![]()
![]()
![]()
La investigación del contenido estomacal ha sido tradicionalmente un sector de actividad de la biología halieútica, pero es un sector en el que existen grandes dificultades para correlacionar los resultados con las investigaciones efectuadas en las otras esferas. Las investigaciones sobre el alimento de los peces no pueden considerarse por separado, sino que tienen que discutirse en relación con todo el medio marino, del que los peces constituyen elementos aislados. Por lo tanto, hay que hacer un breve análisis de los procesos más importantes de la ecología acuática, con referencia especial a la alimentación.
Los organismos vivos actuan unos sobre otros y con su ambiente no vivo (abiótico) de muchas maneras; ningún organismo existe independientemente de su ambiente. El estudio de estas relaciones recíprocas se llama ecología. Se puede estudiar la ecología de una especie con respecto a su ambiente o de un grupo de especies y sus acciones recíprocas entre ellas y con su ambiente. A la ecología no sólo le interesan las disciplinas biológicas sino también las físicas y químicas.
Toda área de la naturaleza en la que los materiales se cambian entre los organismos vivos y su ambiente abiótico, forma un sistema ecológico o ecosistema. Este concepto es importante porque subraya la interdependencia de las partes participantes. Aunque apenas es posible delimitar un área de la naturaleza en la que no influyan las áreas circunvecinas, no obstante se puede considerar un estanque, un lago o aún parte de un bosque como un ecosistema.
Para comprender la dinámica de tal ecosistema independiente como una unidad funcional, primero hay que examinar con cierto detalle las partes que lo forman.
Desde un punto de vista funcional, se pueden reconocer las etapas siguientes en el ecosistema (Figs. 6.1 y 6.2):
1. Recepción de la energía luminosa.
2. Producción de materia orgánica por los productores autótrofos (autoalimentación) utilizando nutrientes inorgánicos.
3. Consumo de la materia orgánica por los consumidores heterótrofos (otros métodos de alimentación).
4. Descomposición de la materia orgánica en nutrientes inorgánicos convenientes para los productores por microconsumidores saprofíticos, principalmente bacterios y hongos.
Los productores principales son las plantas verdes que tienen clorofila; las bacterias sistetizantes generalmente tienen muy poca importancia.
Los consumidores comprenden todos los demás componentes vivos presentes, entre ellos los herbívoros (vegetarianos) que se alimentan directamente de los productores y los carnívoros (predadores) que se alimentan de los herbívoros y de otros carnívoros. También comprenden los parásitos, los necrófagos (devoradores de carroña) y los saprofitos. Aunque conviene incluir en la lista los que causan la descomposición, que comprenden bacterias y hongos, como una entidad separada por su función específica y por ser indispensables, son igualmente consumidores.
Los nutrientes inorgánicos comprenden muchos de los elementos presentes en forma de sales disueltas. Los más importantes son el nitrógeno y el fósforo, seguido del potasio, calcios, azufre y magnesio. Algunos elementos se necesitan en cantidades extremadamente pequeñas y por eso se denominan micronutrientes.
La producción de substancias orgánicas (alimento) por fotosíntesis es un proceso que implica la transformación de la energía luminosa en energía química potencial. La cesión de esta energía alimentaria desde los productores por una serie de consumidores se llama cadena alimentaria y cada organismo por lo cual pasa es un eslabón de esa cadena.
Para simplificar un poco, se reconocen tres cadenas alimentarias diferentes:
1. La cadena de los carnívoros en la que la energía pasa de los organismos más pequeños a los mayores.
2. La cadena de los parásitos en la que la energía pasa de los organismos mayores a los más pequeños.
3. La cadena saprofita, en la que la energía pasa en la mayor parte de los casos de la materia orgánica muerta a los microorganismos.
En realidad, el alimento puede pasar por partes de las tres cadenas antes de que acaben descomponiéndolo en nutrientes inorgánicos las bacterias y hongos que se encuentran al final de todas las cadenas alimentarias. En otras palabras, las poblaciones de especies dentro de una comunidad o ecosistema forman muchas cadenas alimentarias que interconectan, comunican o se cruzan entre ellas de una manera compleja que se denomina usualmente trama alimentaria.
Los organismos que pertenecen al mismo eslabón de la cadena alimentaria contados desde el nivel del productor, se dice que pertenecen al mismo nivel trófico. Las plantas son, por tanto, el primer nivel trófico, los herbívoros el segundo y los carnívoros que se alimentan de herbívoros, el tercero. Los carnívoros secundarios que se alimentan de carnívoros del tercer nivel pertenecen al cuarto nivel trófico y así sucesivamente. No obstante, tiene un límite bien definido el número de posibles eslabones de una cadena alimentaria y, por lo tanto, el número de niveles tróficos de todo ecosistema. La razón es que solamente el 10 por ciento de la energía disponible es asimilada al pasar de un nivel trófico a otro. En el extremo de la cadena alimentaria usualmente no hay más que uno o dos grandes predadores. El número de especies de cada capa trófica aumenta cuando se aproxima al primer nivel dando origen a lo que se llama una pirámide de números. Para los grandes predadores, la introducción de pequeñas cantidades de contaminantes en la primera capatrófica puede tener consecuencias fatales porque finalmente se concentran en ellos.
Las leyes de la termodinámica dicen que la energía no puede crearse ni destruirse, pero también que no puede transformarse de una clase en otra sin dispersión parcial en forma de calor. Esto significa que la transformación de la energía luminosa en energía química potencial, en forma de los compuestos orgánicos de las plantas, no puede ser eficiente al 100 por ciento. Solamente una parte muy pequeña de la energía luminosa absorbida por las plantas verdes se transforma en energía alimentaria (producción bruta), porque la mayor parte se disipa en forma de calor. Además, una parte de la producción bruta la emplean las plantas en sus propios procesos respiratorios, dejando una cantidad todavía menor de energía potencial (producción neta) disponible para ser transmitida al nivel trófico siguiente.
La pérdida de energía se denomina habitualmente pérdida respiratoria porque los organismos utilizan la energía de los alimentos oxidándola. Debido a las pérdidas respiratorias, las cadenas alimentarias no pueden ser muy largas y, por lo tanto, el número de niveles tróficos en las comunidades naturales es raramente más de cuatro o cinco y a menudo sólo tres. Esto significa también que la cantidad total de alimento disponible disminuye al aumentar el nivel trófico. Por esta razón, los animales más grandes se alimentan de plantas o de otros animales que están en un nivel trófico más bajo, como, por ejemplo, la ballena de krill y el elefante de plantas.
En los animales la producción bruta corresponde al alimento asimilado, que significa al alimento ingerido y absorbido por los intestinos. En este caso la producción neta equivale al alimento asimilado menos la respiración.
Aunque casi toda la energía perdida dentro de un ecosistema se debe al proceso respiratorio, hay otras pérdidas que afectan a organismos individuales. Parte de los alimentos potenciales no se ingieren, sino que se descomponen directamente o so almacenan o salen del sistema o de la comunidad. Otro motivo de pérdida es que no se asimila todo el alimento que se ingiere; parte pasa por el conducto alimenticio y se pierde in forma de heces.
Como se ha dicho anteriormente, cuando los organismos mueren son atacados por los descompositores que obtienen de ellos su energía reduciendo su contenido orgánico en nutrientes inorgánicos. Como también se ha indicado, estos elementos nutritivos pueden ser utilizados de nuevo por los productores con el resultado de que los materiales en cuestión circulan continuamente en el sistema. No obstante, la corriente de energía pasa por una “calle de dirección única”. Para mantener el funcionamiento de un ecosistema hay que suministrarlo constantemente de energía luminosa.
Como hemos visto, la verdadera producción de materia orgánica sólo tiene lugar en las plantas que poseen clorofila y ciertas bacterias sintetizantes y a esto se ha denominado producción primaria. Sin embargo, los copépodos y los eufasiaceos, por ejemplo, se denominan con frecuencia productores de carne o animales de la “industria clave”, porque transforman la materia vegetal en proteina que pueden asimilar los animales que se los comen, pero que ellos mismos no podrían subsistir de materia vegetal. En realidad sólo asimilan y almacenan la energía derivada de los productores primarios. Para evitar confusiones sería mejor llamarlos productores secundarios, término que se aplica igualmente bien a los animales de niveles tróficos superiores, porque también ellos - aunque indirectamente - utilizan la producción primaria de las plantas.
Desde el punto de vista práctico, con frecuencia conviene determinar la amplitud de la producción secundaria de ciertos animales de una área dada, digamos un banco de pesca, o lo que es más importante, si puede aumentarse una producción conocida. La estimación de la producción debe basarse en factores como el “cultivo permanente” (biomasa), el índice de remoción de materias y de crecimiento, incluido el de los jóvenes nacidos o eclosionados durante el período del censo. El índice de renovación también es interesante cuando se trata de especies de vida corta, como es casi siempre el caso en el ecosistema del mar. La producción biológica se tiene que expresar en unidades de tiempo. Una gran biomasa no es en absoluto sinónima de un graníndice de producción. Tomamos un ejemplo fácil de imaginar: un prado en el que pastan unas vacas puede tener una biomasa de hierba muy pequeña, porque la producción se consume mientras se produce, pero, no obstante, puede tener una producción mayor que un pasto vecino con una enorme biomasa, pero no consumido.
Las relaciones cuantitativas entre los diversos niveles tróficos se pueden calcular si se conocen los ritmos de producción para cada nivel que interese. También son de enorme interés las relaciones de esta naturaleza dentro de los niveles tróficos. Los resultados de estos cálculos expresados como porcentaje se citan con frecuencia como rendimientos ecológicos porque están en relación con el rendimiento de la transmisión de energía a diferentes puntos de la cadena alimentaria. Por esto son importantes para nuestra comprensión de la dinámica de los ecosistemas. Además, casi todos los índices de rendimiento son significativos en lo que concierne a poblaciones de una sola especie, así como a todos los niveles tróficos.
Por desgracia existe mucha confusión en la terminología empleada por los diversos autores y no es siempre claro a que índice de rendimiento ha deseado un autor referirse realmente. Odum (1959) ha tratado de definir los diversos índices basándose en su diagrama del flujo de la energía y es ciertamente un buen método para ilustrar las complejidades en cuestión (Fig. 6.3).
En biología halieútica la manera más común de describir el rendimiento es por el coeficiente de conversión, es decir: la relación entre el peso del alimento consumido por el pez y su aumento de peso. Algunos autores lo expresan como el coeficiente de nutrición. En la Fig. 6.3 se observa que el coeficiente de conversión es el valor recíproco de lo que Odum denomina rendimiento del crecimiento ecológico. El valor del coeficiente de conversión se define tradicionalmente como 10, pero en realidad puede ser mucho mayor o mucho menor, por ejemplo, en las factorías trutícolas del norte de Europa el coeficiente es de cerca de 5.
No obstante, es importante recalcar que ninguno de los rendimientos ecológicos es constante para ninguna especie o para todo un nivel trófico; depende de factores abióticos como la temperatura y salinidad, así como de factores bióticos como tipo, abundancia, distribución del alimento disponible y edad de los consumidores; por ejemplo, las larvas y los jóvenes de todas las especies investigadas tienen coeficientes de conversión mucho más bajos que los animales de más edad.
Casi todo el trabajo que se ha realizado sobre los alimentos ingeridos por los peces ha sido cualitativo más bien que cuantitativo. Es decir, los investigadores han descrito la presencia del alimento encontrado en el conducto alimenticio, usualmente en el estómago sólo. Esto indica lo que ha comido el pez y en que cantidades aproximadas, pero no describe cuanto de cada clase de alimento ha consumido. La razón de la falta de estudios cuantitativos es que necesitan mucho tiempo.
Lo antes posible después de la captura se diseca el conducto alimenticio y si los investigadores no pueden terminar inmediatamente, quizá por falta de medios de laboratorio, el material se conserva en formol al 4 por ciento tamponado.
Más tarde, en el laboratorio se abren las diversas secciones del conducto alimenticio y los grupos distintos de organismos se separan para identificarlos, lo que se hace muy fácilmente con un microscopio binocular de disección. Para ilustrar los métodos empleados y para describir sus ventajas e inconvenientes se estudiará una serie de resultados basados en el examen del contenido estomacal de dos Lates niloticus (Tabla 6.1).
La presencia de cada alimento se anota y expresa como el porcentaje de la presencia de todos los organismos alimenticios. El método es muy rápido y fácil, pero subestima la importancia de las especies mayores. En el ejemplo, el 40 por ciento de todas las presencias de alimento eran ninfas de Odonata. El contenido del estómago de estas dos L. niloticus pudo haber consistido casi enteramente en ninfas, cada una con una Tilapia galilea muy pequeña, pero los resultados no indicaron si ésta podría ser una verdadera interpretación de los datos.
El número de trozos de cada alimento se anota y el resultado se expresa como porcentaje del número total de trozos de alimento presentes. Este método sobreestima la importancia de organismos pequeños en la alimentación, en este caso las ninfas de Odonata; y no es satisfactorio con alimentos vegetales, excepto para el examen microscópico de las algas. En el ejemplo dado, el material vegetal puede ser un trozo de hierba grande o pequeño.
El método es similar al de la frecuencia de presencias, excepto que los resultados se expresan como el número de peces en los que se ha encontrado el alimento, expresado a su vez como el porcentaje del número total de peces. El método indica cuales son las principales especies alimentarias, pero no su importancia relativa como fuentes de energía.
Se mide el volumen total de lo que contiene el estómago y esto se clasifica en diferentes tipos de alimento, determinándose el volumen de cada uno. Los resultados se comunican usualmente como porcentaje del volumen total.
Este método es más dilatorio que los anteriores, pero describe exactamente la importancia relativa de cada alimento y el volumen total (∑) indica la cantidad de cada alimento consumido.
Este método es idéntico al precedente, excepto que el volumen de cada alimento se expresa primero como porcentaje del volumen total de lo que contiene el estómago del que proviene y después se obtiene la media de todos los peces objeto de muestreo. La ventaja de este método sobre el anterior es que los resultados finales serán más representativos de los hábitos alimentarios de un grupo de peces, si uno o dos ejemplares de ese grupo han consumido mucho más de un alimento determinado. El inconveniente es que se pierde la información sobre el contenido real del volumen de lo que contiene el estómago.
Se determina el peso total de todos los alimentos. Este método es muy parecido al volumétrico, sobre el cual tiene poca ventaja, a menos que la muestra esté seca, en cuyo caso sólo se determina el peso seco. Normalmente los resultados se presentan como porcentaje del total. Si se determinan los pesos en húmedo, los alimentos se ponen en un secante para suprimir el exceso de humedad. Este problema de remover el exceso de humedad hace que el método sea algo subjetivo, por lo que es mejor, aunque mucho más largo, el peso en seco.
Se asignan puntos a cada alimento. El número de puntos depende de que el alimento abunde mucho en el contenido estomacal (mayor número de puntos) o sea muy raro (menor número) y de su tamaño (más puntos para el grande que para el pequeño). El método puede modificarse para tener en cuenta la plenitud del estómago. Es rápido, fácil y no requiere aparatos especiales; con experiencia puede ser muy preciso. En el ejemplo dado (Tabla 6.1) los puntos asignados dan un resultado similar a los métodos volumétrico y gravimétrico. Rounsefell y Everhart (1953) dan más detalles de estos métodos.
Todos estos métodos describen, con más o menos exactitud, la cantidad de alimento que tenían en el estómago las dos L. niloticus empleadas como ejemplo; los mejores fueron los métodos volumétrico y gravimétrico. La suma de los volúmenes reales y de los pesos también indica el grado de repleción del estómago, lo que permite describir los ciclos de alimentación. El sistema de puntos también es satisfactorio si se emplea objetivamente y tiene la ventaja de que es fácil y rápido y puede adaptarse para tener en cuenta la repleción gástrica.
Sin embargo, ninguno de estos métodos describe la cantidad ingerida de cada alimento. En el ejemplo dado, T. galilaea puede tardar mucho en ser digerida y es posible identificarla mucho después que dejan de serlo las ninfas de Odonata. En lo que tarda en digerirse una T. galilaea, de 20 a 30 ninfas de Odonata pueden haber sido ingeridas y digeridas, pero en un momento cualquiera sólo de 4 a 6 pueden ser reconocidas en el estómago. Por otro lado, algunos alimentos pueden haber sido regurgitados en el momento de la captura. Aunque los métodos cualitativos permiten construir las cadenas alimentarias, no describen el flujo de energía por ellas ni permiten determinar la importancia de, digamos, la predación de una especie de valor sobre otra igualmente valiosa. Si se conocieran las soluciones de estos problemas en algunos casos podría considerarse aconsejable reducir la abundancia de predadores, para que aumentara la de las presas, especialmente si estas estuvieran en un nivel trófico más bajo (y de más rendimiento). Estas cuestiones sólo se pueden solucionar con estudios cualitativos.
Los estudios cuantitativos de la alimentación son muy largos y, por lo tanto, la labor de los especialistas. Sólo se esbozarán los métodos empleados. Primero se elije la unidad en que se va a medir el flujo de energía, siendo las dos más comunes las calorías y el nitrógeno; segundo, los experimentos de alimentación se efectúan en acuarios con diferentes alimentos que consume la especie que se estudia a fin de determinar cual es su capacidad para convertir el alimento en carne. En el caso de las calorías, esto significa determinar el valor calórico del alimento seleccionado, dar a los peces un peso conocido de éste, recoger las heces y determinar su valor calórico. Para aplicar las observaciones sobre los pesos y valores calóricos del alimento encontrado en el estómago, hay que establecer la velocidad de la digestión alimentando a los peces y matándolos a intervalos conocidos. Es posible que esto se tenga que hacer a más de una temperatura. Con estos datos se aprende lo que comen al día peces de diversos tamaños de la misma especie, cuanta energía proveniente del alimento utilizan para mantenerse y crecer. Un estudio de Daan (en prensa) muestra que el bacalao grande de la parte sur del mar del Norte probablemente influye mucho en las tallas de las clases de edad al comer bacalao pequeño del grupo O, así como grandes cantidades de arenque y lenguado. Boddeke (1971) también ha demostrado que recientes clases anuales grandes de bacalao han reducido la abundancia del camarón gris (Crangon crangon) en aguas holandesas. Daan ha sugerido que la producción total de especies comerciales de la parte sur del mar del Norte podría aumentarse reduciendo la abundancia de bacalao mediante una pesca intensa de la población. La solución de tales relaciones predador-presa podrían modificar la manera en que se han hecho algunas estimaciones de los peces.
Allee, W.C. et al., 1949 Principles of animal ecology. Philadelphia, W.B. Saunders
Boddeke, R., 1971 The influence of the strong 1969 and 1970 year classes of cod on the stock of brown shrimp along the Netherlands coast in 1970 and 1971. ICES C.M. Shellfish and Benthos Committee. Paper K32:3 p. (mimeo)
Clarke, G.L., 1954 Elements of ecology. Nueva York, Wiley and Sons. 534 p.
Daan, N., A quantitative analysis of the food intake of North Sea cod. Gadus morhua, L. Neth.J.Sea Res., (en prensa)
Hardy, A.C., 1959 The open sea; its natural history. Pt. 2. Fish and fisheries with chapters on whales, turtles and animals of the sea floor. Londres, Collins, 322 p.
Laevastu, T., 1965 (comp.), Manual of methods in fisheries biology. FAO Man.Fish.Sci., (1):pag. var.
Marshall, N.B., 1966 The life of fishes. Cleveland, Ohio, World Publishing Co., 402 p.
Nikolskii, G.V., 1963 The ecology of fishes. Londres, Academic Press, 352 p.
Odum, E.P., 1959 Fundamentals of ecology. Philadelphia, W.B. Saunders, 546 p.
Rounsefell, 1953 G.A. y W.H. Everhart, Fisheries science: its methods and applications. Nueva York, Wiley and Sons, 444 p.
Steele, J.H. (ed.), 1970 Marine food chains. Edinburgh, Oliver and Boyd, 552 p.
TABLA 6.1
Ejemplos de los resultados obtenidos empleando diferentes métodos de estimación del contenido estomacal de dos Lates niloticus
L. niloticus 1. | 1 Tilapia galilaea 20 cm, peso 80 g, volumen 100 ml |
| 6 ninfas de odonata (libélula) cada una de 1 cm, 0,5 g, 1 ml | |
L. niloticus | 1 T. galilaea 10 cm, 10 g, 15 ml |
| 4 ninfas de Odonata cada una de 1 cm, 0,5 g, 1 ml | |
Resultados
| Food | Método | Pez | Ln 1 | Ln 2 | ∑ | % | Total del cual se expresa en % |
|---|---|---|---|---|---|---|---|
| Presencia | |||||||
| T. galilaea | 1 | 1 | 2 | 40 | presencia de todos los alimentos | ||
| Odonata | 1 | 1 | 2 | 40 | |||
| Vegetales | 1 | 0 | 1 | 20 | |||
| Numérica | |||||||
| T. galilaea | 1 | 1 | 2 | 14,4 | todos los organismos alimentarios | ||
| Odonata | 6 | 4 | 10 | 76,9 | |||
| Vegetales | 1 | 0 | 1 | 7,7 | |||
| Dominante | |||||||
| T. galilaea | 1 | 1 | 2 | 100 | todos los peces | ||
| Odonata | 1 | 1 | 2 | 100 | |||
| Vegetales | 1 | 0 | 1 | 50 | |||
| Volumen total | |||||||
| T. galilaea | 100 | 15 | 115 | 87,1 | volumen total de alimentos | ||
| Odonata | 6 | 4 | 10 | 7,6 | |||
| Vegetales | 7 | 0 | 7 | 5,3 | |||
| % volumen | |||||||
| T. galilaea | 88,5 | 79 | 167,5 | 83,8 | volumen de alimentos | ||
| Odonata | 5,3 | 21 | 26,3 | 13,1 | |||
| Vegetales | 6,2 | 0 | 6,2 | 3,1 | |||
| Gravimétrico | |||||||
| T. galilaea | 80 | 10 | 90 | 90 | peso total del alimento | ||
| Odonata | 3 | 2 | 5 | 5 | |||
| Vegetales | 5 | 0 | 5 | 5 | |||
| Puntos | |||||||
| T. galilaea | 50 | 10 | 60 | 85,7 | alimentos | ||
| Odonata | 5 | 4 | 9 | 12,9 | |||
| Vegetales | 1 | 0 | 1 | 1,4 |
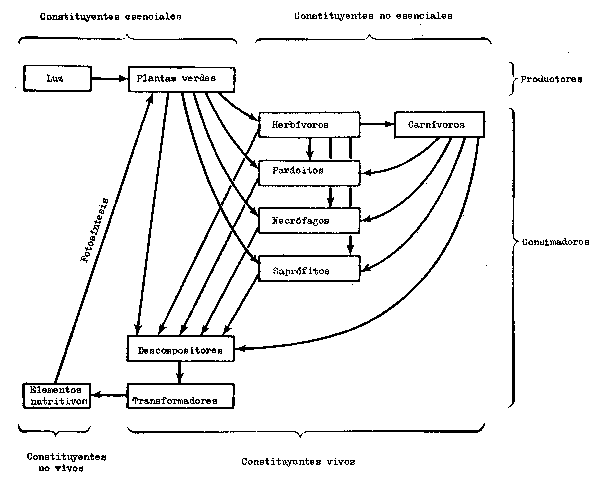
Fig. 6.1 Principales etapas y composición de un ecosistema autónomo (de Clarke, 1954)
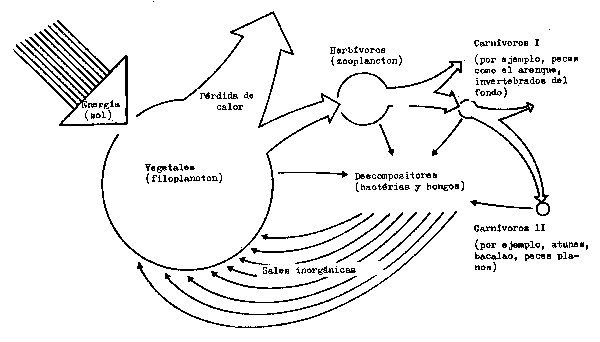
Fig. 6.2 Ejemplo simplificado de un ecosistema marino. El flujo de la energía está representado por la flechas abiertas
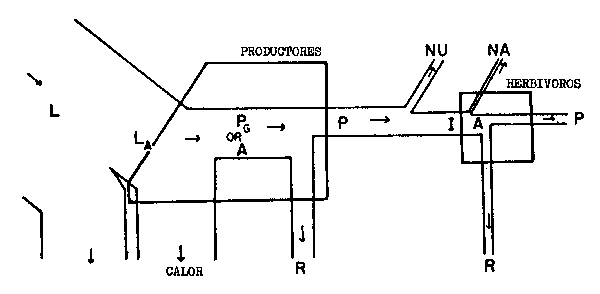
| Relaciones | Designación y explicación | |
|---|---|---|
| A. Relación entre los niveles tróficos | ||
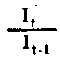 | Nivel trófico de absorción de energía o rendimiento (de Lindeman). Para el nivel primario es | |
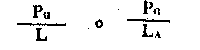 | ||
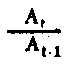 | Nivel trófico de rendimiento de la producción | Para el nivel primario P y A puede ser en función de L o LA como supra; At/At-1=It It-1para el nivel primario, pero no para el secundario |
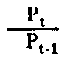 | Nivel trófico de rendimiento de la asimilación | |
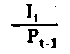 | Rendimiento de utilización | |
| B. Relaciones dentro de los niveles tróficos | ||
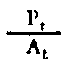 | Rendimiento de crecimiento de los tejidos | |
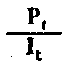 | Rendimiento del crecimiento ecológico | |
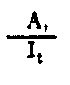 | Rendimiento de la asimilación | |
Fig. 6.3 Diversos tipos de rendimiento ecológico. Los símbolos empleados son (ver diagrama): L = luz (total), LA = luz absorbida, PG = fotosíntesis total (producción bruta), P = producción de biomasa, I = energía absorbida, R = respiración, A = asimilación, NA = ingerido pero no asimilado, NU = no utilizado por el nivel trófico indicado, t = nivel trófico, t-1 = nivel trófico precedente (de Odum, 1959)
![]()
![]()
![]()