the fields of quantitative and population genetics. Fish selection is based on the planned utilization of genetic variation inherent in all living creatures. An understanding of the nature of genetic variation and the laws by which it is governed is indispensable for the formulation of an effective programme of selection work.
In comparison to the breeding of domestic animals and agricultural plants, fish culture is a young science. Though in some eastern Asiatic countries, particularly in China, fish culture has been practised for more than two thousand years, until quite recently, their method of culture was based entirely on culture of young fish caught in natural waters. The domestication of fish and the creation of breeds differing from their wild parents in higher productive properties was actually started only a few centuries ago. The only exception is the decorative goldfish (Carassius auratus); some goldfish varieties have a thousand-year-long history.
The number of domesticated fish species is not great. Besides carp and goldfish, they include Crucian carp (C.auratus gibelio), tench (Tinca tinca), rainbow trout (Salmo gairdneri), lake trout (Salvelinus namaycush), char (S.alpinus), gourami (Osphronemus goramy), several centrarchids, tilapia and several other fish which are bred in some countries of southern and southeastern Asia. Recent years have seen a sharp increase in the domestication of these. This has been brought about by a swift rise in importance of pond fish for the total balancing of fish production. Attempts to reproduce many new species of various fresh water families in ponds and tanks have proved successful. Among these the most important are:
silver carp (Hypophthalmichthys molitrix), feeding on phytoplankton;
grass carp (Ctenopharyngodon idella), feeding on higher aquatic vegetation;
bighead (Aristichthys nobilis), a zooplankton feeder;
black carp (Mylopharyngodon piceus), feeding on benthos, mainly molluscs;
cisco (Coregonus albula), feeding mainly on zooplankton;
peled (Coregonus peled), feeding mainly on zooplankton;
catfish (Ictalurus punctatus, feeding on benthos and is also predatory;
pikeperch (Stizostedion lucioperca), a predator.
buffalo fish (Ictiobus, spp.), feeding on benthos
Domestication of these fish requires selection to increase their productive properties and adaptation to new conditions of existence greatly differing from natural environments.
American and Soviet fishery biologists claim that selection is applicable, not only to domesticated forms but also to natural populations (Riggs and Sneed, 1959; Nikolsky, 1966). When man is able to fully or partially control reproduction in fish inhabiting natural waters, selection becomes possible. An interesting example is the following: at one of the fish cultural farms in Alaska, U.S.A., all chinooks (Oncorhynchus tschawytscha) ascending the river for spawning were caught with the help of traps and the best of them were selected for breeding. The young fish were then liberated so that they could grow under natural conditions and subsequently descend into the sea (Donaldson and Menasveta, 1961). The homing instinct driving the salmon back for spawning into the river from which they descended precluded the selected stock from mixing with other stocks.
Among the principal aims to be accomplished by fish selection are the following (Kirpichnikov, 1966a):
To increase the growth rate, by better utilization of food (physiological decrease of food expenditure per unit of growth increment);
To increase the growth rate, by fuller utilization of natural food in ponds and higher consumption of feed mixtures;
To increase resistance to oxygen deficiency, to high or low temperature, to higher salinity or to other deviations from the normal environmental conditions;
To improve resistance to infectious diseases and to infestation with parasites (to develop new breeds resistant to particular diseases);
To improve the nutritive properties of fish (to increase the calorie content, to decrease the proportional weight of non-edible parts, to decrease the bone content, to increase or decrease the fat content, etc.)
Sometimes it is important to accomplish some other aims. Thus, the aim of carp selection in northern areas is to speed up sexual maturation and to develop the ability to reproduce at relatively low temperatures. In tropical and sub-tropical areas, selection is aimed at slowing down maturation, since early maturation is related to the switching over of the metabolism to the development of sex products and may result in uncontrollable reproduction in ponds. There are many more examples of this kind. Particular attention is to be given to selection for combining ability. As in other branches of animal and plant breeding the utilization of heterosis will become more and more important. The selection of pairs showing maximum heterosis at crossing has acquired a decisive significance in the breeding of carp, tilapia and some American pond fish.
Two more points should be emphasized. In a number of countries there is shortage of water supply as well as of areas suitable for construction of ponds. It is beyond doubt that in the near future fish culture will develop along the lines of growing fish in cages, tanks or aquaria, so that the water running through them can be purified and used many times. Thus fish selectionists are confronted with additional tasks of developing breeds adapted to conditions of existence in small reservoirs at high densities and to feeding on special food mixtures.
The advance of fish culture to countries with a hard, cold climate requires an improvement in the cold-resistance properties of pond fish. Adaptations to the ambient temperature may be referred to the group of the most conservative specific characters.
According to Ushakov (1963), a change in the tempature adaptation is possible if there is a simultaneous change in the level of cold or heat resistance of the great number of various cell proteins. Therefore, the development of breeds having a higher cold resistance (in particular possessing the ability to grow at relatively low temperatures) is one of the most difficult tasks of fish selection.
In mass selection the suitability of each individual is determined by the phenotype, while the genotypic characteristics remain unknown. Response in mass selection (R) is determined by the general equation (Falconer, 1960):
| R=i·б·h2 = S·h2 | (1) |
where S = selective differential (the difference in a certain trait between the individuals selected and the population as a whole);
h2 = heritability of differences (the share of additive genetic variation in the general variation of the character);
i = intensity of selection.
Due to high fecundity of fish, the values of S and i may be much higher than in the selection of domestic animals and poultry. Thus for cattle, intensity of selection is rarely as high as 1, whereas for carp a value as high as i=3 is often obtained. A similar high value of i is sometimes obtained in rainbow trout (Savostjanova, 1969).
In performing mass selection in fish breeding, we usually apply the selection severity coefficient or the rejection rigidity factor (V) which is calculated by applying the equation
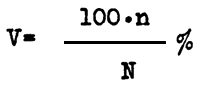 | (2) |
where n is the number of individuals selected, and N is the total number of fish grown. By plotting the intensity of rejection against its severity on a semilogarithmic scale, we obtain a curve which is shown in Fig.1. It is obvious that with a decrease in the severity coefficient within the range of 100 to 10 percent there is a sharp increase in the intensity of selection. A further decrease in V (down to 1 percent results in a considerably lower increase in i; and with a decrease in V down to 0.1 - 0.01, the intensity of selection hardly increases at all. Practically, for fish with high fecundity, selection gives the best results when the severity of selection ranges from 1 percent (1:100) to 0.1 percent (1:1000). A further increase of rejection will produce such a low increase in the response to selection that it becomes hardly practicable.
Response to selection is directly proportional to the heritability of the character (h2). In a number of cases this permits a rather accurate estimate of the value of heritability of the character under selection by applying the equation
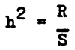 | (3) |
To obtain such an estimate, selection should be conducted in several successive generations; for in the first generation, the results of selection may be distorted by dominance and epistasis. Heritability determined by the results of selection is called realized heritability. Some investigators believe that this method of estimating heritability is the best when working with pond fish (Moav and Wohlfarth, 1967).
Wlodek (1968) recommends that biometric constants should be used in carp selection for weight. Apart from the mean value, the standard deviation by weight is calculated for each group of fish at all ages. Individuals showing a stable deviation in the course of several years (at the age of 0+, 1+, 2+, 3+ etc.) of a maximum value (2 σ or even 3 σ) should be selected for breeding.
Stegman (1968) introduced the concept of “relative growth coefficient” which is calculated by applying the equation
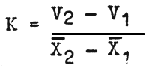 | (4) |
Fig.1 Relation between severity (v) and intensity (i) of selection. | 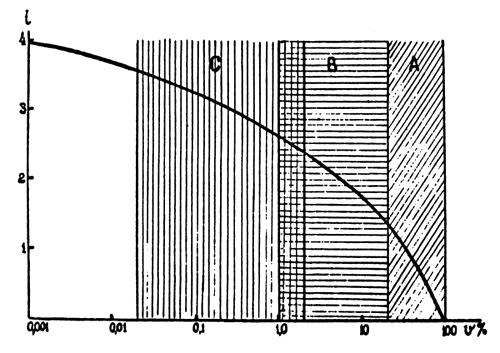 |
A - cattle-breeding B - poultry-breeding C - fish culture | |
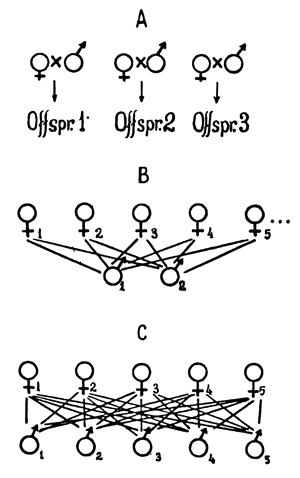 | Fig.2 Progeny testing in fish breeding. |
A - comparison of pairs B - testing of fish of one sex C - diallele crossing |
where V2 and V1 are the final and the initial weights of the given specimen and x2 and x1 are the mean final and initial weights of all fish in the pond. Wlodek and Stegman recommend that fish which have the growth coefficient well over 1 in the course of several years should be maintained for breeding.
In applying selection methods, the fact should not be overlooked that in all cases of selection for growth rate, high intensity of selection is of great importance. This can only be realized when a sufficiently large number of fish are grown. Difficulties involved in individual marking of thousands and tens of thousands of fish limit the applications of these methods on a wide scale.
According to equation (1), selection response depends on three values: i, б and h2. An increase in any of them may bring better results in selection.
Intensity of selection (i): The value of i may be raised to 3 or 4 in highly fecund fish by increasing the number of individuals grown, and through this, increasing the severity of selection. However, it should be noted that very rigid norms of rejection not only result in an increase in the expenditure incurred in the growing of fish but may involve other undesirable effects:
the weight variation curve is often asymmetric (particularly in the first year of life). Its right part becomes longer due to a small number of jumpers (champions), which may not be genetically superior but jump forward only due to a chance (purely paratypic) excess in weight over fish of the same age at the beginning. This weight advantage then increases under conditions of food competition (Nakamura and Kasahara, 1955, 1956, 1957; Moav and Wohlfarth, 1963, 1967; Krjageva, 1966). Heritability of weight differences between the champions and other fish proves to be very poor, and the selection of champions for breeding does not bring good results.
Equally undesirable is the so-called correlated response to selection - an often deleterious change in some characters correlated to the character subjected to selection.
One must be very careful in increasing the intensity of selection and pay due regard to a possible decrease in the heritability of the character subjected to selection and to the possibility of the occurrence of indirect deleterious effects due to very rigid selection.
Variability, expressed by the standard deviation б must be sufficiently high. Only genetic variation is important in mass selection. An increase in the value due to an increase in environmental variation is useless for it results in a proportional decrease in heritability. Equally useless is an increase in the share of the phenotypic variance which is conditioned by the non-additive genetic variation.
Heritability (h2) is an important component of the selection response equation. In carrying out selection, all possible methods of increasing heritability should be used.
Heritability is understood to be the relation between the genetic (additive) variance and the general (phenotypic) variance:
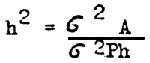 | (5) |
Sometimes this heritability is called heritability in the narrow sense of the word as distinguished from the heritability in the broad sense involving the whole genetic variation irrespective of its nature.
There are two ways of increasing h2:
Genetic (additive) variation increases when non-related individuals are crossed. Inbreeding results in a higher homozygosity of the population, whereas outbreeding increases the degree of heterozygosity, i.e. increases genetic variation.
Inbreeding is unavoidable when the number of spawners is low (20–40 individuals) and when the progeny obtained from one or two crossings is maintained for breeding. Such a situation easily occurs in the selection of highly fecund fish such as carp. In carp breeding every step must be taken to avoid a sharp drop in the abundance of the stock; it must consist of at least 50 to 100 spawners. Measures should be taken to have annually a sufficient number of fish for crossings and to make possible the selection of fish from different crossings for breeding.
Particularly dangerous is the so-called heterozygous balance in a population. In this case, the best specimens maintained for breeding appear to be the most heterozygous due to advantages typical of heterozygotes (the so-called overdominance). Genetic variation ceases to be additive, and heritability sometimes drops to a very low value. There are reasons to believe that a considerably high heterozygous balance occurs in carp taken for body weight character, which makes positive selection not responsive (Kirpichnikov, 1958b; Moav and Wohlfarth, 1967). It can be destroyed, and the share of additive genetic variation (б A2) increased only by means of sufficiently remote crossings.
A new method of increasing genetic variation, which is not yet applied on a large scale, consists in speeding up the process of mutation by means of irradiation and chemical mutagens. Experiments of this kind were started by R.M.Tzoy in 1967, and preliminary results seem to be promising (personal communication).
Paratypic (non-hereditary) variation may be depressed provided certain conditions are observed, such as:
growing of spawners under conditions favourable for the maturation of sex products;
simultaneous crossings;
incubation of eggs under conditions as identical as possible temperature, oxygen regime, illumination, water flow, etc. must be identical for all the eggs;
growing of larvae and later of fry, fingerlings and older fish in water bodies rich in food where food competition may not play an important part. It should be remembered that conditions under which fish are kept should not differ to a great extent from normal conditions; for selection may appear to be effective under certain conditions but give poor results under a different set of conditions;
simultaneous transference of fish from one pond to another without mixing fish grown in different ponds;
selection mainly at the age which is subject to improvement by selection. Experiments show (Kirpichnikov, 1959; Stegman, 1965 a,b) that in carp the covariance of weight in the first, second and third years of life is not very high. The champions of the first year may not appear to be the best in the second year and vice versa.
It is not easy to grow a great number of fish of older ages for subsequent selection. A way out is to carry out selection in two or three stages - with low severity in the first year (10–20 percent) and more rigidly later on, so that only 5–10 percent of the number of fingerlings selected are maintained for breeding. Thus the summary severity of selection will be sufficient (0.5–1.0 percent or 1:200–1:100).
The observance of the conditions mentioned above will result in a considerable reduction of environmental variance and an increase of heritability. It should be added, however, that different fish may require different methods of reducing paratypic variation.
A long-lasting, one-way selection for a certain character often results in a change in other morphogenetically or genetically correlated characters. Indirect changes of this kind often appear to be deleterious and result in a reduction of viability and growth rate. This accounts for the fact that many highly productive animal breeds are too delicate. In nature the only criterion of selection is the ability of survive and to produce viable progeny (fitness). In artificial selection, the suitability is determined by the productive properties in which the selectionist is interested; but which, however, are far from being always positively correlated to fitness.
European cultured carp has undergone selection in the course of several centuries mainly for growth rate, exterior characters and scale pattern. Selection has resulted in the deterioration of a number of physiological and biochemical factors, including the blood factor (Steffens, 1964). Here we undoubtedly have correlated changes.
Recently an experiment was conducted on the selection of carp for height/length ratio. It appeared that the progeny derived from the highest carp was inferior to other progenies in growth rate (Moav and Wohlfarth, 1967).
A study of the Ropsha carp showed that variations in vertebrae number were related to variations in oxygen consumption by fingerlings (Tzoy, 1968). There is undoubtedly a correlation here, which means that selection for one of these characters will result in changes in the other.
An example of correlated response which is very important for the theory of selection is provided by changes in size of gonads and in rate of maturation. This relationship has been established for carp and rainbow trout. According to our observations, the largest carp often show a slow maturation of sex glands and sometimes a marked decrease in the number of eggs or spermatozoa. Severe selection for growth rate may therefore have an adverse effect on fecundity. On the other hand, according to Savostjanova (1969), trout which are superior in growth rate to other trout of the same age show an increase in fecundity; at the same time some undesirable effects of high growth rate occur, such as fatty degeneration of the liver, which seems to be a result of increased artificial feeding (Factorovich, 1969). Selection of large specimens under conditions of severe food competition observed at high stocking densities is accompanied by another correlated change, intensification of aggressiveness. In competition for food, the victory is often gained by those fish which are more active in search of food and which snatch it from their neighbours. If differences in growth rate are mainly determined by this factor, then the effect of selection for weight will be very poor.
Examples of correlated responses in selection of non-selected characters in fish are numerous. This phenomenon should not be overlooked in conducting mass selection, for it may reduce to nought all attempts to improve productive properties of a breed by selection.
There are two aspects of genotypic selection, family selection and progeny testing, both of which are applicable in fish culture.
In applying family selection, several fish families are grown under identical conditions. By comparing the properties of these families it is possible to determine which of them are the best and to maintain these for breeding. In order to obtain a separate progeny (one family) we can use either one male and one female or a small group of spawners.
Family selection may be very effective. The response equation remains essentially the same as in mass selection:
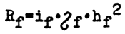 | (6) |
but the magnitude of all terms in the right side of the equation is different. Intensity of selection appears to be lower than in mass selection since it is not possible to grow a great number of families. A reduction is also observed in the standard deviation, which in this case reflects variation in the family means rather than individual variation. On the other hand, the heritability factor becomes much higher, and when the conditions under which all families are kept are equalized it approaches unity.
If it is not necessary to cut the fish open in order to be able to examine it, any specimen may be maintained for reproduction. When it is necessary to cut the fish open or when it is impossible to avoid damaging the fish in examining it, the brothers and sisters of the specimens examined from the best families are maintained for breeding. This is called sib-selection.
Family selection methods in fish culture were considered by Kirpichnikov (1966b). They are as follows:
Crossings. Several pairs or groups of spawners are used to obtain a progeny with the help of artificial fertilization or natural spawning. The number of crossings may be 8–10; and when many aquaria and ponds are available, it may be as high as 15 or 20. All crossings are performed simultaneously and the same methods of culture are applied.
Incubation of eggs. When eggs are placed into a Zuger or a Weiss apparatus or into a crystallizer 1 incubation conditions - temperature, water flow, oxygen content, illumination, density and pattern of distribution of eggs - are equalized. The same requirements apply to development of eggs in ponds, but in this case there is an additional requirement of simultaneous spawning in all ponds.
Larvae. Larvae derived from artificial fertilization are kept in aquaria, cages or ponds at equal densities and under conditions as similar as possible.
Larvae derived from natural spawning are transferred to other ponds not later than 3 or 4 days after hatching.
Communal growing of fish belonging to different families with the aim of testing them. On reaching the size when they can be marked, larvae can be stocked in communal ponds or tanks for further growing. To decrease competition between separate individuals and to eliminate much of the dependence on stocking weights, the following conditions should be observed:
stocking weights (family averages) are equalized as far as possible;
fish belonging to different families are stocked simultaneously;
dense stockings and starvation should be avoided;
growing experiments are replicated three or four times;
at least 50 fish from each crossing are used for each experiment (pond).
Separate growing of fish belonging to different families for testing them. When many identical ponds, tanks or aquaria are available or when it is possible to arrange net enclosures of equal size in a pond, each family is grown separately. The testing of 10 families would require not less than 30 ponds or tanks (when each experiment is replicated in three ponds).
In addition to conditions mentioned above, it should be remembered that when separate growing is applied, the reliability of results depends on the degree to which it is possible to obviate the differences between ponds or between tanks. The best results were obtained when ponds were partitioned into equal parts with the help of capron netting (Moav and Wohlfarth, 1968 b).
The existence of the so-called maternal effect (correlation between egg size, survival of larvae and their growth rate) and paternal effect (correlation between the weight of males and the quality of sperm) makes it necessary to grow fish in testing experiments until they reach the weight of 20 to 30 g, and in large fish such as carp - to the weight of 400 to 500 g (Kirpichnikov, 1966a).
The principal disadvantage of family selection lies in technical difficulties involved in the simultaneous growing of many families.
There are three ways of progeny testing in fish culture. The first method is testing of pairs without testing males and females separately (Fig.2A). The second is to test spawners belonging to one sex (Fig.2B shows the scheme of testing females); and the third (complete diallele crossing) is the testing of both females and males (Fig.20). All three methods have been applied in practice.
Pair testing is widely used in carp selection work in Israel (Wohlfarth, Moav and Lahman, 1961; Moav and Wohlfarth, 1963, 1968a). Progenies of several pairs (up to 15 at a time) are grown together and the combinations are replicated as many as 10 times. The analysis of the results obtained allows the following conclusions to be made:
The data on final weights are to be corrected in accordance with the differences in the initial weights (at stocking). For growing fish weighing 400 to 600 g the correction factor is 3–4.
Comparative experiments on growing fish of the above weight permit an accurate estimation of the worth of pairs and the selection of the best among them. Results obtained from ponds located in different areas are in good accord.
Experiments show that pairs from which the best progenies were derived maintain their superiority when grown separately.
Testing of spawners of either sex was performed repeatedly in carp selection in the U.S.S.R. (Kirpichnikov, 1958b, 1961, 1966b; Kusema, 1961, 1962; Polyksenov, 1962). Artificial fertilization was mainly used. The experiments revealed a number of limitations, mainly of a technical nature (the necessity of having many ponds, distortion of results due to competition, difficulties involved in marking and in equalizing initial weights, etc.). If all the conditions are observed, which are mentioned in the section dealing with family selection, this method will permit the evaluation of the best and the worst spawners. Two or three males are necessary for the evaluation of females and, correspondingly, two or three females are sufficient for the evaluation of males. Experience shows that the number of spawners tested should not exceed 10 or 12, for a greater number of fish would result in a less accurate evaluation. Improvements in the technique of growing fish in aquaria may possibly increase the number of individuals tested to 15–20.
An experiment on diallele crossing of five female and five male grass carp was first conducted by Slutsky in 1967 (personal communication). Twenty five progenies were derived; the investigation is restricted to comparing progenies at embryonic and larval stages of development.
The advantage of progeny testing lies in the fact that it permits the evaluation of separate spawners (or pairs of spawners) and the selection of the best of them for further selection work. Since many fishes are highly fecund, such selection may be of great significance. On the other hand, it should be noted that the great disadvantage of this method is a reduction in the rate of selection. In carp breeding, progeny testing requires one or two years (a considerable period of time is also required for other fish). In carp selection this means a slowing down of selection work by 20 to 30 percent. This is absolutely inadmissible in northern areas where carp mature in the 5th or 6th year of life. A way out is either the use of combined selection or growing of spawners in aquaria where water can be heated at any time of the year.
By comparing two response equations
R= S·h2 and
Rf=Sf·h2f
it is possible to find out which method is more practicable in a given instance. If
S·h2 > Sf·hf2,
then mass selection proves to be more effective than selection for relatives (Kirpichnikov, 1968).
The magnitude of the heritability factor (h2) appears to be different for different characters and different populations. Data are available for carp only (Kirpichnikov, 1958a; Nenashev, 1966; Moav and Wohlfarth, 1967). The following values of h2 have been established:
by the weight of fingerlings and two-year olds, 0.1–0.3;
by exterior characters, 0.3–0.5;
by fat content, 0.2–0.4;
by the number of gill rakers and scales, 0.3–0.5;
by the number of fin rays and vertebrae, 0.4–0.7.
Heritability of weight increases with increasing age and weight of the fish.
In going from mass selection to family selection or progeny testing the heritability factor of weight increases from 0.1–0.3 to 0.8–0.9 and with carefully performed testing to nearly 1. Thus selection for relatives is only advantageous when an increase in h2 is not accompanied by a proportionally greater drop in the selection differential. According to our data in family selection of the Ropsha carp, the selection differential decreases by 3 to 4 times. Thus it follows that with h2 > 0.2, mass selection seems to be more useful; whereas with h2 < 0.2, selection for relatives should be preferred.
In summarizing the results, the following conclusions may be drawn:
Different methods of selection may be applied in fish culture. Selection is the most important method of improving fish breeds.
The choice of the selection method is determined by its response. In most cases the most effective method is that of mass selection.
In selection for weight the choice of method to be applied depends on the level of heritability of individual weight differences.
In selection for exterior characters, as well as for outward countable (diagnostic) characters, mass selection is clearly more effective. The use of mass X-raying (Sengbusch, 1967) permits the application of mass selection for all skeletal characteristics.
In selection for fat content or other characteristics, the determination of which causes fish to die, the only possible method of improvement is selection for parents. However, improvements in live fish investigation techniques may permit the application of mass selection in these cases. A remarkable example is the method of liver biopsy in trout, which can be applied to live fish without damaging them.
As far as the weight character is concerned, it appears practicable to attempt to apply simultaneously different selection methods to the same material. Such a combined method has been applied in Ropsha carp selection. It breaks down into the following stages:
Family selection - several families are obtained and the best of them selected;
Mass selection - superior individuals of the best families are maintained for breeding;
Progeny testing (males of the best families).
Since under our conditions, males reach maturity 1 or 2 years earlier than females, testing of males does not involve any drop in the rate of selection.
In combined selection, responses are summed up by this equation:
| Rc =R1+R2+R3… | (7) |
Combined selection is highly advantageous, particularly when applied to slowly maturing but highly fecund fish.
Donaldson, L.R. and D. Menasveta, 1961 Selective breeding of Chinook Salmon. Trans. Amer.Fish.Soc., 90(2):160–4
Factorovich, K.A., 1968 Histophysiological control over condition of the liver in selected trout. Izv. gosud. nauchno-issled. Inst. ozer. rech. ryb.Khoz., (in press)
Falconer, D.S., 1960 Introduction to quantitative genetics. London, Oliver and Boyd, 365 p.
Kirpichnikov, V.S., 1958a Genetic methods of selection for relatives in carp culture. Dokl. Akad. Nauk SSSR, 121(4):682–5
Kirpichnikov, V.S., 1958b Heterogeneity in populations of wild carp and of hybrid between culture and wild carp. Dokl. Akad. Nauk SSSR, 122(4):716–9
Kirpichnikov, V.S., 1959 Genetic methods of fish selection. Biull.Mosk. Obshck.Ispyt. Prir. (biol.), 64(1):121–37
Kirpichnikov, V.S., 1961 Die genetischen Methoden der Selektion in der Karpfenzucht. Z. Fisch., 10(⅓):137–63
Kirpichnikov, V.S., 1966a Goals and methods of carp selection. Izv. gosud. nauchno.-issled. Inst. ozer. rech. ryb. ryb. Khoz., 61:7–27
Kirpichnikov, V.S., 1966b The methods of progeny testing in carp breeding. Izv. gosud. nauchno.-issled. Inst. ozer. reck. ryb. Khoz., 61:40–61
Kirpichnikov, V.S., 1968 Efficiency of mass selection and selection for relatives in fish culture. FAO Fish.Rep., (44) Vol.4: 179–94
Konradt, A.G. and A.M. Sakharov, 1966 The method of receiving of game fish larvae under hatchery conditions. Izv. gosud. nauchno-issled. Inst. ozer. reck. ryb. Khoz., 61:193–207
Krjageva, K.V., 1966 Influence of stocking rate on viability, growth rate and variability of young hybrid carp. Izv. gosud. nauchno.-issled. Inst. ozer. reck. ryb. Khoz., 61:80–101
Kusema, A.I., 1961 Studies on progeny testing in carp breeding. Trudy ukr. nauch.-issled. Inst. ryb. Khoz., 13:72–84
Kusema, A.I., 1962 Progeny testing in carp selection. Trudy ukr. nauch.-issled. Inst. ryb. Khoz., 14:71–84
Moav, R. and G.W. Wohlfarth, 1963 Breeding schemes for the genetic improvement of edible fish. Prog. Rep. Israel-U.S. Fish Wildl.Serv., 1961–62: 39 p.
Moav, R., 1967 Breeding schemes for the genetic improvement of edible fish. Prog. Rep. Israel, U.S. Fish Wildl. Serv., 1964–65:61 p.
Moav, R., 1968a Genetic improvement of yield in carp. FAO Fish.Rep., (44) Vol.4:12–29
Moav, R., 1968b The relative efficiency of experiments conducted in individual ponds and in ponds divided by nets. FAO Fish.Rep.(44) Vol.4:487–92
Nakamura, N. and S. Kasahara, 1955 On the earliest stage at which the shoot carp appears. Bull. Jap. Soc. sci. Fish., 21(2):73–6
Nakamura, N., 1956 On the effect of particle size and quantity of the food. Bull. Jap. Soc. sci. Fish., 21(3):1022–4
Nakamura, N., 1957 On the result of culturing the modal group and the growth of carp fry reared individually. Bull.Jap.Soc.sci.Fish., 22(2):674–8
Nenashev, G.A., 1966 Heritability of some morphological (diagnostic) characters in Ropsha carps. Izv. gosud. nauchno-issled. Inst. ozer. reck. ryb. Khoz., 61:125–35
Nikolsky, G.V., 1966 On participation of geneticists in working out biological fishery problems. Vestn.mosk. Univ. (Biol.), (6):3–17
Polyksenov, D.P., 1962 Development of highly productive and viable carp stock with purpose of raising new breed for Byelorussia. In Problems of Fish Culture in Byelorussia, Minsk, 5–62
Riggs, C.D. and K.E. Sneed, 1959 The effect of controlled spawning and genetic selection of the fish culture of the future. Trans. Amer. Soc. Fish., 88(1):53–60
Savostjanova, G.G., 1969 Comparative characteristic of different stocks of rainbow-trout (Salmo irideus Gibb.). Trudy Soveshch. Genet., Selekts. Gybridiz. Ryb, (in press)
Sengbusch, V., 1967 Eine Schnellbestimmungsmethode der Zwischenmuskelgräten bei Karpfen zur Auslese von “gratenfreien” Mutanten. Züchter, 37(6):275–6
Steffens, W., 1964 Vergleichende anatomisch-physiologische Untersuchungen und Wild und Teich Karpfen (Cyprinus carpio L.) Z. Fisch. 12(8/10):725–800
Stegman, K., 1965a Variability of weight gains in carp in the first two years of life. Zesz. nauk. Szk. glów Gosp. Wiejsk., 2:67–92
Stegman, K., 1965b Growth of carp in the second and third year. Zesz. nauk. Szk. glów. Gosp. Wiejsk., 2:93–110
Stegman, K., 1968 The estimation of the quality of carp by means of length/height ratio and relative weight gains. FAO Fish. Rep. (44) Vol.4:160–8
Tzoy, R.M., 1968 Correlations between some morphological and physiological traits in Ropsha carp. Izv. gosud. nauchno-issled. Inst. ozer. reck. ryb. Khoz (in press)
Ushakov, B.P., 1963 Change of heat resistance of cells in ontogenesis and the problem of conservatism of cell in cold-blooded animals. In Problems of animal cytoecology, Moscow - Leningrad, 21–42
Wlodek, Y.M., 1968 Studies on the breeding of carp (Cyprinus carpio L.) at the experimental pond farms of the Polish Academy of Science in South Silesia, Poland. FAO Fish.Rep.(44) Vol.4:93–116
Wohlfarth, G.W., R. Moav and M. Lahman, 1961 Genetic improvement of carp, III. Progeny tests for differences in growth rate, 1959 – 1960. Bamidgeh, 13(2):40–54
Woynarovich, E. and H. Kausch, 1967 Hypophysierung und Laicherbrutung bei Karpfen. In Vortragsveranst. über neve Methoden der Fischzüchtung und - haltung, Hamburg, 66–79
by
V.S. Kirpichnikov
Scientific Research Institute on Lake and River Fisheries
Leningrad, U.S.S.R.
Artificial selection and crossing are the principal means of fish improvement. Methods of artificial selection have been dealt with. In the present paper the different methods of the utilization of crossing in fish selection are detailed.
Two principal ways of crossing can be mentioned.
Commercial crossing directed toward breeding of the first-generation hybrids for commercial purposes. The importance of commercial crossing in the fish culture industry is rapidly growing. In crossings of this kind only the first generation hybrid is used, characterized by a heterosis of productive qualities or incorporating the advantageous characteristics of both the parental forms. First generation hybrids, as a rule, are not preserved for further reproduction.
Synthetic crossing (of various degrees of separation, including intergeneric), whose purpose is the development, in the course of long selection, of a new breed. It ought to combine the best qualities of the parents (of 2, 3 or sometimes 4 breeds, 2 or more species or even 2 genera of fish). A particular pattern of crossing in new-breed production should ensure the preservation and perfection of the productive qualities of the breed, the preservation of genetic variability, and the prevention of inbred depression. Lately, use has been made of such crossing patterns that allow heterosis to be utilized to its utmost in every generation.
Both methods of synthetic crossing and the various techniques of crossing in producing new breeds of fish will be concentrated upon later.
Before passing over to them we ought to analyze the role of inbreeding and outbreeding in the fish culture industry.
Inbred depression
Inbreeding is used on a large scale in a wide variety of cattle rearing and plant cultivating industries. It can be used in fish selection as well.
It is known that the inbreeding measure is the coefficient of inbreeding incorporating the degree of the animal's homozygosity. It shows what part of the genes in a group of individuals (expressed in fractions of one, or in percentage of the total genes) in a haploid combination) are in the homozygous state (Falconer, 1960). Close inbreeding, especially sib mating (brothers with sisters and parents with children), causes homozygosity to rapidly increase. Accordingly, the coefficient of inbreeding also increases up to 0.90–0.95 and more (sometimes as high as 1). In most animals with different sexes, inbreeding results in a drop in viability, rate of growth and, quite often in fertility. This deterioration of inbred specimens is termed inbred depression. Inbred depression was found in studies of many fish. A few are innumerated below.
Carp (Cyprinus carpio). A marked drop in the viability and rate of growth of carp during inbreeding has been recorded by many selectionists. It was mentioned by Kuzema (1950, 1953), Shaskolsky (1954), Lieder (1956), Kirpichnikov (1960, 1961, 1966), Schaperclaus (1961), Golovinskaya (1962), and many others in their works.
The problem is dealt with in detail by Moav and Wohlfarth (1963, 1967, 1968). According to their data, the full-sibling mating (brothers and sisters) decreases the growth rate of commercial carp by 10–20 percent. At the same time there is a drop in viability, and the number of anomalous offspring abruptly increases. According to our own data (Kirpichnikov, 1969), even moderate inbreeding, used in the selection of the Ropsha carp, resulted in their deterioration. In certain families, many deviations from normal were observed - the so-called phenodeviants. For instance such deviants were the glass carp, which we observed in a few instances of crossings in the limits of inbred lot M (Kirpichnikov, 1961).
Inbred depression in carp may have a very pronounced manifestation. Estonian carp, for example, subjected to inbreeding for a number of years on small peasant fish farms, have a growth rate of only one-half that of the Ropsha carp.
In accordance with the degree of inbred depression, outbreeding is accompanied by heterosis in growth rate and viability. Especially considerable is heterosis, when fish from different highly inbred groups are crossed. The higher the coefficient of inbreeding in these groups before crossing, the greater the reduction of the coefficient in the outbred offspring. The considerable manifestation of inbred depression in carp can be attributed to the peculiarities of its biology and the structure of its chromosome system, high fecundity (over 1 000 000 eggs), large natural populations, as well as the high natural heterozygosity in this species. The reduction of heterozygosity in these conditions, owing to inbreeding, would always be deterimental.
Common trout and brook trout. As regards the salmon family, data on the effect of inbreeding are scarce and discrepant. In brook trout (Salvelinus fontinalis) the occurrence of inbred depression was established by Cooper (1961). In inbred lines, the growth rate of fish was retarded. However, in the experiments conducted in Ropsha (Savostjanova, 1969), in crossing trout of different origin, heterosis has not been detected. This evidence seems to indicate that there is no inbred depression in these trout. In evaluating the possibility of inbred depression in man-bred salmon, it should be borne in mind that their fecundity is relatively low (up to 2–3 thousand, rarely up to 5–10 thousand eggs from one female). The fish breeder has to keep a great number of spawners, frequently in the thousands range. The number of fish being so great, the hazard of the coefficient of inbreeding increase is insignificant, and the population as a whole remains quite heterogeneous. This fact can easily account for the absence of marked heterosis in crossing different groups of trout. In close inbreeding effected by the selectionist, inbred depression may easily result; this depression however, is eliminated in crossing.
Data pertaining to other fish are very scarce. Gibson (1954) points out that in the guppy (Lebistes reticulatus) there is a drop in viability resulting from inbreeding. Krivoshchekov (1963) believes that the reduction in the size of crucian carp in small lakes can be attributed to inbred depression, resulting from inbreeding with small populations of crucian carp. There are scattered data pointing to the detrimental effect of inbreeding in other kinds of fish. It should be remembered that the breeding of many kinds of aquarium fish is accompanied by very close inbreeding. The inbred depression in this case can be overcome by a careful selection of the best specimens. Probably this peculiarity of aquarium fish is explained by their low fecundity and the occurrence in nature of a great multitude of small isolated populations. In small populations, inbreeding is inevitable. The species gets adapted to the low heterozygosity, and for this reason inbred depression manifests itself to a very insignificant extent.
Now to come back to the principal object of pond breeding. Fish of the carp and salmon families, bullheads, sunfishes, tilapia and other commercial fish are widely cultivated at present. Considerable inbred depression is to be expected in all these species. Crossing patterns used in selection should be designed to overcome the depression and to make maximum use of heterosis.
In fish selection as well as in selection of other animals, distant outbreeding is indispensable. The purpose of such crossings may be described thus:
An overall increase of genetic variability results in an increase of artificial selection response.
The achievement of a combination of characters of two or three breeds or two (rarely three) species.
The improvement of the productive quality of the local breed by making use of the few valuable traits of another breed (improver).
The increase in the viability of the breed by introducing genes responsible for resistance to environmental factors and diseases.
In accordance with the above objectives, following the original crossing, the reproduction of the hybrid population is carried out by means of reproductive, introductory, absorptive or alternating crossing.
This method of crossing is carried out when many valuable properties from both parents are to be combined in the hybrid. It can be easily effected with complete fertility of the hybrids and requires only a meticulous selection in the subsequent generations. In fish breeding, we know of a few instances of successful reproductive crossing. With its help the Ropsha North carp has been obtained, the Ukrainian breeds of carp have been reared, and work is being carried out to rear the Mid-Russian carp. There are reasons to believe that the pre-world war II Hungarian carp was the result of three-way crossing of the local less productive breed with the German Aischgrund carp and the carp brought from Japan. For further reproduction of the hybrids reproductive crossing must be used. It was also applied in the rearing of rainbow trout (after the crossing of two or three American trouts) as well as in similar work with some other fishes.
This system can be used to advantage when one or a few characteristics from a breed must be incorporated in the hybrid. Each generation of the hybrid must be crossed with the local breed of fish. The hazard of losing the useful characteristics of the improved breed in back crossings of this kind is very high. If the characteristics are determined by one or several clearly segregating genes, the problem is solved with little difficulty. In the case of polygenic inheritance of the properties selected, the selection difficulties are sometimes impossible to overcome.
There have been no indications in the literature that introductory crossing has been put into practice. Its application is envisaged in the selection of Israeli carp in two cases; in the crossing of the local highly productive carp with those brought from the Netherlands and in the making of breed strains marked by the colouring genes. The first introductory crossings have already been carried out with the object of introducing colouring markers into the best strains of the Israeli carp (Moav and Wohlfarth, 1967). The markers in this case are three recessive genes, determining gold, blue and grey colouring of the fish.
Introductory crossing has a lot of potentialities when the selectionist attempts the task of increasing a breed's resistance to a certain disease. Resistance to disease often depends on the presence of one or a few genes, and they can be preserved by means of meticulous selection in each generation. Possibly, selectionists will succeed in rearing breeds of fish that will have high resistance to some of the most dangerous diseases, such as infectious dropsy, furunculosis, sanguinicolosis, ichthyophthiriasis, and some others.
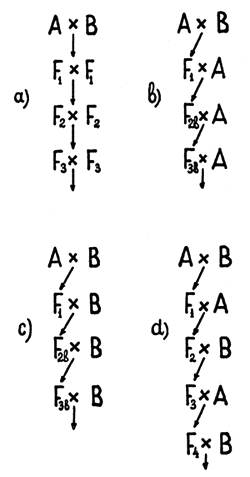
Fig. 1 Types of crossings in synthetic selection.
| a | - | Reproductive |
| b | - | Introductive |
| c | - | Absorptive |
| d | - | Alternative |
This resembles introductory crossing. The only difference is that the purpose of this type of crossing is a nearly complete substitution of the local breed of genotype by the genotype of the improved breed. Only some peculiarities typical of the local breed must be preserved, pertaining mainly to its viability.
In both introductory and absorptive crossing, selection is seriously handicapped if the high viability of the local breed is determined by a large number of genes.
As new high-quality fish breeds are reared, the significance of absorptive crossing in commercial fish breeding will increase.
This method requires the most complicated system of rearing, and so far it has not been used in commercial fish rearing. It is especially advantageous when a combination of many characteristics from two breeds with polygenic inheritance is required. Alternating crossing provides for the preservation of high genetic variability through a number of generations. Owing to this variability, selection efficiency is kept at a high level and selection plateaus do not result.
After several (3,4 or more) alternating crossings, they must be replaced by the conventional reproductive crossings. Otherwise, stabilization of breed characteristics will be difficult to achieve.
It should be noted that with alternating crossing, the coefficient of inbreeding practically does not increase. This enables specimens from each hybrid generation to be used directly for commercial breeding.
Undoubtedly, alternating crossing as represented in Fig.1d and in more complicated forms will be used in the future for fish selection. However, it will require experienced selectionists and well-equipped selection stations. If the selectionist faces the problem of obtaining new hybrid breeds by means of crossings (interspecific or intergeneric), the principal difficulty in hybrid selection is their complete or partial sterility. The restoration of normal fertility takes a lot of hard and time-consuming work. The results of Kuzema and Tomilenko's (1965) experiments with common and crucian carp hybrids indicate that back crossing of the hybrids with one of the original species may help restore the fertility in the hybrid form.
To utilize completely the advantages associated with heterogeneous crossings, fish breeding should be carried out according to a rigid pattern. We shall analyze some of the possible breeding systems which offer good prospects for the breeding of pond fish.
While working with slowly maturing fish, it is practicable to have two or three groups concurrently within a breed, without intermingling, allowing inside each a moderate inbreeding, and carrying out selection in each generation. For commercial purposes, fish from different groups are crossed. The prevention of close inbreeding enables one to avoid considerable exhaustion of hereditary variability within the breed. This method is presently used for the selection of the Ropsha carp.
In each generation a large number of crossings of fish from different groups is carried out (in Israel up to 20 combinations a year). The parents producing the best offspring are used for subsequent commercial crossings. For further reproduction those offspring are selected that have maximum overall combining ability. The weak spot of such a system of breeding is the gradual narrowing of the genetic variability in carrying out family selection.
Moav and Wohlfarth (1967) recommend, that in selecting two groups marked by certain genes, a reserve group of fish, numerous enough, should be kept for each group. In case of a genetic variability drop, it will allow an additional gene pool to be introduced into the exhausted groups. As in the previous case, selection should aim at an increase in combining capacity.
After two or three generations of close inbreeding, the evaluation of hybrids from different inbred lines is performed. The best combinations are used for commercial rearing and among the offspring new inbred lines are established. It seems possible to obtain 4-step hybrids. The practicability of such a system in fish breeding must be corroborated by appropriate experiments.
Linear selection involving inbreeding for superior ancestors was recently started by A.I. Kuzema in 1950 in his experiments with the Ukrainian carp. The problems in this type of selection lie chiefly in the difficulties of accurate evaluation of spawners and the necessity to choose a few of the best of a great number of fish.
The term topcross is applied to a selection technique whereby one or several inbred lines and a large outbred group are maintained at the same time. To preserve the genetic variability and to improve the offspring characteristics, the crossing is done between the best inbred specimens (males for instance) and specimens from the outbred population (females).
Topcross may be applied in commercial fish breeding, especially if inbreeding results in a considerable drop in female fecundity and viability.
Reciprocal recurrent selection being one of the most complicated techniques of animal breeding at present, is not likely to find its way to fish breeding in the near future. In reciprocal selection, the combining capacity of the parents from each of the two breed groups is evaluated by means of a cross with parents from the other group. The specimens, selected as a result, are reproduced without recrossing and their offspring are again tested for combining potential.
The basic peculiarity of all the systems considered is the utmost utilization of heterosis in crossing fish from different groups, lines and breeds. Alongside this, inbreeding is used, varying from quite moderate, in some cases, to very close in others. The future will decide which breeding techniques are most effective in the breeding of pond fish. The correct choice of techniques will be primarily a function of our knowledge of genetics and of the perfection of our knowledge of inbred depression and heterosis in fish crossing.
Apart from selection and crossing, special genetic and cytogenetic techniques are widely used in present day selection, some of which are applicable to fish. A detailed review of such methods can be found in the article on the objectives and techniques of the selection of carp. What follows is a brief outline of this article (Kirpichnikov, 1966).
An illustrative example was given while discussing the marking of selected groups of carp by colouring mutations. Another example is the backcross study of homo-and heterozygosity in the scaled carp in the gene s (mirror and scattered scale). The backcrossing is carried out thus:
| 1) | S(s? )nn scaled | × | ssnn scattered |
| 2) | S(s? )nn scaled | × | Ssnn scaled heterozygous |
In the former case, the scaled and scattered offspring of the heterozygous parents are produced in a 1:1 ratio. In the latter case, the ratio is 3:1 (three scaled per one scattered). The entire offspring of the homozygous scaled parents in any of the two crossings have a completely scaled integument. Within the period from 1956 till 1964, 469 parents of the Ropsha carp were examined by this technique, 247 turning out to be homozygous. This work enabled the selectionists to eliminate completely the occurrence of the scattered carp as early as in the 5th selected generation.
The better we know the genetics of the fish under selection, the more data we have at our disposal on the inheritance of certain quantitative and qualitative characters, rendering selection easier and faster.
Increase in the number of mutations caused by strong mutagenic actions may considerably increase the populations' heterozygosity and selection effectiveness. Research in this direction was initiated in Leningrad in 1967 on the carp (Tzoy, 1969).
The very first experiments with chemical mutagens proved that mutation rates may be multiplied many-fold. Artificial mutagenesis can be used to the best advantage in the selection of pond fish for resistance to certain dangerous infections and parasitic diseases.
These reproduction procedures may be quite useful in fish breeding if it is necessary to rapidly increase the homozygosity of a selection group or line. The problems of gynogenesis are dealt with in another paper in this report.
Polyploidy in fish is feasible. Sex in fish is determined by sex chromosomes (gonosomes) rather than the balance between sex chromosomes and autosomes. This makes it possible, as was shown by Victorovsky (1969), to double chromosome sets in the course of evolution (as it must have occurred in the carp and salmon families). However, the possibility of obtaining practical artificial polyploids in fish is questionable.
A few of the most important selection techniques are described below:
Selection of carp and many other kinds of fish with external fertilization is facilitated by using artificial fertilization. In some fish, such as trout and whitefish, eggs and sperm can be obtained without hormone injections. In other fish, such as carp, crucian carp, grass carp, silver carp and others, injection of gonadotrophic hormones is used, mainly the hormones of the hypophysis. Ovulation occurs at a certain time after the intramuscular injection of the hormone. In carp this period varies from 10 to 30 hours, depending on maturity and water temperature. After the injection, fish are kept in special breeding cages.
Prior to spawn taking, large fish should be anesthetized (by quinaldine, MS-222, or other drugs).
The spawn and milt are placed into basins or china cups and stirred without adding water (the so-called Russian method of fertilization). If the fish have non-adhesive spawn, the fertilization can be performed in a small volume of water. The optimal spawn and sperm ratio varies from species to species. In carp it is from 50:1 to 100:1.
After fertilization, adhesive eggs require washing away of the agglutinating agent, and its further secretion must be stopped. Degumming is done by means of two methods:
Woynarovich's method (Woynarovich and Kausch, 1967).
A solution of a mixture of sodium chloride (4 g /litre H2O) and urea (4 g /litre H2O) is added to the mixture of spawn and milt. One volume of spawn requires two volumes of the solution. During the first 3–5 minutes the spawn is carefully and continuously stirred; fertilization takes place at that time. The subsequent agitation is done intermittently every 2–3 minutes. As the spawn swells, additional batches of the solution are added.
In 1–1.5 hours the excess solution is poured out and the spawn is placed in another solution (1.5 g of tannin / litre of H2O) for 10 seconds to strengthen the membranes. The spawn should be continuously stirred. Following that, the spawn is washed with fresh water. A little later it is treated once or twice with a weaker solution of tannin. As a result, the spawn is completely degummed and can be placed into the Zuger or Weiss apparatus. At the end of this treatment the spawn is much larger in size due to considerable swelling.
The Konradt and Sakharov method (Konradt and Sakharov, 1966)
A solution containing hyaluronidase is added in little portions to the mixture of spawn and milt. The hyaluronidase enzyme can be easily obtained from the testicles of oxen and pigs. (Tetz, 1963). The solution is maintained at a level of 1–1.5 cm above the eggs and the spawn is continuously stirred by a soft feather. In 15–18 minutes, without removing the hyaluronidase, a tannin solution (100 mg / Litre) is added to the spawn, also in little portions. The spawn is degummed 40–50 minutes after the commencement of the treatment. On making sure that the spawn no longer tends to agglutinate, it is transferred to the incubating apparatus.
With the utilization of artificial fertilization, the spawn and milt can be divided into many equal parts, the percentage of fertilization can be accurately determined, as well as embryo survival, rate of development, etc. All these factors make artificial fertilization indispensable in selection, and especially in evaluating the reproductive capacity of the parents.
In fish selection, aquaria are necessary, above all, for checking the quality of spawn, larvae and fry from different crossings. Recently, German and Japanese scientists developed a method of purifying water coming out of fish aquaria and a closed-circuit water supply for aquaria became possible. This allows one to grow fish all year round in aquaria with running water, keeping the temperature at the optimal level without consuming too much fresh water (Sengbusch et al., 1967).
Quite a number of marking techniques have been tested. The best results obtained were by cutting the fins of fish, branding signs on the skin and scales by a red-hot wire, or subcutaneous injections of India ink and some organic dyes (dichlorotriazine and other compounds). For certain fish, suspended tags can be used. Marking makes it possible to identify different varieties or groups of fish, as well as the best individual spawners if numerals are used.
It is noteworthy that great importance is attached to the techniques of examining the fish during their life time, such as X-raying, biopsy, etc. which are successfully being developed now in a number of laboratories.
Cooper, E.L., 1961 Growth of wild and hatchery strains of brook trout (Salvelinus fontinalis). Trans. Amer. Fish. Soc., 90 (4): 424–38
Falconer, D., 1960 Introduction to quantitative genetics. London, Oliver and Boyd, 365 p.
Gibson, M.B., 1954 Upper lethal temperature relations of the guppy, Lebistes reticulatus. Canad.Zool., 32:393
Golovinskaya, K.A., 1962 Selective breeding in pond fish culture. Rybovod. i rybolov (3):7–10
Kirpichnikov, V.S., 1960 Development of selective breeding in carp culture. Nauch.-tech. Biull.GosNIORKH, (ii):38–40
Kirpichnikov, V.S., 1961 Die genetischen Methoden der Selektion in der Karpfenzucht. Z.Fisch., 10(⅓): 137–63
Kirpichnikov, V.S., 1966 Goals and methods of carp selection. Izv. gosud. nauchno-issled. Inst. ozer. rech. ryb. Khoz., 61:7–27
Kirpichnikov, V.S., 1969 The North Ropsha hybrid carp. Izv. gosud. nauchno-issled. Inst. ozer. rech. ryb. Khoz. (in press)
Konradt, A.G., and A.M. Sakharov, 1966 The method of receiving of game fish larvae under hatchery conditions. Izv.gosud. nauchno-issled. Inst.ozer.rech.ryb.Khoz., 61:193–207
Krivoshchekov, G.U., 1963 Inbreeding and heterosis in lake fish culture. Razvit.ozer.ryb. Khoz. Sibiri, Novosibirsk, 45–8
Kuzema, A.I., 1950 Organizations and principles for selective improvement of carp in Ukrainian fish farms. Trudy ukr.nauchno-issled. Inst. prud.ozer.-rech. ryb.Khoz., 7:107–37
Kuzema, A.I., 1953 Ukrainian carp strains. Trudy Soveshch.vop.prudov.rybov., Moscow,65–70
Kuzema, A.I., and V.G. Tomilenko, 1965 The development of new cyprinid fishes by means of intergeneric hybridizations. Rhyb.Khoz., Kiev, 2:3–17
Lieder, U., 1956 Uber einige genetische Probleme in der Fischzucht. Z. Fisch., 5(½):133–42
Moav, R. and G.W. Wohlfarth, 1963 Breeding schemes for the genetic improvement of edible fish. Prog.Rep.Israel - U.S.Fish.Wild.Serv., 1962, 39 p.
Moav, R., 1967 Breeding schemes for the genetic improvement of edible fish. Prog.Rep. Israel - U.S.Fish.Wildl.Serv., 1964–1965, 61 p.
Moav, R., 1968 Genetic improvement of yield in carp. FAO Fish.Rep. (44) Vol.4:12–29
Savostjanova, G.G., 1969 Comparative characteristic of different stocks of rainbow trout (Salmo iridens Gibb.) Trudy Soveshch. Genet., Selects. hybridiz. ryb, Moscow (in press)
Schäperclaus, W., 1961 Lehrbuch der Teichwirtschaft, 2 Aufl., P. Parey, Berlin and Hamburg, 582 p.
Sengbusch, R., et al., 1967 Gewichts-Zunahmen von Karpfen in Kleinstbehältern, Zugleich ein Beitrag zur Aufklarung des Raumfaktors. Z.Fisch., 15(½):45–60
Shaskolsky, D.V., 1954 Inbreeding in commercial carp fish farms. Trudy Vseross.nauchno-issled. Inst.prudov.ryb.Khoz., 7:22–3
Tetz, V.I., 1963 Degumming of fertilized eggs of carp by means of enzyme hyaluronidase. Ryb.Khoz., 7:21–3
Tzoy, R.M., 1969 Mutagenetic action of some chemicals upon eggs and sperm of rainbow trout (Salmo gairdneri) and peled (Coregonus peled). Tsitologia (in press) (In Russian)
Victorovsky, R.M., 1969 The possibility of polyploidy in fish evolution. Trudy Soveshch. Genet., Selects. hybridiz. ryb. (in press)
Woynarovich, E. and H. Kausch., 1967 Hypophysierung und Laicherbrütung bei Karpfen. In Vortragsveranst. über neue Methoden der Fischzüchtung und haltung, Hamburg, 66–79
![]()
![]()
![]()