![]()
![]()
![]()
by
R.L. Welcomme
Fishery Resources Officer
Aquatic Resources Surveys and Evaluation Service
Fishery Resources and Environment Division
FAO, Rome
Abstract
This paper describes the morphology of floodplains and the chemical and physical changes with the flood cycle. Aspects of the biology of the fish species are presented and the principal methods of fishing summarized. The data on productivity and standing stock are collected and assessment made of the effects of differing flood regime on catch. Conclusions are drawn as to possible research and management policies for floodplain fisheries.
Despite the considerable amount of work that has been carried out on the fisheries of African inland waters in recent years, surprisingly little effort has been directed at the biology of the fish of river or floodplain systems. This has resulted in a general lack of understanding of the ecological mechanisms regulating the fish populations of such systems, although some of the most important fisheries in Africa are located on them.
Because of the general world pressure for food and hydro-electric power, rivers and floodplains, like many other ecosystems, are coming under increasing demand for rational management. In the case of floodplains this includes the development of a number of activities other than fisheries, some of which conflict with the maintenance of fish communities in their waters.
To ensure that fisheries play their proper role in the integrated development of such areas, it is vital that the basic information be obtained that is needed to evaluate effects of changes in the ecosystem on the fish. As a preliminary step, the attention of the First Session of the Committee for Inland Fisheries of Africa (Fort Lamy, Chad 1972) was drawn to these problems (Welcomme, 1974) and it was recommended that a Correspondence working group be established for the collection of existing data on floodplain fisheries and for the stimulation of research in these areas (First Session of FAO Committee for Inland Fisheries of Africa).
This paper, which is intended to partially fulfil the first part of the recommendation, is based on a review of the literature, personal observations and unpublished information supplied by various African fishery workers, summarizes what is known of the fisheries ecology of the floodplain systems of the African continent. Some observations are necessarily speculative or based on circumstantial evidence, but it is hoped that these will stimulate interest and discussion and serve as a basis for future work on this very important area.
A floodplain is a strip of relatively low-lying flat land bordering a river which is submerged seasonally. According to Leopold, Wolman and Miller (1964) a typical floodplain includes the following features:
sand splays, deposits of flood debris usually of coarser sand particles in the form of splays or scattered debris.
In view of these authors floodplains are depositional features of a river valley whereby sediment is temporarily stored in the floodplain. Under equilibrium conditions averaged over a number of years net inflow of sediments equals net outflow, although local changes in the morphology of the floodplain occur through silting of old channels and cutting of new ones.
Most of these major features of floodplains can be seen in larger rivers, although in smaller water courses they may be hard to distinguish due to the rapidity with which change occurs in them.
The features of the floodplain that are ecologically more important for fish are (Fig. 1 and Fig. 2):
The main river channel, which may be either simple or braided. Braided channels (see Fig. 3) consist of many anastomosing water courses meandering among sandy and vegetated islands, which are exposed at low and flooded at high water. Such channels are typical of portions of the Zambezi, Niger and Congo Rivers and where they occur the lateral floodplain is often restricted in width. There is some evidence (Svensson, 1933; Gosse, 1963) that ecologically such islands are analogous to the lateral floodplain and fulfil a similar role in the biology of the fish.
The floodplain, which is submerged during the flood phase and dries out during the low water phase. The majority of floodplains are on rivers of the “Sudanian” type, i.e., with grass savannah vegetation, although some rivers of the “Guinean” type have forested floodplains which give the flooded forest typical of many Zairian rivers.
Swamps and pools (lagoons) situated in depressions of the floodplain which may be permanent, persisting from one flood phase to the next, or temporary, drying out progressively as the low water phase advances. The distinction between swamps and pools depends largely on the amount of vegetation which can alter throughout a season. There is a tendency for permanent pools to become swamps as the water level in them is lowered. In their most extreme form these features comprise a considerable portion of the floodplain area; systems such as the Luapula (Fig. 1B) and Pongola consist of strings of large permanent lakes, and the Elephant marshes of the Shire River floodplain are extensive persistant swamps.
Channels and creeks, which connect the depressions of the floodplain with each other and with the river form a network over the plain. Some of these penetrate the levée and represent the main paths of water movement during the earlier periods of rising water and the later phases of falling water.
Levées or raised banks bordering the main river channel, which may be significant in the way in which they direct or deflect the water flow over the floodplain and delay both the onset of flooding and the drainage and drying of the plain.
The importance of these features is frequently acknowledged by the peoples living on floodplain systems and is reflected in their language where each is named (Holden, 1963; Hurault, 1965; CTFT, 1972; Rzoska, 1974). For example, in the Senegal the levée is “Fondé” and the depressions of the floodplain “Oualo” are fed by channels “tiangol” and retain in their lowest part a permanent pool or marsh “Vindou”.
Svensson (1933) distinguishes three main types of floodplain swamp depending on the manner of their inundation. His type 1 swamps are formed from local precipitation or drainage only and become connected with the river or other lower lying swamps by a single channel or creek. Such swamps are never influenced by variations in river height, but fish may migrate to them by ascending the connecting channel. Type 2 areas are situated immediately alongside the river and are flooded directly by it. Swamps of the third type (type 3) are intermediate between the other two and are located in the shallow depressions of the floodplain separated from the river by levées. Access to them is by channel and flooding results in the first instance from local rainfall and drainage and only later by a rise in river level.
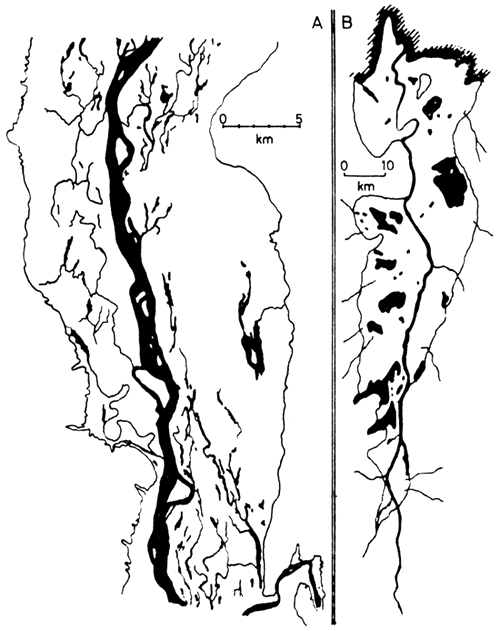
Figure 1 Features of floodplain systems
A Niger River (FAO 1969)
B Luapula river
Systems shown at low water, black - permanent swamp
and lagoon, main river channel and creeks. The limits
of flooding at high water are also indicated
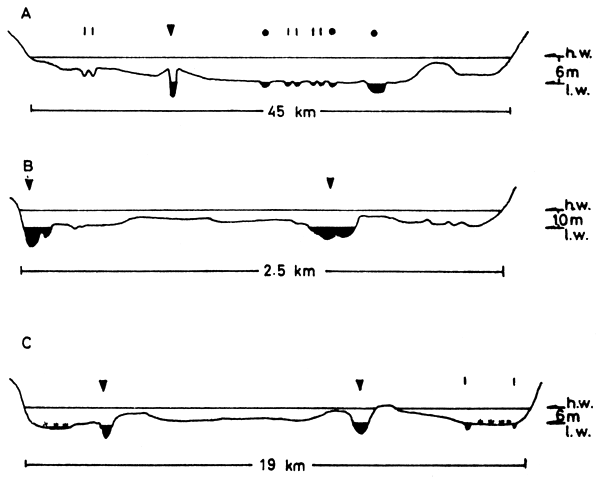
Figure 2 Cross sections of three river/floodplain systems
(vertical scale exagerated)
A Kafue (UNDP/FAO 1968)
B Niger at Onitsha (NEDECO 1959)
C Oueme at Zinvie
▼ Main river channel
• Lagoon
 Stream or secondary channel
Stream or secondary channel
 Permanent swamp
Permanent swamp

Figure 3 Braided channel, Niger River
Floodplains may also be separated by the size and permanence of the water body with which they are associated as the dynamics of fish populations of river/floodplain systems connected with large lakes (e.g., the Yaérés of the Chari-Logone Rivers (Durand, 1970) or the floodplain of the Kenyan rivers flowing into Lake Victoria) are likely to differ greatly from those of rivers that break into a series of isolated pools in the dry season, if only by virtue of the carrying capacity of the water mass available during low water (Van Someren, 1961).
A further distinction can be made between fringing floodplains that are normal features of the longitudinal evolution of the river, and those areas that arise by geographical accident. The former are normally very long but of relatively restricted width, whereas the latter of which the central delta of the Niger or the Kafue flats of Zambia are typical, are usually large areas, disproportionate to the rest of the system, which arise due to the particular configuration of the land and may occur at any point of the river. Further floodplains of this type are common in deltaic areas toward the mouth of a river, for example, the Ouémé and Senegal.
Rivers, themselves, vary in nature regionally within the African continent. Daget and Iltis (1965) classify West African rivers into (i) Guinean and (ii) Sudanian types.
(i) Guinean rivers are those which drain the forested areas of West Africa. They run their course through gallery forest and have floodplains which may extend to inundate large areas of forest floor. These are reputedly less productive than Sudanian rivers.
(ii) Sudanian rivers drain the open savannah regions and have little or no fringing forest. The floodplain is usually extensive and grass-covered.
Any absolute classification of floodplains into types based on their morphology is, however, difficult as the range of variation encompasses too many intermediate forms. It is, nevertheless, useful to bear these differences in mind when interpreting the behaviour of the different regions.
Fringing floodplains may be regarded as normal developmental features of rivers and they are present to a greater or lesser extent in most African water courses. Major floodplains arising from local geographical anomalies are somewhat rarer and are confined to only a few river systems. The flooded and dry areas of some representative floodplains are given in Table I.
Less data are available on the composition of the low-water components of the system (Table II) although the figures that are shown indicate the relatively low proportion of the total area that is contributed by the main channel.
TABLE I
Characteristics of some African floodplains
| Floodplain | A1 Area at peak flood (ha) | A2 Area during low water (ha) | 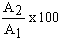 | Authority | |
| Barotse (Zambesi) | 512 000 | 32 915 | 6 | 1 | MacDonald |
| Benue* (Nigeria) | 310 000 | 129 000 | 42 | FAO/UN, 1970 | |
| Kafue flats | 434 000 | 145 560 | 34 | 1 | Kapetsky |
| Massilli* | 1 500 | 200 | 13 | 1 | Barry |
| Niger (Central Delta) | 2 000 000 | 300 000 | 15 | 1 | Barry |
| Niger* (Niger) | 90 704 | 27 000 | 30 | FAO/UN, 1971 | |
| Niger* (Dahomey) | 27 440 | 3 200 | 12 | FAO/UN, 1971 | |
| Niger* (Nigeria) | 480 000 | 180 000 | 38 | FAO/UN, 1970 | |
| Okavango | 1 600 000 | 312 000 | 20 | 1 | Cross |
| Ogun* | 4 250 | 2 500 | 59 | 1 | Tobor |
| Oshun* | 3 740 | 2 000 | 53 | 1 | Tobor |
| Ouémé | 100 000 | 5 170 | 5 | Pers. obs. | |
| Pongolo* | 10 416 | 2 927 | 28 | 1 | Coke |
| Senegal* | 1 295 000 | 78 700 | 6 | 1 | Lessent, Fall, Sere |
Elephant and Ndinde Marshes (Shire)2 | 67 300 | 46 049 | 68 | 1 | Hastings |
| Sudd (Nile) | 9 200 000 | 1 000 000 | 11 | Rzóska, 1974 | |
| Volta* (White Volta)2 | 85 324 | 10 221 | 12 | 1 | Vanderpuye |
| Yaérés (Logone)2 | 460 000 | 2 450 | 0.5 | 1 | Ali Garam |
* Fringing floodplains
1 Replies to CIFA Working Party forms
2 Part of a larger system
TABLE II
Dry season composition of floodplain systems
| Floodplain | River plus channels | Swamp | Lagoon/lake | Total | Dry area (ha) | |||
| Kafue | 5 380 | ( 4) | 130 000 | (89) | 10 180 | ( 7) | 145 560 | 288 440 |
| Shire | 724 | ( 2) | 15 500 | (84) | 6 000 | (14) | 46 049 | 21 251 |
| Pongolo | 427 | (15) | - | 2 500 | (85) | 2 927 | 7 489 | |
| Senegal | 28 100 | (36) | - | 50 600 | (64) | 78 700 | 1 216 300 | |
| Ouémé | 1 402 | (27) | - | 3 768 | (73) | 5 170 | 94 830 | |
() Percentage of total
The essential feature of the floodplain, which distinguishes it from other aquatic systems, is the alternation of flood and dry phases. Flooding may be brought about by two main agencies, (i) local rainfall and (ii) overspill from the river.
Local rainfall and drainage can inundate depressions in the plain independently of any rise in river level. This type of flooding has been noted from the Gambia River by Svensson (1933) and the Kafue by Carey (1971) and is probably common to all systems to a greater or lesser degree. Extensive areas can be flooded in this way and the inundation of the Upper Chari (Bar Salamat and Bar Azoum) appears to be almost entirely of this type (Durand, pers. comm.).
Overspill from the river occurs as a wave of high water travels down the river channel overflowing the banks and levées to flood the low-lying plain (Fig. 4).
The two types of flooding are quite distinct, one resulting in flow from the plain to the river and the other from river to plain. In many regions they may occur at different times of the year. In long rivers, for instance, the arrival of the flood crest may be delayed until the dry season. Such a delay is typical of the Niger River where at the level of Malanville (Dahomey) two floods occur, one in August–November caused by rainfall and drainage of local rivers and a second in December–March due to the arrival of the flood from the headwaters of the river.
The tendency for floods to arrive at different times from the diverse tributaries of large rivers may lead to complex flood patterns or eventually to the effective suppression of the fluctuation in water height.
Typical flood patterns of major African floodplains shown in Fig. 5 indicate that the flooded phase of the annual cycle lasts about six months (range 4.5–8 months). It is however difficult to give absolute figures for flood duration for any one floodplain due to the time taken for the flood crest to pass the length of the area. Fig. 4 shows the hydrological regime of three floodplains where the crest takes about a month to travel downstream from one end of the plain to the other. The same phenomenon makes assessment of absolute flooded areas difficult as the water level may be falling in one part of the system while rising in another and in extreme cases, such as the Okovango, one portion of the plain is dry while the other is flooded.
Hydrological Indices: Apart from simple water height as measured at a gauge, a number of other measures of flood intensity have been used to assess the effects of flooding on the dynamics of fish in the system. Chapman et al. (University of Idaho, 1971) and Dudley (1972) have used a flood index (FI) by measuring the area under a graph of water depth for the flooded phase of the water cycle. Kapetsky (1974) uses a modification of this index to obtain three indices (Fig. 6):
Index 1: the area below a water level curve at which the floodplain initially becomes inundated until it finally becomes dry (shown in black in Fig. 6). This is a measure of the annual extent and duration of the inundation of the floodplain
Index 2: the area above the water level curve for the time when the floodplain becomes dry until it is once again flooded. This index measures the extent of the aquatic environment when the floodplain is dry and includes the duration of the dry phase
Index 3: the area above the water level curve only until the minimum annual water level is reached (indicated by white line in Fig. 6). This is a measure of the extent and duration of the contraction of the aquatic environment after the floodplain has dried.
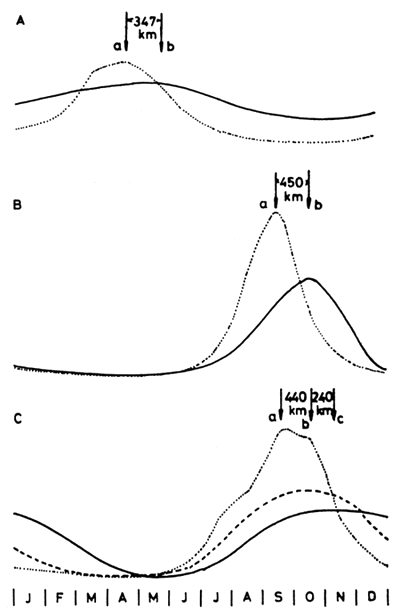
Figure 4 Representative hydrological regimes of three African floodplains
A. Kafue River at (a) Namwala and (b) Kasaka
B. Senegal River at (a) Bakel and (b) Dagana
C. Niger River at (a) Kolikoro, (b) Mopti and (c) Diré
| S. Hemisphere | 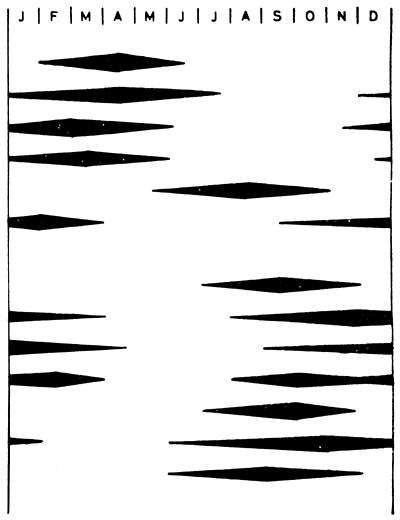 | ||
| Barotse (Zambezi) | |||
| Kafue | |||
| Shirem | |||
| Okovango Upper Basin | |||
| Lower Basin | |||
| Pongolo | |||
| N. Hemisphere | |||
| Benue | |||
| Niger (Central Delta) | |||
| Niger (Niger) | |||
| Niger (Dahomey) | |||
| Ouémé | |||
| Yaeres | |||
| Senegal | |||
Figure 5 Flood regimes of some major African floodplains
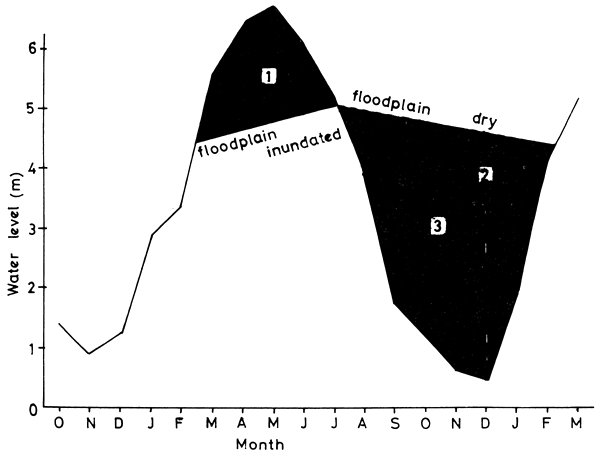
Figure 6 Calculation of the three hydrological indices (from Kapetsky, 1974)
Index 3 is similar to the DDF (Draw-Down Factor) used by Lagler et al. (University of Michigan, 1971) which is the sum of the values for flood storage volume of the Kafue flats for the months September through December.
A moderate amount of data from various African systems indicates the similarities of hydrological behaviour existing between them. Physical and chemical changes follow the flood cycle closely. There is little or no distinction between conditions in the river and on the floodplain during the floods, but differences do arise after the separation of the floodplain pools from the main river channel.
Dissolved oxygen: Dissolved oxygen levels are generally lower in both river channel and floodplain pools during the dry season than during the floods (e.g., Egborge, 1971; Carey, 1971), although variations do occur induced by wind action or by photosynthetic activity of aquatic plants (Holden and Green, 1960). Lowered dissolved oxygen levels can be induced in the river early in the floods by the flushing out of stagnant waters from the swamps (Holden and Green, 1960; Tait, 1967). The smaller pools of floodplains may become completely de-oxygenated in the dry season (Welcomme, 1971) and catastrophic fish mortalities ascribed to wind induced overturn of de-oxygenated lagoon waters, have been described by Tait (1967a).
pH: pH is the most variable of factors between different river systems. In general pH is lower in the flood season and rises in the dry in the river channel (Tait, 1967; Egborge, 1971). Changes in the floodplain pools during the dry season seem to depend on the type of soil; thus in the marshy type 1 swamps of the Gambia system the water was more acid than the river (Johnels, 1954), little difference was noted between pools and river in the Senegal (Centre Technique Forestier Tropical, 1972a) and water in the lagoons of the Sokoto River tended to alkalinity due to the concentration of calcium by evaporation (Holden and Green, 1960).
Conductivity: A general tendency for conductivity to be higher in the dry season than the wet is noted by the Centre Technique Forestier Tropical (1972a) for the Senegal, Duerre (FAO, 1969) for the Barotse plain, Egborge (1971) for the Oshun River and Carey (1971) for the Kafue. This does not necessarily mean that the total quantity of salts is less during the floods, rather it is thought by Holden and Green (1960) that the total remains unchanged but is more diluted due to the greater quantity of water in the system. Anomalies in productivity in Lake Chad (Lamoalle, in press) correlated with the failure of the Yaéré floods might suggest that there is an increase in ions coming down the Chari River which would normally remain in some bound form on the floodplain (Daget, pers. comm.). The role of nutrients in the floodplain/river system, however, remains largely univestigated, particularly in conjunction with variations in primary productivity (see 3.3).
Current: It is perhaps commonly thought that floodplain waters are largely static. This does not appear to be the case as currents of varying intensity flow across the plains during the whole period of inundation. These currents are probably characteristic for the plains and are regulated by the openings in the levée. In some rivers, for example, in the Yaérés, the water spilling over from one river, the Logone, is voided by a second channel, the El Beid In other rivers water entering the floodplain system upstream emerges again many kilometres downstream. Evidently such flow patterns will condition the behaviour of the fish inhabiting the system.
Phytoplankton: Blache (1964) wrote of the Yaérés system of the Logone River that the immediate entry into solution of organic and mineral nutrients (faeces, dried vegetation, ash, etc.) from the soil leads to an explosive increase in phyto-and zooplankton. This view is not upheld by subsequent workers in other areas of Africa who are in general agreement that the maximum production of phytoplankton occurs during the dry season, and that during the floods concentrations are low (Centre Technique Forestier Tropical, 1972a; Holden and Green, 1960; Talling and Rzóska, 1966). In fact, Egborge (1974) found phytoplankton production to be strongly correlated with conductivity and transparency, and inversely correlated with water level and current velocity. The main location of phytoplankton production within a river-floodplain system appears to be in the lagoons after separation from the river (Centre Technique Forestier Tropical, 1966, 1972; Holden and Green, 1960) and Egborge (1974) has found that all forms of phytoplankton except diatoms are dominant in the backwaters of the Oshun River in the dry season. Occasional phytoplankton blooms have been recorded from rivers during the dry season (Carey, 1971). Maximum phytoplankton production therefore appears to occur in the lagoons and pools of the floodplain in the dry season. However, the suggestion has been advanced by Holden and Green (1960) that the total mount of phytoplankton in the system changes little and that there is merely a dilution which results in a reduced density per unit area.
Zooplankton: There is little information on zooplankton production although a marked correlation between this and conductivity has been noted (FAO/UN, 1969). High production of zooplankton organisms has been noticed in larger floodplain lakes during low water, but this remains low in the rivers (FAO/UN, 1970). Zooplankton would therefore appear to follow similar patterns as phytoplankton.
Higher vegetation: The most notable feature of floodplains during the period of rising water is the rapidity of growth of the higher vegetation. Often this is burnt off completely in the dry season, but rapidly colonizes the inundated zone and growth rates of 75–100 cm in a fortnight have been recorded (Kapetsky, pers. comm.). Estimates of production of grasses indicate a mean weight of 16 000 kg/ha after about 5 weeks of flooding (Kapetsky, 1974). Correlated with the increased amount of vegetation is the periphyton, including filamentous algae, which appears attached to most surfaces (Carey, 1971; personal observation), and it appears likely that during periods of high flow where adequate supports are to be found, periphyton (or “Aufwuchs”) replaces the phytoplankton at least to a certain extent.
Other invertebrates: Insect and other invertebrate populations of floodplains remain relatively poorly studied. Carey (1967) states that the largest concentrations of macro-invertebrates was found in weeds in lagoons, but that they were generally widely distributed and abundant in flooded areas. Personal observations have shown the enormous increase in macro-invertebrates that are possible during the flooded phase, when on three successive floods in the Ouémé River, pulmonate gastropods appeared in great quantities during the second month of flooding and persisted until the end of the floods; these organisms are generally absent from the environment during the dry season.
It seems that little of the traditional richness of the floodplain is discernable from either the chemical changes in the water or the plankton production during the flooded part of the cycle. Indeed both these factors would seem to indicate greater productivity at low water. However, this period is one of intense growth activity for higher vegetation and it is possible that the utilization of salts by the growing plants is so rapid that few free ions enter the aqueous component of the system. Growth of fish, and also aquatic macro-invertebrates is equally rapid and although few species are planktonophage, an adequate source of nutrient must be available to sustain their growth. This would appear to be located in the detritus and possibly in the epiphytic organisms. In general little information exists on food chains and energy inter-relationships of floodplain organisms and the subject deserves considerable further study.
Apart from the use of the permanent waters of the river channel and floodplain pools for fisheries, the plain itself performs a number of functions which may influence the productivity of the aquatic phase. In its wild state the plain is colonized with grasses, which support populations of game animals including hippopotamus and on some plains the wild population is now maintained in parks or reserves. The dung dropped by such animals remains on the plain as a depository of organic material. In many areas the grasses are burnt off by natural or humanly induced fires toward the end of the dry season. This practice renders the organic components of the dessicated grassland into inorganic ash, thereby probably making it more easily available during the floods and also reducing the de-oxygenation which would result from the immersion of decaying grasses. Grazing is widely practised on the plain and traditional nomadic cycles have grown up around such areas, particularly the Central Delta of the Niger and Sudd of the Nile. The dung of the cattle performs a similar function to that of the game animals. Various forms of agriculture are common but by no means as widespread as they might be due to the difficulties of life in such areas. Only on certain rivers, such as the Ouémé, do stilt villages exist which are adapted to life during the flood period. Maize, cassava, millet and a variety of vegetables are grown, and in some areas small irrigated agriculture projects have been initiated. Rice is moderately common but harvesting and most of the growth occurs during the high-water phase. Agriculture may effect the ecology of the plain adversely through the use of insecticides and the modification of the flood regime by irrigation schemes. The role of fertilizers remains obscure although these could increase productivity of both dry and wet season phases.
The dry season uses listed by respondants to the CIFA questionnaire are given in Table III
TABLE III
Dry season uses of floodplains
| Floodplain | Agriculture | Grazing | Game or forestry reserves | ||
| Rice culture | Vegetable gardening | General | |||
| Kafue | x | x (58%) | x (2%) (93 000 head of Lechwe) | ||
| Massili | x | x | |||
| Shire | x | x | x (148 000 head) | x | |
| Gambia | x (50%) | x | x | x (300 000 head) | |
| Central Delta | x (1%) | x | |||
| Okavango | x | x | x | ||
| Ouémé | x | x | x | ||
| Ogun | x | x | x | ||
| Oshun | x | x | |||
| Pongolo | x | x | |||
| Senegal | x | x | x | x | x |
| Yaérés (Logone) | x | x | |||
In general fish are concentrated in permanent bodies of water in the dry season and spread over the floodplain in the floods. Some species, however, appear to remain in the main river channel throughout their life in some systems. In the Niger, for example (Daget, 1954), there are a number of pairs of homologous species within the different genera, one form of which breeds in the floodplain, the other in the river channel.
Many species commonly encountered on the floodplain have adaptation to survival in low dissolved oxygen conditions. These include: cocoon formation in Protopterus sp. and dormant eggs among various Nothobranchius and Aphyosemion species, both of which permit the survival of the species over dry periods. External gill filaments are present in the young of both Heterotis and Gymnarchus and complete external gills are found in Protopterus and Polypterus. Species of Clarias, Heterobranchus, Ctenopoma, Parophiocephalus and Protopterus all have accessory breathing organs which permit aerial respiration. Such species form a group that is well adapted to swamp life and tend to concentrate in the more de-oxygenated small pools and swamps of the floodplain during low water, when other more active species are to be found in the river or larger lagoons. An example of such associations is given in Table IV for three size groups of floodplain pools in the Ouémé River (data from unpublished personal observations).
Migration of fish within tropical river systems are a well established phenomenon and have been widely described from all areas of Africa. Daget (1960) identifies two components of such movements:
lateral migrations onto and off of floodplains.
Each of these requires a different type of behaviour and probably different sets of physiological stimuli and both must be regarded as “active” migrations. The movement of fish upstream against currents is well known (Whitehead, 1959; Matagne, 1950; Welcomme, 1969), but fish entering the floodplain frequently have to move against equally strong current (Johnels, 1954).
Various authors (Blache, 1964; FAO/UN, 1970; Williams, 1971) classify movements into four main phases correlated with flood regime:
rising water - waters confined within banks; at this time fish may undertake longitudinal migrations within the river channel, either from parts of the river with no floodplain or from adjacent lakes
rising water - waters extending over floodplain; the fish spread by lateral migration over the floodplain
falling water - water draining from floodplain; as the flooded area diminishes there is a movement to the river and other permanent water bodies
falling water; once arrived in the river the fish disperse to the dry season habitats.
Following the dispersal there is apparently a period of stability with little movement.
The longitudinal migratory phase in the river may be particularly spectacular, and intense migrations of potamodromous species, particularly of the genus Labeo have been recorded both for the Nzoia and Yala Rivers, Kenya (Cadwalladr, 1965) and for the Luapula (Matagne, 1950), where the fish ascend the rivers from lakes. In these cases there is an additional lacustrine component to the migration.
TABLE IV
Differences in species composition of catches from permanent floodplain pools of different areas
| Species | Percentage representation by weight | |||
| Small pools (up to 500 m2) | Medium pools (500–5 000 m2) | Large pools (over 5 000 m2) | ||
| Swamp dwelling | ||||
| Clarias ebriensis | 72.2 | 20.0 | 1.3 | |
| Clarias lazera | 5.0 | 13.6 | 3.4 | |
| Ctenopoma kingsleyae | 0.9 | 7.2 | P | |
| Gymnarchus niloticus | - | P | 2.1 | |
| Heterotis niloticus | - | 26.0 | 2.6 | |
| Parophiocephalus obscurus | 23.8 | 27.2 | 1.6 | |
| Polypterus senegalus | P | 0.3 | 0.7 | |
| Protopterus annectens | P | 0.8 | 0.7 | |
| Xenomystus nigri | P | 0.2 | P | |
| Generally distributed | ||||
| Citharinus latus | - | 0.1 | 1.2 | |
| Distichodus rostratus | - | 0.7 | 8.1 | |
| Hepsetus odoe | - | 2.3 | 2.6 | |
| Hemichromis (2 species) | P | P | P | |
| Hyperopisus bebe | - | P | 5.4 | |
| Lates niloticus | - | - | 10.1 | |
| Labeo senegalensis | - | - | P | |
| Mormyrops deliciosus | - | - | 18.4 | |
| small mormyrids | - | - | 18.4 | |
| Pelmatochromis güntheri | P | P | P | |
| Schilbe mystus | - | - | 6.0 | |
| Synodontis (2 species) | - | - | 15.2 | |
| Tilapia (4 species) | - | 1.6 | 2.2 | |
P = Present at less than 0.1 percent
That movement in rivers can be extensive is demonstrated by Williams (1971) who tagged fish in the Kafue River some of which were returned from up to 120 km away. Fish from the Central Delta of the Niger also migrate for great distances (up to 400 km - Daget, 1957) and reached the area around Bamako before the construction of the Markala Dam interrupted their passage (Daget, 1960a). Similar long-distance migrations have been noted for the Chari/Lake Lake Chad system (Durand, 1970) and the Nzoia/Lake Victoria system (Cadwalladr, 1965).
There is evidence that migrations onto and off of floodplains are not random and that the different species tend to arrive at characteristic times and in particular groups. Thus, Williams (1971) and the University of Idaho (1971) note that Clarias spp., Schilbe mystus, Barbus spp., and Tilapia spp.1 begin migration earlier than other species in the Kafue River and in some Tilapia females tend to migrate earlier than males. Migration off of the floodplains are even more specifically segregated. There is firstly a tendency for adult fish to leave the plain early, as soon as there are indications of diminishing water level (FAO/UN, 1970). Secondly the young fish leave in a precise order. Fishes sensitive to de-oxygenated conditions, such as Alestes, leave the plain first and other more resistant genera such as Clarias or Polypterus at a later date (Daget, 1957). Different species of the same genus may also have different migration times and Welcomme (1969) has shown that two species of Barbus behave very differently in this respect. Durand (1971) also demonstrates that fish leaving the Yaérés floodplain by way of the El Beid River are associated in groups each with a characteristic time of migration.
1 Including Sarotherodon sp. (Trewavas, 1973)
The reproductive cycle of riverine fish species in Africa is closely allied to the seasons, reproduction occurring almost universally just prior to or during the floods (Blache, 1964; Daget, 1954; Svensson, 1933; Carey and Bell-Cross, 1967; FAO, 1968; Durand, 1970). Fish normally reproduce in the grass swamps at the edge of the advancing floods (Daget, 1957) although a number of species including Heterotis niloticus, Gymnarchus niloticus and Hepsetus odoe, construct floating nests which permit deeper-water spawning. Some species, however, may spawn in the river channel before the water overflows onto the floodplain. This has been noted particularly for Clarias and Tilapia species in the Niger (Daget, 1957). Cichlids from the Kafue River may reproduce before the floods and carry eggs and fry onto the floodplain (Williams, 1971; University of Idaho, 1971; Dudley, 1972) and personal observation also shows this to be true of Hemichromis and Tilapia in the Ouémé River, Dahomey.
Reproductive activity continues for a number of weeks and in some species throughout the entire flood season although it clearly peaks during the earlier phases of flooding and stops entirely by the end of the floods (Daget, 1957). Flooding appears to be essential to the completion of the reproductive cycle of most species and the failure of the floods due to the Sahelian drought has brought about a decline in reproductive success of fish, both from the Central Delta of the Niger (Macher - provisional report) and Lake Chad (Stauch, pers. comm.).
There are some indications that the intensity of flooding influences reproductive success as stronger year classes have been noted from those years when particularly intense floods occurred in the Kafue River (Dudley, 1972).
The feeding habits of many species of riverine fish have been widely described (Daget, 1954; Svensson, 1933; Blache, 1964; Carey, 1971; FAO/UN, 1970) and the adults cover the complete range of trophic types from previous predators to planktonophage, although species fee feeding solely on plankton are generally rare in rivers. Juvenile fish appear to feed mainly on periphyton, detritus, zooplankton and small insects. There is considerable seasonal variation in feeding intensity correlated with the flood regime. During the floods the release of nutrients, rapid growth of vegetation and increased availability of other sources of food such as seeds, young shoots, leaves, insects and molluscs forms the basis for a particularly intense feeding activity (Blache, 1964). Most fish are in peak condition during this season and this is reflected in high condition factors and abundant fat deposits in the body cavity (FAO/UN, 1970). During low water feeding intensity is reduced (Johnels, 1954 Daget, 1957, 1960) and in certain areas may cease altogether. This phenomenon, at least in part, is responsible for the formation of clear rings on the scales of many riverine species.
According to some authors (Johnels, 1954; Kapetsky, 1974) piscivorous predators are an exception to the above pattern, showing reduced feeding during the initial stages of flooding when fish are widely dispersed on the floodplain and are protected by the heavy growth of vegetation, and increased activity during low water when individuals are crowded together into a diminishing volume of water.
Mean growth rates are given for several species of riverine fish by FAO/UN (1970) and Kapetsky (1974). However, these are very variable and change both seasonally and from year to year.
Seasonal fluctuations: Changes in growth rate may be expected during the year due to the expansion and contraction of the aquatic environment, seasonality of temperature and rainfall and flood-associated changes in the abundance of food (Kapetsky, 1974). Growth rates are usually slower in the dry season (Daget, 1957; Johnels, 1954) and Dudley (1972) found that about 75 percent of the expected first-year growth of juvenile Sarotherodon andersoni and S. macrochir occurs within six months after spawning (about six weeks after peak floods on the Kafue River). Kapetsky (1974) also found little growth in length during the dry season, especially during the period of maximum drawdown.
Annual fluctuations: Variations in intensity and duration of flooding give differences in growth from year to year. University of Idaho (1971), Dudley (1972) and Kapetsky (1974) have all found high correlations between various indices representative of flood intensity (FI and index 1 - see 3.1), severity of drawdown (indices 2 and 3) and temperature in the Kafue system on first and second-year growth increments of various Cichlid fishes (Table V). In general growth is positively correlated with the intensity of flooding and poor growth results from more severe drawdown conditions.
Very little detailed information is vailable on mortality rates of riverine species in Africa, particular with regard to seasonality.
Annual: Daget et al., (1973) found annual values of Z = 2.691 and 2.656 for two zones of the Bandama River, but few definite conclusions as to the true mortality rate of the fish could be drawn from these figures as they represent total loss from the area, including loss by emigration. In fact a third zone of the river had a negative value of Z, indicating considerable migration into the area. Figures are quoted by Kapetsky (1974) for several Cichlid species ranging from 99 percent (Z = 4.6090) for third-year T. rendalli to 33 percent (Z = 1.7918) for seventh-year S. andersoni, although it is felt that mortalities as high as 92–99 percent are slightly over-estimated. Nevertheless these figures, together with those of Daget indicate the high mortality rates that are to be encountered in riverine species.
Seasonal variations: Due to the severity of conditions when water areas are diminishing and fish populations becoming more concentrated, mortality rates in the dry season may be expected to be higher than during the flood phase. At this period fish are more vulnerable both to natural mortalities from predation and environmental stress, and to fishing mortalities induced by the greater accessibility of the stocks. Observations by the University of Michigan (1972) indicate the high losses encountered in floodplain lagoons (between 75 and 85 percent over 3 months) and an overall decrease in biomass of 40 percent is estimated by these workers for a three-month period (July–September 1970) on the Kafue system. The Kafue figures and those of Daget et al. (1973) are very similar, indicating dry season losses of about 15 percent per month.
TABLE V
Parameter estimates for simple and multiple linear regressions of first and second year growth increments of various cichlid species on temperature and on hydrological indices
| Species | Year of growth | Sex | Model | R | Authority |
| S. andersoni | 1 | M | Growth = 0.02 FI + 12.87 | 0.92 | Dudley, 1972 |
| 1 | M | TL(mm) = 146.51 - 0.11(H12) | 0.94 | Kapetsky, 1974 | |
| 1 | F | Growth = 0.014FI + 13.4 | Dudley, 1972 | ||
| 2 | M | TL(mm) = -29.47 + 1.98(TI) | 0.90 | Kapetsky, 1974 | |
| 2 | F | TL(mm) = 38.24 - 0.30(H13) + 0.83(TI) | 0.93 | Kapetsky, 1974 | |
| S. macrochir | 1 | M | Growth = 0.2FI + 11.02 | 0.9 | Dudley, 1972 |
| 1 | M | TL(mm) = 130.39 - 0.13(H12) | 0.92 | Kapetsky, 1974 | |
| 1 | F | TL(mm) = 130.13 - 0.32(H12) | 0.85 | Kapetsky, 1974 | |
| 2 | M | TL(mm) = 74.72 - 0.10(H13) | 0.58 | Kapetsky, 1974 | |
| 2 | F | TL(mm) = 14.69 - 0.18(H13) | 0.95 | Kapetsky, 1974 | |
| T. melanopleura | 1 | M | Growth = 0.029 FI + 12.8 | 0.80 | Dudley, 1972 |
Kapetsky (1974) models growth and mortality for floodplain systems by rotating and reversing traditional exponential models. This gives low mortality rates for the initial part of the year (flood season) and an increasing mortality toward the end of the year (dry season). While this model is a useful approximation for the purpose of calculating production, it appears likely that mortality curves consist of two or three different components corresponding to the different phases of the flood cycle.
Variation between years: There is, at present, no information on variations in mortality rate between years. It is however known that periods of more intense flooding, which produce better recruitment and growth, also produce improved survival (University of Idaho, 1972). It also appears likely that differences in the amount of water remaining in the system in the dry season, together with its duration, would strongly influence survival from one year to the next. This would also depend on the intensity of the preceding flood as an intense flood with good spawning success followed by a prolonged dry period may result in a greater mortality than would occur during the same dry period if preceded by a poor spawning.
In general, little is known of the seasonal and annual variations in mortality rates, and prolonged observations on several systems are needed to clarify this position to any degree.
Many functions of the biology of fish living in floodplain systems vary with the hydrological cycle. Within any one year a period of high recruitment and growth rate, intense feeding and low mortality rate during the floods alternates with a period of high mortality, low growth and negligible feeding and reproduction. Variations in the intensity and duration of the floods, and the severity of drawdown conditions during low water produce corresponding fluctuations in many biological parameters. Thus, year-classes from years of good flooding and slight drawdown show greater numerical strength, growth and survival than year-classes from years of poorer flood conditions.
A great range of fishing methods are used on African floodplains and in rivers (Stauch, 1966; FAO, 1969; Welcomme, 1970; Centre Technique Forestier Tropical, 1972; Ratcliffe, 1972). Most of these are, however, general methods which are practised almost universally through-out the continent. The purpose of this chapter is to examine those methods which profit by some morphological feature of the river/floodplain system or from the behaviour patterns of the fish on them. These generally fall into two categories: (i) those methods which use the confinement of fish within limited areas during low water; and (ii) those methods which exploit the migrations of the fish for their capture.
Dry season: During the season when fish are confined to the permanent lagoons, swamps and river channels, there is intense fishing activity. Communal fish drives are common in the floodplain pools and are described from many regions of Africa. Poisoning of pools is also well known and generally such areas are fished out completely at some time during the period. Fishing in the major river channel is usually pursued with a wide variety of nets, traps and long lines. The effectiveness of the fishery in the dry season varies inversely with the amount of water remaining in the system. In periods of extreme drawdown fish are more concentrated and easier to capture. In this way fishing mortality increases with increases of HI2 as well as natural mortality.
Period of rising water: Initially fishing activity is intense and is particularly aimed at the capture of fish migrating on the river channel. As the floods increase, the technical difficulties of fishing in the high currents and the dispersion of the fish over wide areas of floodplain make yields low and in most parts of Africa activity is at a minimum during this period. Effectiveness of fishing depends largely on the intensity of the initial stages of flooding. Rapid and high floods disperse the fish more quickly and currents interfere with the efficient working of some types of gear. Thus catches tend to be negatively correlated with HI1 (Muncy, 1973).
Period of falling water: As the waters decrease in area fish become concentrated into depressions of the floodplain and into the main drainage channels. At this time there is once again a very active fishery aimed at the capture of migrating fish particularly juveniles and attempts are made to retain fish on the plain by dams or ponds.
Fishing by damming creeks has been described from the Gambia River (Svensson, 1933; Johnels, 1954) where the exit streams from the swamps are blocked by earth dams. Water is than allowed to drain through a hole in the dam where a trap is placed for the capture of the fish. Such weirs prevent the draining of the water at normal speed and the process is prolonged. Reed (FAO, 1969) describes a similar experimental method applied to the floodplain of the Niger River, where exit streams from swamps are blocked by earth dams thus retaining more water than usual (Fig. 7). The dams are left in place for several weeks allowing an increased time for the growth of the fish. In the dry season the dam is breached and traps placed in the opening. A doubling of the normal yield of the swamps has been achieved by this method.
Fishing in the permanent lagoons and ponds of the floodplain is common and in the Delta floodplain of the Ouémé River the number of such natural features of the floodplain has been considerably augmented by trench-like fish holes “whedos” which have been excavated to a depth of about 1 m in a network all over the plain (Welcomme, 1971). On average, these cover about 3 percent of the dry area of the plain and in some areas reach very dense concentrations (Fig. 8). The ponds are fished, after removal of the vegetation covering the water surface, with the aid of fences which are advanced in such a manner as to confine the fish in a small area from which they are removed. Yields from these will be discussed in Section 7.
The capture of fish migrating within the channels draining the floodplain has been described from several parts of Africa. This is principally done with barriers or weirs of the type described by Whitehead (1958); however, of these fisheries two can be cited as representative:
The “maalelo” fishery of the Barotse floodplain, which consists of long low earth and grass dams in which conical valve basket traps are placed, has been described by FAO (1968) and FAO/UN (1969; 1970a) and Bell-Cross (1971). This fishery crops mainly juvenile fish on their migration to the river.
Cross channel traps of the El Beid River are described by Durand (1970). Here dams made of enmeshed branches and tree trunks completely block the main river channel diverting the flow over the shallow fringing plain where a series of V-shaped enclosures are constructed. The weir is fished with a type of clapnet designed to seal the space between the enclosures. Weirs are installed in such density as to almost completely bar the passage of the fish migration from the Yaérés floodplain to Lake Chad. The majority of the fish caught are juveniles in their first year.
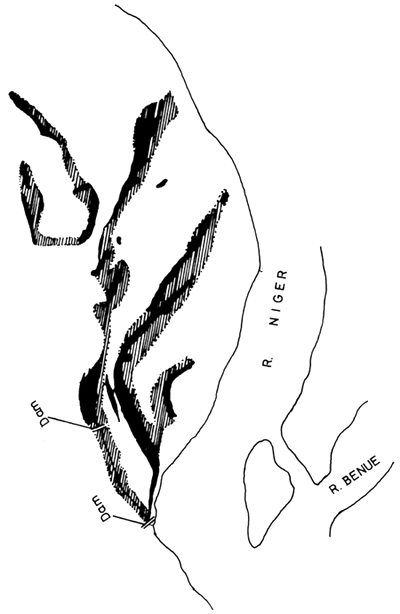
Figure 7 Tracing from aerial photographs of the Niger floodplain retention dams, showing the original water area (solid black) and the area flooded after the closure of the dams (hatched). (After FAO 1969)

Figure 8 Portion of the Ouémé floodplain showing the
distribution of the “whedo” floodplain pools.
White areas are pools that have been fished;
unfished vegetation-covered areas may also be
seen.
(Air photograph by courtesy of I.G.N., Paris)
Kapetsky (1974) derives the productivity of five species of fish from the Kafue River using a “floodplain” model for mortality and growth as detailed in Section 5.6. The total figures for fish of age two and older range from 76 kg/ha/year and 56 kg/ha/year for S. andersoni and S. macrochir, two herbivorous and long-lived species, to 12 kg/ha/year and 6 kg/ha/year for Hepsetus odoe and Serranochromis augusticeps two short-lived predators. A very large percentage of the production (85–99 percent) of fish older than age two occurs during the third year of life. As a general rule, the production during the first two years of life contributes a substantial protion of lifetime production. Thus, the total lifetime production of the five species investigated by Kapetsky is probably considerably superior to the 199 kg/ha/year estimated for fish of age two and older.
A limited number of standing stock estimates have been made either by poisoning or exhaustive fishing in several river and floodplain areas.
Main river channel: During high water the standing stock of fish in the main river channel is assumed to be similar to that of the open water lagoon of the floodplain (University of Michigan, 1971), i.e., 337 kg/ha. During low water Kapetsky (1974) found 106, 386 and 576 kg/ha for three stretches of the Kafue River and a mean value of 348 kg/ha for the whole river.
At the beginning of the dry season estimates of 341, 227 and 141 individuals/ha were made from three reaches of the Bandama River by Daget et al. (1973). The equivalent ichthyomass for one part of the river in January was 125 kg/ha and in May estimates of 50, 177 and 305 kg/ha were made.
Lagoons and floodplain: During high water the University of Michigan (1971) report an ichthyomass of 2 682 kg/ha from the vegetated waters of the lagoon, 337 kg/ha from open waters and 64 kg/ha from grass marsh.
During the dry season when the water had fallen and the lagoons separated from the main channel, estimates of 3 306 and 2 693 kg/ha were made for two different lagoons. Within three months these had fallen to 684 and 501 kg/ha respectively. Kapetsky (1971) found the largest concentration of fish (879 kg/ha) in vegetated lagoons and lower concentration of 575 kg/ha in open water lagoons.
The University of Michigan (1971) estimates were combined to give estimate of total ichthyomass in dry and wet season for the Kafue system (Table VI).
TABLE VI
Estimates of ichthyomass for dry and flood seasons for the Kafue Flats system /after University of Michigan (1971)/
| 1970 | Ichthyomass (metric tons) | |||
| Lagoon | Floodplain | River | Total | |
| High water June–July | 85 374 | 8 704 | 1 786 | 95 864 |
| Low water September–October | 56 426 | - | 979 | 57 405 |
Holden (1963) found that the standing stock in floodplain pools of the Sokoto River varied widely with bottom type. The intermediate (mixed sand-mud) bottomed pools supported 1 029 ± 536 kg/ha; sand bottomed pools 475 ± 148 kg/ha and mud bottomed pools 144 ± 83 kg/ha. The mean ichthyomass from all types was 415 kg/ha which spread over the whole floodplain (of which the pools comprise 3 percent) amount to 12–17 kg/ha. Differences in pool form were also found to influence ichthyomass in the Senegal River (Centre Technique Forestier Tropical, 1972a). Here yields were high from elongated channel type pools (375–200 kg/ha) and much reduced from circular pools 1.5–20 kg/ha. Mean yield was low (37.5 kg/ha) from this area and a total tonnage of 260 tons was estimated for the 6 600 ha of floodplain pools.
The most complete studies carried out on floodplain pools have been done on the artificial whedos of the Ouémé delta. Here observations by a number of workers have been summarized by FAO (1971) and are here presented (Table VII; Fig. 9) for comparison with other areas. Ichtyomass is generally high although it is of the same order as that found in the intermediate pools of the Sokoto and some Kafue lagoons.
TABLE VII
Standing stock of some “whedo” floodplain pools of the Ouémé floodplain
| Year | No.of samples | Yield (t/ha) |
| 1955 | 17 | 2.06 ± 1.14 |
| 1956 | 14 | 2.14 ± 1.39 |
| 1958 | 3 | 2.13 |
| 1968 | 2 | 1.59 |
| 1970 | 32 | 1.57 ± 0.41 |
The total ichthyomass in the floodplain pool system of the Ouémé is estimated at between 2 255 and 1 719 tons for the 1 095 ha of whedos.
As mentioned by Holden (1963) the high yields of floodplain and pool fisheries, and particularly the whedos, are achieved by the “drain-in” principle which concentrates fish from the whole floodplain area. If the ichthyomass of the fish from the pools is extended over the whole area of the plain, the standing stock is between 61.8 and 47.1 kg/ha.
The catch from 12 floodplains is listed in Table VIII.
The mean catch from all floodplains was 32.1 ± 24.2 kg/ha/year. There is however reason to believe that the Barotse and Okavango floodplains are considerably underfished. Similarly Motwani (FAO/UN, 1970) report that the present levels of yield are only moderate with respect to potential in the Niger and Benue Rivers in Nigeria. Of the other floodplain mentioned in the table, all appear to be fished at a high level. Considering only these, a mean catch of 50.4 ± 19.24 is estimated. It seems, therefore, reasonable to expect catches of this order from most floodplain areas (i.e., a potential catch of 834 000 tons for the floodplains listed in Table I).
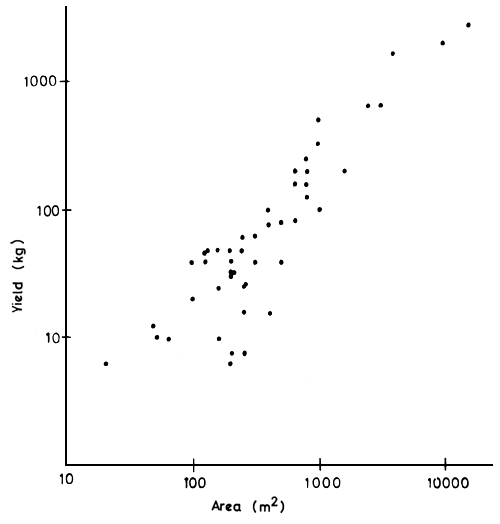
Figure 9 Relationship between surface area and yield in the “whedo” floodplain pools of the Ouémé delta
TABLE VIII
Commercial catch from 12 African floodplains
| Name of floodplain | Area at peak flood (ha) | Catch (t) | Yield (kg/ha) | |
| Barotse | 512 000 | 2 395 | 4.68 | |
| Kafue Flats | 434 000 | 6 078 | 14.00 | |
| Shire | (total) | 140 000 | 8 972 | 64.08 |
| Central Delta | 2 000 000 | 134 000 | 67.00 | |
| Massilli | 1 500 | 465 | 31.00 | |
| Okavango | 1 600 000 | 800 | 0.50 | |
| Ouémé | 100 000 | 6 484 | 64.84 | |
| Senegal | 600 000 | 36 000 | 60.00 | |
| Niger | (Niger) | 90 704 | 5 200 | 57.33 |
| (Dahomey) | 27 440 | 1 200 | 43.73 | |
| (Nigeria) | 480 000 | 14 350 | 29.90 | |
| Benue | 310 000 | 9 570 | 30.87 | |
Two sets of data exist upon which an assessment of the effect of differences in hydrological regime upon the fish catch can be based. Neither of these sets of data have any reliable measure of effort; however, extremely good correlation between some of hydrological index and catch can be obtained.
Kafue flats: Water level and catch records are available for this region from 1954 until the present. However, the fishery is not judged to have reached its maximum expansion until 1958 and the hydrological regime of the river was altered by a downstream dam in 1972. Calculations have therefore been based largely on the years 1958–71. University of Michigan (1971) found a correlation (r = 0.77) for catch against the amount of water retaining in the system represented by Drawdown (DDF) in the preceding season with the following relationship: Y'n = -6 630 + 1 830 loge DDFn - 1. University of Idaho (1971) found correlations of a similar order for Flood Index (FI) as a representation of intensity of flood. These were T = 0.72 on the preceding year and 0.71 on two years previously. Muncy (1973) presents a complete analysis of these factors. In summary he found a very weak negative correlation (r = -0.171) for water stage and catch in the same year. This is attributed to a lessened efficiency of fishing during more intense floods. A good positive correlation r = 0.61 exists between catch and the flood intensity of the preceding year which is improved to r = 0.74 if a log-log plot is adopted. Good correlation r = 0.70 also exists between catch and the flood intensity two years previously.
Shire River (Elephant and Ndinde Marshes): Five years catch data and eight years hydrological data have been supplied by Dr. Hastings of the Shire Valley Agricultural Development Project. Analysis of these show a good correlation r = 0.84 to exist between catch and the flood regime (HI1) of the previous year (Fig. 10). No significant correlations are obtained with HI1 of the same year or two years previously. the regression catchn = 60.98 + 29.57 HI1n-1) is not improved by calculation in the logarithmic form (log Catchn = 3.54 + 0.21 log HI1n-1) as r here = 0.68. A weaker, but still significant, correlation exists between Catch and Hydrological indices 2 and 3. Here logarithmic relationship F12 log Catchn = 4.3-0.16 log HI2n-1 has a correlation coefficient r = -0.54 and the arithmetic form Catchn = 100.37 - 4.47 FI2n-1 has a weaker correlation r = -0.43.
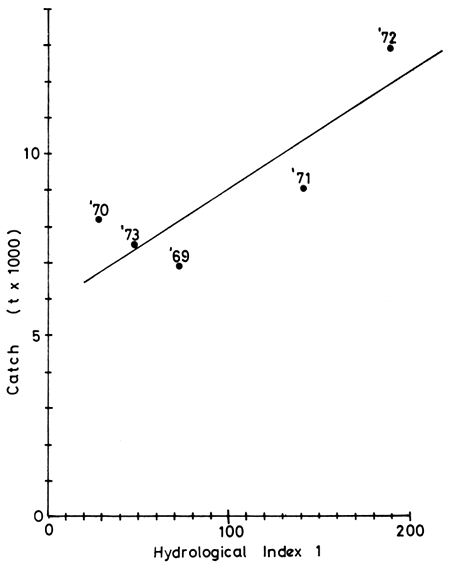
Figure 10 Relationship between Hydrological Index 1 and
Catch in the River Shire.
Regression line Catch = 6098+29.57 (HI1 previous year)
The above results, while still being rather imprecise, indicate clearly that there is a relationship between the hydrological regime and catch although much further data on other systems is needed to define the relationships more fully. The main interest here centres on whether the factor responsible for the fluctuation is in fact the intensity of the flooding or the severity of the drawdown period (or some complex of both). The similarity of the correlation obtained in the Kafue is hardly surprising as intensity of flooding (FI1) is inversely correlated with severity of drawdown (FI2) (r = -0.78). The correlation between these factors in the Shire is much lower (r = -0.45) which may account for the lowered correlation of catch with FI2. A further factor influencing this may be the area of water remaining in the system relative to that at peak floods. In the Kafue this is about 34 percent whereas, in the Elephant and Ndinde Marshes, it is 68 percent. It may be supposed that the more stringent the drawdown, as reflected by lessened percentage of water remaining in the system during the dry season, the greater the influence on catch of fluctuation in this factor.
As may be expected from the biology of the fish (Section 5) the productivity and yield patterns of floodplain fisheries fluctuate considerably both during one year and between successive years. Populations of fish increase through the flood season reaching a maximum at peak floods. Populations then decline both numerically and by weight during the dry season reaching a maximum just prior to the next flood. Between year fluctuations in catch (and probably in population) can be correlated with differences in flood intensity in previous years, the most influencial being that of the year preceding the current year. Similarly the degree of drawdown on the amount of water remaining in the system would appear to exert an effect. Thus, years of optimum catch follow one or more years of good flooding and here the amount of water remaining in the system during the dry season is relatively abundant.
![]()
![]()
![]()