![]()
![]()
![]()
par
Veikko Koski
The Finnish Forest Research Institute
Unioninkatu 40 A
SF-00170 Helsinki 17
Finlande
INTRODUCTION
Les buts assignés aux vergers à graines sont de produire des semences d'essences forestières génétiquement améliorées, favoriser de fréquentes récoltes de graines, et faciliter la récolte. Sur un plan international, ce n'est qu'assez récemment qu'ont été publiées des règles précises sur les normes minimales pour les vergers à graines, sous la forme de directives pour les pays participant au Système de l'OCDE 1 pour le contrôle des matériels forestiers de reproduction destinés au commerce international (OCDE 1974). Bien que ces règles spécifient que l'autopollinisation et la contamination par un pollen étranger doivent être réduites au minimum, elles ne donnent aucune directive chiffrée et ne fixent aucune exigence précise. La responsabilité de la définition de critères spécifiques pour les vergers à graines appartenant aux diverses catégories reconnues dans le Système est laissée aux autorités nationales chargées de son application.
Il est normal que l'on ne puisse énoncer des exigences détaillées universelles qui soient communes à tous les vergers à graines; il faut prendre en considération les diverses catégories et les divers types de vergers, les caractéristiques biologiques des diverses essences, les conditions de milieu. D'un autre côté, nos connaissances en matière de génétique quantitative de même que de biologie de la floraison pour de nombreuses essences forestières s'est accrue de manière appréciable depuis que le concept de verger à graines a été reconnu comme partie intégrante des activités de génétique forestière. En fait, diverses études en provenance des pays scandinaves indiquent que le fonctionnement d'un verger à graines en tant qu'unité génétique peut être étudié quantitativement (Sarvas 1970, Eriksson et al. 1973, Jonsson et al. 1976, Koski 1975, Bhumibhamon 1978, Chung 1981).
Une législation imposant l'enregistrement officiel des vergers à graines et autres sources de matériel forestier de reproduction est déjà en vigueur dans de nombreux pays à une échelle nationale, et le nombre de ces pays a des chances de s'accroître dans l'avenir. Si l'on veut que la notion de semences génétiquement améliorées soit respectée, il importe que certaines garanties soient assurées en ce qui concerne la composition et les caractéristiques des lots de semences fournis en conformité avec cette législation. Dans la plupart des cas, une certification par le moyen d'une évaluation réelle de tests de descendance suffisamment âgés signifierait un délai d'une à plusieurs décennies. Pour éviter de tels délais, les chercheurs forestiers sont souvent contraints de se reposer sur la théorie génétique dans une mesure plus forte que leurs collègues travaillant dans le domaine de l'agriculture ou de l'horticulture.
Divers facteurs influent sur la composition génétique des semences récoltées dans un verger à graines. Nombre d'entre eux peuvent être chiffrés, tels que le nombre de génotypes présents dans le verger, l'intensité de la sélection, la probabilité de consanguinité, le degré de contamination par un pollen étranger. En se basant sur des études antérieures, Sarvas (1970) et Bhumibhamon (1978) ont proposé d'utiliser un système d'indice pour l'inscription des vergers à graines. Chaque facteur contribuant à l'indice serait évalué séparément suivant une échelle de points, le nombre total de points déterminant la valeur génétique des semences récoltées dans le verger à graines, ce qui permettrait un classement homogène des différents vergers.
Bien qu'un système d'indice soit plus objectif qu'un jugement intuitif, il a des limitations dans l'application pratique: une notation élevée pour un facteur peut compenser une valeur basse inacceptable pour un autre facteur. On peut par conséquent se demander si les différentes valeurs obtenues pour cet indice reflètent bien la valeur génétique du matériel de reproduction et sa valeur monétaire, même relatives. L'auteur de cet article estime qu'il est plus sûr et plus pratique de i) classer les vergers à graines (exemple: vergers clonaux de Pinus sylvestris; vergers de semence de P. sylvestris; vergers clonaux de Picea abies, etc.); ii) sur la base de cette classification, définir les exigences minimales pour certains facteurs mesurables qui influent sur la qualité génétique des semences.
Parmi les différents types de vergers à graines identifiés par le Système OCDE (voir par ex. Barner 1976), les vergers à graines clonaux de première génération, qui se rangeraient dans la catégorie de l'OCDE “Vergers régionaux” 1, sont actuellement parmi les plus courants à l'échelle mondiale. C'est également le cas pour la Finlande, qui a officiellement adhéré au Système OCDE en 1980 (Hagman 1982).
Des exigences minimales pour les vergers à graines clonaux de Pinus sylvestris en Finlande ont récemment été arrêtées et publiées (Koski 1980). Ces directives sont maintenant en vigueur. Il est fort probable que des directives techniques analogues pour le contrôle de qualité des semences de vergers à graines existent aussi ailleurs, bien que l'auteur n'ait pas connaissance d'informations publiées sur ce point. L'objet du présent article est de faire connaître plus largement ce qui existe en Finlande en matière d'exigences minimales, et de solliciter des commentaires et suggestions fondés sur l'expérience acquise dans d'autres pays et avec d'autres essences. Bien qu'il soit peu probable que les présentes directives trouvent une application directe ailleurs, elles pourront sans doute aider d'autres pays en suggérant une méthodologie pour définir des normes quantitatives à respecter dans l'installation et la classification des vergers à graines.
Manuscrit reçu en février 1982.
EXIGENCES MINIMALES POUR LES VERGERS A GRAINES CLONAUX EN FINLANDE
Généralités
Le principal objectif des spécifications établies est d'assurer que le gain génétique attendu sera bien réalisé lorsqu'on utilisera des semences récoltées dans les vergers à graines enregistrés. Il y a trois points principaux à considérer: 1) l'identification des ramets dans le verger; 2) la composition clonale du verger; 3) les parents mâles des semences. Quoique importantes, des caractéristiques de forme, telles qu'un nombre égal de ramets par clone ou un dispositif de plantation défini, ne sont pas considérées comme essentielles pour notre propos.
1) Le strict respect de la règle selon laquelle on doit pouvoir identifier chaque greffe individuellement dans un verger à graines a parfois été considéré comme pédantesque et d'intérêt douteux dans un projet visant à produire des semences en quantités commerciales. Pourtant il faut bien voir que les éclaircies sélectives, les tests de descendance et les calculs de probabilités de croisement n'auraient aucun sens si le marquage des arbres et la documentation n'étaient pas dignes de confiance. Il est par conséquent indispensable de procéder à une inspection complète des vergers à graines pour déceler les erreurs possibles et les altérations au dispositif originel, et s'assurer que les porte-greffes n'ont pas éliminé les greffons et pris leur place.
2) La composition clonale se rapporte i) au nombre de clones; ii) au nombre de ramets par clone; iii) à l'origine des clones. Ces facteurs influent sur la diversité génétique des semences, sur la distribution des croisements par suite des synchronismes de floraison, et sur la répartition spatiale des greffes. Ils déterminent par ailleurs la zone dans laquelle les semences provenant du verger peuvent être utilisées.
3) L'exigence la plus complexe est celle qui se rapporte au parent qui a fourni le pollen. Toutes les spécifications concernant les vergers à graines s'accordent à dire que tant l'autopollinisation que la contamination par un pollen extérieur doivent être réduites au minimum. Cela signifie implicitement que l'on cherche à encourager les croisements entre les génotypes sélectionnés présents dans le verger. Ainsi, dans l'idéal, tous les clones devraient contribuer en proportions égales à la formation des gamètes femelles et mâles. En pratique, l'autopollinisation et la contamination par du pollen provenant de sources extérieures ne peuvent jamais être éliminées totalement et, en raison des différences individuelles dans l'abondance et l'époque de la floraison, la contribution des différents clones est loin d'être égale. Ce n'est qu'une fois que les caractéristiques phénologiques de chacun des clones présents dans le verger auront été déterminées que l'on pourra évaluer avec précision la répartition des croisements à l'intérieur du verger; les études visant à éclaircir cet aspect doivent donc démarrer le plus tôt possible. Les mesures réelles du degré d'autopollinisation et de consanguinité qui en découle doivent également être effectuées pour chaque clone, et leurs effets défavorables chiffrés au moyen de tests de descendance.
Le moyen le plus simple pour réduire la consanguinité est d'assurer une distance suffisante entre les ramets d'un même clone. L'importance de la contamination par du pollen extérieur peut être mesurée avec une relative précision sur le terrain, mais sa part réelle dans la pollinisation est difficile à déterminer. En règle générale, il est difficile de réduire le degré de pollinisation ambiante 1 dans les vergers à graines déjà existants. Dans le cas où elle est importante, la seule mesure à prendre pour réduire ses effets nuisibles est d'améliorer l'efficience des croisements à l'intérieur du verger, en favorisant une floraison abondante et en augmentant la taille du verger.
Spécifications minimales
En Finlande, l'enregistrement des vergers à graines non contrôlés se fonde sur sept considérations principales. Les valeurs numériques indiquées ont été calculées pour Pinus sylvestris. Ces exigences et ces valeurs numériques doivent être envisagées comme un tout: toutes les exigences doivent être satisfaites simultanément. Les valeurs numériques ont été estimées en tenant compte de la possibilité que l'un ou plusieurs des autres facteurs soient au voisinage du niveau minimum acceptable.
1. Le verger à graines doit être en bonne condition physique, et tous les documents exacts et à jour
Un plan détaillé indiquant le numéro de clone de chaque ramet ainsi que l'espacement est une condition indispensable pour toute recherche génétique. Outre le plan de position réelle, il doit y avoir également une liste indiquant le numéro de clone et la position (file et rangée) de chaque greffe et les distances entre files et rangées. En Finlande le tracé, l'inventaire, le calcul du taux de consanguinité, etc. se font à l'aide d'un programme d'ordinateur conçu à cet effet. Avant que les évaluations et calculs ne commencent, l'autorité nationale 1 doit vérifier l'exactitude du plan de position en visitant des placettes échantillons aléatoirement réparties sur l'ensemble du verger, afin de contrôler les greffes une par une.
2. L'aire géographique d'origine des ortets (= arbres plus originels) doit être limitée de façon à favoriser le synchronisme de la floraison et faciliter la détermination de la zone d'utilisation des semences 2
Lorsque les premiers vergers à graines furent établis en Finlande, des opinions divergentes furent exprimées quant à la taille acceptable pour la zone d'origine des ortets à y inclure. Le souci d'accroître la diversité génétique et de bénéficier des effets d'hétérosis incitait à inclure des arbres provenant d'une très vaste zone; l'hypothèse de l'existence de races locales de taille réduite, hautement différenciées, militait en faveur d'une zone étroite.
Grâce à des études menées en Finlande sur le rythme physiologique annuel de la croissance chez Pinus sylvestris, on a pu montrer que pour cette essence comme pour d'autres essences commercialement importantes il existe une forte variation à l'intérieur d'un même peuplement pour ce caractère. Si la variation dans le rythme annuel est maintenue dans le verger à graines telle qu'elle existe dans n'importe quel peuplement naturel continu, toutefois, les époques de floraison de tous les clones se chevaucheront, et par conséquent s'accorderont assez bien. L'explication supposée est que la sélection phénotypique pratiquée, favorisant les arbres de croissance et de qualité supérieures, n'a pas changé le mode de répartition du rythme annuel.
Cherchant à quantifier les limites recommandables de la zone d'origine des ortets, Sarvas (1967, 1969, 1972) est arrivé à la conclusion que dans la région tempérée fraîche la durée et la somme de températures cumulées de la saison de végétation sont d'une importance décisive pour le déclenchement de processus physiologiques déterminés tels que la floraison. Sur la base des moyennes et des écarts-types de la somme de températures 1 calculés pour des populations naturelles synchronisées, et du coefficient de variation dans ces populations, il est maintenant spécifié en Finlande que les ortets inclus dans un verger à graines doivent être originaires de zones ne différant pas plus de ± 130 d.j. En fonction de l'expérience acquise à ce jour, il est probable que ces limites seront encore rétrécies pour la génération suivante de vergers à graines.
3. Le nombre de clones doit être d'au moins 30
Cette exigence se fonde sur les faits suivants: i) chez P. sylvestris, les clones ne fleurissent pas en abondance chaque année, et comme une floraison abondante n'a pas été retenue comme critère de sélection en Finlande, certains clones, en conséquence, fleurissent plus que d'autres (voir par ex. Erickson et al. 1973; Jonsson et al. 1976; Bhumibhamon 1978; Koski et Tallqvist 1978). Si le nombre total de clones est réduit, on aura une probabilité accrue que la contribution de certains clones à la production de pollen et de semences d'une année sur l'autre soit exagérément importante. Bien qu'il n'ait été présenté aucun modèle théorique pour le nombre optimal de clones, plusieurs auteurs recommandent un chiffre minimum variant de 20 à 30 clones; ii) un petit nombre de clones augmente la probabilité de consanguinité, parce que la distance entre les ramets d'un clone quelconque se trouve réduite quel que soit le dispositif de plantation. Ce fait sera aggravé s'il y a une grande variation dans le nombre de ramets par clone; un grand nombre de ramets dans un clone, combiné avec un faible nombre dans un autre, entraîne inévitablement la constitution de bouquets de certaines clones, et par conséquent un risque accru de consanguinité. En Finlande, la variation admise dans le nombre de ramets est ± 30 pour cent du nombre moyen dans le verger.
4. Les ramets de chaque clone doivent être répartis dans l'espace de telle sorte que la probabilité d'autopollinisation soit toujours inférieure à 20 pour cent
En raison de la présence chez P. sylvestris de gènes létaux embryonnaires, la proportion d'embryons vivants issus d'autopollinisation est bien inférieure à la proportion réelle d'autopollinisation. L'efficacité de ce triage biologique de la consanguinité varie selon les espèces et les individus (voir par ex. Koski 1973). Lindgren (1975) et l'auteur (Koski 1982) ont calculé la probabilité d'embryons consanguins à la suite de différentes proportions d'autopollinisation. Dans un génotype modal 2 de Pinus sylvestris ou de Picea abies, ces auteurs indiquent que 20 pour cent d'auto-pollinisation produiront 3 à 4 pour cent d'individus autogames dans la descendance. Il n'existe aucun critère absolu pour fixer la limite à 20 pour cent; toutefois une limite plus basse serait difficile à appliquer en pratique.
On peut estimer la proportion d'autopollinisation en se basant sur i) les valeurs moyennes de l'autopollinisation naturelle et ii) la dispersion de pollen. Il n'est pas douteux que dans certains cas les valeurs réelles diffèrent considérablement des prévisions. Un système plus raffiné exigerait des informations détaillées sur l'abondance de la floraison mâle, le nombre de gènes létaux embryonnaires dans chaque clone, les conditions de vent, etc. Au stade actuel des connaissances on admet que la proportion d'autopollinisation dans la cime de chaque arbre est de 10 pour cent dans un peuplement normal. Cette hypothèse se relie à l'abondance de la floraison mâle (point 5) et à la superficie minimale du verger (point 6). La présence de ramets contigus d'un même clone accroît naturellement la probabilité d'autopollinisation.
Dans le programme d'ordinateur actuellement en usage en Finlande, l'ordinateur vérifie le voisinage de chaque greffe dans une circonférence de 30 mètres 1. S'il détecte un arbre ayant le même numéro de clone, il calcule la distance exacte entre les deux ramets. Le programme comporte une fonction pour la densité de pollen en relation avec la distance: P = 10 e-d/8,5, dans laquelle P est la probabilité d'autopollinisation (en %), e la base des logarithmes naturels, d la distance (en mètres). Le coefficient 8,5 a été modifié par rapport aux premières valeurs empiriques (Koski 1970, Müller 1977, Shen et al. 1981). Si la probabilité cumulée d'autopollinisation excède 20 pour cent, il faut prendre des mesures en conséquence sur le terrain pour abaisser cette estimation, en éliminant un ou plusieurs ramets. La limite de 20 pour cent est assez généreuse; en fait, elle est environ deux fois plus élevée que l'autopollinisation naturelle estimée chez P. sylvestris, telle que la démontrent des études biochimiques récentes (voir par ex. Müller 1976; Müller-Stark 1979; Shen et al. 1981; Rudin et Ekberg 1982).
5. La quantité de pollen produite à l'hectare et par an dans le verger doit être d'au moins 20 kg
La pollinisation par le vent ne joue convenablement que lorsque la quantité de pollen est suffisamment importante. D'une manière générale, une pollinisation insuffisante a pour résultat une excision des jeunes cônes et un avortement des ovules, en d'autres termes une diminution de la récolte de semences (Sarvas 1962). Dans un verger à graines clonal, une faible production de pollen accroît la probabilité d'autopollinisation dans les clones à floraison relativement abondante, et surtout elle accroît la probabilité de contamination par du pollen extérieur.
En règle générale, les greffes de Pinus sylvestris ne commencent à produire des fleurs mâles que lorsqu'elles ont atteint une certaine taille. Une fois que la floraison mâle a démarré, son abondance est pendant plusieurs années proportionnelle à l'âge et à la taille des greffes. On peut prédire en gros que lorsque les greffes auront atteint une hauteur de 6–8 mètres, ou un diamètre à hauteur d'homme de 14–16 cm, la production de pollen sera satisfaisante, à la condition que le nombre d'arbres à l'hectare soit d'au moins 250. D'un autre côté, un espacement trop faible dans le verger provoque la mort des basses branches, sur lesquelles sont produits la majorité des strobiles.
Bien que l'on puisse prédire approximativement la production de pollen en se basant sur la taille des greffes, il faut procéder également à une estimation plus précise une fois que les greffes ont effectivement commencé à fleurir. Un comptage direct de tous les strobiles mâles serait trop laborieux pour être appliqué en pratique. Chez P. sylvestris, les inflorescences mâles peuvent être comptées jusqu'à deux ou trois ans après la floraison, en comptant les cicatrices, ou espaces dépourvus d'aiguilles, sur les rameaux, verticille par verticille. En règle générale, il suffit de compter le nombre de cicatrices sur une branche par verticille, et de multiplier par le nombre de branches dans le verticille. Il faut toutefois s'assurer au préalable de la précision de cette méthode en comparant le nombre de cicatrices sur différentes branches.
Comme chaque centimètre de cicatrice correspond à une certaine quantité de pollen, qui peut être déterminée expérimentalement, le poids moyen de pollen produit par greffe peut être aisément calculé. En Finlande, l'estimation de la quantité de pollen produite par clone se fonde sur le comptage de 5 ramets par clone. Le total de tous les clones représente la production de pollen de l'ensemble du verger. Dans le sud de la Finlande, la production maximum de pollen enregistrée dans les vergers à graines de Pinus sylvestris est de l'ordre de 90 kg/ha/an 2.
6. La superficie du verger à graines doit être d'au moins 5 hectares, et la plus petite largeur d'au moins 150 mètres
Il faut une surface relativement étendue de greffes pour engendrer un nuage de pollen suffisamment dense. Dans un petit “îlot” d'arbres, le pollen est immédiatement transporté en dehors du peuplement, et la densité de pollen demeure faible. Dans ce cas, la proportion d'autopollinisation et éventuellement de pollinisation ambiante sera élevée, même si la floraison mâle dans le verger est abondante. Des valeurs numériques ont été obtenues par une équation de régression de l'efficience de la pollinisation en fonction du diamètre du peuplement (Koski 1975).
7. La proportion de contamination par du pollen extérieur (pollinisation ambiante) doit être inférieure à 20 pour cent
L'existence de migrations importantes de pollen n'est souvent pas suffisamment prise en considération. On recommande souvent une bande d'isolement de 100–200 mètres comme protection totale contre la contamination par du pollen extérieur. La production de semences viables peu après l'installation d'un verger à graines, alors que la floraison mâle n'a pas encore commencé ou est peu abondante, est une preuve convaincante d'un isolement insuffisant. Les quantités de pollen étranger et les périodes de pointe pour la contamination peuvent être aisément mesurées durant le jeune âge d'un verger.
En Finlande, où la distance minimum aux arbres pouvant s'hybrider avec ceux du verger est fixée à 100 mètres, on a constaté que la quantité de pollen extérieur peut être en relation directe avec l'abondance de l'essence dans la région, plutôt qu'au voisinage immédiat. Si la pollinisation ambiante est importante, les seules mesures efficaces pour la réduire sont i) d'accroître la pollinisation à l'intérieur du verger à graines ou ii) de créer des barrières phénologiques entre le verger à graines et les forêts environnantes. Les directives données aux points 2, 5 et 6 ci-dessus tiennent compte également du problème de la pollinisation ambiante.
On peut parfois créer une barrière phénologique en transférant le verger à graines dans une région où sa floraison sera déphasée par rapport aux populations locales. Toutefois, l'effet d'un tel transfert n'est pas toujours en corrélation linéaire avec la distance (Chung 1981); cela signifie que la probabilité de croisements indésirables ne peut pas toujours être exactement prédite, et que de tels transferts comportent par suite un élément d'incertitude.
L'exigence minimum proposée de 20 pour cent de contamination ou moins peut sembler à beaucoup de généticiens forestiers être excessivement généreuse. Dans de nombreuses régions, on peut certainement appliquer des limites plus strictes. En Finlande, cependant, il serait illusoire de placer la limite plus bas, si l'on considère les vastes surfaces existantes de peuplements naturels de P. sylvestris. D'un autre côté, la diminution de gain génétique due à une pollinisation ambiante de 20 pour cent est, si on la calcule théoriquement, seulement de un dixième par comparaison avec le cas d'un isolement total. Si le gain attendu était, mettons, de 15 pour cent, une pollinisation ambiante de 20 pour cent le ramènerait donc à 13,5 pour cent. Considérant le fait que nous parlons de vergers de première génération, dans lesquels les fréquences de gènes n'ont pas encore été modifiées de façon importante par rapport aux populations originelles, une diminution de cet ordre de grandeur est considérée comme acceptable. Toutefois, lorsqu'on projettera de nouvelles générations de vergers à graines, il faudra assurer un isolement plus effectif.
REFERENCES
Barner, H. 1976 Certification of Forest Reproductive Material. In Report on the FAO/DANIDA Training Course on Forest Seed Collection and Handling. Vol. II, pp. 65–78.
Bhumibhamon, S. 1978 Studies on Scots Pine Seed Orchards in Finland with Special Emphasis on the Genetic Composition of the Seed. Commun. Inst. For. Fenn. 94:4: 1–118.
Chung, M.S. 1981 Flowering Characteristics of Pinus sylvestris L: with Special Emphasis on the Reproductive Adaptation to Local Temperature Factor. Acta For. Fenn. 169: 1–69.
Eriksson, G., Jonsson, A and Lindgren, D. 1973 Flowering in a Clone Trial of Picea abies Karst. Stud. For. Suec. 110: 1–45.
Hagman, M. 1982 Die neue Finnische Gesetzgebung über den Handel von forstlichem Verjüngungsmaterial und die dazu gehörige Regelung der Versorgung von Waldbaumsamen. Allgem. Forstz. (Jahrgang 1982, in press).
Jonsson, A. Ekberg, I. and Eriksson, G. 1976 Flowering in a Seed Orchard of Pinus sylvestris L. Stud. For. Suec. 135: 1–38.
Koski, V. 1970 A Study on Pollen Dispersal as a Mechanism of Gene Flow in Conifers. Commun. Inst. For. Fenn. 70.4: 1–78.
Koski, V. 1973 On Self-Pollination, Genetic Load and Subsequent Inbreeding in Some Conifers. Ibid. 78.10: 1–42.
Koski, V. 1975 Natural Pollination in Seed Orchards with Special Reference to Pines. Forestry Commission, Bull. 54: 83–91.
Koski, V. 1980 Minimivaatimukset männyn siemenviljelyksille Suomessa. Summary: Minimum Requirements for Seed Orchards of Scots Pine in Finland. Silva Fenn. 14: 136–149.
Koski, V. 1982 How to Study the Rate of Inbreeding in Populations of Pinus sylvestris and Picea abies. Ibid. 16 (in press).
Koski, V. and Tallqvist, R. 1978 Tuloksia monivuotisista kukinnan ja siemensadon määrän mittauksista metsäpuilla. Summary: Results of Long-Time Measurements of the Quantity of the Flowering and Seed Crop of Forest Trees. Folia For. 364: 1–60.
Lindgren, D. 1975 The Relationship between Self-Fertilization, Empty Seeds and Seeds Originating from Selfing as a Consequence of Polyembryony. Stud. For. Suec. 126: 1–24.
Müller, G. 1976 Untersuchungen über die natürliche Selbstbefruchtung in Beständen der Fichte (Picea abies (L.) Karst.) und Kiefer (Pinus sylvestris L.) Silvae Gen. 26: 207–217.
Müller-Stark, G. 1979 Estimates of Self- and Cross-Fertilization in a Scots Pine Seed Orchard. Proc. Confer. Bioch. Gen. For. Trees. Umea. Sweden 1978: 170–179.
OCDE 1974 Système de l'OCDE pour le contrôle des matériels forestiers de reproduction destinés au commerce international. Organisation de coopération et de développement économiques, 2 rue André Pascal, 75016 Paris, France.
Rudin, D. and Ekberg, I. 1982 Genetic Structure of Open Pollinated Progenies from a Seed Orchard of Pinus sylvestris. Silva Fenn. 16. (In press).
Sarvas, R. 1962 Investigations on the Flowering and Seed Crop of Pinus sylvestris. Commun. Inst. For. Fenn. 53.4: 1–198.
Sarvas, R. 1967 The Annual Period of Development of Forest Trees. Proc. Finn. Acad. Sci. Lett. 1965: 211–231.
Sarvas, R. 1969 Adaptation génétique des arbres forestiers au facteur thermique du climat. Documents de la Deuxième Consultation mondiale sur l'amélioration des arbres forestiers, Washington, FAO/IUFRO, FO, FTB 69/2:1–11.
Sarvas, R. 1970 Establishment and Registration of Seed Orchards. Folia For. 89: 1–24.
Sarvas, R. 1972 Investigations on the Annual Cycle of Development of Forest Trees, Active Period. Commun. Inst. For. Fenn. 76.3: 1–110.
Shen, H.H., Rudin, D. and Lindgren, D. 1981 Study of the Pollination Pattern in a Scots Pine Seed Orchard by Means of Isozyme Analysis. Silva Genetica 30:7–15.
1 Organisation de coopération et de développement économiques.
1 L'écartement de plantation est généralement de 5 × 5 m dans les vergers à graines de Finlande.
![]()
![]()
![]()
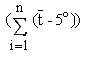 (
(