![]()
![]()
![]()
| Mr. A. ASSABIR | Mr. F. MENNES |
| ISPM | Projet MOR/81/002 |
| Casablanca | Casablanca |
| MAROC | MAROC |
| Mr.H. BELVEZE | Mr. M. RAMI |
| IFREMER | ISPM |
| Nantes | Casablanca |
| FRANCE | MAROC |
| Mr.S. BENCHERIFI | Mlle R. REFK |
| ISPM | ISPM |
| Casablanca | Casablanca |
| MAROC | MAROC |
| Mr. M. CHBANI IDRISSI | Mr. F. ROEST |
| ISPM | Projet COPACE |
| Casablanca | Dakar |
| MAROC | SENEGAL |
| M.T. DO CHI | Mr. J. ROULLOT |
| Projet MOR/81/002 | Projet MOR/81/002 |
| Casablanca | Casablanca |
| MAROC | MAROC |
| Mr. L. FOSKINE | Mr. M. O/SOUEILIM |
| Ministère de la Pêche, URSS | CNROP |
| Casablanca | Nouadhibou |
| MAROC | MAURITANIE |
| Mr. S. HOLZLOHNER | Mr. I. STAICU |
| IfH | IRCM |
| Rostock | Constantza |
| REPUBLIQUE DEMCRATIQUE D'ALLEMAGNE | ROUMANIE |
| Mr. C. KOUKHORENKO | Mr. M. VAN DER KNAAP |
| Ministère de la Pêche,URSS | Projet COPACE |
| Casablanca | Dakar |
| MAROC | SENEGAL |
| Mr. M. LAMBOEUF | Mr. M. ZOUIRI |
| Projet MOR/81/002 | ISPM |
| Casablanca | Casablanca |
| MAROC | MAROC |
BUREAU REGIONAL DE LA FAO POUR L'AFRIQUE (RAFR)
PROJET POUR LE DEVELOPPEMENT DES PECHES DANS L'ATLANTIQUE CENTRE-EST (INT/81/014)
GROUPE DE TRAVAIL AD HOC SUR LA SARDINE
(Sardina pilchardusWalb.)
I.S.P.M. Casablanca, 26 Novembre - 5 Décembre 1984
ORDRE DU JOUR ANNOTE
OUVERTURE
ADOPTION DE L'ORDRE DU JOUR
RESUME DES DONNES DISPONIBLES
Chaque participant est prié d'apporter à réunion des données détaillées sur :
3.1 Données sur les pêches
3.1.1 Description de la pêcherie, son évolution, nombre de navires, caractéristiques, zones et saisons de pêche, limitations d'accès, engins et maillages utilisés, ports et structures à terre.
3.1.2 Captures et effort, captures totales par espèce cible et secondaire. Effort de pêche, essais de standardisation, tendances. Captures par unité d'effort, indices d'abondance.
3.2 Données biologiques
Clé âge-longeur ou longueur-âge ;
Composition en longueurs des captures ;
Paramètres de croissance (K,L∞,to lecture de l'âge ;
Taux de mortalité (F, M, Z);
Rapport longuer-poids;
Taille et âge de (50%) maturité sexuelle et périodes et zones de reproduction;
Distribution, migrations et distinction de stocks (electrophorèse, etc…)
Abondance/capturabilité en fonction de l'amplitude et du site de l'upwelling et d'autres conditions climatologiques, sélectivité des engins de pêche utilisés.
3.3 Bibliographie
Il est demandé à chaque participant de bien vouloir apporter une liste exhaustive de références d'articles sur la sardine.
4. EVALUATION DES STOCKS
4.1 Modèles globaux de production
4.2 Modèles analytiques, analyses de cohorte
4.3 Prospections acoustiques, des juvéniles et autres études spéciales.
5. MESURES D'AMENAGEMENT 6. RECOMMANDATIONS EN MATIERE DE RECHERCHE ET D'AMENAGEMENT DES PECHERIES PELAGIQUE
ETAT DES STOCKS DE LA SARDINE (Sardina pilchardus Walb.)
DANS LA DIVISION STATISTIQUE 34.1.3
par
N.A. Barkova et L.N. Domanevsky
AtlantNIRO, Kaliningrad, URSS
1. CONDITIONS DU MILIEU
La sardine commune (Sardina pilchardus (Walb.)) vit dans la zone côtière néritique. Du point de vue de leurs conditions climatiques, les eaux côtières de la division 34.1.3 font partie de la zone aride Nord (Atlas physico-géographique du monde, 1964). Les côtes de cette région sont baignées par les eaux superficielles du courant des Canaries. Les caractéristiques hydrologiques de la région sont conditionnées par son appartenance à la zone des alizés à upwelling intense ainsi que par le fait que la région est située sur la limite de deux zones ayant leurs propres systèmes d'upwelling. Nord du Cap Blanc, les eaux de l'upwelling côtier sont celles provenant de la masse d'eau de la partie centrale du Nord Atlantique ; Sud de ce Cap, elles proviennent de l'Atlantique Sud (Tomozak et Miosga 1976). Nord du Cap Blanc, l'upwelling s'intensifie durant la période de réchauffement, limitant ainsi l'augmentation de la température dans cette région (Sorkina 1974). En division 34.1.3 un fort upwelling est observé durant toute l'année (Wooster et al, 1976 ; Sédykh, 1972). L'intensité de l'upwelling change avec la latitude, même à l'intérieur de la région 20–25°N, étant trois fois plus intense à 20°30'N qu'à la latitude 23°30'N (Hagen et Weiss, 1980). Sur toute l'étendue de la région 20–°N, l'upwelling change d'un type à l'autre, créant ainsi un gradient dans les conditions du mdilieu entre la partie Nord de la région aux affinités marocaines et la partie Sud ayant un upwelling du type tropical (Burukovsky 1983). Dans la région 20–25°C, deux saisons hydrologiques peuvent être distinguées (Bernikov, 1969) : la saison froide de la mi-Décembre à la mi-Juillet, quand la température de l'eau dans les strates superficielles est inférieure à la moyenne annuelle, c'est à dire quand elle atteint 17–18°C ; la saison chaude commence en Juillet, quand la température est supérieure à 18°C. Les températures moyennes mensuelles minimum de l'ordre de 17°C sont observées en Février-Mars. Au mois d'Août, la température superficielle atteint son maximum annuel de 19,9°C. La division 34.1.3 est caractérisée pendant toute l'année par les températures plus basses de ses strates superficielles dans la zone côtière par rapport à la zone du large. Entre 20 et 25°, une grande anomalie a été trouvée pendant toute l'année, atteignant un maximum négatif (plus de 6,5°C) en Mai-Juin. Entre Août et Novembre, quand le front tropical est situé plus au Nord, l'anomalie diminue jusqu'aà 2,5°C (Wooster et al., 1976). Pendant le dernier trimestre de l'année, l'anomalie de la température côtière reste élevée. Les valeurs moyennes de la salinité sur plusieurs années sont beaucoup plus élevées que celles dans les eaux océaniques. La période dessalée dure d'Octobre à Février (salinité inférieure à la moyenne annuelle de 35,60%o). Un minimum de 35,05%o est atteint en Novembre.
2. DISTRIBUTION DE LA SARDINE ET MOUVEMENTS SAISONNIERS
Dans la division 34.1.3 la sardine vit au-dessus de fonds de 20–180 m. On ne trouve que quelques bancs isolés dans la zone pélagique au-dessus de fonds supérieurs à 200 m. Dès 1970, lors de l'accroissement de l'abondance de la sardine à la périphérie Sud de son aire de distribution, les profondeurs au-dessus desquelles vit la sardine ont augmenté de 125 à 220 m (Barkova et Domanevsky, 1976). Il doit cependant être mentionné qu'en automne-hiver la gamme des fonds occupés (25–170 m) est beaucoup plus grande qu'en été-automne (30–100 m). Ce phénomène s'explique probablement par le fait qu'en hiver sur la bordure du plateau continental se forme une zone frontale entre les eaux côtières et océaniques caractérisée par de hauts gradients horizontaux en paramètres hydrologiques, particulièrement la température (Kolesnikov et Overko, 1971). La température de l'eau dans cette saison froide est proche de la température préférée de cette espèce (16–19°C. En été la température monte à 29°C au large, restant à 18°C près des côtes.
La distribution de la sardine durant l'année peut être résumée comme suit : en hiver-printemps, quand l'isotherme de 20°C atteint les côtes du Sénégal, se forme entre le Cap Spartel et le Cap Blanc une zone d'eaux froides de 17°C créée par le courant froid des Canaries et l'arrivée des eaux froides des courants de convection (Staicu et al., 1977). Durant cette période, la sardine est répartie sur tout le plateau jusqu'à 19°N. Les plus grandes concentrations de la sardine se forment dans la région où l'upwelling est le plus intense dans la zone 20–25°N. En Avrial-Mai, la sardine se déplace Nord de 22°N en raison du réchauffement de la température au Sud de la région. Ce réchauffement intense des eaux commencant au mois de Juin et durant jusqu'en Septembre avec un maximum de 21–28°C au niveau du Cap Blanc en Août) provoque un déplacement des adultes au Nord de 25°N où ils trouvent leurs températures préférées de 17–19°C. Les plus jeunes individus qui préfèrent de plus hautes températures restent dans la région du Cap Blanc pendant l'été, sans toutefois former des concentrations denses. En Octobre-Novembre, la baisse de température jusqu'à 17–19°C dans toute la région, provoque le déplacement inverse de la sardine vers le Sud (Figure 1).
Cependant, quelques déviations ont été observées ces dernières années dans ce schéma de distribution saisonniére de la sardine. A notre avis, elles sont provoquées par les changements des températures dans la région.
3. LA PONTE
La saison de ponte est relativement longue en division 34.1.3 et les femelles pondeuses sont observées pratiquement le long de l'année. Deux pics principaux sont néanmoins observées annuellement : un pic hivernal en Novembre-Février et un pic printanier en Avril-Mai. Comme remarqué plus haut, leurs périodes et intensités peuvent varier d'année en année. La sardine commence à devenir mature à la taille de 12–13 cm, à la deuxième année de sa vie. La maturation massive a lieu quand la sardine mesure 15–16 cm et la maturation totale à 17–18 cm.
Nous avons examiné de manière détaillée la ponte pendant un nombre d'années (Figure 2). Le début de la ponte, comme déjà remarqué, est observé en Octobre-Novembre. En 1977 la ponte n'a commencé qu'au mois de Décembre, le pic tombant en Février 1978. Le plus grand retard dans le début de la ponte a été observée en 1981/82, quand les femelles pondeuses n'ont pas été observées en grandes quantités qu'au mois de Février. L' intensité de la ponte a en même temps été la plus basse de celle de toutes les années examinées. Dans les années où l'intensité de la ponte était le plus élevé, elle commencait impétuesement en Novembre-Décembre et passait rapidement. Le pic hivernal était alors beaucop plus important que celui du printemps.
Parmi les facteur provoquant la ponte doit être cité la vitesse de refroidissement de l'eau. Dans les années où la ponte commencait tardivement par exemple en 1977, l'évolution annuelle de la température a été peu spectaculaire : en Juin-Octobre, la temperature était d'environ 20°C (19,9–20, 6°C) dans la zone 10–25°N et 15–20°W ; un faible refroidissement a commencé en Novembre et durant la période jusqu'en Avril 1978, la température n'a baissé que de 2,5°C. En hiver, le refroidissement a été faible également. Le réchauffment printanier de 1981–1982 aégalement été faible et la ponte n'a commencé qu'au mois de Février. Contrairement, la ponte est précoce dans les années de refroidissement considérable en automne. Elle commencait à des températures d'en ,pyenne 18,5–21, 7° C pour la zone 20–25°N, 15–20°W. Un deuxiéme pic est observé quand la température moyenne était minimale : 18,0–19,6°C. Dans les années , ce deuxiéme pic a été enregistré en Avril-Mai et une fois en Juillet (1975). Dans le dernier cas, la température montait très lentement à partir du minimum et faisait un saut brusque au mois d'Août seulement (Figure 2).
Etant donné que la sardine pond près des côtes, directement dans les zones de remontée des eaux, nous allons regarder plus en détail l'évolution annuelle de l'upwelling.
En règle générale, le premier pic de la ponte coincide avec la période d'intensité minimale des alizés et de l'upwelling côtier. D'habitude l'upwelling commence à s' intensifier à partir du mois de Février voire Mars. A ce moment prend fin le pic hivernal de la reproduction. En Avril-Mai l'on observe souvent un affaiblissement de l'upwelling, qui se renforce de nouveau au mois de Juin quand il atteint son maximum saisonnier. Dans la période d'affaiblissement en Avril-Mai on observe le deuxième pic de reproduction. La figure 2 montre le pic de printemps est faible lorsque le second minimum d'upwelling n'existe pas ou est déplacé. Evidemment, la perturbation géomagnétique exerce son influence. Dans tous les cas, les pics de ponte observés succèdent à l'affaiblissement de la perturbation géomagnétique (KR). Dans les années où KR était très élevé et il n'y avait pas d'affaiblissement de l'intensité de l'upwelling en Avril-Mai, le deuxième pic de reproduction était très faible, comme en 1982.
4. DESCRIPTION DE LA PECHE
La pêche de la sardine dans la division 34.1.3 a commencé en 1972. Les prises depuis 1979 sont présentées au Tableau 1.
A partir de l'année 1973, cette pêche a étê pratiquée par les RTM des types “Superatlantique” et “Atlantique” et dans une moindre mesure par les BMRT. Les capacités de pêche des deux premiers types de bateaux sont considérablement plus élevées (Tableaux 2-4). La pêche est effectuée à l'aide de chaluts et de sennes. Les concentrations les plus denses et les plus stables de la sardine se forment dans la période, de ponte intense en Décembre-Février et Avril-Mai. Pendant cette période, le rendement des bateaux atteint sont niveau maximum. Dans les prises des chalutiers, la sardine présente 40–60% et dans celles des senneurs 95–100%. Jusqu' en Juin pratiquement, les concentrations commerciales sont présentes le sardine de tout le plateau jusqu'en Mauritanie. De Novembre à Mai, la sardine constitue la base des captures dans la région (jusqu'à 80–95% pendant la pêche spécialisée). En Mars-Avril la sardine commence à se déplacer vers le Nord. Elle se tient en petits bancs isolés et mobiles. La prise des senneurs diminue brusquement. les chalutiers ne peuvent traviller productivement qu'aux petits fonds de 30–60 m. Vers le mois d'Avril, les principales concentrations commerciales sont cantonnées entre 22°40 et 23°45N et plus au Nord. En Juin, les concentrations se trouvent essentiellement au Nord de 23°N. En Mai-Juin, la part de la sardine dans les prises diminue jusqu'à 10–30%, la situation de la pêche étant difficile dans cette période. Au secteur Nord du plateau, les chalutiers capturent la sardine jusqu'en fin Juin, mais les captures deviennent faibles.
Il est à noter que la pêche de la sardine par l'URSS est pratiquée dans un secteur constituant environ 20–30% de l'aréal de la population “saharienne”.
5. EVALUATION DES STOCKS
Une évaluation des stocks de la sardine a été effectuée par la méthode d'analyse des cohortes. Le coefficient de mortalité par pêche choisi est de F = 0,8 pour les générations entièrement capturées.
L'effectif de la génération de 1983 a été déterminé suivant la formule de Domanevsky et Dubrovin (1984) :
V = 8,7 - 2,1 . X1 - 0,01X2
où V = génération de la sardine de l'année i
X1 = intensité de l'upwelling (X . 10-2)
X2 = abundance des sardines matures de l'année i - 1 en milliards d' individus
L'analyse des données sur 11 ans (Tableaux 5–7) fait ressortir plusieurs périodes bien définies : 1) 1972–1974 : explosion de l'abondance, augmentation jusqu'en 1974, quand la biomasse atteint 3 x 106 tonnes (phase initiale du développement de la pêche) ; 2) à partir de l'année 1978, un retour de la biomasse à sa valeur initiale ; 3) à partir de l'année 1979 on observe une stabilisation du stock au niveau de 1.1–1.2 million de tonnes.
A l'heure actuelle, compte tenue de l'apparition d'une génération abondante née en 1982–1983, on peut s'attendre à une nouvelle augmentation de l'abondance de la sardine commune atteignant le niveau des années 1973–1974.
6. REFERENCES BIBLIOGRAPHIQUES
Anonyme, 1964 Informations hydrométéorologiques sur les eaux des côtes Ouest-africaines. Hydrométéoisdat. 1964
Barkova, 1976 N.A., et L.N. Domanevsky, Some peculiarities of sardine distribution and spawning along Northwest Africa. ICES. C.M. 1976/J:6 1–11
Bernikov, 1969 R.G., Variations saisonniéres de la température et la salinité des eaux de la région tropicale du cap Blanc jusqu'au Port de St. Louis. Tr. AtlantNIRO 22 : 13–29
Burukovsky, R.N., 1983 Crevettes des eaux Ouest-Africaines. Distribution, nutrition, relations biocénotiques, valeur commerciale. Thèse Doct.Kaliningrad.
Domanevsky, L.N., 1984 et B.I. Dubrovin, Au sujet du problème des prévisions à long terme de l'abondance des poissons dans la zone d'upwelling côtier. Recueil “Facteurs océanologiques dans les prévisions des captures”, Kaliningrad
Hagen, E., 1980 et R. Weiss, Zur Abhängigkeit der Variabilität der Thunmakrelen-konzentration von der mesoskalen Auftriebsdynamik vor der Küste Nordwestafrikas. Fischerei-Forschung 18 (1):19–31
Kolesnikov, V.G., 1971 et S.M. Overko, Habitat and distribution of horse mackerel in the area off Northwest Africa depending on the oceanological environment. ICES C.M.
Sedykh, K.A., 1972 Circulation verticale des eaux dans la partie Nord-Est de l'Atlantique Tropical en Septembre 1971–Mai 1972. AtlantNIRO 61:3–19
Sorkina, A.I., 1974 Fluctuations saisonnières et interannuelles de l'intensité de la remontée des eaux au large des côtes de l'Afrique du NordOuest en fonction des particularités du régime des alizés : Tr.GOIN 120 : 76–99
Staicu, I. et al., Resurçele biologice ale Oceanului Atlantic est-central. Consiliul national pentre Stiinta si tehnologie. Bull.Inf.Pesc. 4,110p. IRCM, Constantza
Tomczak, M., 1976 et G. Miosga, The sea surface temperature as detected by airborne radiometer in the upwelling region off Cape Blanc NW-Africa. “Meteor” Forsch.Ergebn. A 17 : 1–20
Wooster, W.S., 1976 A.Bakun et D.R. McLain. The seasonal upwelling cyle along the eastern boundary of the North Atlantic. Journ.Mar.Res. 34(2): 131–141
| Année | 1979 | 1980 | 1981 | 1982 | 1983 |
|---|---|---|---|---|---|
| Prises | 110.4 | 168.2 | 185.9 | 164.2 | 173.7 |
| ('000 t) |
| Type de bateau | MOIS | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | ||
| RTM-S Nombre de jours de pêche | 356 | 324 | 148 | 114 | 82 | 131 | 389 | 442 | 335 | 342 | 330 | 228 | |
| Chalut Prise par unité d'effort | 28.8 | 25.9 | 37.1 | 28.9 | 42.2 | 15.4 | 0.1 | 1.6 | 0.1 | 2.3 | 1.4 | 9.7 | |
| RTM-A Nombre de jours de pêche | 254 | 218 | 211 | 184 | 183 | 246 | 364 | 388 | 326 | 352 | 335 | 358 | |
| Chalut Prise par unité d'effort | 23.3 | 22.7 | 23.1 | 21.7 | 28.7 | 18.2 | 3.9 | 0.4 | 0.3 | 3.5 | 2.5 | 5.7 | |
| BMRT-PT Nombre de jours de pêche | 517 | 424 | 404 | 235 | 114 | 85 | 192 | 283 | 226 | 165 | 239 | 246 | |
| Chalut Prise par unité d'effort | 22.7 | 22.0 | 15.6 | 8.7 | 12.6 | 2.4 | - | - | 0.1 | 2.5 | 2.5 | 10.6 | |
| Type de bateau | MOIS | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | ||
| RTM-S Nombre de jours de pêche | 359 | 304 | 246 | 115 | 169 | 169 | 191 | 260 | 381 | 254 | 176 | 228 | |
| Chalut Prise par unité d'effort | 37.0 | 28.0 | 25.9 | 38.1 | 43.6 | 17.0 | - | - | - | - | - | 0.1 | |
| RTM-A Nombre de jours de pêche | 601 | 457 | 373 | 290 | 287 | 302 | 391 | 417 | 590 | 463 | 367 | 214 | |
| Chalut Prise par unité d'effort | 26.3 | 24.4 | 22.1 | 33.6 | 33.1 | 9.7 | 0.02 | - | - | - | 0.4 | 0.2 | |
| BMRT-PT Nombre de jours de pêche | 237 | 197 | 237 | 136 | 146 | 191 | 223 | 368 | 643 | 556 | 239 | 206 | |
| Chalut Prise par unité d'effort | 24.6 | 28.2 | 21.9 | 28.8 | 26.0 | 9.4 | - | 0.2 | - | - | 0.1 | 0.4 | |
| SRTR Nombre de jours de pêche | 666 | 225 | 89 | 18 | - | - | - | 47 | 80 | 288 | 274 | 361 | |
| Sennes Prise par unité d'effort | 25.0 | 23.1 | 5.3 | 26.3 | - | - | - | - | - | 3.8 | 2.9 | 7.4 | |
| Type de bateau | M O I S | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | ||
| RMS-S Nombre de jours de pêche | 235 | 223 | 259 | 306 | 241 | 327 | 306 | 332 | 298 | 335 | 394 | 529 | |
| Chalut Prise par unité d'effort | 3.1 | 27.4 | 35.3 | 36.3 | 36.3 | 18.6 | 0.5 | 0.7 | 0.7 | 1.0 | 4.1 | 11.3 | |
| de pêche RTM-A Nombre de jours de pêche | 186 | 147 | 214 | 286 | 276 | 312 | 248 | 411 | 410 | 334 | 327 | 563 | |
| Chalut Pris par unité d'effort | 2.9 | 23.3 | 24.9 | 23.6 | 26.9 | 19.5 | 1.8 | 1.7 | 9.0 | 21.5 | |||
| BMRT-PT Nombre de jours de pêche | 122 | 78 | 70 | 100 | 91 | 155 | 142 | 332 | 320 | 402 | 412 | 596 | |
| Chalut Prise par unité d'effort | 0.3 | 10.2 | 22.9 | 16.0 | 18.4 | 8.6 | 0.1 | 0.3 | 7.3 | 15.1 | |||
| SRTR Nombre de jours de pêche | 302 | 200 | 155 | 5 | 5 | - | - | 92 | 122 | 106 | 234 | 453 | |
| Sennes Prise par unité d'effort | 7.0 | 13.9 | 9.6 | 6.6 | 15.6 | - | - | 6.3 | 0.05 | - | 12.0 | 18.0 | |
| Année de pêche | GROUPE D'AGE | Total | ||||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | ||
| 1972 | 1 149.97 | 239.62 | 34.02 | 4.11 | 1 427.72 | |||
| 1973 | 538.86 | 906.86 | 178.20 | 16.67 | 0.36 | 2 640.95 | ||
| 1974 | 1 245.00 | 1 197.22 | 585.95 | 57.08 | 0.59 | 3 085.84 | ||
| 1975 | 710.92 | 971.64 | 792.89 | 246.40 | 10.63 | 2 732.48 | ||
| 1976 | 535.98 | 552.11 | 658.34 | 406.07 | 85.62 | 2 238.12 | ||
| 1977 | 472.26 | 442.30 | 403.53 | 251.34 | 76.35 | 1 625.78 | ||
| 1978 | 416.28 | 351.24 | 301.44 | 162.17 | 41.93 | 1 273.06 | ||
| 1979 | 414.00 | 325.20 | 257.78 | 167.36 | 35.87 | 1 200.21 | ||
| 1980 | 434.53 | 309.09 | 244.10 | 164.76 | 62.74 | 7.14 | 0.09 | 1 222.45 |
| 1981 | 431.89 | 334.93 | 221.39 | 133.32 | 52.39 | 12.53 | 0.97 | 1 187.42 |
| 1982 | 356.06 | 334.59 | 237.56 | 108.98 | 24.11 | 5.49 | 4.61 | 1 071.40 |
| 19831 | 1 312.00 | 280.55 | 242.63 | 108.77 | 17.64 | 0.42 | 0.09 | 1 962.10 |
| Année de pêche | GROUPE D'AGE | Total | ||||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | ||
| 1972 | 35 936.57 | 5 209.21 | 531.50 | 48.36 | 41 725.64 | |||
| 1973 | 48 089.23 | 19 714.26 | 2 784.43 | 196.13 | 3.50 | 70 787.55 | ||
| 1974 | 38 906.34 | 26 026.49 | 9 155.47 | 671.52 | 5.77 | 74 765.59 | ||
| 1975 | 22 216.38 | 21 122.68 | 12 388.93 | 2 898.78 | 103.25 | 58 730.02 | ||
| 1976 | 16 749.40 | 12 002.37 | 10 286.53 | 4 777.35 | 831.25 | 44 646.90 | ||
| 1977 | 14 758.13 | 9 180.34 | 6 305.09 | 2 956.94 | 741.30 | 33 941.80 | ||
| 1978 | 13 008.81 | 7 635.76 | 4 709.95 | 1 907.85 | 407.12 | 27 669.49 | ||
| 1979 | 12 504.33 | 7 069.53 | 4 027.76 | 1 968.89 | 348.24 | 25 918.75 | ||
| 1980 | 13 579.06 | 6 719.40 | 3 755.39 | 1 938.31 | 609.09 | 55.32 | 0.70 | 26 657.27 |
| 1981 | 13 496.44 | 7 281.00 | 3 405.95 | 1 568.43 | 508.60 | 97.14 | 7.17 | 26 364.73 |
| 1982 | 11 126.73 | 7 273.66 | 3 654.75 | 1 282.13 | 234.11 | 42.58 | 34.12 | 23 648.08 |
| 19831 | 43 000.00 | 6 098.85 | 3 732.73 | 1 279.70 | 173.30 | 3.29 | 0.70 | 54 288.57 |
1 Données préliminaires
| Année de pêche | GROUPE D'AGE | ||||||
|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | |
| 1972 | 0.00 | 0.03 | 0.40 | 2.03 | |||
| 1973 | 0.01 | 0.17 | 0.82 | 2.93 | 0.80 | ||
| 1974 | 0.01 | 0.14 | 0.55 | 1.27 | 0.80 | ||
| 1975 | 0.02 | 0.12 | 0.35 | 0.65 | 0.80 | ||
| 1976 | 0.00 | 0.04 | 0.65 | 1.26 | 0.80 | ||
| 1977 | 0.06 | 0.07 | 0.60 | 1.38 | 0.80 | ||
| 1978 | 0.01 | 0.04 | 0.27 | 1.10 | 0.80 | ||
| 1979 | 0.02 | 0.03 | 0.13 | 0.57 | 0.80 | ||
| 1980 | 0.02 | 0.08 | 0.27 | 0.74 | 1.24 | 1.44 | 0.80 |
| 1981 | 0.02 | 0.09 | 0.38 | 1.30 | 1.80 | 0.45 | 0.80 |
| 1982 | 0.00 | 0.07 | 0.45 | 1.41 | 3.67 | 3.51 | 0.80 |
| 1983 | 0.16 | 0.53 | 0.85 | 1.67 | 3.17 | 0.80 | |
| Longueur cm | |||
|---|---|---|---|
| 1981 | 1982 | 1983 | |
| 8 | + | - | + |
| 9 | - | - | 0.1 |
| 10 | - | - | 0.1 |
| 11 | + | - | 0.7 |
| 12 | 0.2 | + | 1.1 |
| 13 | 0.4 | 0.07 | 0.5 |
| 14 | 1.2 | 0.05 | 0.2 |
| 15 | 1.3 | 0.08 | 0.7 |
| 16 | 1.5 | 0.3 | 1.8 |
| 17 | 4.4 | 2.9 | 7.4 |
| 18 | 14.5 | 14.9 | 24.1 |
| 19 | 33.7 | 22.2 | 32.8 |
| 20 | 30.2 | 18.3 | 22.7 |
| 21 | 10.2 | 21.8 | 6.6 |
| 22 | 2.1 | 14.2 | 1.0 |
| 23 | 0.3 | 4.3 | 0.1 |
| 24 | + | 0.9 | - |
| 25 | - | + | - |
| Longueur moyenne, cm | 19.70 | 20.59 | 19.28 |
| Masse moyenne, kg | 0.095 | 0.105 | 0.090 |
| Nombre d'exemplaires | 42 246 | 14 635 | 49 481 |
| Age, ans | 1981 | 1982 | 1983 |
|---|---|---|---|
| 1 | 3.1 | 0.2 | 3.2 |
| 2 | 11.4 | 9.8 | 18.1 |
| 3 | 27.3 | 38.2 | 43.7 |
| 4 | 38.7 | 37.2 | 28.1 |
| 5 | 17.7 | 10.6 | 6.7 |
| 6 | 1.8 | 2.4 | 0.2 |
| 7 | 0.03 | 1.6 | 0.03 |
| Age moyen, ans | 4.07 | 4.20 | 3.73 |
| Groupe d'âge | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Suspension (kg) | 0.032 | 0.046 | 0.064 | 0.085 | 0.103 | 0.129 | 0.135 |
Paramètres de croissance de la sardine d'après l'équation de Bertalanffy
k = 0.48 to = -0.39 L = 25.6 W = 147.7
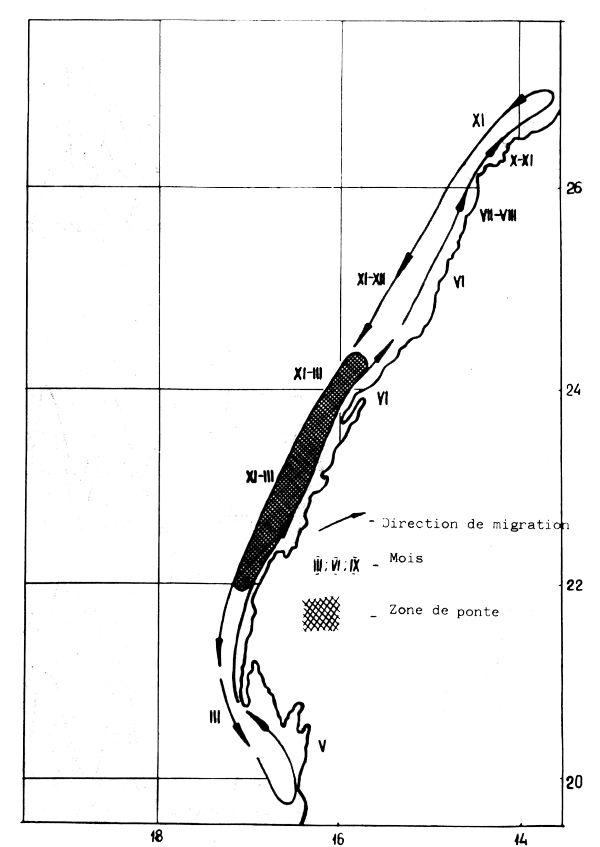
Figure 1 : Schéma annuel de déplacements de la sardine dans la division statistique 34.1.3
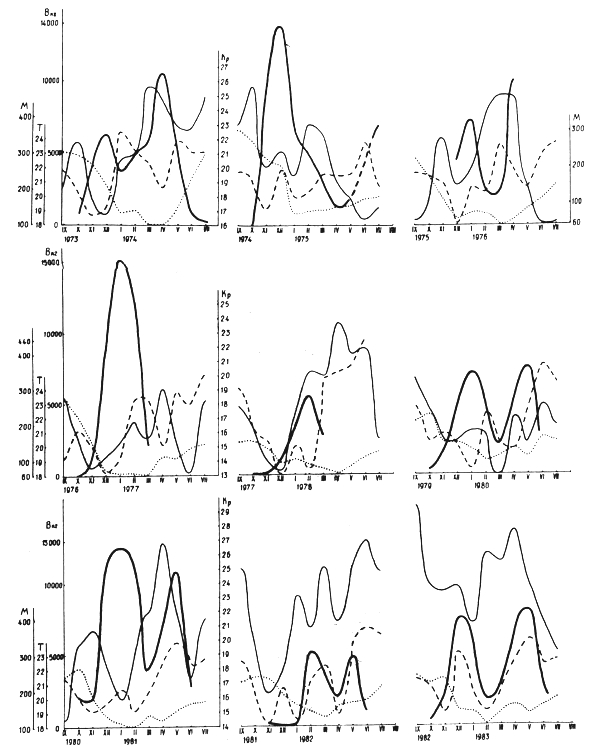
Figure 2 : Intensité de la ponte de la sardine et variations de quelques paramètres du milieu
 | Prises de femelles pondeuses (kg/bateau/jour de pêche) |
 | Perturbation géomagbétique (KR) |
 | Transport d'Ekmann (t/seconde par 100 m de côtes) |
 | Température de la surface |
LA PECHE ROUMAINE DE LA SARDINE (Sardina pilchardus(Walbaum)) DANS LA REGION DU COPACE en 1967–1983
par
I. Staicu et C. Maxim
Institut Roumain de Recherches Marines, Constantza, Roumanie
1. MATERIEL ET METHODES
Dans cette étude de la sardine, on s'est basé principalement sur les observations effectuées directement à bord des chalutiers roumains qui ont pêché dans les divisions 34.1.3 et 34.3.1 pendant la période 1973–1983.
L'échantillonnage a été effectué en conformité avec les instructions de l'Institut (IRCM 1970, 1977, 1980), le matériel étant collecté seulement du secteur Sud de la zone C.
Les échantillons analysés constituaient généralement d'unités de 100–200 exemplaires choisis de quatre points différents d'une capture d'au moins une tonne déchargée sur le pont. Pour la détermination de l'âge, des échantillons furent prélevés selon la méthode stratifiée. La longueur, mesurée au centimétre le plus près est celle à la fourche (lf), les intervalles entre les classes de longueur étant donc de l centimétre.
Le poids moyen par classe de longueur a été déterminé en pesant ensemble tous les individus de cette classe, ainsi que par des pesages individuels, permettant une erreur de plus ou moins 5 grammes. Le sex ratio fut obtenu après dissection.
Pour la détermination de l'âge, des écailles furent prélevées de la zone comprise entre la tête et la nageoire dorsale, au dessus de la ligne latérale. Afin de permettre une bonne lecture, les écailles ont été dégraissées, lavées et fixées entre deux lamelles de verre. Ensuite, elles ont été examinées dans un lecteur du type “Lector-1” de fabrication roumaine (objectif 3,2/50 et illumination 100 W/12v). Le premier Janvier a été considéré comme date de naissance.
Les nombres d'échantillons sont donnés au Tableau 1.
2. LA PECHE
2.1 Description de la pêcherie
Pendant la période 1967–1969, la flottille roumaine était représentée dans la zone du COPACE par deux chalutiers de 3800 TJB, qui pêchaient uniquement avec le chalut de fond d'un maillage étiré de 110 mm, étant donné la faible abondance de la sardine qui constituait une espèce secondaire dans les captures de chinchards et de maquereaux. Bien que les chalutiers roumains fussent également présents dans l'intervalle suivant (1970–1972), la sardine ne fut pas signalée dans les captures. Pendant la période suivante, 1973–1976, les bateaux roumains présents dans la zone mesuraient 2156–3800 TJB et utilisaient le chalut pélagique d'un maillage étiré de 40 mm. Il faut mentionner que l'intervalle 1975–1976 a été consacré à la pêche spécialisée de la sardine. Durant la période 1973–1979, des navires de 2156 TJB ont dominé et en 1980–1983 ceux de 3917 TJB. L'engin utilisé a été surtout le chalut pélagique à mailles étirées de 60 mm.
Les principaux secteurs de pêche pendant la période 1967–1983 sont situés dans la zone C, comprise entre le Sud du Cap Bojador (26°00'N) et le Nord du Cap Blanc (20°00'N). L'activité des chalutiers roumains s'est limitée aux secteurs de concentration maximale de la sardine, situés entre 20°00 et 26°00'N, aux profondeurs de 70–90 m (Figure 1).
2.2 Captures et effort
La sardine a été signalée dans les captures roumaines à partir de l'année 1967, l'on observe une diminution des captures, bien que l'effort total de pêche ait beaucoup augmenté. La sardine disparait ensuite pratiquement des captures roumaines à partir de 1981, quand les chalutiers roumains déplacent leurs activités vers le Sud du Cap Blanc, dans la ZEE de la Mauritanie. En Janvier 1983, des sardines furent signalées brièvement dans le secteur situé près de la limite Nord de la zone à 20°36'N.
Les captures roumaines de la sardine dans la période 1967–1983 sont présentées au Tableau 2, où l'on observe que la plupart des captures provient effectivement de la division 34.1.3 (zone C).
L'effort de pêche du Tableau 3 correspond à l'effort total des chalutiers roumains dans la division 34.1.3 où ont été signalées les plus grandes captures de sardines.
En analysant les captures par unité d'effort, il est évident que la sardine apparait en grandes quantités en Janvier-Avril et Novembre-Décembre, quand l'espèce forme des concentrations de préponte et de ponte (Tableau 4). Pendant ces périodes, la sardine constitue l'espèce principale dans les captures roumaines de la zone Nord du COPACE.
3. DONNEES BIOLOGIQUES
3.1 Structure par longueur et par âge
Dans les captures roumaines, des sardines ont été rencontrées de 11–23 cm de long, âgées de 1–6 ans. Les individus de 17–20 cm, âgés de 3–5 cm sont dominants dans les captures (Tableaux 5 et 6).
3.2 Croissance
Les longueurs et poids moyens par classe d'âge sont présentés aux Tableaux 7 et 8. On observe nettement une diminution de la croissance avec l'âge (Tableau 9).
Les paramètres de croissance ont été calculés par année (Tableau 10).
3.3 Sex ratio
Pendant toute la période d'étude, le sex ratio des sardines dans les captures roumaines a été très voisin de l, avec une légère domination de femelles en 1973 et 1977 et de mâles en 1978 et 1983 (Tableau 11).
4. BIBLIOGRAPHIE
IRCM, 1970 Instructiuni de prelevare a esantioanelor la bordul navelor de pescuit (en roumain). Constantza, 23p.
IRCM, 1977 Recoltarea esantioanelor si datelor de productie pe navele de pescuit oceanic in scopul studierii populatiilor pescaresti (en roumain). Constantza, 62p.
IRCM, 1980 Esantionajul populatiilor exploatabile (en roumain). Constantza, 23p.
Staicu, 1975 I. C. Maxim et I. Panait, Recherches ichtyologiques roumaines dans la zone de l'Océan Atlantique est-central, en 1974. Cercetari Marine IRCM 8:119–135
Staicu, 1976 I. C. Maxim et I. Panait, Recherches roumaines sur l'ichtyofaune de l'Atlantique en face du Sahara espagnol, pendant la période 1973– 1975. Cercetari Marine IRCM 9:191–204
Staicu, I. 1977 et al., Resursele biologice ale Oceanului Atlantic est-central (en roumain). Bul. de inf. pesc. nr.4:110p.
Staicu, I., 1981, Ihtiofauna zonei Oc. Atlantic est-central (en roumain) Halieutica nr. 3(9):72–144.
Staicu, I., 1983 Raport asupra rezultatelor cercetarilor efectuate pe platforma continental mauritaniano in perioada ianuarie-mai 1983. (en roumain) Halieutica nr.3(27) : 5–66
| Année | Total exemplaires | Analyse | lengueur poids | |
|---|---|---|---|---|
| âge | sexe | |||
| 1973 | 100 | 100 | 60 | 100 |
| 1974 | 3.657 | 3.657 | 856 | 3.657 |
| 1975 | 4.797 | 4.797 | 994 | 3.508 |
| 1976 | 20.248 | 20.248 | 3.659 | 20.248 |
| 1977 | 3.995 | 3.995 | 815 | 3.995 |
| 1978 | 3.107 | 3.107 | 700 | 1.900 |
| 1979 | ||||
| 1980 | Il n'y a pas de dennées biologiques | |||
| 1983 | 800 | 800 | 70 | 800 |
| Année | Total | 34.1.3. | 34.3.1. |
|---|---|---|---|
| 1967 | 2 | 2 | - |
| 1968 | 518 | 5 | 513 |
| 1969 | 591 | 256 | 335 |
| 1970 | 122 | 122 | - |
| 1974 | 21.275. | 21.275 | - |
| 1975 | 34.939 | 34.922 | 17 |
| 1976 | 17.641 | 17.641 | - |
| 1977 | 26.968 | 26.771 | 197 |
| 1978 | 10.566 | 10.377 | 189 |
| 1979 | 678 | 599 | 21 |
| 1980 | 4.543 | 4.131 | 410 |
| 1983 | 2.510 | 2.492 | 18 |
| Année | Jours | Noures |
|---|---|---|
| 1967 | - | 1.430 |
| 1968 | - | 787 |
| 1969 | - | 2.590 |
| 1973 | 1.909 | 17.040 |
| 1974 | 2.914 | 27.655 |
| 1975 | 2.818 | 24.556 |
| 1976 | 1.457 | 13.867 |
| 1977 | 2.733 | 29.007 |
| 1978 | 1.379 | 14.327 |
| 1979 | 486 | 4.426 |
| 1980 | 1.036 | 11.371 |
| 1983 | 127 | 1.910 |
| Mois /AnnesAnnées | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1973 | 0,00 | 0,04 | - | - | 0,02 | 0,02 | - | - | - | - | - | - | - |
| 1974 | 0,07 | 0,08 | 0,52 | 1,17 | 2,12 | 2,96 | 3,02 | 1,18 | 0,28 | 0,00 | 0,00 | 0,01 | 0,03 |
| 1975 | 1,37 | 0,62 | 1,73 | 3,65 | 3,37 | 4,11 | 0,78 | 0,16 | 0,04 | 0,14 | - | 0,03 | 0,18 |
| 1976 | 1,27 | 1,39 | 1,75 | 0,90 | 2,91 | 0,82 | - | 0,14 | 0,10 | 0,14 | 0,17 | 1,17 | 2,00 |
| 1977 | 0,83 | 2,70 | 2,52 | 1,62 | 0,05 | 0,04 | 0,01 | 0,33 | 0,00 | 0,14 | 0,00 | 1,25 | 1,24 |
| 1978 | 0,73 | 1,28 | 1,52 | 1,32 | 0,48 | 0,15 | 0,05 | 0,17 | 0,02 | - | 0,03 | 0,08 | 0,28 |
| 1979 | 0,15 | 0,33 | 0,00 | - | 3,40 | 0,00 | 0,04 | 0,01 | 0,10 | 0,21 | 0,50 | - | - |
| 1980 | 0,40 | 4,10 | 3,40 | 0,49 | 0,11 | 0,09 | 0,13 | - | - | - | - | - | - |
| 1983 | 1,31 | 1,31 | - | - | - | - | - | - | - | - | - | - | - |
| Classe de long gueur(cm) | 1973 | 1974 | 1975 | 1976 | 1977 | 1978 | 1979– 1982 | 1983 |
|---|---|---|---|---|---|---|---|---|
| 11 | 0,4 | PAS DE DONNEES | ||||||
| 12 | 1,1 | 0,1 | + | |||||
| 13 | 2,8 | 0,2 | + | 0,2 | ||||
| 14 | 3,4 | 0,5 | + | 1,0 | ||||
| 15 | 4,7 | 0,9 | 0,1 | 1,4 | + | |||
| 16 | 9,6 | 1,6 | 0,4 | 1,8 | 2,4 | |||
| 17 | 21,5 | 4,7 | 1,7 | 5,1 | 8,2 | |||
| 18 | 6,0 | 31,1 | 20,5 | 17,3 | 22,4 | 16,7 | 0,1 | |
| 19 | 32,0 | 15,1 | 42,1 | 40,5 | 40,0 | 32,0 | 23,5 | |
| 20 | 45,0 | 7,3 | 20,7 | 26,6 | 21,0 | 28,4 | 55,6 | |
| 21 | 7,0 | 2,7 | 8,3 | 11,2 | 5,7 | 10,1 | 18,6 | |
| 22 | 2,0 | 0,3 | 0,4 | 2,1 | 1,4 | 1,8 | 1,9 | |
| 23 | 1,0 | 0,1 | + | 0,3 | 0,2 | |||
| Lfm | 19,77 | 17,47 | 18,57 | 19,33 | 18,87 | 19,14 | 19,90 | |
| No.expl. | 100 | 3657 | 4797 | 20248 | 3995 | 3107 | 800 |
| Age/ Année | 1 | 2 | 3 | 4 | 5 | 6 | Age moyen |
|---|---|---|---|---|---|---|---|
| 1973 | 30,0 | 50,0 | 15,0 | 5,0 | 3,95 | ||
| 1974 | 2,9 | 10,7 | 31,4 | 44,2 | 10,8 | - | 3,53 |
| 1975 | 1,5 | 4,0 | 32,9 | 58,8 | 2,8 | - | 3,57 |
| 1976 | - | 0,4 | 19,7 | 55,3 | 22,1 | 2,5 | 4,07 |
| 1977 | - | 1,1 | 28,6 | 59,0 | 7,2 | 4,1 | 3,85 |
| 1978 | - | 0,3 | 13,3 | 60,3 | 22,8 | 3,3 | 4,16 |
| 1979–1982 | PAS DE DONNEES | ||||||
| 1983 | - | - | 8,6 | 77,1 | 10,1 | 4,3 | 4,10 |
| Âge/ Anné | 1 | 2 | 3 | 4 | 5 | 6 | No. exempl. |
|---|---|---|---|---|---|---|---|
| 1973 | - | - | 18,35 | 20,13 | 21,20 | 21,84 | 60 |
| 1974 | 12,87 | 16,61 | 18,56 | 20,12 | 21,17 | - | 856 |
| 1975 | 12,91 | 18,86 | 18,78 | 20,03 | 20,95 | - | 994 |
| 1976 | - | 16,24 | 18,31 | 19,75 | 20,80 | 21,37 | 3659 |
| 1977 | 14,02 | 16,81 | 18,59 | 19,84 | 20,75 | 21,33 | 815 |
| 1978 | - | 16,82 | 18,72 | 19,89 | 20,63 | 21,17 | 700 |
| 1979 | |||||||
| 1980 | PAS DE DONNEES | ||||||
| 1981 | |||||||
| 1982 | |||||||
| 1983 | - | - | 18,33 | 20,38 | 21,15 | 22,00 | 70 |
| Âge/ | 1 | 2 | 3 | 4 | 5 | 6 | No. |
|---|---|---|---|---|---|---|---|
| Anné | exempl. | ||||||
| 1973 | - | - | 79,3 | 110,6 | 124,5 | 135,0 | 60 |
| 1974 | 23,1 | 52,2 | 73,8 | 92,4 | 114,9 | - | 856 |
| 1975 | - | 53,0 | 77,0 | 96,0 | 112,0 | - | 994 |
| 1976 | - | 53,5 | 76,3 | 96,6 | 110,7 | 120,9 | 3659 |
| 1977 | 33,2 | 56,4 | 78,3 | 93,7 | 113,3 | 124,4 | 815 |
| 1978 | - | 58,0 | 79,8 | 92,9 | 109,4 | 125,0 | 700 |
| 1979 | |||||||
| 1980 | |||||||
| 1981 | PAS DE DONNEES | ||||||
| 1982 | |||||||
| 1983 | - | - | 74,4 | 101,2 | 115,0 | 127,8 | 70 |
| 1/2 | 2/3 | 3/4 | 4/5 | 5/6 | |
|---|---|---|---|---|---|
| 1973/74 | - | - | 1,77 | 1,04 | - |
| 1974/75 | 3,99 | 2,17 | 1,47 | 0,83 | - |
| 1975/76 | 3,33 | 1,45 | 0,97 | 0,77 | 0,42 |
| 1976/77 | - | 2,35 | 1,53 | 1,00 | 0,53 |
| 1977/78 | 2,80 | 1,91 | 1,30 | 0,79 | 0,42 |
| Année | a | b | r | k | to | L∞ |
|---|---|---|---|---|---|---|
| 1973 | 2,536 | 0,055 | 0,998 | 0,51 | -0,205 | 22,80 |
| 1974 | 2,824 | 0,018 | 0,996 | 0,42 | -0,950 | 23,08 |
| 1975 | 3,098 | 0,009 | 0,997 | 0,49 | -0,826 | 22,17 |
| 1976 | 2,827 | 0,021 | 0,994 | 0,43 | -0,954 | 22,49 |
| 1977 | 3,106 | 0,009 | 0,991 | 0,39 | -1,480 | 22,56 |
| 1978 | 3,114 | 0,009 | 0,992 | 0,42 | -1,402 | 22,16 |
| 1979 | PAS DE DONNEES | |||||
| 1980 | ||||||
| 1981 | ||||||
| 1982 | ||||||
| 1983 | 2,910 | 0,016 | 0,994 | 0,33 | -1,523 | 23,98 |
| Age/ Année | F | M | Total | ||
|---|---|---|---|---|---|
| no.ex. | % | nr.ex. | % | ||
| 1973 | 65 | 65,0 | 35 | 35,0 | 100 |
| 1974 | 1.860 | 50,8 | 1.794 | 49,2 | 3.657 |
| 1975 | 1.866 | 53,2 | 1.642 | 46.8 | 3.508 |
| 1976 | 10.148 | 50.1 | 10.100 | 49,9 | 20.248 |
| 1977 | 2.045 | 51,1 | 1,950 | 48,9 | 3.995 |
| 1978 | 839 | 44,2 | 1.061 | 55,8 | 1.900 |
| 1983 | 373 | 46,6 | 427 | 53,4 | 800 |
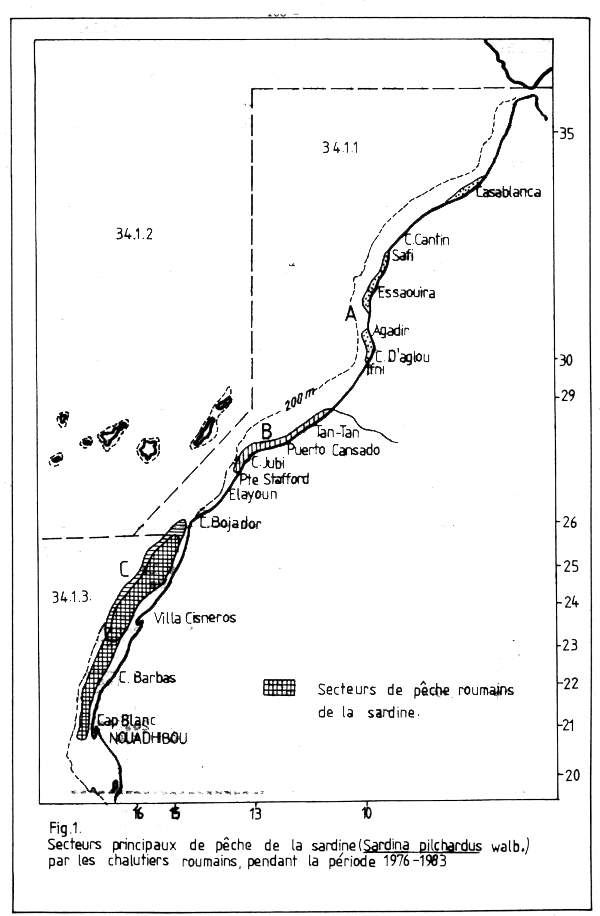
Fig. 1. Secteurs principaux de pêche de la sardine (Sardina pilchardus walb.) par les chalutiers roumains, pendant la période 1976–1983
SARDINE FISHERY AND RESEARCH (Division 34.1.3) OF THE GERMAN DEMOCRATIC REPUBLIC
by
S. Holzlöhner
Institute of Deep Sea Fishery and Fish Processing
1. THE FISHERY
Since 1965–1966 the sardine was fished in the western part of the English Channel using small vessels (Kutter, Logger) (Holzlöhner, 1968 (a and b).
A sardine fishery off Northwest Africa took place from 1974 to 1980 in which the following types of vessels were used : Tropic, 1340 HP; FVS Ib, 2300 HP and FVS III, 3880 HP.
The sardine catches from Northwest Africa waters (division 34.1.3) are presented in Table 1.
| 1974 | 1561 |
| 1975 | 1963 |
| 1976 | 7410 |
| 1977 | 13159 |
| 1978 | 196 |
| 1979 | 2709 |
| 1980 | 10370 |
In Table 2 catches per hour and per vessel type are presented for the main fishing months (February-July). Table 3 summarizes the catches per unit effort per vessel type for the period 1974–1977.
Table 2 : GDR Sardine catches per fishing hour in West African waters 1974–1977
| Vessels type | Year | Month | Catch per hour (t) | Latitude (°N) |
|---|---|---|---|---|
| Tropik | 1974 | IV | 3,2 | 25°00' – 26°15' |
| 1975 | II | 3,2 | 23°20' – 25°30' | |
| IV | 3,4 | 22°50' – 23°45' | ||
| V | 2,7 | 22°30' – 23°40' | ||
| 1976 | II | 3,6 | 23°00' – 25°10' | |
| III | 3,6 | 23°40' – 25°10' | ||
| FVS Ib | 1976 | II | 6,4 | 23°40' – 25°10' |
| III | 6,2 | 23°40' – 25°10' | ||
| 1977 | II | 2,5 | 22°45' – 23°55' | |
| III | 2,1 | 22°55' – 24°00' | ||
| IV | 2,6 | 22°45' – 24°00' | ||
| V | 2,8 | 23°00' – 23°45' | ||
| FVS III | 1977 | I | 3,3 | 23°35' – 24°00' |
| II | 4,0 | 22°55' – 24°00' | ||
| III | 4,2 | 20°35' – 21°00' | ||
| 22°50' – 23°45' | ||||
| IV | 5,1 | 22°50' – 24°00' | ||
| V | 4,8 | 22°55' – 24°00' | ||
| VI | 4,6 | 22°50' – 23°40' |
| 1974 | 1975 | 1976 | 1977 | |
|---|---|---|---|---|
| Tropik (III–IV) | 3,2 | 3,4 | 3,6 | |
| FVS Ib (II–III) | 6,3 | 2,3 | ||
| FVS III (II–III) | 4,1 |
The differences from year to year are mainly caused by stock fluctuations in zone C.
From 1974 to 1976 the stock was supported by the very strong yearclass born in 1971, and by the above average classes 1972 and 1983.
In 1977 the size of the stock decreased sharply which was illustrated by the fact that the FVS III vessels caught only 66% of the amount fished by the smaller FVS Ib vessels in 1976.
In the sardine fishery only pelagic trawling gear was used, before 1975 the mesh trawl and afterwards the line trawl.
The catches consisted for nearly 100 per cent of sardine, the by-catch being of no great importance.
2. RESEARCH
The Institute of Deep Sea Fishery and Fish Processing of the GDR based at Rostock started its first biological observations on Northwest African fish stocks in 1959, and from 1971 more specifically worked on sardine. Some experience in sardine research was already obtained before 1971 as the GDR had investigated and sampled the sardine stock in the western part of the English Channel since 1965 (Holzlöhner, 1974, Borrmann and Holzlöhner, 1976).
From 1971/72 to 1976/77 research off Northwest Africa concentrated on the development of the sardine stock in relation with the decrease of the traditional target species in this region, Scomber japonicus (Holzlöhner, 1975).
During the cruises of the R/V “Ernst Haeckel” in the spring of 1972 and 1973, first observations were made of the pelag trachurus and Sardinella aurita were years, Scomber japonicus, Trachurus trachurus and Sardinella aurita were studied. Surveys of the R/V “Alexander von Humboldt” of the Institute of Marine Research (IfM) of the Academy of Sciences of the GDR (1971–1973) contributed very much to the study of the annual variations in upwelling strength.
From 1974 the research on the sardine stock was intensified. Beside stock analysis the feeding habits of the sardine were also studied.
During the GDR cruises in the Northwest African region from 1974 to 1976, investigations were made on school formation and the structure of the sardine stock. In these years the virtual population analysis (VPA) was introduced. This method and the results of some hydroacoustic surveys were used in order to estimate the sardine stock between Cape Bojador and Cape Blanc.
In 1976 was studied the relation between the sardine and its environment. Based on some preliminary experiments in 1974, biological and oceanographical data could be combined. Oceanographical observations were made at anchor stations as well as during a hydroacoustic survey.
The short-term variations in oceanographical and biological parameters were studies as well as the food intake, the formation of schools and availability of the sardine.
In the period from 1982 to 1984 investigations on sardine were limited to some by-catches of factory trawlers off the northern part of Mauritania.
Every year during spring time (February-April) some sardine schools migrate southwards into northern Mauritanien waters down to 20°N.
In the beginning of 1980 the surface temperature off Mauritania was measured continuously. These observations are of great interest for the study of the long-term variations in the abundance of sardine (see Annex 6).
In 1983, sardines imported from Morocco were sampled in the GDR. The sardines were caught by the Moroccan-GDR joint venture PIMARDA, using 1000 HP factory trawlers.
The samples from 1971 to 1975 and 1983 (partly, except November) originate from factory and research ships inside the main distribution area of the sardine of zone 34.1.1.
The samples from 1982, from November 1983 and from 1984 are to be processed with caution because they originate from the southern periphery of the distribution area off North Mauritania. They are only useful in combination with samples from countries fishing in the centre of the distribution area.
In Table 4 is summarized the age structure of the sardine stocks resulting from all GDR observations. Detailed length and age tables for the years 1971–1975 can be found in the ICES paper of Holzlöhner (1975), and for the years 1982–1984 are available upon request at the CECAF Project.
Important conclusions from these data are that the year class of 1971 was very strong, and that the year classes 1972 and 1973 were above average. Probably again above average was year class 1981, but it appeared from hydroacoustic observations and catch and catch effort levels that it never reached the abundance of 1971 class.
Table 5 shows the result of the VPA for the period of 1972–1980 based on GDR data and the international catch data available in 1981.
The maximum biomass in 1974 of somewhat less than 4 million tons is obviously connected with the very good year class of 1971. The above average classes 1972 and 1973 were probably responsible for the stock levels of 1975 and 1976 of 3.7 and 2.6 million tons respectively. Because of the lack of good year classes, in 1977–1980 the biomass dropped to between 1.1 and 1.8 million tons.
Hydroacoustic observations in the years 1972–1975 measured the distribution area of the main stock C. The results are given in Table 6.
| 1972 | 1973 | 1974 | 1975 |
|---|---|---|---|
| 450 | 2100 | 3200 | 2600 |
| Year | Month | Latitude (°N) | Water depth (m) | Age groups (°/00) | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | ||||
| 1971 | V VI | 20°30'–21°05' | 40 – 96 | 0 | 30 | 970 | 0 | 0 |
| VI | 20°12 | 80 – 90 | 0 | 1000 | 0 | 0 | 0 | |
| 1972 | VI VII | 24°21' – 24°28' | 35 – 50 | 0 | 500 | 490 | 10 | 0 |
| VI | 21°17' | 80 | 0 | 1000 | 0 | 0 | 0 | |
| VI | 28°19' | 44 | 0 | 0 | 1000 | 0 | 0 | |
| 1973. | V. | 23°25'–25°15' | 40 – 78 | 0 | 0 | 950 | 50 | 0 |
| 1974. | V | 23°15–25°25' | 30 – 65 | 0 | 10 | 290 | 700 | 0 |
| 1975. | III V | 22°40'–25°20'. | 30 – 75 | 20 | 0 | 240 | 670 | 70 |
| 1982. | X. | 20°00–20°35' | 30 | 0 | 1000 | 0 | 0 | 0 |
| 1983. | II. | 24°N | 60 | 0 | 138 | 854 | 8 | 0 |
| III. | 24°N | 60 | 0 | 0 | 1000 | 0 | 0 | |
| V. | 24°N | 30 – 45 | 0 | 0 | 1000 | 0 | 0 | |
| VI. | 24°N | 30 – 45. | 0 | 0 | 996 | 4 | 0 | |
| XI. | 20°00'–20°35' | 38 | 0 | 991 | 9 | 0 | 0 | |
| 1984. | III. | 20°00'–20°35' | 25 – 32 | 20 | 611 | 223 | 28 333 | 8 |
| III. | 20°00'–20°35' | 80 – 87 | 0 | 0 | 921 | 97 921 | 0 | |
| IV. | 20°00'–20°55' | 38 – 55 | 69 | 490 | 37 | 40 | 40 | |
| Year | Catch (t) | STOCK IN NUMBERS (106) | STOCK | WEIGHTED (×1000t) | FISHING MORTALITY | |||
|---|---|---|---|---|---|---|---|---|
| N1 | N1+ | N3+ | P1+ | P3+ | F1+ | F3+ | ||
| 1972 | 138.6 | 39394 | 48484 | 2212 | 1742 | 180 | 0.083 | 1.430 |
| 1973 | 238.8 | 50246 | 75597 | 3731 | 3077 | 302 | 0.076 | 0.740 |
| 1974 | 422.5 | 45839 | 84741 | 11518 | 3930 | 930 | 0.109 | 0.544 |
| 1975 | 793.1 | 27212 | 69547 | 17303 | 3734 | 1418 | 0.148 | 0.477 |
| 1976 | 660.0 | 6044 | 39628 | 18784 | 2639 | 1560 | 0.390 | 0.777 |
| 1977 | 618.0 | 3812 | 20003 | 12930 | 1394 | 1065 | 0.737 | 1.026 |
| 1978 | 212.9 | 21614 | 27715 | 4281 | 1110 | 372 | 0.194 | 1.187 |
| 1979 | 115.8 | 17553 | 31000 | 1621 | 1370 | 139 | 0.073 | 0.574 |
| 1980 | 207.5 | 25000 | 40800 | 6700 | 1820 | 539 | 0.108 | 0.439 |
These hydroacoustic findings correspond with the results of the VPA (Table 5), the age structure (Table 4) and with the catches per unit effort (Table 3).
The distribution area which is correlated with the biomass shows a maximum value in 1974, followed by 1975.
3. LITERATURE
See Annexe 9
CONTRIBUTION TO THE STUDY OF SURFACE TEMPERATURE VARIATIONS
IN THE NORTHWEST AFRICAN UPWELLING REGION, 1981–1983
FROM OBSERVATIONS OF GDR FACTORY TRAWLERS
by
S. Holzlöhner
Monthly mean surface temperatures were calculated from the 12 daily measurements made by the factory trawlers on the fishing grounds. Days of absence from the grounds were excluded from the calculations.
We assume that monthly mean values based on over 300 single observations correctly describe surface temperatures variations above the main fish concentrations. The typical mesoscale variations in temperatures of short duration will thus be eliminated. We can also assume that on the average a factory trawler will show a fixed movement based on the results of hydroacoustic research and exchange of catch information. The target species in all years and months investigated was Trachurus trecae. We may therefore assume that temperatures are comparable annually. Surface temperatures above concentrations of T. trecae are higher than above sardines. However, in the present paper we limit our discussions only to the obvious tendencies in temperature development throughout the year. It is necessary to compare our data from the northern Mauritanian waters with those obtained North of Cape Blanc by coastal stations making similar continuous measurements.
The surface temperature data in Table 1 were obtained from the factory trawlers fishing within the area of 20°00'N–20°35'N, between the isobathes 30–80 m, during the warmest time of the year (August-November) :
| 1980 | 1981 | 1982 | 1983 | |
|---|---|---|---|---|
| August | 22.1 | 25.5 | 24.6 | |
| September | 21.0 | 27.0 | 23.6 | 23.4 |
| October | 20.1 | 21.6 | 22.4 | |
| November | 19.7 | 21.7 | 20.1 | |
| Mean | 20.6 | 24.0 | 22.7 |
These data show that 1981 was apparently a very warm year off northern Mauritania. In the warmest months, August and September, the tropical font moved North of Cape Blanc. The years 1982–1983 show a gradual decrease to lower temperatures, although their absolute values were higher than those for 1980.
GDR INVESTIGATIONS INTO THE RELATIONSHIP
BETWEEN ENVIRONMENTAL CONDITIONS AND THE DISTRIBUTION
of Sardina pilchardus (Walb.)
by
S. Holzlöhner (*)
1. INTRODUCTION
The majority of the results presented in this paper was obtained during the seventh expedition of R/V “ALEXANDER VON HUMBOLDT”, which took place between 28 January and 14 April 1976. In addition to oceanographic measurements, the vessel also made hydroacoustic observations. Biological data on fish stocks were simultaneously collected onboard the factory trawlers “ERICH WEINERT” and “GRANITZ”.
2. RESULTS OF THE COMBINED OCEANOGRAPHICAL-HYDROACOUSTIC SURVEY
The transects followed by the R/V “ALEXANDER VON HUMBOLDT” on 25–31 March and the position of the sardine fishing stations in February and March are shown in Figure 1.
Oceanographic conditions prevailing during the survey were caracterized by upwelling processes decreasing in intensity from South to North.
Figure 2 shows the North-South profile along the 100 m isobath of the parameters temperature, phosphate, nitrate and chlorophyll-a. It appears from this Figure that there is a general decreasing trend in the upwelling intensity from South to North. In the southern area, whirls occur, which according to Hagen (1977) can enhance the vertical upwelling speed by one order of magnitude. In the North, at station 245 a secondary maximum in chlorophyll-a a concentration disturbs the general trend. This can be explained from the local topography : the 40–80 m isobaths change direction here, turning from parallel to perpendicular to the coastline. The slope thus created leads within the water masses of the Canary Current to a local upwelling phenomenon.
(*) Traduit de l'Allemand au Projet COPACE Translated from German at the CECAF Project
The hydroacoustic observations made during this survey showed interesting parallels to the oceanographic measurements. The quantities of sardines (expressed in school volume) and chlorophyll-a determined lead to two sets of comparable data, one for the southern part and one for the northern part of the area (Holzlöhner and Nehring, 1978) as is shown in Table 1.
| School volume (m3) | Mean chlorophyll-a concentration (mg/m2) | |
|---|---|---|
| Southern area (22–24°N) | 650 × 103 | 76.8 |
| Northern area (24–26°N) | 366 × 103 | 31.2 |
Interesting conclusions can also be drawn from the comparision of chlorophyll-a/phaeopigment concentration and sardine distribution (Holzlöhner and Nehring, 1978). The ratio chlorophyll-a/phaeopigment is a measure of the photosynthetically active chlorophyll-a. It was found that in the South on the average more living plankton was present than in the North. At station 245 mithin the northern area the above mentioned anomaly was again apparent. According to the results of the hydroacoustic survey, sardines appear to prefer such thermally restricted areas.
All sardine schools detected during the sruvey were compared with the upwelling indicators temperature and phosphate measured within the schools (Holzlöhner and Nehring 1978, Nehring 1978). Above average sardine concentrations were found in water bodies with temperatures of 15.0°–16.5° (-17.0°C and phosphate concentrations of 0.2–0.8 μugat/1). On transect II in the southern part of the study area, which was much richer in sardine occurence than the neighbouring transects, the main concentrations were found at 13.5°–16°OC and 0.4–1.4 μgat/1 of phosphate. In the area of maximum sardine occurrence, over water depths of 80–100 m, water of 15.0°–16.0°C and phosphate concentrations of 0.5–0.6 μugat/1 rose to the surface. Along transect XIV where hardly any fish was found, the 16.0°C isotherm and the 0.4 ugat/1 phosphate isoline were located at about 200 m depth.
3. RESULTS OF ANCHOR STATION OBSERVATIONS
From 19–23 February and 8–11 March 1976, R/V “ALEXANDER VON HUMBOLDT” occupied an anchor station at 23°19'N and 16°48'W above 65 m depth. Simultaneously, biological data were collected and test fishing undertaken on board the factory trawler “ERICH WEINERT (Holzlöhner and Nehring, 1978).” During the first period at anchor, fishing took place in the immediate vicinity, between 22°45' and 23°30'N over water depths of 60–90 m. During the second anchor stationfishing was done somewhat more northerly, between 23°45' and 24°35'N above 60–120 m depth. Figure 3 presents the chlorophyll-a concentrations found and the catches per unit effort of sardine caught by the factory trawler. In general, a good correlation was found between these two parameters, often even on a daily basis. During the second anchor station, when fishing was done more northerly, the correlation was less pronounced but still easily recognizable. Apparently, within half a degree of latitude, temporary mesoscale variations overlap spatial variations.
These parallel fluctuations of chlorophyll-a content and catches per unit of effort are explained by Holzlöhner and Peters (1976), Holzlöhner (1978) and Nieland (1980) on the basis of the fact that phytoplankton constitutes the preferred food of the sardine. Daily fluctuations in chlorophyll-a concentration are apparently caused by mesoscale variations in upwelling intensity.
4. REFERENCES
See bibliography, Annexe 9
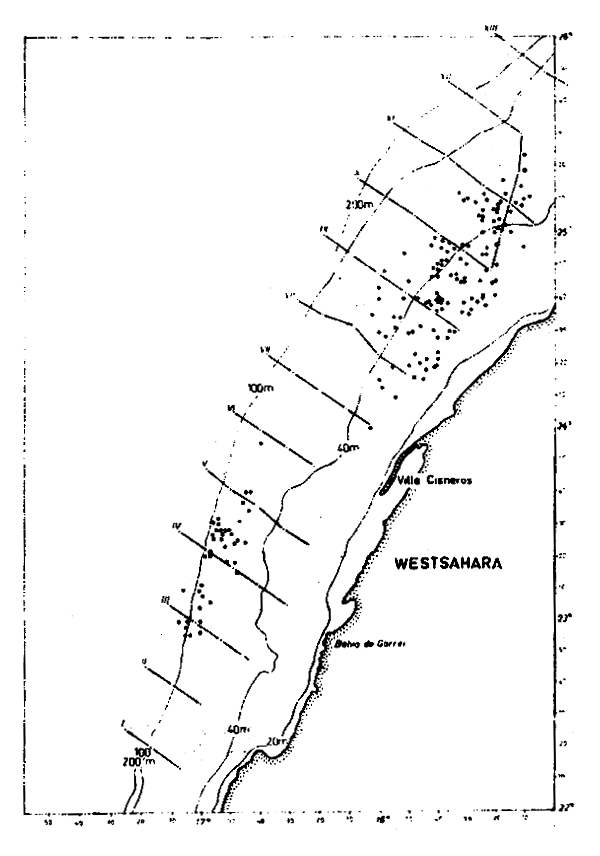
Figure 1 : Position of oceanographic and hydroacoustic transects followed the R/V “ALEXANDER VON HUMBOLDI” survey (25–31 March 1976) and sardine fishing stations in February/March 1976
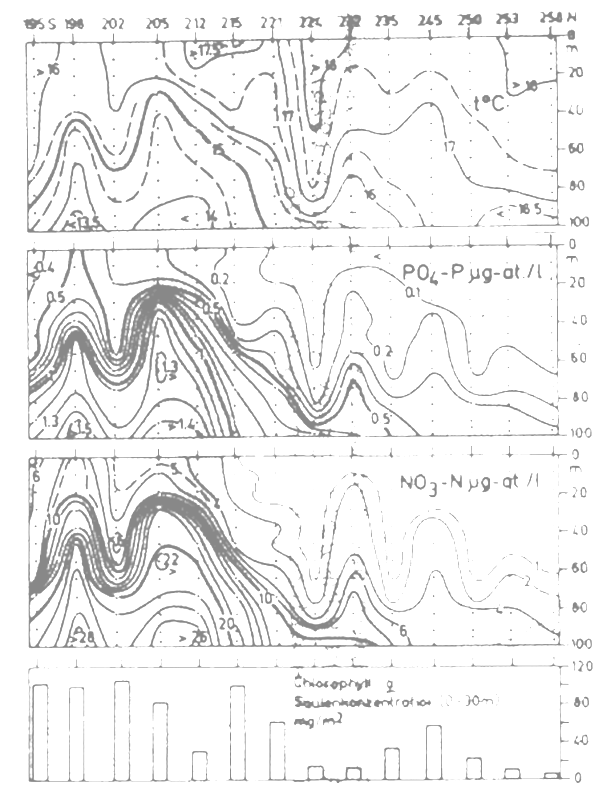
Figure 2 : Vertical distribution of temperature phosphate nitrate and chlorophylla-a, during the R/V “ALEXANDER VON HUMBOLDI” survey, 25–31 March 1976
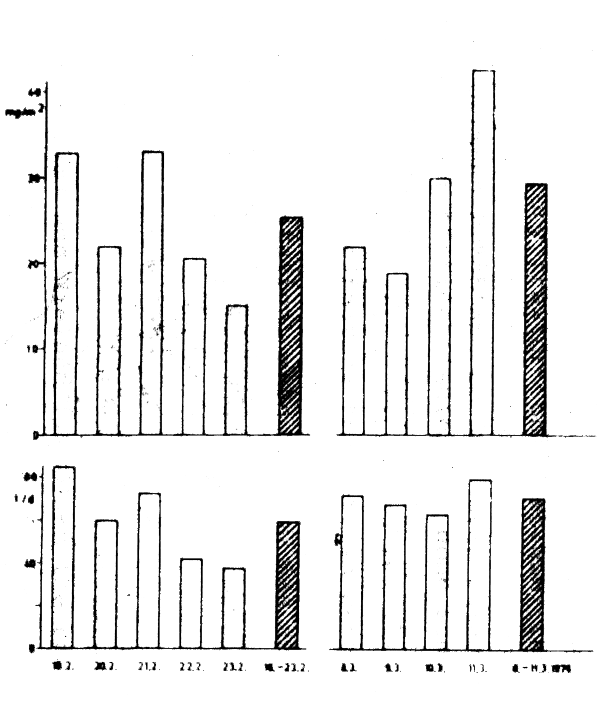
Figure 3 : Variability of chlorophyll-a concentration during the anchor station measurements in March 1976 Above : average chlorophyll-a content in water column Below : catch per fishing day
A NON LINEAR CATCH AT AGE ANALYSIS OF THE CENTRAL SUBPOPULATION OF THE ATLANTIC MOROOCAN SARDINE (Sardina pilchardus) (1)
by
Mohammed Rami
ISPM, Casablanca
The sardine, Sardine pilchardus, is commercially and ecologically the most important pelagic resource of the coast of northwest Africa. Estimation of parameters of this population has been the principal concern of the CECAF ad hoc Working groups and the fisheries scientists from the countries involved in this fishery (FAO, 1978 and 1980). To obtain these estimates, this committee employed mainly catch-effort methods and cohort analysis techniques. These procedures, however, cannot be relied upon for the following reasons.
The scientific community agree now that traditional fishery methods employing catch per unit of effort (CPUE) measurements especially when applied to schooling stocks, have been insufficiently developed as yet to allow rigorous assessments. In multi-gear and multispecies fisheries the inability to standardize and repartition effort make it difficult to maintain time series of consistent CPUE estimates. Interpretation of these data become further complicated by the changing nature of fisheries and variability in the availability of the target species. These problems together with a desire to describe stock abundance in absolute numbers rather than by a relative index contributed directly to the development of theory to estimate fish stock sizes and fishing mortalities from catch-at-age data without reliance on CPUE. These techniques, referred to in the literature as VPA and/or cohort analysis, are due toFry (1949), Gulland (1965), Pope (1972) and others.
In general these methods require on assumption about the fishing mortality (or the population size) for the oldest age of the year class for whicha catch number is available. Estimates of population number and fishing mortality are afterwards calculated for earlier ages of the year class. But, in addition to the arbitrary choice of this terminal rate of fishing mortality which can lead to serious errors, particularly with older fish and most recent year classes, there are more parameters to be estimated than there are observations. For instance, each mortality estimate is supported by somewhat less than one observation.
(1) Summary, Master of science thesis, University of Washington, 1984, 87p., (unpublished)
Therefore, a methodology that fits all the catch-at-age data points simultaneously rather than in a piece-meal fashion and that has a higher observation-to-parameter ration would be more appropriate.
Pope (1974) suggested a model based on Baranov's catch equation and assumed that the instantaneous fishing mortality rate may be partitioned into selectivity (S) as a function of age and full recruitment fishing mortality (F) as a function of year. Since the same S and F are applied to several year classes in this procedure, the parameter estimates are supported by more than one observation each. This permits a meaningful goodness of fit measure to be calculated. Later in 1976, Doubleday attempted linearization techniques, a process of iterative linear approximation and estimation, to obtain least squares estimates of selectivity, fishing mortality rate and population size. This resulted in what became known as Doubleday's model. Recently, Deriso, Quinn and Neal (1984) employed the Marquardt non-linear least squares regression technique to solve the Doubleday's mathematical formulation. This new method of catch-at-age analysis (CAGEAN) was codified in FORTRAN 77. The CAGEAN system is very flexible and accommodates itself easily to the inclusion of extra information such as fishing effort and stock recruitment relationships, and has many other features such as the determination of the variability of the parameter estimates by means of the Monte Carto bootstrap technique (Efron, 1982). Other features are discussed in Rami (1984).
This non-linear method was applied to the 1976–1982 catch-at-age data from the sardine central subpopulation (Table 1). Results of this age structured population model indicated that the exploitable annual surplus production (EASP) was smaller than the commercial catch during the years 1976 and 1977, and greater thereafter (Figure 1). The current average EASP, estimated for the period 1978–1981, was found to range (in 1000's of metric tons) from 341 to 524.
Furthermore, the conventional Gulland's VPA was used to test the goodness fit of the non-linear results (Pope and Shepherd, 1982). This method, initiated with the estimates of the terminal population number obtained by CAGEAN, produced nearly identical results (Figure 2).
Finally, the trend in the available CPUE values was shown to correspond with that of exploitable biomass (Figure 3). Effort was than incorporated in the non-linear method are found to improve the variances of the parameters estimates. Results from this run are shown in Table 2 along with the definition of some of the terms used.
LITERATURE CITED
Deriso, R.B., 1984 T.J. Quinn II and P.R. Neal, Catch-at-age Analysis with auxiliary information. Internation.Pacific Halibut Commission, working paper
Efron, B., 1982 The Jacknife, the bootstrap, and other resampling plans. CBMS-NSF Regional Conference Series in Applied Mathematics, 38 SIAM Arrowsmith Ltd.
FAO, 1978 Comité des Pêches pour l'Atlantique Centre-Est. Rapport du groupe de travail ad hoc sur la sardine COPACE/PACE Series 78/7
FAO, 1980 Comité des Pêches pour l'Atlantique Centre-Est, Rapport de la deuxième réunion du Groupe de travail sur la sardine. COPACE/PACE Series 78/15
Rami, M., 1984 A non-linear Catch-at-age Analysis of the Central Subpopulation of the Atlantic Moroccan Sardine (Sardine pilchardus Walbum). Master of Science Thesis
| Age | |||||||
|---|---|---|---|---|---|---|---|
| Year | 0 | 1 | 2 | 3 | 4 | 5 | 6+ |
| 1976 | 398 | 1404 | 1970 | 544 | 497 | 102 | 10 |
| 1977 | 477 | 693 | 860 | 633 | 164 | 207 | 23 |
| 1978 | 840 | 662 | 663 | 430 | 305 | 222 | 189 |
| 1979 | 202 | 3088 | 923 | 263 | 122 | 108 | 85 |
| 1980 | 2088 | 1007 | 2043 | 432 | 79 | 53 | 89 |
| 1981 | 825 | 2733 | 1062 | 1152 | 69 | 26 | 25 |
| 1982 | 1175 | 860 | 1865 | 563 | 700 | 77 | 46 |
| Years | Total Biomass | Exploitable Biomass | EASP | Fishing Mortality | Age | Selectivity Coefficients |
|---|---|---|---|---|---|---|
| 1976 | 1675(460) | 1450(433) | -27(74) | 0.191(0.0584) | ||
| 1977 | 1324(406) | 1131(370) | 45(34) | 0.191(0.0646) | 0 | 0.14(0.0220) |
| 1978 | 1359(468) | 942(352) | 433(195) | 0.222(0.0800) | ||
| 1979 | 1682(647) | 1159(525) | 474(117) | 0.172(0.0613) | 1 | 0.55(0.0736) |
| 1980 | 1866(769) | 1379(619) | 489(223) | 0.176(0.0598) | ||
| 1981 | 2213(941) | 1602(776) | 444(143) | 0.168(0.0591) | ||
| 1982 | 2174(942) | 1750(821) | 0.236(0.091) |
Age - selectivity is the proportion of age class vulnerable to fishing gear, it results from both partial recruitment of year classes into the fishery and selectivity of gear.
Exploitable biomass is the sum over age classes of age specific exploitable biomass. The latter is the product of age-specific biomass times the proportion of this class fully vulnerable to fishing gear.
Annual surplus production (ASP) represents the annual change in biomass adjusted for the removals by the fishery, it is calculated as the biomass at the biginning of the year minus the biomass at the start of the prior year plus annual catch. Exploitable annual surplus production (EASP) is calculated in a similar manner, except that this applies only to the exploitable part of the stock.
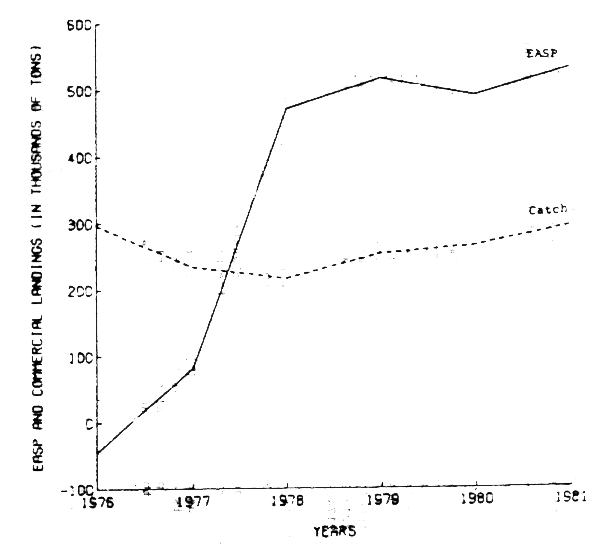
Figure 1. comparison of commercial catch with EASP obtained from nonlinear catch age analysis
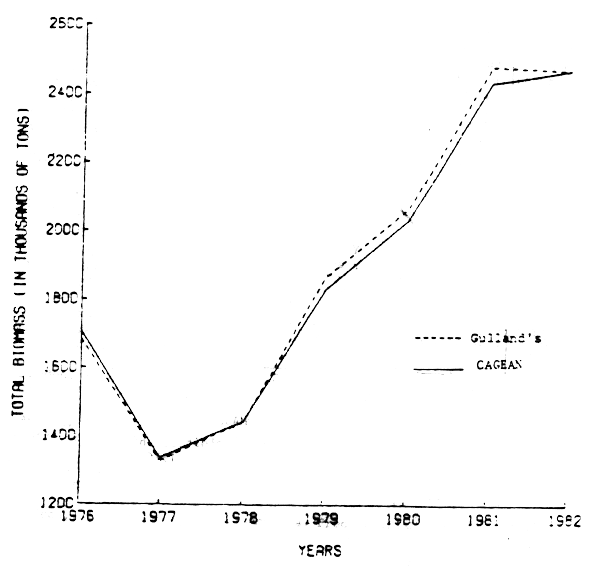
Figure 2. Comparison of total biomass estimates from Gulland's method with those from nonlinear catch age analysis.
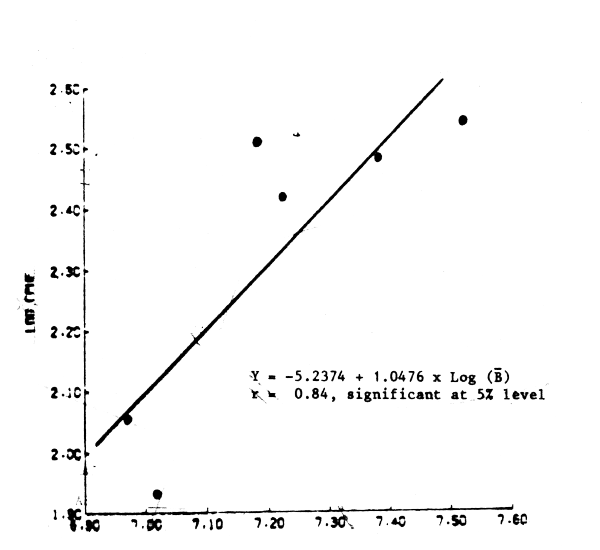
Fig. 3 Relationship between mean exploitable biomass and cpue of the Moroccan seiners, 1976–1982
![]()
![]()
![]()