![]()
![]()
![]()
por
Veikko Koski
The Finish Forest Research Institute
Unioninkatu 40 A
SF-00170 Helsinki 17
Finlandia
INTRODUCCION
Los objectivos de los huertos de semillas son producir semilla genéticamente mejorada de árboles forestales, fomentar las cosechas frecuentes de la misma y facilitar la recolección. A escala internacional, la publicación de normas específicas sobre niveles mínimos para huertos de semillas de árboles es relativamente reciente, como directrices para los países que participaron en el “OECD 1 Scheme for the Controlf of Forest Reproductive Material Moving in International Trade” (Proyecto para el control del material de reproducción forestal que se mueve en el mercado internacional, OCDE 1974). Aunque en dichas normas se especifica que hay que evitar en lo posible la autopolinización y la contaminación por polen externo, no se da ninguna norma cuantitativa ni se establecen exigencias específicas. La responsabilidad del establecimiento de criterios específicos para los huertos de semillas pertenecientes a las distintas categorías reconocidas en el proyecto queda en manos de las autoridades nacionales que tienen a su cargo su ejecución.
Es natural que no se puedan establecer requerimientos detallados de carácter general, comunes para los huertos de semillas; hay que tener en cuenta las diversas categorías y tipos de huertos de semillas, las características biológicas de las distintas especies de árboles y las condiciones del medio. Por otra parte, nuestros conocimientos en materia de genética cuantitativa y de biología de la floración de numerosas especies de árboles forestales han aumentado considerablemente desde que el concepto de huertos de semillas comenzó a formar parte integrante de las actividades de la fitogenética forestal. En realidad, en varias investigaciones realizadas en los países nórdicos se indica la posibilidad de aplicar criterios cuantitativos al estudio del funcionamiento de un huerto de semillas como unidad genética (Sarvas 1970, Eriksson et al. 1973, Jonsson et al. 1976, Koski 1975, Bhumibhamon 1978, Chung 1981).
En numerosos países está ya en vigor una legislación nacional que exige el registro oficial de los huertos de semillas y otras fuentes de material de reproducción forestal, y es probable que en el futuro aumento el número de estos países. Si se desea respetar el concepto de semilla genéticamente mejorada, son imprescindibles ciertas garantías en cuanto a la composición y propiedades de los lotes de semillas, de acuerdo con la legislación mencionada. En la mayoría de los casos, la certificación por medio de una evaluación real de pruebas de descendencia con suficiente antigüedad supondría un retraso que podría llegar a ser de uno o varios decenios. Con objeto de evitar estas demoras, los científicos que trabajan con especies de árboles forestales se ven obligados muchas veces a fiarse de la teoría genética más que sus colegas de los sectores de la agricultura y la horticultura.
Son varios los factores que influyen en la composición genética de la semilla cosechada en un huerto de semillas. A muchos de ellos se les puede aplicar un criterio cuantitativo, por ejemplo al número de genotipos del huerto, la intensidad de la selección, la probabilidad de intracruzamiento y la cuantía de la contaminación por polen externo. Basados en estudios anteriores, Sarvas (1970) y Bhumibhamon (1978) han propuesto el uso de un índice para el registro de huertos de semillas. Cada factor que se tiene en cuenta en el índice se evaluaría por separado siguiendo una escala de puntos; el número total de puntos indicaría el valor genético de la semilla cosechada en el huerto de semillas, haciendo posible así una clasificación coherente de los distintos huertos.
Aunque el sistema del índice es más objetivo que la apreciación intuitiva, en la prática tiene sus limitaciones: una puntuación elevada en un factor podría compensar el valor inaceptablemente bajo de otro. Es, pues, discutible si los distintos grados alcanzados por la utilización de dicho índice reflejarían o no el valor genético del material de reproducción y su valor económico, incluso en términos relativos. El autor de este artículo considera más fiable y práctico: (i) clasificar los huertos de semillas (por ej. huertos de clonos de Pinus sylvestris; huertos de árboles para semilla de Pinus sylvestris; huertos de clonos de Picea abies, etc.); y (ii) a partir de esta clasificación, definir los requerimientos mínios de determinados factores evaluables que influyen en la calidad genética de la semilla.
Entre los diversos tipos de huertos de semillas señalados en el proyecto OCDE (ver, por ejemplo, Barner 1976), los clonos de primera generación, que pertenecerían a la categoría de la OCDE de “huertos regionales” 1, son actualmente de los más corrientes a escala mundial. Este es también el caso Finlandia, que se incorporó oficialmente al proyecto OCDE en 1980 (Hagman, 1982).
En Finlandia se han establecido y publicado en fecha reciente (Koski 1980) los requerimientos mínimos para huertos de semillas de clonos de Pinus sylvestris. Estas directrices ya están en vigor. Lo más probable es que en otras partes también existan directrices técnicas semejantes para el control de calidad de los huertos de semillas, aunque el autor no tiene conocimiento de información publicada al respecto. El objeto de este artículo es difundir los requerimientos mínimos finlandeses, y solicitar comentarios y sugerencias que tengan como base las experiencias de otros países y otras especies. Aunque probablemente no se podrá dar a estas directrices una aplicación directa más amplia, pueden servir de ayuda a otros países como base metodológica en el establecimiento de normas cuantitativas específicas para la creación y clasificación de huertos de semillas.
1 Organización de Cooperación y Desarrollo Económicos. Recibido en febrero de 1982.
REQUERIMIENTOS MINIMOS FINLANDESES PARA LOS HUERTOS DE SEMILLAS DE CLONOS DE PINO
Consideraciones generales
El principal objetivo de los requerimientos establecidos es asegurar el logro de la ganancia genética esperada cuando se utiliza semilla cosechada en huertos de semillas registrados. Son tres los principales aspectos que hay que tener en cuenta: (1) la determinación de las plantas de clono en el huerto; (2) la composición clonal del huerto; y (3) la procedencia del polen de la semilla. Hay aspectos formales, como el número idéntico de plantas por clono y un diseño específico de la plantación que, aunque importante, no se consideran decisivos en este contexto.
1. La aplicación estricta de la norma de que cada uno de los vástagos de un huerto de semillas deba ser identificable, se ha considerado en ocasiones formalista y de dudoso valor en un proyecto orientado a la producción de cantidades comerciales de semilla. Sin embargo, hay que tener en cuenta que las claras selectivas, los ensayos de descendencia y el cálculo de las probabilidades de cruzamientos carecerán de sentido sin una clasificación y documentación exactas. Es, por consiguiente, fundamental la realización de encuestas exhaustivas en los huertos de semillas, para descubrir posibles errores y alteraciones en los datos originales y asegurar que los patrones no han sustituido y eliminado los vástagos injertados.
2. La composición clonal se refiere a: (i) el número de clonos, (ii) el número de plantas por clono, y (iii) el origen de los clonos. Estos factores influyen en la diversidad genética de la semilla, en la pauta de cruzamiento, a través de la sincronización de la floración, y en la distribución espacial de los vástagos. También depende de ellos la zona en que se podrá utilizar la semilla del huerto.
3. El requerimiento más complejo es el que se refiere a la procedencia del polen de la semilla. Todas las definiciones de huertos de semillas coinciden en la necesidad de evitar en lo posible tanto la autopolinización como la contaminación por polen externo. Esto significa de manera implícita que se fomenta el cruzamiento entre los genotipos seleccionados incluidos en el huerto. De esta manera, en teoría todos los clonos deberían aportar una proporción igual de gametos masculinos y femeninos. En la práctica nunca se pueden eliminar completamente la autopolinización y la contaminación por polen de otras procedencias y las diferencias entre los distintos clonos en cuanto a la abundancia y el tiempo de la floración hacen que la contribución de todos ellos diste mucho de ser igual. La pauta de cruzamiento dentro del huerto solamente se podrá evaluar de manera exacta una vez determinadas las características fenológicas de cada uno de los clonos del huerto; es necesario iniciar lo antes posible estudios que aclaren este aspecto. También se deberían realizar medidas reales del grado de autopolinización y el consiguiente intracruzamiento, tomando como base un solo clono, y habría que determinar el valor cuantitativo de sus efectos perjudiciales por medio de ensayos de descendencia.
La manera más sencilla de reducir el intracruzamiento es asegurar una distancia adecuada entre las plantas del mismo clono. El alcance de la contaminación por polen externo se puede medir de manera relativamente exacta sobre el terreno, pero resulta difícil determinar su importancia en la polinización real. La reducción de la cuantía de la polinización de fondo 1 en los huertos de semillas ya existentes suele ser difícil. En el caso de que sea abundante, la única medida que se puede tomar para reducir sus efectos perjudiciales es mejorar la eficacia del cruzamiento dentro de huerto de semillas, estimulando una floración abundante y ampliando el huerto.
Requerimientos mínimos
El registro de huertos de semillas de clonos no probados en Finlandia se basa en siete puntos principales. Los valores numéricos que se dan se han calculado para Pinus sylvestris. Hay que considerar en conjunto los requerimientos y los valores numéricos: Se deben cumplir simultáneamente todos los requerimientos. Los valores numéricos se han calculado contando con la posibilidad de que entre los otros factores haya uno o varios que estén próximos al nivel mínimo aceptable.
1. El huerto de semillas ha de estar en buenas condiciones físicas y todos los documentos deben ser exactos y estar actualizados
Para cualquier investigación genética es imprescindible un mapa detallado, en el que se indique el número de clono de cada planta de multiplicación, así como la separación entre ellas. Además del mapa de posición, debe existir también una lista con el número de clono y la posición (= fila/columna) de cada vástago, y las distancias entre las filas y las columnas. La disposición, enumeración y cálculo de intracruzamiento, etc., se hacen en Finlancia con la ayuda de un programa de computador preparado al efecto. Antes de realizar cualquier evaluación o cálculo, la autoridad nacional 2 ha de comprobar la exactitud del mapa de posición visitando “parcelas de prueba” distribuidas al azar en todo el huerto, para examinar los vástagos uno por uno.
2. El ámbito geográfico de los pies clonales (= árboles sobresalientes originales) debe ser limitado, de manera que se facilite la sincronización de la floración y la determinación de la zona de utilización de la semilla 1
Al establecerse por primera vez huertos de semillas en Finlandia, las opiniones eran dispares por lo que se refiere al tamaño de un ámbito aceptable. Quienes deseaban incrementar la diversidad genética y conseguir efectos de heterosis, abogaban por la inclusión de árboles de una zona muy amplia; quienes mantenían la hipótesis de la existencia de pequeñas razas locales muy diferenciadas, apoyaban la idea de un ámbito reducido.
A través de estudios realizados sobre el ritmo del crecimiento fisiológico anual de Pinus sylvestris en Finlandia, se ha demostrado que en ésta y en otras especies importantes desde el punto de vista comercial varía mucho esta característica dentro de una misma masa. Si las variaciones del ritmo anual son en un huerto de semillas del mismo orden que en una masa natural continua, se superpondrán los períodos de floración de todos los clonos y, por consiguiente, se emparejarán bastante bien. Según esto, la selección fenotípica que favorece a los árboles de mayor crecimiento y calidad no ha cambiado los patrones de distribución del ritmo anual.
En la determinación cuantitativa de los límites de un ámbito recomendado, Sarvas (1967, 1969, 1972) llegó a la conclusión de que la duración y la suma del calor acumulado de la temporada de crecimiento tienen en la región templado-fría una importancia decisiva para la iniciación de procesos fisiológicos específicos, como la floración. Tomando como base las medias y las desviaciones típicas de la suma 2 calculadas para poblaciones naturales sincronizadas, y el coeficiente de variación en dichas poblaciones, actualmente se especifica en Finlandia que los pies clonales incluidos en un huerto de semillas deben proceder de zones con diferencias no superiores a ± 130 g.d. De acuerdo con la experiencia obtenida hasta la fecha, es probable que estos límites se reduzcan todavía más en la próxima generación de huertos de semillas.
3. El número de clonos ha de ser por lo menos de 30
Este requerimiento se basa en los siguientes hechos: (i) en P. sylvestris, la floración de los clonos no es abundante todos los años y, habida cuenta que la floración abundante no se ha utilizado como criterio de selección en Finlandia, algunos clonos siempre florecen más que otros (ver p. ej. Erickson et al. 1973; Jonsson et al. 1976; Bhmibhamon 1978; Koski y Tallqvist 1978). Si el número total de clonos es pequeño, aumentará la probabilidad de que la contribución de un año para otro de determinados clonos a la producción de polen y semilla sea desproporcionadamente grande. Aunque no se ha presentado ningún modelo teórico para establecer el número óptimo de clonos, varios autores recomiendan un número mínimo comprendido entre 20 y 30; (ii) si el número de clonos es pequeńo, aumenta la probabilidad de intracruzamiento, debido a que las distancias entre las plantas de multiplicación de cada clono serán pequeñas en cualquier diseño de plantación. Esto se verá agravado si existe mucha variación en el número de plantas por clono; un elevado número de las mismas en algunos clonos, unido a un número pequeño en otros, lleva inevitablemente al agrupamiento de algunos clonos, y por consiguiente aumenta el riesgo de intracruzamiento. La variación aceptada en Finlandia en cuanto al número de plantas de clono es de ± 30 por ciento del número medio del huerto.
4. La distribución espacial de las plantas de multiplicación de cada clono debe ser tal que la probabilidad de autopolinización sea siempre inferior al 20 por ciento
Debido a la presencia de factores letales para los embriones en P. sylvestris, la proporción de embriones vivos que aparecen por autopolinización es muy inferior a la de la autopolinización real. La eficacia de esta selección biológica del intracruzamiento varía entre las distintas especies e individuos (ver p.ej. Koski 1973). Lindgren (1975) y el autor de este artículo (Koski 1982) han calculado las probabilidades de formación de embriones por intracruzamiento después de distintas proporciones de autopolinización. En un genotipo de valor predominante 1 de Pinus sylvestris o Picea abies. el 20 por ciento de autopolinización mencionado produciría del 3 al 4 por ciento de individuos autofecundados en la descendencia. No existe un criterio absoluto para establecer el límite en el 20 por ciento; sin embargo, un límite inferior más estricto sería difíci de aplicar en la práctica.
El cálculo de la proporción de autopolinización puede basarse en: (i) los valores medios de la autopolinización natural, y (ii) la dispersión del polen. No cabe duda que en algunos casos los valores reales sufren una desviación considerable con respecto a las expectativas. Para un método más complejo sería necesaria una información detallada acerca de la abundancia de la floración masculina, el número de factores letales para el embrión en cada clono, las condiciones del viento, etc. Con los conocimientos actuales, se supone que la proporción de autopolinización dentro de la copa de cada árbol es del 10 por ciento en una masa normal. Esta suposición tiene relación con la abundancia de floración masculina (punto 5) y la superficie mínima del huerto (punto 6). Las plantas adyacentes del mismo clono aumenta la probabilidad de autopolinización.
En el programa de computador que se utiliza actualmente en Finlandia, el computador examina la zona que rodea cada vástago en una circunferencia de 30 metros 2. Si detecta un árbol con el mismo número clonal, calcula la distancia exacta entre las dos plantas. El programa contiene una función para la densidad del polen en relación con la distancia P= 10 e - d/8,5, en la que P es la probabilidad de autopolinización (%); e es la base del logaritmo neperiano y d es la distancia (metros). El coeficiente 8,5 se ha modificado a partir de valores empíricos anteriores (Koski 1970, Müller 1977), Shen et al. 1981). Si la probabilidad acumulativa de autopolinización supera el 20 por ciento, hay que tomar las medidas oportunas sobre el terreno para disminuir esta estimación, eliminando una o varias plantas. El límite del 20 por ciento es bastante amplio; en realidad es aproximadamente el doble de la autopolinización natural estimada en P. sylvestris, tal como han puesto de manifiesto estudios bioquímicos recientes (ver, por ejemplo, Müller 1976; Müller-Stark 1979; Shen et al. 1981; Rudin y Ekberg 1982).
1 Valor predominante = Norma: valor, magnitud o resultado que se presenta el mayor número
de veces en una serie determinada de observaciones.
2 La distancia de plantación suele ser de 5 m × 5 m en los huertos de semillas de Finlandia.
5. El huerto debe producir un mínimo de 20 kg de polen por año y hectárea
La anemogamia solamente es satisfactoria cuando existe una gran cantidad de polen. La polinización insuficiente suele provocar el desprendimiento de pequeños conos y el aborto de óvulos, es decir, una reducción en la cosecha de semilla (Sarvas, 1962). En un huerto de semillas de clonos, la escasez de polen hace que aumente la probabilidad de autopolinización en los clonos con una floración relativamente abundante, y sobre todo hace más probable la contaminación por polen externo.
Por lo general, los vástagos de Pinus sylvestris no comienzan a producir flores masculinas mientras no han alcanzado un tamaño determinado. Una vez iniciada la floración masculina, durante varios años su cantidad es proporcional a la edad y tamaño de las plantas. Se puede predecir que la producción de polen será satisfactoria aproximadamente cuando los injertos hayan alcanzado una altura de 6–8 metros o un diámetro a la altura del pecho de 14–16 cm, con tal que el número de árboles por hectárea sea de 250 como mínimo. Por otra parte, la proximidad excesiva de las plantas en el huerto provoca la muerte prematura de las ramas más bajas, productoras de la mayoría de los estróbilos masculinos.
Aunque se puede hacer una predicción aproximada de la producción de polen en función del tamaño del vástago, es necesario hacer también una valoración más exacta una vez que se ha iniciado la floración. El recuento directo de todos los estróbilos masculinos resultaría demasiado laborioso para llevarlo a la práctica. Las inflorescencias masculinas de P. sylvestris se pueden contar incluso dos o tres años después de la floración, contando las “cicatrices” o espacios sin agujas en las ramas que forman cada uno de los verticilos. En general basta contar el número de escamas de una rama por verticilo y multiplicar por el número de ramas del mismo. Sin embargo, se ha de comprobar previamente la exactitud de este método comparando el número de cicatrices en diferentes ramas.
Como a las cicatrices de cada centímetro corresponderá una cantidad específica (= peso) de polen, que se puede determinar experimentalmente, resulta fácil el cálculo de la cosecha media de polen por planta. En Finlandia, la cosecha de polen de un clono se calcula haciendo el recuento de cinco plantas del mismo. El total de todos los clonos corresponde a la producción de polen de todo el huerto. En el sur de Finlandia, las mayores cosechas de polen registradas en huertos de semillas de P. sylvestris han sido del orden de 90 kg/ha/año 1.
6. El huerto de semillas debe tener una superficie mínima de 5 ha, y el diámetro más corto debe ser de 150 m por lo menos
Se necesita una superficie relativamente grande de plantas para producir una nube de polen con una densidad satisfactoria. En una pequeña “isla” de árboles, el polen es arrastrado inmediatamente fuera de la masa, y la densidad polínica se mantiene baja. En este caso la proporción de autopolinización y posible polinización de fondo será alta, aunque la floración masculina del huerto sea abundante. Los valores numéricos indicados se obtuvieron a partir de una regresión de la eficacia de polinización en el diámetro de la masa (Koski 1975).
7. La proporción de contaminación por polen externo (polinización de fondo) ha de ser inferior al 20 por ciento
A menudo no se tiene suficientemente en cuenta la existencia de una considerable migración de polen. Se recomiendan con frecuencia franjas aislantes de 100–200 m como protección total contra la contaminación por polen externo. La producción de semilla germinable al poco tiempo de establecer un huerto de semillas, cuando todavía no se ha iniciado la floración masculina o es escasa, es una prueba evidente de un aislamiento inadecuado. Durante la fase juvenil de un huerto se pueden medir fácilmente la cantidad de polen externo y los períodos de contaminación máxima.
En Finlandia, donde la distancia mínima establecida con respecto a árboles hibridantes es de 100 m, se ha comprobado que existe una relación directa entre la cantidad de polen externo y la abundancia de especies arbóreas en la región, más que en las proximidades inmediatas. Si la polinización de fondo es abundante, las únicas medidas eficaces para reducirla consisten en (i) potenciar la polinización dentro del huerto de semillas, o (ii) crear barreras fenológicas entre el huerto de semillas y los bosques circundantes. En las orientaciones que se han dado en los puntos 2, 5 y 6 se ha tenido también en cuenta el problema de la polinización de fondo.
En ocasiones se pueden crear barreras fenológicas físicas trasladando un huerto de semillas a una región donde su floración no coincida con la de las poblaciones locales. Sin embargo, no siempre existe una correlación lineal entre los efectos de un traslado y la distancia del mismo (Chung 1981): esto significa que no se puede predecir con exactitud la probabilidad de los cruzamientos indeseables, lo cual introduce un elemento de incertidumbre en dichos traslados.
El requerimiento mínimo sugerido, del 20 por ciento de contaminación o menos, puede parecer excesivo a muchos seleccionadores de árboles. Es indudable que en numerosas regiones se pueden aplicar límites más estrictos. No obstante, pretender establecer un límite más bajo en Finlandia sería engañarse, teniendo en cuenta las grandes superficies existentes de masas naturales de P. sylvestris. Por otra parte, la disminución en la ganancia genética a causa de una polinización de fondo del 20 por ciento es, en cálculos teóricos, solamente de una décima parte cuando se compara con una situación de aislamiento total. Si suponemos que la ganancia esperada es del 15 por ciento, una polinización de fondo del 20 por ciento la haría disminuir al 13,5 por ciento. Habida cuenta que hablamos de huertos de primera generación, en los que no se han producido grandes cambios en las frecuencias genéticas en comparación con las poblaciones originales, una disminución de esta magnitud se ha considerado aceptable. Sin embargo, cuando se proyecten huertos de semillas de generaciones avanzadas habrá que asegurar un aislamiento más eficaz.
BIBLIOGRAFIA
Barner, H. 1976 Certification of Forest Reproductive Material. En Report on the FAO/DANIDA Training Course on Forest Seed Collection and Handling. Vol. II, págs. 65–78.
Bhumibhamon, S. 1978 Studies on Scots Pine Seed Orchards in Finland with Special Emphasis on the Genetic Composition of the Seed. Cmmun. Inst. For. Fenn. 94.4: 1–118.
Chung, M.S. 1981 Flowering Characteristics of Pinus sylvestris L. with Special Emphasis on the Reproductive Adaptation to Local Temperature Factor. Acta For. Fenn. 169: 1–69.
Eriksson, G., Jonsson, A. y Lindgren, D. 1973 Flowering in a Clone Trial of Picea abies Karst. Stud. For. Suec. 110: 1–45.
Hagman, M. 1982 Die neue Finnische Gesetzgebung über den Handel von forstlichem Verjüngungs-material und die dazu gehörige Regelung der Versorgung von Waldbaumsamen. Allgem. Forstz. (Jahrgang 1982, en prensa).
Jonsson, A. Ekberg, I. y Eriksson, G. 1976 Flowering in a Seed Orchard of Pinus sylvestris L. Stud, For. Suec. 135: 1–38.
Koski, V. 1970 A Study on Pollen Dispersal as a Mechanism of Gene Flow in Conifers. Commun. Inst. For. Fenn. 70.4: 1–78.
Koski, V. 1973 On Self-Pollination, Genetic Load and Subsequent Inbreeding in Some Conifers. Ibid. 78.10: 1–42.
Koski, V. 1975 Natural Pollination in Seed Orchards with Special Reference to Pines. Forestry Commission, Bull. 54: 83–91.
Koski, V. 1980 Minimivaatimukset männyn siemenviljelyksille Suomessa. Summary: Minimum Requirements for Seed Orchards of Scots Pine in Finland. Silva Fenn. 14: 136–149.
Koski, V. 1982 How to Study the Rate of Inbreeding in Populations of Pinus sylvestris and Picea abies. Ibid. 16 (en prensa).
Koski, V. y Tallqvist, R. 1978 Tuloksia monivuotisista kukinnan ja siemensadon määrän mittauksista metsäpuilla. Summary: Results of Long-Time Measurements of the Quantity of the Flowering and Seed Crop of Forest Trees. Folia For. 364: 1–60.
Lindgren, D. 1975 The Relationship between Self-Fertilization, Empty Seeds and Seeds Originating from Selfing as a Consequence of Polyembriony. Stud, For. Suec. 126: 1–24.
Müller, G. 1976 Untersuchungen über die natürliche Selbstbefruchtung in Beständen der Fichte (Picea abies (L.) Karst.) und Kiefer (Pinus sylvestris L.) Silvae Gen. 26: 207–217.
Müller-Stark, G. 1979 Estimates of Self- and Cross-Fertilization in a Scots Pine Seed Orchard. Proc. Confer. Bioch. Gen. For. Trees. Umea. Sweden 1978: 170–179.
OCDE 1974 OECD Scheme for the Control of Forest Reproductive Materials Moving in International Trade. Organization for Economic Cooperation and Development, 2 rue André Pascal, 75016 París, Francia.
Rudin, D. y Ekberg, I. 1982 Genetic Structure of Open Pollinated Progenies from a Seed Orchard of Pinus sylvestris. Silva Fenn. 16. (en prensa).
Sarvas, R. 1962 Investigations on the Flowering and Seed Crop of Pinus sylvestris. Commun. Inst. For. Fenn. 53.4: 1–198.
Sarvas, R. 1967 The Annual Period of Development of Forest Trees. Proc. Finn. Acad. Sci. Lett. 1965: 211–231.
Sarvas, R. 1969 Adaptación genética de especies forestales al factor térmico del clima. Documentos de la segunda Consulta mundial sobre mejora de árboles forestales, Wáshington, FAO/UIOIF, FO, TTB 69/2: pág. 191.
Sarvas, R. 1970 Establishment and Registration of Seed Orchards. Folio For. 89: 1–24.
Sarvas, R. 1972 Investigations on the Annual Cycle of Development of Forest Trees, Active Period. Commun. Inst. For. Fenn. 76.3: 1–110.
Shen, H.H., Rudin, D. y Lindgren, D. 1981 Study of the Pollination Pattern in a Scots Pine Seed Orchard by Means of Isozyme Analysis. Silva Genetica 30:7–15.
![]()
![]()
![]()
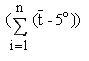 . De esta manera
refleja los efectos conjuntos de la temperatura y el
tiempo.
. De esta manera
refleja los efectos conjuntos de la temperatura y el
tiempo.