![]()
![]()
![]()
OBSERVACIONES GENERALES
El pino tea (Pinus taeda L.) se extiende muchísimo por el sur y el sudeste de los Estados Unidos. De gran adaptabilidad, buena madera y ordenación fácil, por su extensa distribución nativa es genéticamente complejo y consta de varias razas geográficas. Hibridiza tan fácilmente con otras especies, que cuesta trabajo decir cuál es tea puro. Sin embargo, parece ser siempre la especie “dominante”.
Dada su amplia variabilidad geográfica, el pino tea no se ha utilizado nunca como especie exótica con el éxito y la amplitud que merece. Muchas plantaciones fracasaron porque en los ensayos se empleó semilla de origen desconocido, además de haberla sembrado en medios a los que no se adaptaba. De ahí que en muchas zonas subtropicales se crea erróneamente que el pino antillano (P. elliottii), de mucha más adaptación, es mejor, e inferior el tea.
No siempre ha sido sencillo obtener fuentes apropiadas de pino tea, pero el Centro de semillas recientemente organizado por el Servicio Forestal de los Estados Unidos es de muchísima ayuda 3. Según se vaya disponiendo de semilla original genéticamente mejorada y así que reconozcamos y utilicemos su diversidad geográfica, este pino sera cada vez más apreciado como especie exótica. En este trabajo se exponen, primero, algunas características importantes del pino tea y, después, consideraciones a tener en cuenta al utilizarlo como exótico.
AREA DE DISPERSION
El área de dispersión del pino tea viene a formar un rectángulo en sentido E.-O. en el sudeste de los Estados Unidos. Pero a lo largo de la costa atlántica hay extensiones nordorientales y, hacia el S., hasta Florida (fig. 1). No se da en la zona ribereña del Mississippí. Expondremos parajes de fuentes semilleras con su grado distintivo de productividad, de las que, entre otras varias, se tratará más adelante. En la figura 1 se muestran regiones fisiográficas de tanta importancia para nuestra exposición como pueden serlo la llanura costera, fall line + o Sandhills (“Las dunas”) y Piedmont (“a pie de monte”). Damos también (fig. 2) climógrafos que pueden utilizarse, con la latitud y la longitud, para la estimación de los homoclimas cuando el pino tea se emplee como exótico. En la Florida las precipitaciones son elevadas en épocas de altas temperaturas; o sea, lo contrario que en Texas, que con más calor llueve menos. Las letras E, A y O representan en cada gráfica los meses de enero, agosto y octubre. Los datos proceden de resúmenes climatológicos pormenorizados que figuran en una separata (Hocker, 1955).
1 Profesor de Genética Forestal, Universidad de Carolina del Norte, Raleigh.
Figura 1: Area botánica del pino tea y sus principales regiones fisiográficas.
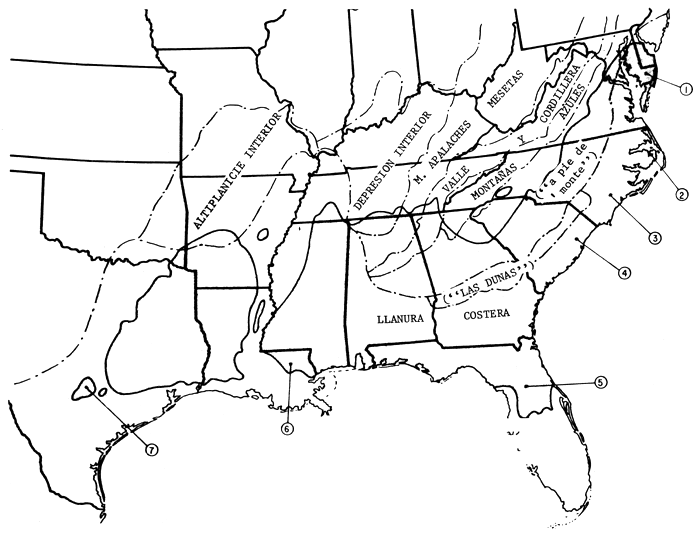
KEY TO SEED SOURCES
1 - MARYLAND
2 - DARE COUNTY, CAROLINA DEL NORTE
3 - ONSLOW COUNTY CAROLINA DEL NORTE
4 - GEORGETOWN COUNTY, CAROLINA DEL SUR
5 - MARION COUNTY, FLORIDA
6 - LIVINGSTON PARISH, LOUISIANA
7 - “PINOS PERDIDOS”, TEXAS
SUBSTITUYE LAS PAGINAS 4 Y 5
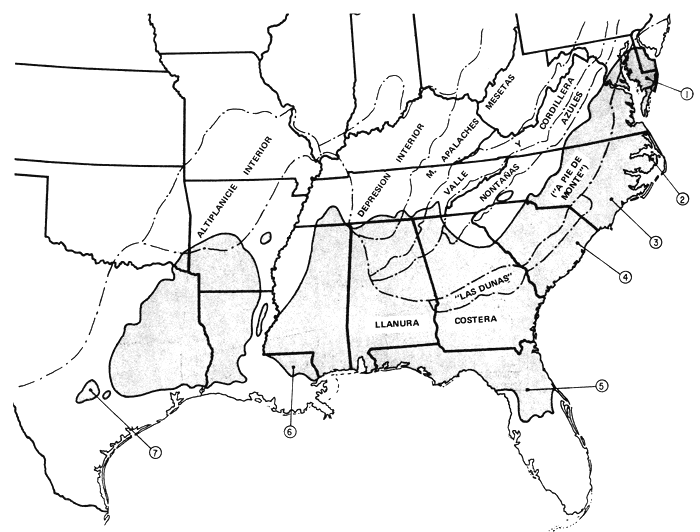
LLAVE DE LAS FUENTES DE SEMILLAS
1 - MARYLAND
2 - DARE COUNTY, CAROLINA DEL NORTE
3 - ONSLOW COUNTY, CAROLINA DEL NORTE
4 - GEORGETOWN COUNTY, CAROLINA DEL SUR
5 - MARION COUNTY, FLORIDA
6 - LIVINGSTON PARISH, LOUISIANA
7 - “PINOS PERDIDOS”, TEXAS
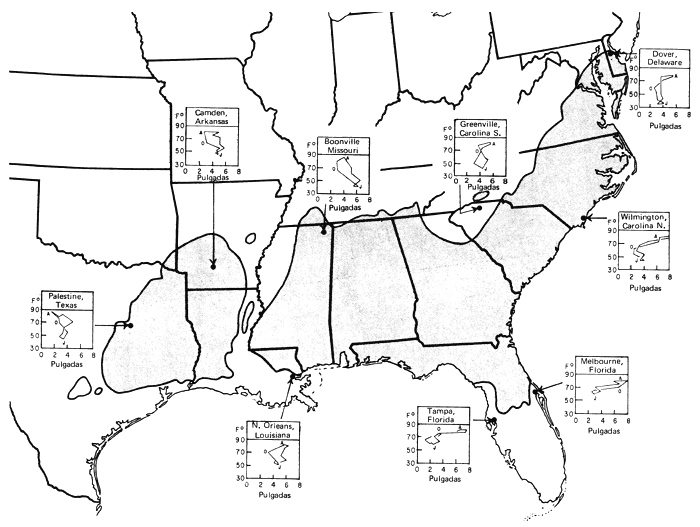
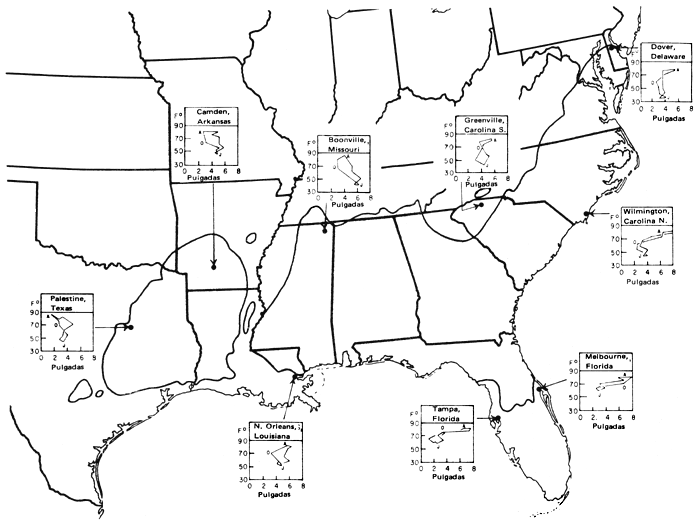
Figura 2 - Area de tipos climáticos en toda la región del pino tea (según W.G. Wahlenberg y C.E. Ostrom, en “Geographie variation in climate in the Loblolly pine region”. Res. Note S.E. For Exp. Sta. No. 94 (1956)
CARACTERISTICAS DEL PINO TEA
La magnitud y clase de sus variaciones tienen la importancia suficiente para que este pino desempeñe un papel relevante en todo programa de mejora de árboles que entrañe la introducción de semilla, la selección racial y la de árboles plus, o hibridación. Se sabe más de esta especie que de cualquier otro pino del sur de los Estados Unidos, pero la información sobre su variación es más completa que la que haya concretamente sobre cada uno de sus caracteres. En cuanto vamos a exponer seguidamente figuran sólo pocas referencias para cada carácter o tema, si bien hay más detalles informativos en un trabajo más extenso: The Genetics of Loblolly Pine (Dorman y Zobel, 1973).
Variación y herencia
Porcentaje de crecimiento
El extenso estudio de áreas semilleras, a sus 5, 10 y 15 años, patrocinado por el Comité sobre la mejora de especies forestales, reveló profundas variaciones clinales en crecimiento, con árboles de zonas costeras que, por lo general, medran más de prisa que todos los del interior, salvo si se trata de localidades muy frías (Wakeley, 1961; Wells y Wakeley, 1966, y Wells, 1969). Trasladados de uno a otro extremo de su habitat, produjeron menos de la mitad del volumen alcanzado por árboles de semilla local, debido a una doble circunstancia: supervivencia mediocre y crecimiento lento; sin embargo, al llevar árboles de origen costero hacia el interior, hasta 225 millas (unos 360 kilómetros) al sur de su área, crecieron con mayor rapidez que los continentales.
Cuatro de las quince fuentes de pino tea comprendidas en este vasto estudio tenían muchísima menos roya fusiforme que las otras. Estos árboles, resistentes hasta cierto punto, eran los originarios del oeste del río Mississippí, crecidos en Livingston Parish (Louisiana) y en la costa oriental de Maryland. Otro estudio igualmente amplio y que abarca 36 portagranos crecidos en Crossett (Ashley County), Arkansas, revela también el rápido crecimiento que presentan en potencia los pinos tea de los litorales, especialmente los de ambos estados de Carolina (Grigsby, 1973).
Los resultados de estos extensos estudios han sido corroborados y ampliados por otros, con mucha recolección de muestra de pino tea en Georgia (Kraus, 1967), Alabama (Goggans, Lynch y Garin, 1972), Mississippí (Wells y Switzer, 1971) y Carolina del Norte (Lantz y Hofmann, 1969). Un estudio de varias poblaciones de pinos de Louisiana que se plantaron por doquier en el mismo estado revelaba que los de Livingston Parish y los del oeste de Louisiana eran resistentes a la roya, pero las diferencias de altura entre los del norte y los del sur, si bien interesantes, estadísticamente, en cambio, no lo eran (Crow, 1964; Merrifield, 1965).
Estudios hechos dentro de los límites del estado han permitido determinar que, además de las estructuras de variación establecidas por los estudios más amplios, el pino tea varía clinalmente en supervivencia, en crecimiento potencial - hasta cierto punto - y en resistencia a la roya fusiforme. Los de las costas del Golfo y del Atlántico Sur sobreviven algo menos y tienen menor resistencia a la roya que los pinos de las zonas interiores.
Las diferencias inherentes en crecimiento entre árboles padres individuales son grandes, como también lo son las que se observan entre una y otra fuente geográfica. En Texas, progenies de padres de pino tea vienen a crecer en zonas semilleras casi al mismo promedio que los testigos, y aunque esas progenies eran más uniformes, entre las familias había amplias diferencias. (Van Buijtenen, 1969). Las diez familias de producción más elevada dieron 1,82 cuerdas por acre y año; los testigos, 1,54 cuerdas, y el rendimiento de las diez familias menos productoras fue de 1,14 cuerdas +. El crecimiento más bajo de una familia arrojó 0,90 cuerdas por acre y año, y el más alto, 2,05. En familias procedentes de ciertos árboles padres por polinización libre, seleccionadas para huertos industriales de semillas, hay grandes diferencias de rendimiento en leña por acre. (Zobel, Kellison y Matthias, 1969). Se han registrado rendimientos que variaban de 1,8 a 2,7 toneladas de leña por acre y año entre familias de siete años sobre terreno excelente. En general, se espera que el crecimiento de progenies procedentes de huertos de semillas de clones indeseables sea del 10 al 20 por 100 superior al de las plantas destinadas a plantaciones comerciales. Cuando las progenies de once árboles elegidos en el sudeste, centro y noroeste de Georgia se plantaron en la región central de aquel mismo estado, la familia de crecimiento más lento en un 17 a 31 por ciento al cabo de cinco períodos vegetativos (Barber, 1966). La familia de crecimiento más lento, procedente de semilla también más lenta en crecer, daba como promedio 8,4 pies; en cambio, la de crecimiento más rápido, de semilla asimismo más rápida, 12,0 pies (casi el 43 por 100 más en altura), prueba de que los efectos de la selección racial y de las selecciones de árboles plus pueden ser acumulativos.
Copa y tronco. El pino tea es una especie muy vigorosa y de forma mediocre, relativamente. De ahí que algunos tengan ramas largas, robustas y con mucha inclinación, mala poda, tronco ahorquillado o torcido. Lo cual no quita, si embargo, que en otros se dé una excelente combinación de copa y tronco. En estos debe centrarse la selección.
Ensayos de progenie revelan grandes diferencias en cuanto a forma de tronco y copa. En una progenie de siete años polinizada de árboles con copas bien conformadas, unos, y mal conformadas, otros, la heredabilidad era del 34 por 100 (Trousdell, Dorman y Squillace, 1963). En Texas, la relación “longitud media de ramas/altura del tronco” era apenas menor que en los testigos en la progenie de tres de cada cinco árboles seleccionados por su buena calidad, pero en la inclinación media de las ramas no hubo diferencias apreciables (Goddard, Brown y Campbell, 1959).
En Florida de efectuaron polinizaciones reguladas entre pinos tea torcidos, para comparar después las progenies con las de troncos derechos (Perry, 1960). A los dos años de edad, las progenies de cruzamientos entre árboles derechos tenían cuatro veces más plantones de padres torcidos. A los siete años había casi la misma proporción de árboles torcidos y derechos (Goddard y Strickland, 1964). Conclusión: aptitudes combinatorias para dar troncos derechos, lógicamente marcadas, sea en general que en concreto.
Propiedades madereras. Por su fuerza y resistencia, la madera del pino tea tiene múltiples aplicaciones en la industria transformadora y en la del papel. Para conservar su calidad y uniformidad, mejorando al par el crecimiento y rendimiento, y, además, de ser posible, las características especiales para cada producto, han hecho falta estudios de variación de la madera y la fibra. Como base para determinar las diferencias inherentes existen procedimientos técnicos que, sin destruir el árbol, permiten un muestreo de características en relación con su tamaño, edad y factores ambientales. Los resultados de todo ello han sido resumidos (Zobel, 1961, 1964) y no hay por qué repetirlos. La variación geográfica en la calidad de la madera ha sido investigada extensamente por su importancia en orden al rendimiento en pasta y su clase, y en su clasificación para sierra. Estudios emprendidos en todo el Sudeste por el Laboratorio de Productos Forestales, del Departamento de Agricultura de EE.UU. (1965), y por industrias y universidades, revelaron que el peso específico de la madera decrece en sentido SE.-NE., desde la costa hasta la región a pie de monte. Se advertían diferencias bastante grandes y uniformes, las cuales influían mucho en el rendimiento y en la calidad de la pasta, revelándose también importantes para las actividades forestales y las estructuras de la propiedad terrera de esta región.
En las dos Carolinas se observó que los árboles “a pie de monte” era superados por los de llanuras costeras en peso específico, porcentaje de madera de verano, extractos de alcohol bencílico, rendimiento en pasta y resistencia a la tracción (Einspahr, Peckham y Mathes, 1964). En Maryland y Delaware, peso específico y longitud de las traqueidas disminuían de Norte a Sur (Whitesell, Zobel y Roberds, 1966). Aunque en el sudeste de los Estados Unidos se hallaron diferencias en peso específico de la madera y varios tipos de celulosa, entre las localizaciones geográficas eran menores, sin embargo, que entre cada árbol (Zobel y McElwee, 1958; Zobel, Thorbjornsen y Henson, 1960).
Peso específico y contenido de humedad suelen estar estrecha e inversamente relacionados entre árboles de un área determinada. O sea que uno y otro, estructuralmente, corren paralelos. Pero esta relación no siempre rige para las distribuciones geográficas: En los pinos tea de las montañas de Tennessee, con menor peso específico, el contenido de humedad era alto, como en efecto se esperaba; mas en ciertos sectores de la llanura costera de Georgia, con peso específico elevado, la humedad era mayor en estos árboles que en los de la región adyacente “a pie de monte” (Zobel, Matthias, Roberds y Kellison, 1967).
La variación de árbol a árbol en orden al peso específico de la madera, longitud de las traqueidas y otros caracteres morfológicos ha sido estudiada en lo que al crecimiento se refiere. El primer trabajo importante en su género, de Zobel y Rhodes (1955), demuestra que lo mismo entre árboles que entre zonas geográficas y aun dentro de ellas existen amplias variaciones en peso específico, pero que entre el crecimiento diamétrico y el peso específico hay poca correlación. Otros muchos trabajos han sido publicados con idénticas conclusiones. Aunque no puedan citarse aquí, tales conclusiones figuran en documentos de van Buijtenen (1963), Einspahr, van Buijtenen y Peckham (1969) y Zobel, et al. (1969). En su conjunto, pueden resumirse así:
Para todas las características de la madera estudiadas se observan grandes diferencias de árbol a árbol entre los de igual edad y de una misma estación. Gran parte de esas diferencias se pueden corregir genéticamente, con muy insignificante heredabilidad; por lo general, del orden de 0,5 a 0,6.
Hay muchas características importantes no muy correlacionadas (p.ej., peso específico y longitud de las traqueidas). Por tanto, cabe la posibilidad de mejorarlas genéticamente por separado.
La edad del árbol tiene importante influencia en las propiedades de la madera, pues en los más jóvenes supone menor densidad, traqueidas más cortas y paredes de la célula menos gruesas.
La calidad de la madera resulta afectada por diversos ambientes, entre los que figuran lo abonos. Para la madera del pino tea, los abonos nitrogenados parecen ser los que más influyen.
En árboles más viejos, porcentaje de crecimiento y peso específico no están muy correlacionados. Esta escasa correlación permite el desarrollo de razas de crecimiento rápido con mucho o poco peso específico.
Los de zonas con elevadas precipitaciones estivales y climas cálidos tienden a dar maderas más densas que los de áreas secas o climas más fríos. Esta tendencia es una prueba documental para el peso específico y otras características morfológicas de la madera.
Es tal la estructura de la variación de la madera, singularmente las diferencias de árbol a árbol, que pueden hacerse buenos progresos mediante mejoras genéticas de la calidad.
+ Cuerda (“cord” = 1,55 m3). “Acre” = 0,4046 ha.
Resistencia a la sequía y al frío
En ciertas comarcas al oeste del área natural del pino tea, las precipitaciones son escasas en épocas de elevada temperatura. La resistencia a la sequía figura entre los caracteres deseados en los programas de mejoramiento genético. También hay sequía en los arenales de las áreas donde más llueve. La selección para mejorar la resistencia a la sequía ha dado resultados satisfactorios, como lo prueba el que las progenies de los árboles seleccionados sobreviven más que los testigos (van Buijtenen, 1966; Gilmore, 1957), y sin pérdida de crecimiento en altura (Goddard y Brown, 1959; Banks, 1966).
Resistencia a las plagas
La susceptibilidad a los daños provocados por insectos y enfermedades es importante en la mejora genética del pino tea, habida cuenta de sus efectos sobre la producción de semilla, el crecimiento del árbol, y el rendimiento en productos madereros por unidad de superficie, y la calidad de los mismos. En orden a la susceptibilidad a una enfermedad se han descubierto relevantes diferencias raciales y de árbol a árbol, y se van a emprender estudios sobre otras plagas.
La plaga más importante del pino tea es la roya fusiforme (Cronartium fusiforme Hedge. et Hunt ex Cumm.). Tiene gran poder destructivo en cientos de miles de acres, algunos de ellos infectados en más del 90 por 100, sobre una vasta franja que se extiende desde Carolina del Sur y Georgia hasta el 0. de Louisiana (Roth y McComb, 1971).
Un dato de gran utilidad para los proyectos de mejora genética es que los ensayos de progenies de clones de huertos de semillas demuestran una vasta gama de variación en la resistencia a la roya entre algunas familias (Woessner, 1955). En un extenso estudio con material parental de una población selvática, Kinloch y Stonecypher (1969) descubrieron heredabilidades de resistencia a la roya de 0,65 a 0,85, variación del 17,3 al 100 por ciento para la infección entre familias y correlación positiva para la resistencia entre padres y progenies. También se ha advertido variación en el germen patógeno con la latitud y la longitud (Snow, Kais y Dinus, 1972), y se han descrito formas albinas (Kais, 1966). El panorama que la importancia de las razas de roya ofrece está muy lejos de ser claro, pero los estudios actualmente en curso servirán de algo.
Un buen ejemplo de variación geográfica lo tenemos en la susceptibilidad a la roya fusiforme. La incidencia de la infección en el pino tea es atribuible al medio y a factores genéticos indistintamente (Wells y Wakley, 1966), En los que crecen en el E. de Carolina del Norte es poca, pero, plantados esos mismo árboles en el centro de Georgia, se infectan mucho. Se trata de una zona muy castigada. También hay poca infección en los pinos tea del E. de Texas, y si se les lleva al centro de Georgia, igual. Acaso el descubrimiento más importante de las actividades genéticas desarrolladas en los pinares del Sur sea el grado de resistencia a esta enfermedad apreciado en algunos árboles padres. Para sacar partido a este descubrimiento inicial se han establecido huertos de semillas resistentes a enfermedades determinadas o especiales (Blair, 1970; Zobel, Blair y Zoerb, 1971).
Rhyacionia frustrana Comst. (“Nantucket pine tip moth”) es una plaga común, pero de haber en el pino tea variación intraespecífica en la resistencia al ataque, al parecer debe ser poca. Se está estudiando la resistencia a los barrenillos de la corteza.
Asociación entre caracteres
La clase y magnitud de la variación entre individuos de pino tea es amplia, pero entre los caracteres hay pocas correlaciones estadísticas elevadas. Así, en la selección de árboles plus hay que tener en consideración muchos caracteres, como por un sistema de índice, para conseguir una buena combinación de aquellos silviculturalmente importantes. Toda selección para obtener múltiples caracteres ayudará a evitar que uno no satisfactorio pueda neutralizar varios que lo son, lo cual podría occurrir con el método de selección en tandem. Cierto es, sin embargo, que en el pino tea, al igual que en otros pinos meridionales, se da una vasta gama de correlaciones, tales como aumento diamétrico del tronco, y en la longitud de ramas y copa con mayor espacio ocupado. Pero hay excepciones, de modo que, para poder trabajar, el mejorador de árboles, el arboricultor, buscarán aquellos que tengan elevado porcentaje de crecimiento y otros muchos caracteres interesantes en orden a conformación de copa y tronco, producción de semilla, resistencia a las plagas y calidad de la madera.
Híbridos naturales y artificiales
Los pinos meridionales más importantes y algunas otras especies menores hibridizan naturalmente con el tea. La hibridación contribuye a la variación entre zonas geográficas y entre árboles de una misma masa. Tanto el número de caracteres como la medida en que la hibridación influye en ellos son económicamente importantes. La hibridación es, pues, un factor trascendental en la selección de razas geográficas o de árboles para plantaciones o para proyectos de mejora genética.
Pruebas de por lo menos tres generaciones de hibridación de pino longifolio y pino tea, con formas intermedias, fueron halladas en una masa natural de Louisiana (Namkoong, 1966). La existencia de individuos intermedios de pino tea y pino de hoja corta (Pinus echinata Mill.), en sus caracteres morfológicos (Zobel, 1953) y en las estructuras electro-foréticas de las proteínas de la semilla (Hare y Switzer, 1969), es señal de que este híbrido se da en Texas, donde hay poca infección por roya fusiforme. En Mississippí fueron hallados árboles parecidos a los híbridos artificiales de pino tea y pino de hoja corta en dos localidades (Mergen, Stairs y Snyder, 1965). En el nordeste del área natural hay híbridos de pino tea con pino resinoso (P. serotina Michx.) y pino rígido (P. rigida Mill.) (Little, Little y Doolittle, 1967; Smouse, 1970).
En los híbridos artificiales con pino tea, muchos caracteres morfológicos son intermedios (Little y Righter, 1965; Little et al., 1967; Schmitt, 1968), pero el rendimiento en semilla, por cono, puede llegar a ser reducido en extremo (Snyder y Squillace, 1966), y la resistencia a la roya fusiforme, grande. Los híbridos con pino rígido son comercialmente importantes en Corea, porque están mejor conformados que éste y resisten también al frío mejor que el tea (Hyun, 1970). Allí prefieren polen de pino tea de fuentes septentrionales.
Mutaciones
El pino tea es normalmente diploide, pero, aunque raramente, en las eras se dan plántulas mixo y poliploides. Las plantas aberrantes son poco vigorosas y por lo regular no maduran, por lo cual en masas son raras. En 30 de un total de 119 familias autógamas de pino tea se observó que 22 eran formas mutantes fenotípicamente distintas (Franklin, 1969). Por lo general, las formas mutantes no son lo suficientemente apetecibles ni tan comunes como para que constituyan la meta de un programa de selección.
EL PINO TEA COMO ESPECIE EXOTICA
Pese a su pasada “mala fama”, por ser de escaso rendimiento cuando se emplea semilla originariamente errónea, el pino tea se viene utilizando cada vez más como árbol exótico en varios países. Ha ganado en importancia en las zonas adecuadas, como, por ejemplo, en el Transvaal (Sudáfrica), en Nueva Gales del Sur (Australia) o en las grandes elevaciones o latitudes brasileñas y áreas adyacentes de la Argentina.
Pero para que el pino tea triunfe como especie exótica habrá que poner sumo cuidado al elegir la fuente de semillas. Por ejemplo, en regiones cálidas y sin heladas, regiones a las que el pino tea se adapta bien, la semilla de Florida es la que mejor crece, siguiéndole muy de cerca la de Livingston Parish (Louisiana) y, quizá, la del sur de Georgia. La diferencia en crecimiento entre el pino tea de Florida y el de la zona a pie de monte, más septentrional, en área de poca helada, puede ser tanta como de dos a uno; en regiones más frías se dan mejor las fuentes del litoral atlántico, como las de Onslow County (Carolina del Norte), mientras que en climas realmente fríos se darán bien las procedentes de elevaciones mayores, como las de Tennessee o de las grandes latitudes de Maryland. Por regla general, las fuentes de pino tea más meridionales son las mejores para las zonas heladas, o casi, y crecen más de prisa que las de regiones septentrionales o a pie de monte. Habrá también que poner buen cuidado para no confundir las fuentes de las llanuras costeras, más húmedas, con el pino tea que crece naturalmente en el interior, a pie de monte y en las dunas. El pino tea se da ocasionalmente en el norte de Georgia y Alabama, a casi 3 000 pies de altitud (más de 900 metros), pero, principalmente, más abajo de los mil pies (300 m.). Los que crecen en las mayores alturas y en el Norte son mucho más resistentes al frío que los del Sur y necesitan de noches frías para prosperar. Algunas fuentes occidentales, como, verbigracia, las de la región de “Los pinos perdidos”, en Texas, resisten mejor a la sequía que muchas de las fuentes orientales; la fuente tejana es también más resistente a las enfermedades.
Subdivisiones útiles
Entre las fuentes de las regiones a pie de monte y las de la llanura costera se aprecian muchas diferencias. La región o zona a pie de monte ocupa las estribaciones interiores desde la planicie y a lo largo de la costa; allí las temperaturas son más bajas, llueve menos y la primavera es seca, con suelos o franco arcillosos o arcillosos, mientras que en la llanura costera suelen ser arenosos sobre arcilla. Entre las dos hay una estrecha faja de dunas, con suelos profundos y áridos a los que el pino tea no se adapta bien. Suelen hacerse subdivisiones entre la llanura costera septentrional y la meridional, a lo largo del litoral atlántico. En la mejora de especies árboreas se establecen huertos de semillas separados para la región a pie de monte, y para las planicies costeras septentrional y meridional.
Divisiones parecidas se emplean más al Oeste, a lo largo de la costa del Golfo, donde reciben los nombres de baja llanura costera, alta llanura costera y, a veces, zona a pie de monte o estribaciones. También en esta área hay una región de dunas o de “fall line”. Entre las procedencias de la región a pie de monte (y la alta llanura costera) y las de llanura costera se aprecian claras diferenciaciones. El pino de la procedencia de la llanura costera crece más a prisa, lo mismo en habitats de aquella llanura que en los de la región a pie de monte, pero en años excepcionalmente secos o con ocasión de fuertes heladas repentinas, sucumbirá, en tanto que el de “a pie de monte” sobrevive.
Estudios del espesor de la corteza demuestran que en los árboles jóvenes procedentes de la llanura costera es más delgada que en los de procedencias de la región a pie de monte (25 por 100 frente al 33, en árboles de diez años). Aún está por ver si esta diferencia sigue dándose en pies adultos, para los cuales la media aproximada es del 12 al 16 por ciento. Las procedencias de la región a pie de monte dan semilla más pronto y con mayor abundancia que las de la llanura costera, incluso cuando unas y otras se han plantado en el mismo terreno. Casos ha habido en que un huerto de semillas de una procedencia de la región a pie de monte ha llegado a producir ocho o diez veces más que otro de una procedencia de la llanura costera de la misma edad e idéntico tratamiento. El conocimiento de esta diferencia en producción de semilla es importantísimo para el tamaño preciso que el huerto ha de tener.
Una amplia definición práctica de las procedencias útiles de pino tea incluirá las siguientes:
Norte de la Florida central. Esta área comprende la marga caliza que junto a la costa recibe el nombre de “Gulf Hammock” y los suelos profundos de Marion County. Esta fuente es apropiada para las condiciones subtropicales; de crecimiento muy rápido y madera de mucho peso específico, es sumamente susceptible al frío y útil sólo, por tanto, en zonas libres de heladas o con temperaturas no demasiado bajas. También sucumbe a la sequía con bastante facilidad. La fuente del norte de Florida es la procedencia de pino tea que crecía más de prisa sometida a prueba en Queensland, partes del Brasil y Sudáfrica. Tiende a dar árboles más deformes y ramas mayores que las especies más septentrionales. En su área nativa produce poca semilla, mientras que en algunos clones de huertos la producción es bastante satisfactoria, aunque tardía. Parece ser que, en las condiciones del sudeste de los Estados Unidos, una abundante producción de semilla de pino tea será un problema.
Livingston Parish (Louisiana) y sua alrededores. Al parecer, el pino tea de esta área ha resultado ser una fuente extraordinaria que ha dado rendimiento muy satisfactorio en un estudio realizado por todo el Sur, según una comunicación de O.O. Wells (1969). Buen porcentaje de crecimiento, conformación asimismo buena en general, y, por lo visto, abundante floración. Aunque pocas comparaciones directas han alcanzado ya la edad deseada, a nuestro modo de ver la fuente de Livingston Parish no crece con tanta rapidez como el pino tea de la Florida. Parece ser único en cuanto a resistencia a la roya fusiforme, incluso si se planta en las zonas más infectadas que haya. Actualmente se está ensayando en varias zonas del sudeste de los Estados Unidos, pero aún no ha pasado el tiempo suficiente para determinar su reacción al frío, a la sequía o a los suelos arcillosos de la zona a pie de monte, tierra adentro y lejos del área costera. Estimamos que esta es, ciertamente, una fuente preceptiva para ensayos en áreas subtropicales o regiones de poca helada.
Otras fuentes de la llanura costera. Los árboles de la llanura costera meridional de Georgia o de Carolina del Sur crecen muy bien, y en muchos casos pueden ser casi tan buenos como los expuestos en los apartados (a) y (b). Padres y masas deberán ser cuidadosamente elegidos si se desea sacarles el mayor partido posible. Ensayos con árboles de Onslow County (Carolina del Norte) han dado buenos resultados; conforme a nuestra propia experiencia, los árboles buenos procedentes de masas igualmente buenas crecen bien en condiciones subtropicales. Aunque no crecen tan de prisa como el de la Florida, son mucho más adaptables y resistentes.
El pino tea de la región a pie de monte y de la alta llanura costera (fuentes interiores). Este grupo comprende una extensa área continental o interior que llega hasta las llanuras costeras del Golfo y el Atlántico (figura 1). Los árboles de todas estas zonas interiores deberán ensayarse allí donde haya rigurosas condiciones climáticas, tales como sequías o heladas. Aunque de crecimiento más lento que algunos pinos tea de fuentes costeras, las fuentes interiores son las que deben utilizarse cuando se intende ampliar el área en que vaya a plantarse pino tea. No obstante aparecer juntas las fuentes de la región a pie de monte y las de la alta llanura costera, entre ellas se dan diferencias, de modo que decidir si conviene utilizar material de Virginia, de Carolina del Norte o del Sur, de Georgia, de Alabama, u otro más occidental, depende de la rigurosidad del ambiente en cuestión. De toda esta región hay muchísima semilla genéticamente mejorada, procedente de huertos de portagranos.
Fuente del litoral oriental. En Virginia y Delaware el pino tea crece en una península sobre terreno bajo y tendido, con suelo arenoso y, a veces, de arcilla húmeda. Esta fuente del litoral oriental no ha sido ensayada, pero, según parece, en condiciones de cierta salinidad se da bien. Las masas naturales están bien conformadas y producen grandes volúmenes por acre.
Fuentes septentrionales o de las grandes elevaciones. El pino tea no suele crecer a más de 1 000 pies (unos 300 metros), pero a veces se encuentran masas cerca de los 3 000 pies (900 m.). Estos árboles han sido ensayados y resultan ser más resistentes al hielo y la nieve que el pino tea corriente, aunque crecen con más lentitud que los de la costa. Este pino de las grandes altitudes sería adecuado para las regiones con temperaturas muy extremadas y en altiplanicies de zonas subtropicales donde los pinos propios no crecen bien. Las fuentes de pino tea de latitudes más septentrionales, como las de Maryland y Delaware, son también muy resistentes al frío, pero su crecimiento deja mucho que desear.
Estaciones excesivamente húmedas o secas. El pino tea ha dado pruebas de tolerancia a los grados extremos de humedad. Por ejemplo, los resistentes a la sequía que proceden de zonas occidentales como la región de los “Pinos perdidos”, en Texas (Bastrop County), crecen y sobreviven en condiciones cálidas y de sequía. La elección de los árboles padres es muy importante porque en algunos ensayos se han obtenido pies o muy ramosos o demasiado torcidos. La fuente de gran resistencia a la humedad la tiene también a la roya fusiforme. El empleo del pino tea resistente a la sequía puede ampliar considerablemente la gama de aplicaciones valiosas de esta especie.
Ciertas poblaciones de pino tea crecen en turberas profundas, tanto que a veces hay que ahondar hasta los seis metros para encontrar suelo mineral. Gracias a su excepcional sistema radical, este pino forma masa y crece bien en tales medios. Ensayado en condiciones de humedad excesiva, promete buenos resultados si el agua no está estancada. Se han establecido huertos de pinos “hidrófilos”, principalmente para las regiones costeras de Carolina del Norte y del Sur.
RESUMEN
El pino tea es una especie extraordinaria que crece en muy diversos medios y que ofrece una vasta gama de adaptabilidad. Muy variable también, hibridiza naturalmente con otras varias especies pinariegas. Responde bien a las manipulaciones genéticas y en orden a su mejoramiento se han dado ya muchos pasos adelante.
Como especie exótica no ha sido propiamente utilizada. Dada su extensa adaptabilidad, el empleo de la fuente geográfica exacta y precisa es fundamental para conseguir éxitos; en muchos de los primeros ensayos se ignoraba esta circunstancia, y los deficientes rendimientos conseguidos por tal causa ha hecho que algunos forestales tachen al pino tea de especie indeseable. Por lo general, se trata de un árbol de crecimiento rápido con buena madera pero de conformación un tanto defectuosa. De ahí que la selección de los árboles padres sea preceptiva.
Las fuentes meridionales son singularmente valiosas y crecen muy de prisa en los climas subtropicales. La madera del pino tea es, en general, de calidad satisfactoria, independientemente del medio en que el árbol crezca. Suele ser muy resistente a las enfermedades y los insectos, pese a su carácter exótico. El pino tea se empleará cada vez más, sobre todo ahora que se dispone de material genéticamente mejorado.
BIBLIOGRAFIA
Banks, P.F. 1966. “Early height growth in Rhodesia of progenies of P. taeda from Texas.” Rhod. Zambia Mal. Jour. Agri. Res. 4(1): 3–7
Barber, J.C. 1966. “Variation among half-sib families from three loblolly pine stands in Georgia.” Ga. For. Res. Council Res. Pap. 37. 5 págs.
Blair, R.L. 1970. “Quantitative inheritance of resistance to fusiform rust in loblolly pine.” Ph. D. Thesis, N.C. State Univ. 87 págs.
Crow, A.B. 1964. "Ten-year results from a local geographic seed source study of loblolly pine in southeastern Louisiana. La. State Univ. and A & M Coll., Sch. For. & Wildl. Manage., L.S.U. For. Note 57. 4 págs.
Dorman, K.W. y B.J. Zobel. 1973. Genetics of loblolly pine. USDA For. Ser. Res. Pap. W.O. 19. 21 págs.
Einspahr, D.W., J.R. Peckham y M.C. Mathes. 1964. Base lines for judging wood quality of loblolly pine. For. Sci. 10:165–173.
Einspahr, D.W., J.P. van Buijtenen y J.R. Peckham. 1969. “Pulping characteristics of ten-year loblolly pine selected for extreme wood specific gravity.” Sil. Gen. 18(3): 57–61.
Franklin, E.C. 1969. “Mutant forms found by self-pollination of loblolly pine.” Jour. Heredity 60(6): 315–320.
Gilmore, A.R. 1957. “Physical and chemical characteristics of loblolly pine seedlings associated with drought resistance.” Fourth South. Conf. For. Tree Improv. Proc. 1957:34–39.
Goddard, R.E. y C.L. Brown. 1959. “Growth of drought-resistant loblolly pines.” Tex. For. Ser. Res. Note 23. 7 págs.
Goddard, R.E., C.L. Brown y T.E. Campbell. 1959. An evaluation of growth and form in 5-year-old open-pollinated progeny from selected loblolly pine. Fifth South. Conf. For. Tree Improv. Proc. 1959:35–43.
Goddard, R.E. y R.K. Strickland. 1964. “Crooked stem form in loblolly pine.” Sil. Gen. 13(5):155–157.
Goggans, J.F., K.D. Lynch y G.I. Garin. 1972. “Early results of a loblolly pine seed source study in Alabama.” Auburn Univ., Agri. Expt. Sta. Cir. 194. 19 págs.
Grigsby, H.C. 1973. “A test of 36 geographic sources of loblolly pine seed in southern Arkansas.” USDA For. Ser. Res. Pap. SO. (En prensa).
Hare, R.C. y G.L. Switzer. 1969. “Introgression with shortleaf pine may explain rust resistance in western loblolly pine.” USDA For. Ser. Res. Note SO-88. págs.
Hocker, H.W., Jr. 1955. “Climatological summaries for selected stations in or near the southern pine region, 1921 – 1950.” USDA For. Ser. Southeast. For. Expt. Sta. Pap. 56. 11 págs.
Hyun, S.K. 1970. The growth performance of pitch-loblolly hybrid pine produced by different geographic races of loblolly pine in their early age. FAO/IUFRO Second World Consult. For. Tree Breed. 1969:803–814. FAO FO-FTB-69 7/7.
Kais, A.G. 1966. “Persistence of albinism in Cronartium fusiforme.” USDA Plant Dis. Rep. 50:842.
Kinloch, B.B., Jr. y R.W. Stonecypher. 1969. “Genetic variation in susceptibility to fusiform rust in seedlings from a wild population of loblolly pine.” Phytopathology 59(9):1246–1255.
Kraus, J.F. 1967. “A study of racial variation in loblolly pine in Georgia — tenth-year results.” Ninth South. Conf. For. Tree Improv. Proc. 1967:78–85
Lantz, C.W. y J.G. Hofmann. 1969. “Geographic variation in growth and wood quality of loblolly pine in North Carolina.” Tenth. South. Conf. For. Tree Improv. Proc. 1969:175–188.
Little, E.L., Jr., S. Little y W.T. Doolittle. 1967. “Natural hybrids among pond, loblolly and pitch pines.” USDA For. Ser. Res. Pap. NE-67. 22 págs.
Little, E.L., Jr. y F.I. Righter. 1965. “Botanical descriptions of forty artificial pine hybrids.” USDA For. Ser. Tech. Bull. 1345. 47 págs.
Mergen, F., G.R. Stairs y E.B. Snyder. 1965. “Natural and controlled loblolly-x-shortleaf pine hybrids in Mississippi.” For. Sci. 11(3):306–314.
Merrifield, R.G. 1965. “Loblolly pine seed sources in north Louisiana.” North La. Hill Farm Expt. Sta. For., Hill Farm Facts 6:4.
Namkoong, G. 1966. “Statistical analysis of introgression.” Biometrics 22(3):488–502.
Perry, T.O. 1960. “The inheritance of crooked stem form in loblolly pine” (Pinus taeda L.). Jour. For. 58(12):943–947.
Roth, E.R. y W.H. McComb. 1971. “Georgia fusiform rust survey 1971.” USDA For. Ser., Southeastern Area and Georgia Forestry Commission. 4 págs.
Schmitt, D. 1968. “Performance of southern pine hybrids in south Mississippi.” USDA For. Ser. Res. Pap. SO-36. 15 págs.
Smouse, P.E. 1970. “Population studies in the genus Pinus L.” Ph. D. Thesis, N.C. State Univ. 126 págs.
Snow, G.A., A.G. Kais y R.J. Dinus. 1972. “Further evidence of geographical variation in Cronartium fusiforme.” (Abst.) Phytopathology 62:790.
Snyder, E.B. y A.E. Squillace. 1966. “Cone and seed yields from controlled breeding of southern pines.” USDA For. Ser. Res. Pap. SO-22. 7 págs.
Trousdell, K.B., K.W. Dorman y A.E. Squillace. 1963. “Inheritance of branch length in young loblolly pine progeny.” USDA For. Ser. Res. Note SE-1. 2 págs.
USDA Forest Products Laboratory. 1965. “Southern wood density survey — 1965 status report.” USDA For. Ser. Res. Pap. FPL-26. 38 págs.
van Buijtenen, J.P. 1963. Inheritance of wood properties and their relation to growth rate in Pinus taeda. FAO World Consult. For. Genet. Tree Breed. Proc. 1963. FAO Forgen 63/ -7/2.
van Buijtenen, J.P. 1966. “Testing loblolly pines (Pinus taeda) for drought resistance.” Tex. For. Ser. Tech. Rept. 13. 15 págs.
van Buijtenen, J.P. 1969. “Seventeenth progress report of the cooperative tree improvement program.” Tex. For. Ser. Cir. 200. 14 págs.
Wakeley, P.C. 1961. Results of the Southwide pine seed source study through 1960–61. Sixth South. Conf. For. Tree Improv. Proc. 1961:10–24.
Wells, O.O. 1969. “Results of the Southwide pine seed source study through 1968–69.” Tenth South. Conf. For. Tree Improv. Proc. 1969:117–129.
Wells, O.O. y G.L. Szitwer. 1971. “Variation in rust resistance in Mississippi loblolly pine.” Eleventh South. Conf. For. Tree Improv. Proc. 1971:25–30.
Wells, O.O. y P.C. Wakeley. 1966. Geographic variation in survival, growth and fusiform rust infection of planted loblolly pine. Soc. Am. For., For. Sci. Mongr. 11. 40 págs.
Whitesell, C.D., B.J. Zobel y J. Roberds. 1966. “Specific gravity and tracheid length of loblolly pine in Maryland and Delaware.” N.C. State Univ., School For. Tech. Rept. 29. 11 págs.
Woessner, R.A. 1965. “Growth, form and disease resistance in four-year-old control- and five-year-old open-pollinated progeny of loblolly pine selected for use in seed orchards.” N.C. State Univ. Sch. For. Tech. Rept. 28. 67 págs.
Zobel, B.J. 1953. “Are there natural loblolly-shortleaf pine hybrids?” Jour. For. 51(7):494–495.
Zobel, B.J. 1961. “Inheritance of wood properties in conifers.” Sil. Gen. 10(3):65–70.
Zobel, B.J. 1964. “Breeding for wood properties in forest trees.” Unasylva 18(⅔):89–103.
Zobel, B.J., R. Blair y Zoerb. 1971. Using research data — disease resistance. Jour. For. 68(8):486–489.
Zobel, B.J., R.C. Kellison y M. Matthias. 1969. “Genetic improvement in forest trees — growth rate and wood characteristics in young loblolly pine.” Tenth South. Conf. For. Tree Improv. Proc. 1969:59–75.
Zobel, B.J. y R.L. McElwee. 1958. “Variation of cellulose in loblolly pine.” Tappi 41(4):167–170.
Zobel, B.J., M. Matthias, J.H. Roberds y R.C. Kellison. 1967. “Moisture content of southern pine trees.” Fourth. For. Biol. Conf. Proc. 1967:11–35.
Zobel, B.J. y R.R. Rhodes. 1955. “Relationship of wood specific gravity in loblolly pine to growth and environmental factors.” Tex. For. Ser. Tech. Rept. 11. 32 págs.
Zobel, B.J., E. Thorbjornsen y F. Henson. 1960. “Geographic, site and individual tree variation in wood properties of loblolly pine.” Sil. Gen. 9(6):149–158.
![]()
![]()
![]()