![]()
![]()
![]()
par
Bruce J. Zobel 1 et Keith W. Dorman 2
GENERALITES
Il y a de très nombreuses plantations de pin à l'encens (Pinus taeda L.) aux Etats-Unis dans les régions du sud et du sud-est. C'est une espèce qui s'adapte très aisément, facile à entretenir et dont le bois est de bonne qualité. En raison de l'étendue considérable de son aire naturelle, elle présente une certaine complexité génétique, et comprend un certain nombre de races géographiques. Elle s'hybride si aisément avec plusieurs autres espèces qu'il est difficile de définir un pin à l'encens pur. Il semble cependant qu'elle soit toujours l'espèce “dominante” et les régions peuplées d'essences mélangées tendent à devenir des peuplements de pin à l'encens.
A cause de sa grande variabilité géographique, l'introduction du pin à l'encens n'a pas été aussi réussie ou aussi répandue qu'elle aurait dû l'être. Un grand nombre de tentatives de plantation ont échoué parce que les semences utilisées étaient d'origine inconnue et avaient été plantées dans des milieux auxquels elles n'étaient pas adaptées. Ces échecs sont à l'origine de l'opinion regrettable qui est répandue dans de nombreuses régions subtropicales, et selon laquelle le pitchpin américain (Pinus elliottii), qui s'adapte à des conditions plus diverses, est une essence meilleure que le pin à l'encens.
Autrefois, il n'était pas toujours facile d'obtenir des sources convenables de pin à l'encens, mais le Centre de graines forestières récemment institué par le Service des forêts des Etats-Unis a rendu de grands services 3. Au fur et à mesure que des lignées améliorées deviennent disponibles et que la diversité géographique de l'espèce est bien reconnue et mise à profit, la valeur du pin à l'encens continuera d'augmenter en tant qu'essence exotique. Cet exposé met en relief: (1) certaines des principales caractéristiques du pin à l'encens, et (2) quelques notions à prendre en considération lorsqu'on est appelé à l'employer comme essence introduite.
1 Professor of Forest Genetics, North Carolina State University, Raleigh, N.C.
AIRE DE DISTRIBUTION DU PIN A L'ENCENS
L'aire de distribution du pin à l'encens présente à peu près la forme d'un rectangle orienté d'est en ouest dans le sud-est des Etats-Unis. Il y a cependant des prolongements vers le nord-est de long de la côte atlantique et vers le sud en Floride (Fig. 1). On ne trouve pas ce pin dans le bassin du Mississippi. Les emplacements des sources de semence qui se sont distinguées par leur performance sont illustrées; les sources mentionnées ainsi que d'autres sont examinées plus loin. De même, la Figure 1 montre les régions physiographiques: Coastal Plain, Fall Line ou Sandhills et Piedmont, qui sont importantes pour notre discussion.
Figure 1: Aire botanique du pin à l'encens (Pinus taeda L.) et régions physiographiques importantes
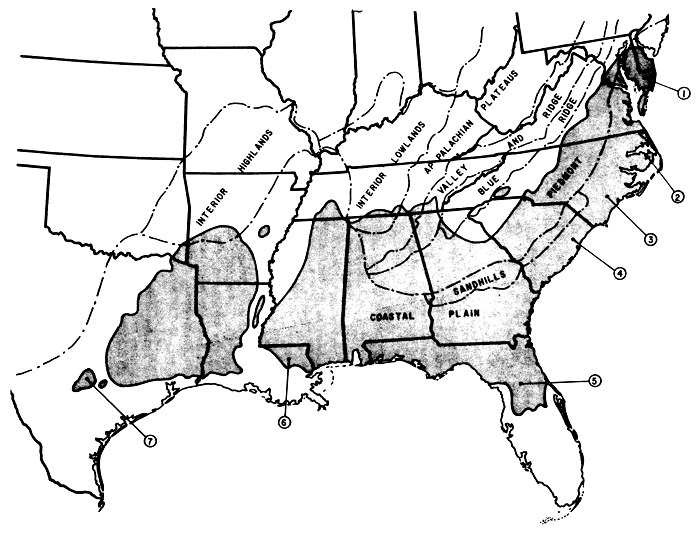
Légende des sources de semences
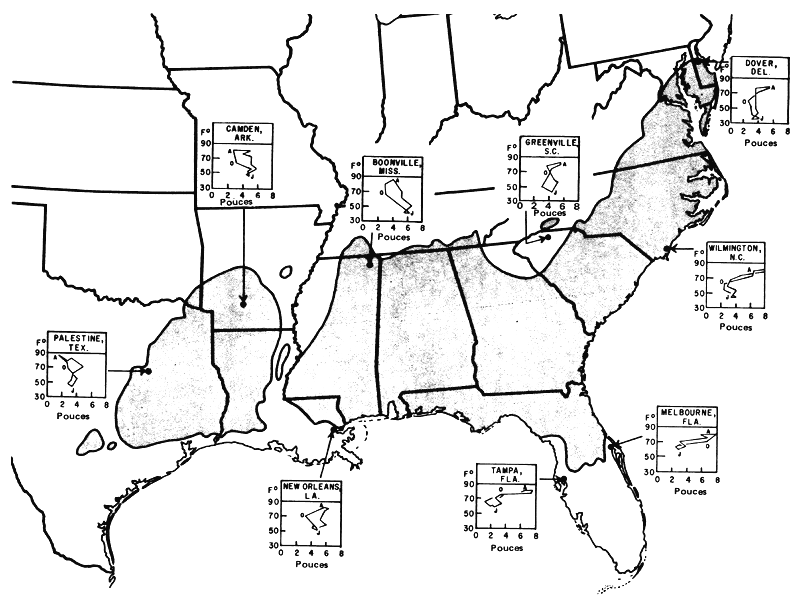
Figure 2: Eventail des types de climat dans toute la région du pin à l'encens (Pinus taeda L.) (d'après Wahlenberg W.G. et Ostrom, C.E., 1956 “Geographic variation in climate in the loblolly pine region”; Res. Note S.E. For. Exp. Sta. No 94).
On trouve également dans la Figure 2 des climographes qu'on peut utiliser conjointement avec les longitudes et les latitudes pour l'évaluation des similitudes de climat lorsque le pin à l'encens doit être utilisé comme essence introduite. En Floride, les précipitations sont élevées pendant les périodes de haute température, contrairement qu'au Texas où elles sont faibles dans les périodes chaudes. Dans chaque graphique, les lettres J, A et O indiquent les mois de janvier, août et octobre. Ces données sont tirées de résumés climatologiques détaillés figurant dans une autre communication (Hocker, 1955).
CARACTERISTIQUES DU PIN A L'ENCENS
Le degré et le genre de variation du pin à l'encens sont d'une importance telle qu'ils sont appelés à jouer un rôle de premier plan dans n'importe quel programme d'amélioration d'arbres, qu'il s'agisse d'introduction de semences, de sélection de races, de sélection d'arbres d'élite ou d'hybridation. On sait à présent beaucoup plus long sur cette espèce que sur n'importe quel autre pin austral, mais les connaissances relatives au genre de variation sont bien plus complètes que celles qui concernent le degré précis de variation de chaque caractère. La discussion qui suit se borne à citer quelques références choisies se rapportant à chaque caractère ou sujet, mais on trouvera des renseignements complémentaires dans une communication plus détaillée intitulée “La génétique du pin à l'encens” (The Genetics of loblolly pine) (Dorman and Zobel, 1973).
Variation et hérédité
Taux de croissance. L'étude d'un vaste échantillonnage de sources de graines, patronnée par le Comité de l'amélioration des arbres forestiers, a démontré qu'à l'âge de 5, 10 et 15 ans il y avait de fortes variations clinales dans la croissance, les arbres des régions côtières se développant généralement plus rapidement que ceux de l'intérieur, et cela partout à l'exception des plantations situées dans les zones les plus froides (Wakeley, 1961; Wells et Wakeley, 1966; Wells, 1969). Lorsque des arbres étaient transplantés d'une extrêmité de l'habitat à l'autre, leur production en volume était inférieure à la moitié de ce que produisent les arbres issus de semences locales, à cause de leur médiocre survivance combinée à la lenteur de leur croissance; mais lorsque des arbres d'origine côtière étaient transportés à une distance de 360 km vers l'intérieur dans la parte méridionale de l'habitat, ils se développaient plus rapidement que les arbres originaires des régions intérieures.
Quatre des 15 sources de pin à l'encens faisant l'objet de cette étude ont été considérablement moins attaqués par la rouille fusiforme que les autres. Ces arbres relativement résistants provenaient de la région située à l'ouest du Mississippi, à Livingston Parish, Louisiane, et sur le littoral oriental du Maryland. Une autre étude portant également sur un vaste échantillonnage qui comprenait 36 sources de semence plantées à Crossett (comté de Ashley) dans l'Arkansas, a aussi démontré le potentiel de croissance rapide du pin à l'encens provenant des régions côtières et en particulier d'arbres originaires des Carolines (Grigsby, 1973).
Les résultats de ces études étendues ont été encore corroborés et complétés par d'autres recherches dans lesquelles un très grand nombre d'échantillons de pin à l'encens a été recueilli dans les Etats de Géorgie (Kraus, 1967), de l'Alabama (Goggans, Lynch et Garin, 1972), du Mississippi (Wells et Switzer, 1971) et de la Caroline du Nord (Lantz et Hofmann, 1969). Une étude effectuée sur plusieurs populations de pin à l'encens de l'Etat de la Louisiane qui ont été plantées en d'autres endroits dans le même Etat a révélé que celles provenant de Livingston Parish et de Louisiane occidentale étaient résistantes à la rouille, mais que les différences en hauteur entre les sources septentrionales et méridionales de la Louisiane, bien qu'elles pussent paraître suggestives, ne sont pas statistiquement significatives (Crow, 1964; Merrifield, 1965).
Les recherches effectuées séparément à l'intérieur des Etats ont démontré qu'en plus des modes de variation établis par l'étude du vaste échantillonnage de sources mentionné, le pin et l'encens est sujet à des variations clinales en ce qui concerne la survie, le potentiel de croissance jusqu'à un certain degré, et la résistance à la rouille fusiforme. Les pins à l'encens du Golfe du Mexique et des côtes méridionales de l'Atlantique survivent un peu moins bien à la plantation et possèdent moins de résistance propre à la rouille que ceux des régions intérieures.
Il existe de grandes différences individuelles de croissance entre les arbres-mères et qui leur sont inhérentes, en plus de celles que l'on observe entre les sources géographiques. Au Texas, la croissance des descendants d'arbres-mères de pins à l'encens dans les régions de production de semences était en moyenne à peu près la même que celle des arbres témoins, mais ces descendants étaient plus uniformes, et il y avait de grandes différences entre les diverses familles (van Buijtenen, 1969). Les dix familles dont le rendement était le plus grand ont produit 1,82 cordes (1 corde = 3 625 m de circonférence) par acre par an, les témoins 1,54 cordes et les dix familles dont la croissance était la plus faible, 1,14 corde. La croissance la plus faible observée dans une famille était de 0,90 corde par acre et par an, et la croissance la plus élevée était de 2,05 cordes. On a observé de grandes différences dans la production de bois sec par acre entre les familles à pollinisation libre - de certains arbres-mères sélectionnés pour des vergers à graines de type industriel (Zobel, Kellison et Matthias, 1969). La production variait entre 1,8 et 2,7 tonnes de bois sec par acre et par an (4,5 et 6,75 tonnes par ha et par an) chez des familles âgées de 7 ans dans des stations d'excellente qualité. D'une manière générale, on s'attend que la croissance des descendants provenant de vergers à graines clonaux épurés soit d'environ 10 à 20 pour cent supérieure à celle des réserves de plants pour usage commercial. A la suite de la plantation en Géorgie centrale des descendants de onze arbres sélectionnés en diverses régions du sud-est, du centre et du nord-ouest de cet Etat, on a observé que la hauteur des arbres de la famille qui a accusé la croissance la plus rapide dans chacune des trois régions mentionnées était, au bout de cinq saisons de croissance, de 17 à 31 pour cent supérieure à celle des arbres de la famille dont la croissance était la plus lente (Barber, 1966). Les arbres de la famille ayant la croissance la plus lente et provenant de la source dont la croissance était la plus lente, avaient une hauteur moyenne de 2,5 m, et les arbres de la famille accusant la croissance la plus rapide et provenant de semences de la source à laquelle était attribuée la croissance la plus rapide avaient une hauteur moyenne de 3,65 m - hauteur supérieure d'environ 43 pour cent - ce qui indique que les effets de la sélection de la race et de la sélection d'arbres d'élite peuvent être cumulatifs.
Forme de la cime et du fût. Le pin à l'encens est une espèce extrêmement vigoureuse et de port relativement médiocre; certains arbres peuvent par conséquent avoir de longues et lourdes branches et un élagage naturel médiocre, des branches formant un grand angle avec l'horizontale, des tiges fourchues ou tordues. Il y a cependant des arbres qui sont certainement dotés d'une cime de bonne classe combinée à de bonnes qualités de fût, et c'est sur ces sujets que la sélection devrait se concentrer.
On trouve dans les essais de descendance de grandes différences dans la forme du fût et de la cime. La transmissibilité héréditaire de la largeur de la cime chez des descendants âgés de 7 ans produits par fécondation libre d'arbres à cimes bien formées et d'arbres à cimes de conformation médiocre était de 34 pour cent (Trousdell, Dorman et Squillace, 1963). Au Texas, on a observé que le rapport de la longueur moyenne des branches à la hauteur de l'arbre était inférieure à celui des témoins chez les descendants de trois sujets sur cinq choisis pour leur bonne qualité; mais il n'y avait pas de différences significatives dans les moyennes des angles que forment les branches (Goddard, Brown et Campbell, 1959).
En Floride, on a effectué des fécondations contrôlées parmi des pins à l'encens tordus et on a comparé les descendants avec ceux issus d'arbres droits (Perry, 1960). A l'âge de deux ans, les descendants issus de croisements entre des arbres droits comportaient quatre fois plus de plants droits que les descendants issus de croisements entre arbres-mères tordus. A l'âge de sept ans, il y avait à peu près la même proportion d'arbres droits et tordus (Goddard et Strickland, 1964). On en a conclu qu'il y a des aptitudes de combinaisons générales et spécifiques assez fortes en matière de rectilignité du fût.
Propriétés du bois. Le bois dense et résistant du pin à l'encens est employé pour la fabrication de nombreux articles en bois et types de papier. En même temps qu'on cherchait à améliorer la croissance et le rendement, et lorsque cela était possible les caractéristiques de divers produits du bois, des études étaient faites sur les variations des caractéristiques du bois et de la fibre, afin de maintenir une qualité de bois supérieure et uniforme. Et pour pouvoir déterminer les différences de qualité inhérentes au bois, on a élaboré des techniques non destructives d'échantillonnage permettant d'évaluer les différentes caractéristiques du bois par rapport à la taille et à l'âge de l'arbre et aux facteurs de son environnement. Les résultats de l'étude de ces techniques ont été publiés en résumé (Zobel, 1961, 1964) et il n'y a pas lieu d'y revenir ici. Des recherches très étendues ont été faites sur les variations géographiques de la qualité du bois à cause de leur importance pour le rendement en pâte et la qualité de celle-ci, comme pour le classement du bois d'oeuvre. Des recherches effectuées par le laboratoire des produits forestiers des Etats-Unis (1965) dans tout le sud-est du pays, par les industries et par les universités, il ressort que le poids spécifique du bois diminue du sud-est au nord-est et de la côte au Piedmont. Des différences assez importantes et régulières ont été observées; ces différences concernent principalement le rendement et la qualité de la pâte, et elles ont joué un rôle important dans les activités forestières et la structure de la propriété foncière dans cette région.
Dans les Carolines, on a observé que le poids spécifique du bois, le pourcentage de bois d'été, les matières extractibles par un mélange alcool-benzène, le rendement en pâte et la résistance à la tension à la portée zéro étaient plus élevés chez les arbres de la Plaine littorale que chez ceux du Piedmont (Einspahr, Peckham et Mathes, 1964). Dans le Maryland et le Delaware, le poids spécifique et la longueur des trachéides diminuent du sud au nord (Whitesell, Zobel et Roberds, 1966). Dans le sud-est des Etats-Unis, on a trouvé que le poids spécifique du bois n'était pas toujours le même et qu'il y avait différents types de cellulose, mais les différences observées étaient plus faibles entre les divers emplacements géographiques qu'entre les arbres pris individuellement (Zobel et McElwee, 1958; Zobel, Thorbjornsen et Henson, 1960).
Dans une région donnée, le poids spécifique et la teneur en humidité des différents arbres sont d'ordinaire en rapport étroit (et inverse); par conséquent le poids spécifique est lié de très près à la teneur en humidité. Cependant, cette corrélation n'est pas toujours valable pour les distributions géographiques; on a trouvé en effet que la teneur en humidité du pin à l'encens à faible poids spécifique dans la région montagneuse du Tennessee était élevée comme prévu, tandis que dans certaines régions de la Plaine côtière de Géorgie, où le bois a un poids spécifique élevé, la teneur en humidité était plus élevée que celle des arbres du Piedmont adjacent (Zobel, Matthias, Roberds et Kellison, 1967).
Des études poussées ont été faites sur les variations individuelles de la longueur des trachéides et d'autres caractères morphologiques en fonction de la croissance. La première publication importante dans ce domaine, par Zobel et Rhodes (1955), fait état d'amples variations dans le poids spécifique d'un arbre à l'autre, d'une région géographique à l'autre et à l'intérieur d'une même région, mais qu'il n'y a qu'une faible corrélation entre la croissance en diamètre et le poids spécifique. Des constatations similaires figurent, en outre, dans de nombreux autres travaux publiés. Faute de pouvoir les citer tous, mentionnons les communications de van Buijtenen (1963), Einspahr, van Buijtenen et Peckham (1969) et Zobel et al. (1969). On peut résumer les conclusions de la majeure partie des publications comme suit:
Pour toutes les caractéristiques du bois étudiées, on a observé de grandes variations d'un arbre à l'autre parmi les arbres du même âge appartenant à la même station. Ces différences dépendent en grande partie de facteurs génétiques avec une héritabilité au sens strict qui est habituellement de 0,5 à 0,6 environ.
Un grand nombre de caractéristiques du bois (par ex., le poids spécifique et la longueur des trachéides) ne sont pas en étroite corrélation. Il est donc possible d'opérer une sélection séparée.
Les propriétés du bois dépendent principalement de l'âge de l'arbre, les jeunes sujets ayant un bois de moindre densité, des trachéides plus courtes et des parois cellulaires plus minces.
Les propriétés du bois sont affectées par les divers facteurs de l'environnement, y compris les fertilisants. Ce sont les engrais azotés qui semblent avoir le plus d'influence sur le bois du pin à l'encens.
Chez les arbres d'un certain âge, le taux de croissance et le poids spécifique du bois ne sont pas en étroite corrélation. Ce manque de corrélation permet le développement de lignées d'arbres à croissance rapide dont le bois peut avoir un poids spécifique élevé ou faible.
Les arbres des régions à hautes précipitations estivales et à climat chaud tendent à avoir du bois de densité plus élevée que les arbres des régions sèches ou à climat plus froid. Cette tendance s'est bien vérifiée, tant en ce qui concerne le poids spécifique que d'autres caractères morphologiques du bois.
Le mode de variation des caractéristiques du bois, et en particulier les différences d'un arbre à l'autre, est tel que la sélection en vue d'améliorer les qualités du bois doit donner d'encourageants résultats.
Résistance à la sécheresse et au froid. Dans certaines zones de la partie ouest de l'habitat du pin à l'encens, les précipitations sont faibles pendant les périodes de chaleur, ce qui a amené à inclure, dans les programmes de sélection, la résistance à la sécheresse parmi les caractères souhaitables. La sécheresse existe aussi dans les sables profonds des régions à précipitations plus fortes. La sélection en vue d'augmenter la résistance à la sécheresse a été couronnée de succès, comme on l'a observé chez les descendants d'arbres sélectionnés dont la survivance était meilleure que celle des témoins (van Buijtenen, 1966; Gilmore, 1957) sans aucune diminution de croissance en hauteur (Goddard et Brown, 1959, Banks, 1966).
Résistance aux insectes nuisibles et aux maladies. Dans les travaux d'amélioration du pin à l'encens, il est important de tenir compte de la résistance aux insectes et aux maladies, en raison des effets que peuvent avoir ces agents sur la production de graines, sur la croissance des arbres attaqués, ainsi que sur le rendement à l'hectare et la qualité des produits tirés du bois. On a pu observer des différences notables d'une race à l'autre et aussi d'un arbre à l'autre dans la réceptivité à l'égard d'une maladie déterminée, et des études sur d'autres ravageurs sont en cours.
Le principal ravageur du pin à l'encens est la rouille fusiforme (Cronartium fusiforme Hedgc. & Hunt ex Cumm.). Elle fait des ravages sur des centaines de milliers d'hectares - certaines zones sont infestées à plus de 90 pour cent - dans une large ceinture qui s'étend vers l'Ouest de la Caroline du Sud et la Géorgie à la Louisiane (Roth et McComb, 1971).
Les essais de descendance des clones dans les vergers à graines ont démontré que la résistance à la rouille des familles qui en sont issues est très variable, phénomène dont on doit absolument tenir compte dans les projets de sélection expérimentale (Woessner, 1965). Dans une étude de grande envergure sur l'héritabilité avec des arbres-mères pris dans une population à l'état sauvage, Kinloch et Stonecypher (1969) ont trouvé des valeurs de 0,65 à 0,85 pour l'héritabilité de la résistance à la rouille; des variations d'intensité de l'infection parmi les familles allant de 17,3 à 100 pour cent; une corrélation positive entre arbres-mères et descendants en ce qui concerne la résistance à la rouille. Les variations de cet agent pathogène avec la latitude et la longitude ont été illustrées (Snow, Kais et Dinus, 1972) et des formes albino ont été décrites (Kais, 1966). On ne discerne pas encore très clairement l'importance des races de rouille, mais les études actuellement en cours seront certainement très utiles.
La réceptivité à la rouille fusiforme est un bon exemple de variation géographique. Chez le pin à l'encens, l'incidence de l'infestation par la rouille fusiforme est imputable à des facteurs tant génétiques que de milieu (Wells et Wakeley, 1966). Les pins à l'encens de la Caroline du Nord ne sont que peu atteints par la rouille, mais le taux d'infestation est élevé chez les arbres originaires de cette région et plantés au centre de la Géorgie, région où ce pathogène est très répandu. On a aussi observé qu'il y a peu d'infestation par la rouille chez les pins à l'encens du Texas oriental, mais que l'attaque demeure faible lorsque les arbres texans sont plantés dans le centre de la Géorgie. Il se peut que le résultat le plus important que l'on ait obtenu concernant les travaux sur la génétique dans l'aire méridionale du pin soit la découverte du degré de résistance de quelques arbres-mères à cette maladie. On a établi des vergers à graines provenant de sujets particulièrement résistants pour mettre à profit les premières connaissances obtenues (Blair, 1970; Zobel, Blair et Zoerb, 1971).
La pyrale des pousses du pin de Nantucket (Rhyacionia frustrana Comst.) est un insecte nuisible commun, mais il y a apparemment peu de variation intraspécifique, si tant est qu'il y en ait, en ce qui concerne la résistance à l'attaque par cet insecte chez les pins à l'encens. L'étude de la résistance aux bostryches est actuellement en cours.
Associations de caractères. La nature et l'ampleur des variations d'un pin à l'encens à un autre diffèrent beaucoup, mais il y a peu de cas de corrélations statistiques étroites parmi les divers caractères. Ainsi, la sélection d'arbres d'élite devrait-elle être basée sur un grand nombre de caractères, par exemple par l'application d'un système de coefficients, de manière à assurer une bonne combinaison de caractères importants du point de vue sylvicole. La sélection de caractères multiples peut permettre d'éviter qu'un caractère indésirable n'ait pour effet de contrebalancer plusieurs caractères recherchés, comme cela peut être le cas avec la méthode de sélection en tandem. Il est cependant exact qu'il existe chez le pin à l'encens de nombreux liens entre les caractères, de même que chez d'autres pins du sud, comme par exemple l'accroissement du diamètre du tronc, de la longueur des branches et de la longueur de la cime en fonction de l'augmentation de l'espace de croissance. Il y a cependant des exceptions à cela et on doit arriver à trouver des arbres qui pourront servir dans les travaux de sélection et d'amélioration et qui ont un taux de croissance élevé et de nombreux autres caractères souhaitables concernant la forme de la cime et de la tige, la production de graines, la résistance aux maladies et la qualité du bois.
Hybrides naturels et artificiels
Les principaux pins du sud et certaines autres espèces de moindre importance s'hybrident naturellement avec le pin à l'encens. L'hybridation contribue à créer des variations parmi les régions géographiques et parmi les arbres à l'intérieur d'un même peuplement. Le nombre de caractères et le degré d'influence que l'hybridation peut avoir sur ceux-ci sont importants du point de vue économique. L'hybridation compte donc parmi les facteurs importants dans la sélection de races géographiques ou d'arbres particuliers soit pour être plantés, soit pour être utilisés dans des projets d'amélioration.
La présence d'au moins trois générations d'hybridation de pin longleaf et de pin à l'encens avec production de formes intermédiaires a été vérifiée dans un peuplement naturel dans l'Etat de la Louisiane (Namkoong, 1966). L'existence d'arbres intermédiaires entre le pin à l'encens et le shortleaf (Pinus echinata Mill.) quant aux caractères morphologiques (Zobel, 1953) et le module électrophorétique des protéines de la graine (Hare et Switzer, 1969) indique que cet hybride se rencontre au Texas où il y a peu d'infestation par la rouille fusiforme. On a trouvé, dans deux localités différentes de l'Etat du Mississippi, des arbres ressemblant aux hybrides artificiels de pins à l'encens et pins shortleaf (Mergen, Stairs et Snyder, 1965). Dans la partie nord-est de l'habitat, on trouve des hybrides de pin à l'encens avec Pinus serotina Michx. et P. rigida Mill. (Little, Little et Doolittle, 1967; Smouse, 1970).
On trouve de nombreux caractères morphologiques intermédiaires chez les hybrides artificiels dans lesquels entre le pin à l'encens (Little et Righter, 1965, Little et al., 1967; Schmitt, 1968), mais la production de graines par cône peut être extrêmememt faible et la résistance à la rouille fusiforme élevée. Les hybrides avec le pin longleaf ont une importance commerciale en Corée parce que la forme est meilleure que celle du longleaf et la résistance au froid supérieure à celle du pin à l'encens (Hyun, 1970). En Corée, on préfère le pollen des pins à l'encens de source nordique.
Mutations
Le pin à l'encens est normalement diploïde, mais il arrive que l'on trouve quelquefois des plants mixoploïdes et polyploïdes dans les planches à semis de pépinières. Les plantes aberrantes ont peu de vigueur et ne parviennent pas à maturité; elles sont donc rares dans les peuplements. On a trouvé 22 formes de mutants dissemblantes du point de vue phénotypique dans 30 familles sur 119 de pins à l'encens auto-fécondées (Franklin, 1969). D'une manière générale, les mutants ne sont pas suffisamment intéressants pour qu'on cherche à les produire, ni assez répandus pour qu'on cherche à les éviter dans les programmes de sélection.
Le pin à l'encens en tant qu'essence exotique
Malgré sa mauvaise réputation antérieure, due à sa performance médiocre lorsqu'on employait des semences de source erronée, le pin à l'encens est actuellement en voie d'être très largement introduit dans un grand nombre de pays. Son importance s'est accrue dans certaines régions qui lui conviennent comme le Transvaal en Afrique du Sud, la Nouvelle Galles du Sud (Australie) ou encore aux hautes altitudes et à la latitude du Brésil et des régions adjacentes de l'Argentine.
Pour réussir l'introduction du pin à l'encens, il faut soigneusement choisir la source de la semence. Par exemple, pour les régions chaudes et qui sont essentiellement des zones sans gelées auxquelles le pin à l'encens est adapté, la meilleure croissance est obtenue avec la source Floride, suivie de près par la Livingston Parish, la Louisiane et peut-être aussi le pin à l'encens de la Géorgie du Sud. Les différences du taux de croissance entre la source Floride et les sources plus septentrionales du Piedmont, dans des régions où il gèle peu, peuvent aller du simple au double. Dans les zones plus froides, les sources de la région du littoral atlantique comme le comté d'Onslow en Caroline du Nord viennent bien, tandis que dans les climats réellement froids, ce sont les sources des hautes altitudes du Tennessee ou des hautes latitudes du Maryland qui peuvent réussir. D'une manière générale, les sources les plus méridionales de pin à l'encens viennent le mieux dans les régions où il ne gèle jamais ou presque jamais, et ces sources donnent lieu à une croissance plus rapide que celle des sources nordiques ou du Piedmont. Il faut bien distinguer entre les sources de la Plaine littorale plus humide et le pin à l'encens qui pousse naturellement dans l'intérieur du Piedmont et dans les régions des dunes. On trouve parfois le pin à l'encens dans le nord de la Géorgie et de l'Alabama à une altitude d'environ 900 m., mais on le rencontre principalement au-dessous de 300 m. d'altitude. Les sources des hautes altitudes et latitudes septentrionales sont beaucoup plus résistantes au froid que les sources méridionales et il leur faut des nuits fraíches pour croître convenablement. Quelques-unes des sources occidentales, comme celles de la région de Lost Pines au Texas, supportent mieux la sécheresse que les sources plus orientales; la source texane est aussi plus résistante aux maladies.
Subdivisions d'intérêt pratique
Il y a de nombreuses différences entre le pin à l'encens de source Piedmont et celui de la Plaine littorale. La zone du Piedmont occupe les contreforts qui surplombent la plaine bordant la côte; les températures y sont plus basses, les précipitations ordinairement plus faibles et les printemps secs; les sols sont argileux ou de limon argileux, tandis que dans la Plaine littorale, l'argile est souvent recouverte de sable. Il y a une zone étroite entre le Piedmont et la Plaine littorale qu'on appelle “les Dunes” où la terre est aride et profonde et où le pin à l'encens ne s'adapte pas bien. D'autres subdivisions sont également faites entre la Plaine littorale septentrionale et la Plaine littorale méridionale le long de la côte atlantique. Aussi, dans les programmes d'amélioration des arbres, a-t-on installé des vergers à graines séparés pour chacune des régions du Piedmont, de la Plaine littorale septentrionale et de la Plaine littorale méridionale.
Des distinctions similaires sont faites plus à l'ouest le long de la côte du golfe du Mexique où les subdivisions ont pour nom: Plaine littorale inférieure, Plaine littorale supérieure et quelquefois une zone dénommée Piedmont ou contreforts. On trouve aussi dans cette région une zone de dunes appelée Fall Line. Il y a quelques différences évidentes entre les provenances du Piedmont (et la plaine littorale supérieure) et celles de la plaine littorale. Les pins à l'encens de la provenance de la plaine littorale croissent rapidement tant dans les stations de la plaine côtière que dans celles du Piedmont, mais ils périssent dans les années de sécheresse anormale ou lorsqu'il se produit des gelées sévères et intempestives, tandis que les provenances du Piedmont survivent. Les recherches effectuées sur l'épaisseur de l'écorce indiquent que pour la provenance de la plaine littorale l'écorce des jeunes arbres est plus mince que chez les sujets de la provenance du Piedmont (25 pour cent contre 33 pour cent chez les arbres âgés de dix ans). Il reste à savoir si cette différence persiste chez les arbres arrivés à l'âge d'exploitation pour lesquels la moyenne s'établit approximativement entre 12 et 16 pour cent. La production de graines a lieu plus tôt et est plus abondante chez la provenance du Piedmont que chez celle de la plaine littorale, même lorsque l'une et l'autre sont plantées dans la même station. On a vu dans certains cas un verger à graines d'une provenance du Piedmont produire jusqu'à 8 ou 10 fois plus de graines qu'un verger d'une provenance de la plaine littorale du même âge et ayant été soumis à un traitement similaire. Il est essentiel de connaître l'existence de pareilles différences, lorsqu'on projette d'installer un verger à graines, pour lui donner la dimension voulue.
Une ébauche de définition, au sens large du mot, des provenances de pin à l'encens susceptibles d'être utiles, pourrait comprendre:
Le centre nord de la Floride, qui englobe la zone de marnes calcaires avoisinant la côte qu'on désigne sous le nom de Gulf Hanmock et les sols plus profonds du comté de Marion. Cette source convient aux conditions subtropicales; croissance très rapide; bois de poids spécifique élevé. Elle est très sensible au froid et ne peut être utilisée que dans les régions où il ne gèle pas et où le froid n'est pas rigoureux. Elle ne résiste pas à la sécheresse et succombe assez facilement. La source du nord de la Floride est la provenance de pin à l'encens dont la croissance est la plus rapide parmi celles qui ont été mises à l'essai dans un certain nombre de régions comme le Queensland, quelques parties du Brésil et de l'Afrique du Sud. Cette source a tendance à avoir des tiges plus sinueuses et des feuilles plus larges que les sources plus septentrionales. Dans son aire naturelle, cette source n'est qu'un médiocre producteur de graines, bien que la production de certains clones plantés dans des vergers à graines soit passablement bonne mais tarde à débuter. Il semble que dans les conditions climatique du sud-est des Etats-Unis, la production massive de graines du pin à l'encens de Floride soit problématique.
Le Livingston Parish, Louisiane et ses environs. Le pin à l'encens de cette région semble représenter une source de grande valeur dont la performance, dans l'étude effectuée sur un grand nombre de sources méridionales, a été excellente, comme l'a signalé O.O. Wells (1969). Il a un bon taux de croissance, il est d'un bon port et il semble avoir une floraison convenable. Bien qu'on ne dispose encore que peu de données directes sur des sujets arrivés à l'âge souhaité, nous avons l'impression que la source Livingston Parish n'a pas une croissance aussi rapide que le pin à l'encens de Floride. Il semble également que la source Livingston Parish soit unique en son genre pour la résistance à la rouille fusiforme, même lorsqu'elle est plantée dans les régions les plus infestées. Elle fait actuellement l'objet d'essais dans un certain nombre de régions du sud-est des Etats-Unis, mais ces essais sont encore trop récents pour permettre de déterminer sa réaction au froid, à la sécheresse ou aux sols argileux secs du Piedmont qui succède à la zone côtière. Nous sommes d'avis que c'est une source qui doit obligatoirement être testée pour la plantation dans des zones subtropicales ou des régions où les gelées sont légères.
Les autres sources de la Plaine littorale. Les arbres de la Géorgie méridionale ou de la Plaine littorale méridionale de la Caroline du Sud poussent très bien et peuvent être, dans beaucoup de cas, aussi bons que les sources (a) et (b) mentionnées ci-dessus. Pour en tirer le meilleur profit, il est nécessaire de choisir avec soin les arbres-mères aussi bien que les peuplements. Des essais effectués sur des arbres du comté d'Onslow en Caroline du Nord ont donné de bons résultats; à notre connaissance, les bons arbres des bons peuplements des zones côtières des Carolines et de la Géorgie viennent bien dans les conditions des régions subtropicales. Bien que leur croissance ne soit pas aussi rapide que celle de la source Floride, ces sources ont une faculté d'adaptation et de résistance beaucoup plus grande.
Le pin à l'encens du Piedmont et de la Plaine littorale côtière supérieure (sources de l'intérieur). Ce groupe comprend une vaste région située en deçà des plaines côtières du golfe du Mexique et de l'Atlantique (Fig. 1). Les arbres de ces régions intérieures doivent faire l'objet d'essais là où les conditions climatiques, sécheresse ou gel, sont rigoureuses. Bien que leur croissance soit plus lente que celle de certains pins à l'encens de source côtière, les sources de l'intérieur devraient être utilisées dans les tentatives d'extension de l'aire de plantation du pin à l'encens. Bien que, d'autre part, les sources du Piedmont et de la Plaine côtière supérieure soient rangées dans le même groupe, il est certain qu'il existe des différences entre elles et les décisions quant à l'emploi de matériel génétique de Virginie, de Caroline du Nord, de Caroline du Sud, de Géorgie, d'Alabama ou de régions situées plus à l'ouest, dépendent de la rigueur de l'environnement considéré. Des quantités considérables de semences génétiquement améliorées sont disponibles dans les vergers à graines de cette région.
La source de la côte orientale. En Virginie et dans le Delaware, le pin à l'encens pousse sur une péninsule basse aux sols sableux et quelquefois argileux et humides. Cette source n'a pas été testée, mais il semble qu'elle s'accommode bien d'une salinité pas trop forte. Les peuplements naturels ont un bon port et produisent un cubage important par unité de superficie.
Les sources septentrionales ou des hautes altitudes. Le pin à l'encens ne pousse ordinairement pas au-dessus de 350m d'altitude, mais on trouve parfois des peuplements à des altitudes voisines de 1 000 m. Ces arbres ont fait l'objet de tests et l'on a constaté qu'ils étaient plus résistants à la glace et à la neige que le pin à l'encens normal, mais leur croissance est moins rapide que celle des sources côtières. Le pin à l'encens des hautes altitudes conviendrait bien aux zones à températures extrêmes des hautes terres des régions subtropicales où les pins subtropicaux ne viennent pas bien. Les sources de pin à l'encens des latitudes plus septentrionales, comme celles du Maryland et du Delaware, supportent également très bien le froid, mais leur croissance a été décevante.
Les stations excessivement humides ou excessivement sèches. Le pin à l'encens est devenu manifestement tolérant aux degrés extrêmes d'humidité. Par exemple, les pins résistants à la sécheresse originaires de l'ouest, comme la région de Lost Pine dans le Texas (comté de Bastrop) ont été testés à grande échelle. Ils supportent très bien les sécheresses extrêmes et poussent et survivent tout aussi bien dans les environnements chauds et secs. Le choix des arbres-mères joue un rôle capital, certains essais ayant produit des arbres excessivement branchus ou tordus. Il s'est avéré que la source résistante à la sécheresse pourrait considérablement étendre l'aire où le pin à l'encens peut avoir une certaine valeur.
Certaines populations de pin à l'encens poussent dans une tourbe profonde, qui atteint parfois 7 m. avant d'arriver au sol minéral. C'est à cause de leur système radiculaire unique que ces pins se maintiennent et viennent bien dans cet environnement. Les essais auxquels ils ont été soumis laissent entrevoir de bonnes chances de réussite dans des conditions d'humidité excessive à condition que l'eau ne soit pas stagnante. On a établi des vergers à graines de ces pins à l'encens “tolérants à l'eau” principalement pour les régions côtières des Carolines du Nord et du Sud.
RESUME
Le pin à l'encens est une espèce remarquable qui pousse dans de nombreux environnements différents et qui a une grande capacité d'adaptation. Il a une grande amplitude de variation et s'hybride naturellement avec un certain nombre d'autres espèces de pins. Il réagit bien aux manipulations génétiques et de grands progrès ont été réalisés dans l'amélioration de cette espèce.
Il n'a pas été assez judicieusement employé comme essence introduite. En raison de sa grande capacité d'adaptation, le choix de la source géographique correcte est un facteur essentiel pour la réussite de l'introduction. Ce fait a été ignoré lors de nombreux essais effectués et la performance médiocre qui en a résulté a incité certains forestiers à le considérer comme une espèce peu désirable. C'est un arbre à croissance rapide, donnant du bon bois, mais qui a un port médiocre. Il en résulte que la sélection du pied-mère est indispensable.
Les sources du sud du pays sont particulièrement précieuses et ont une croissance rapide sous les climats subtropicaux. Le bois du pin à l'encens est généralement d'une qualité satisfaisante quel que soit l'environnement dans lequel il pousse. Pour une essence exotique, il est en général résistant aux maladies et aux insectes. Enfin, ce pin est appelé à connaître une utilisation plus large, surtout depuis que l'on commence à disposer de matériel génétique amélioré.
REFERENCES BIBLIOGRAPHIQUES
Banks, P.F. 1966. Early height growth in Rhodesia of progenies of P. taeda from Texas. Rhod. Zambia Mal. Jour. Agri. Res. 4(1): 3–7
Barber, J.C. 1966. Variation among half-sib families from three loblolly pine stands in Georgia. Ga. For. Res. Council Res. Pap. 37. 5 pp.
Blair, R.L. 1970. Quantitative inheritance of resistance to fusiform rust in loblolly pine. Ph. D. Thesis, N.C. State Univ. 87 pp.
Crow, A.B. 1964. Ten-year results from a local geographic seed source study of loblolly pine in south-eastern Louisiana. La. State Univ. and A & M Coll., Sch. For. & Wildl. Manage., L.S.U. For. Note 57. 4 pp.
Dorman, K.W. and B.J. Zobel. 1973. Genetics of loblolly pine. USDA For. Ser. Res. Pap. W.O. 19. 21 pp.
Einspahr, D.W., J.R. Peckham and M.C. Mathes. 1964. Base lines for judging wood quality of loblolly pine. For. Sci. 10: 165–173.
Einspahr, D.W., J.P. van Buijtenen and J.R. Peckham. 1969. Pulping characteristics of ten-year loblolly pine selected for extreme wood specific gravity. Sil. Gen. 18(3): 57–61.
Franklin, E.C. 1969. Mutant forms found by self-pollination of loblolly pine. Jour. Heredity 60(6): 315–320.
Gilmore, A.R. 1957. Physical and chemical characteristics of loblolly pine seedlings associated with drought resistance. Fourth South. Conf. For. Tree Improv. Proc. 1957:34–39.
Goddard, R.E. and C.L. Brown. 1959. Growth of drought-resistant loblolly pines. Tex. For. Ser. Res. Note 23. 7 pp.
Goddard, R.E., C.L. Brown and T.E. Campbell. 1959. An evaluation of growth and form in 5-year-old open-pollinated progeny from selected loblolly pine. Fifth South. Conf. For. Tree Improv. Proc. 1959: 35–43.
Goddard, R.E. and R.K. Strickland. 1964. Crooked stem form in loblolly pine. Sil. Gen. 13(5):155–157.
Goggans, J.F., K.D. Lynch and G.I. Garin. 1972. Early results of a loblolly pine seed source study in Alabama. Auburn Univ., Agri. Expt. Sta. Cir. 194. 19 pp.
Grigsby, H.C. 1973. A test of 36 geographic sources of loblolly pine seed in southern Arkansas. USDA For. Ser. Res. Pap. SO. In press.
Hare, R.C. and G.L. Switzer. 1969. Introgression with shortleaf pine may explain rust resistance in western loblolly pine. USDA For. Ser. Res. Note SO-88. 2 pp.
Hocker, H.W., Jr. 1955. Climatological summaries for selected stations in or near the southern pine region, 1921 – 1950. USDA For. Ser. Southeast. For. Expt. Sta. Pap. 56. 11 pp.
Hyun, S.K. 1970. The growth performance of pitch-loblolly hybrid pine produced by different geographic races of loblolly pine in their early age. FAO/IUFRO Second World Consult. For. Tree Breed. 1969:803–814. FAO FO-FTB-69 7/7.
Kais, A.G. 1966. Persistence of albinism in Cronartium fusiforme. USDA Plant Dis. Rep. 50:842.
Kinloch, B.B., Jr. and R.W. Stonecypher. 1969. Genetic variation in susceptibility to fusiform rust in seedlings from a wild population of loblolly pine. Phytopathology 59(9): 1246–1255.
Kraus, J.F. 1967. A study of racial variation in loblolly pine in Georgia — tenth-year results. Ninth South. Conf. For. Tree Improv. Proc. 1967:78–85
Lantz, C.W. and J.G. Hofmann. 1969. Geographic variation in growth and wood quality of loblolly pine in North Carolina. Tenth. South. Conf. For. Tree Improv. Proc. 1969:175–188.
Little, E.L., Jr., S. Little and W.T. Doolittle. 1967. Natural hybrids among pond, loblolly and pitch pines. USDA For. Ser. Res. Pap. NE-67. 22 pp.
Little, E.L., Jr. and F.I. Righter. 1965. Botanical descriptions of forty artificial pine hybrids. USDA For. Ser. Tech. Bull. 1345. 47 pp.
Mergen, F., G.R. Stairs and E.B. Snyder. 1965. Natural and controlled loblolly-x-shortleaf pine hybrids in Mississippi. For. Sci. 11(3):306–314.
Merrifield, R.G. 1965. Loblolly pine seed sources in north Louisiana. North La. Hill Farm Expt. Sta. For., Hill Farm Facts 6:4.
Namkoong, G. 1966. Statistical analysis of introgression. Biometrics 22(3):488–502.
Perry, T.O. 1960. The inheritance of crooked stem form in loblolly pine (Pinus taeda L.). Jour. For. 58(12):943–947.
Roth, E.R. and W.H. McComb. 1971. Georgia fusiform rust survey 1971. USDA For. Ser., Southeastern Area and Georgia Forestry Commission. 4 pp.
Schmitt, D. 1968. Performance of southern pine hybrids in south Mississippi. USDA For. Ser. Res. Pap. SO-36. 15 pp.
Smouse, P.E. 1970. Population studies in the genus Pinus L. Ph. D. Thesis, N.C. State Univ. 126 pp.
Snow, G.A., A.G. Kais and R.J. Dinus. 1972. Further evidence of geographical variation in Cronartium fusiforme. (Abst.) Phytopathology 62:790.
Snyder, E.B. and A.E. Squillace. 1966. Cone and seed yields from controlled breeding of southern pines. USDA For. Ser. Res. Pap. SO-22. 7 pp.
Trousdell, K.B., K.W. Dorman and A.E. Squillace. 1963. Inheritance of branch length in young loblolly pine progeny. USDA For. Ser. Res. Note SE-1. 2 pp.
USDA Forest Products Laboratory. 1965. Southern wood density survey — 1965 status report. USDA For. Ser. Res. Pap. FPL-26. 38 pp.
van Buijtenen, J.P. 1963. Inheritance of wood properties and their relation to growth rate in Pinus taeda. FAO World Consult. For. Genet. Tree Breed. Proc. 1963. FAO Forgen 63/ -7/2.
van Buijtenen, J.P. 1966. Testing loblolly pines (Pinus taeda) for drought resistance. Tex. For. Ser. Tech. Rept. 13. 15 pp.
van Buijtenen, J.P. 1969. Seventeenth progress report of the cooperative tree improvement program. Tex. For. Ser. Cir. 200. 14 pp.
Wakeley, P.C. 1961. Results of the Southwide pine seed source study through 1960–61. Sixth South. Conf. For. Tree Improv. Proc. 1961:10–24.
Wells, O.O. 1969. Results of the Southwide pine seed source study through 1968–69. Tenth South. Conf. For. Tree Improv. Proc. 1969:117–129.
Wells, O.O. and G.L. Szitwer. 1971. Variation in rust resistance in Mississippi loblolly pine. Eleventh South. Conf. For. Tree Improv. Proc. 1971:25–30.
Wells, O.O. and P.C. Wakeley. 1966. Geographic variation in survival, growth and fusiform rust infection of planted loblolly pine. Soc. Am. For., For. Sci. Mongr. 11. 40 pp.
Whitesell, C.D., B.J. Zobel and J. Roberds. 1966. Specific gravity and tracheid length of loblolly pine in Maryland and Delaware. N.C. State Univ., School For. Tech. Rept. 29. 11 pp.
Woessner, R.A. 1965. Growth, form and disease resistance in four-year-old control- and five-year-old open-pollinated progeny of loblolly pine selected for use in seed orchards. N.C. State Univ. Sch. For. Tech. Rept. 28. 67 pp.
Zobel, B.J. 1953. Are there natural loblolly-shortleaf pine hybrids? Jour. For. 51(7):494–495.
Zobel, B.J. 1961. Inheritance of wood properties in conifers. Sil. Gen. 10(3):65–70.
Zobel, B.J. 1964. Breeding for wood properties in forest trees. Unasylva 18(2/3):89–103.
Zobel, B.J., R. Blair and M. Zoerb. 1971. Using research data — disease resistance. Jour. For. 68(8): 486–489.
Zobel, B.J., R.C. Kellison and M. Matthias. 1969. Genetic improvement in forest trees — growth rate and wood characteristics in young loblolly pine. Tenth South. Conf. For. Tree Improv. Proc. 1969:59–75.
Zobel, B.J. and R.L. McElwee. 1958. Variation of cellulose in loblolly pine. Tappi 41(4): 167–170.
Zobel, B.J., M. Matthias, J.H. Roberds and R.C. Kellison. 1967. Moisture content of southern pine trees. Fourth. For. Biol. Conf. Proc. 1967:11–35.
Zobel, B.J. and R.R. Rhodes. 1955. Relationship of wood specific gravity in loblolly pine to growth and environmental factors. Tex. For. Ser. Tech. Rept. 11. 32 pp.
Zobel, B.J., E. Thorbjornsen and F. Henson. 1960. Geographic, site and individual tree variation in wood properties of loblolly pine. Sil. Gen. 9(6):149–158.
![]()
![]()
![]()