![]()
![]()
![]()
by
J.M. Stretta
The study of the environment of tuna in the Gulf of Guinea began with the works of Postel (1955 and 1955a) on the tuna boats Gerard Treca and Albacore. Postel defined the ranges of distribution of the principal species and defined the major features of the relation between tuna and their environment. Afterwards, Bane (1963) analyzed data of the Columbia cruise in 1959–1960 on the African coasts of Liberia to Angola. The study of the tuna/environment relationship was continued at ORSTOM by the works of Le Guen et al., (1965). However as Postel (1967) pointed out, “information acquired on distribution and abundance of tuna in this region of the gulf are due much more to the data analysis of commercial fishing than to the study from oceanographic expeditions”. At present, there is a good statistical coverage of tuna fishing for surface fisheries as well as for those by longline. This large quantity of information permits much more precise analyses in space and time than if there were only data issued from oceanographic expeditions.
The availability in space and time of tuna is subject to strong variations, and it is only when certain environmental conditions are linked that tuna gather. In other terms, tuna distribution in a given ecosystem depends directly on physico-chemical and biological parameters and their interactions, that is to say their synergetic action. These environment parameters will have direct and indirect effects on tuna. Direct effects on mortality of the species, on their growth by influencing factors favoring the presence of food and recruitment; indirect on migration, on the availability of tuna to fishing gears and therefore on fishing effort.
Tuna can be considered as the penultimate link in the food chain, man being the last; exploitation by man should not be thought as a removal “from the exterior” but as an insertion of an additional predator in the ecosystem. Schematically this chain starts from a high primary production activated by the arrival of nutrients in the euphotic layer, ending as prey for tuna (micronecton, pelagic fish, etc…), then finally, tuna and man.
Among the parameters currently used to describe the environment and tuna distribution, temperature, and more specifically that of the surface, has been the object of numerous works to define thermal preferenda where different species of tuna are found. The surface temperature is a parameter taken routinely for a number of years (by fishing boats, merchant and oceanographic vessels, buoys, satellites, coastal stations) and from this there are reliable surface temperature maps over long periods of time. Other parameters that potentially influence tuna distribution and catchability should also however be taken into account: we will cite the thermocline depth, its temperature gradient, the concentration of dissolved oxygen, salinity, speed of the current, the state of the sky and the sea the speed of the wind, etc.. Furthermore, experiments done in the Pacific on tuna physiology permits the establishment of physiological limits in regard to certain environment parameters (chapter 6); the quest for food and reproductive behavior are also going to influence tuna distribution within the boundaries defined by these physical and physiological parameters (Blackburn, 1965 and 1965a; Sund et al., 1981; Albaret, 1977).
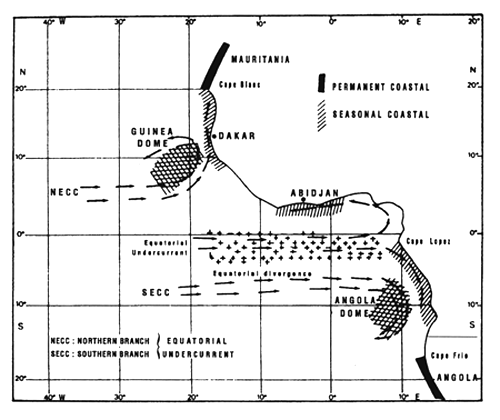
Figure 7.1 Various enrichment zones in the eastern tropical Atlantic. These are principally due to the vertical movement of deep water to the surface (upwelling) between July and September. These are localized along the coast and at the equator (Herbland et al., 1983).
The limiting values of environment parameters generally taken into account to study tuna distribution are evaluated mostly from fishing data. By using commercial fishing data, one describes only however incompletely the actual tuna distribution in the ocean, and rather their availability to different fishing gears; information remains null outside of space/times strata explored by the fisheries. In this chapter, we will analyze the principal zones and fishing seasons of tuna fleets in the eastern tropical Atlantic and will attempt to link them to principal productive systems known in the zone.
These productive systems, described by Voituriez and Herbland (1982) and presented in figure 7.1. are the following:
In their “Comparison of tropical Atlantic productive systems”, Voituriez and Herbland (1982) emphasized the fact that “these systems are not independent from one another: the counter current system, that participates dynamically with each of them and controls them to a certain extent, creates a remarkable link that gives this region a unity that one should keep in mind when one studies such an ecosystem and its variations”.
All of the statistical catch and effort data by fleet according to 1° - month strata that have been used in chapter 4 are used. These data cover the period 1969 – 1984.
For detailed analysis of spatio-temporal stratum of fishing, we will use the detailed data from long books of FIS and Spanish fleets (pole and line boats and purse seiners) for the period 1980 – 1983. These data are available by fishing day and with precise positions taken by satellite; furthermore temperatures measured during fishing operations are frequently available (for around 50% of catches), as well as size estimations of individuals caught. Log books used cover around 80 – 90% of the catches (depending on year) of FIS and Spanish fleets.
One must not loose sight of the fact that these data constitute visual estimations reported in the ships log books by tuna masters and thus present various limitations:
Thermal data used come from:
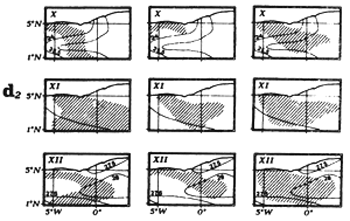
For this study, we have demarcated eight regions (figure 7.2), each corresponding to a particular hydrological structure and to a particular fishing zone.
The eight regions utilized present several types of hydrological situations and different productive systems:
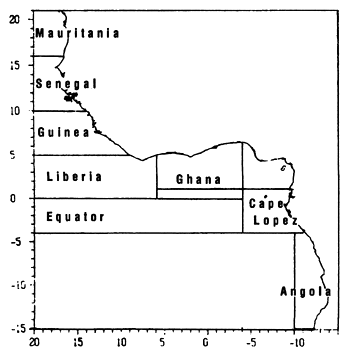
Figure 7.2 The 8 fishing zones used for the study of tuna-environment relationships.
We have fishing seasons during which catches are larger for each one of these regions, from FIS and Spanish fleet catches. Later, we will employ the term fishing zone and period of fishing, and not fishing concentration, in order to refer to these strata. This latter term has a much more restrictive sense and corresponds to a large aggregation of fish exploited in a short duration and in a zone of a few hundred square miles.
We have thus recognized the following fishing periods:
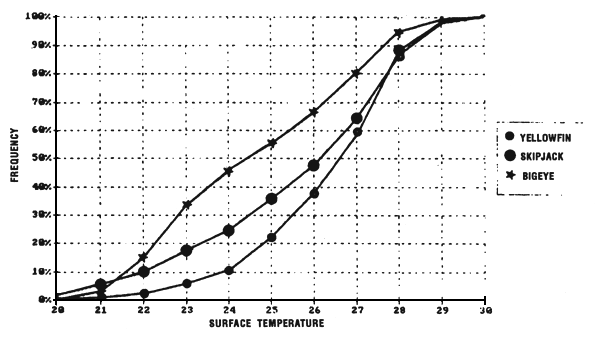
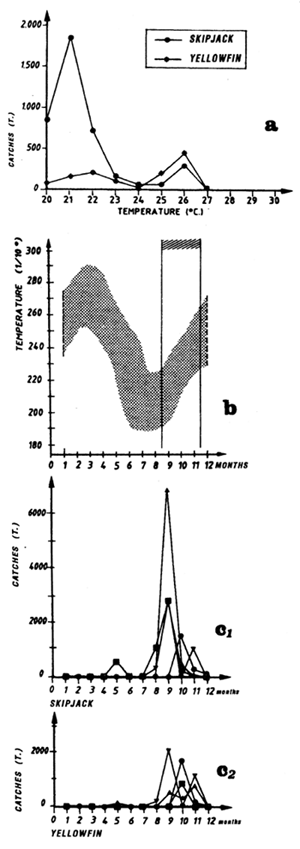
Figure 7.3 Cumulative percentages of yellowfin, skipjack and bigeye catches as a function of surface temperature (same sample as figures 7.3 to 7.5).
In the eastern tropical Atlantic, Postel (1955) fixed the thermal limits of yellowfin at 21° C and 28° C based on pole and line fishing between the Mauritanian coasts and the Bijagos archipelago. In the Pointe-Noire (Congo) region, Le Guen et al., (1965) note null or poor fishing in water less than 23° C or above 27° C. According to these authors, the majority of catches and the largest daily catch rates by pole and line boats and small purse seiners are obtained in waters where the surface temperature was between 24° C and 26° C. Bages and Fonteneau (1979) emphasize, for the year 1977, yellowfin fishing between 22° C and 29° C in the Gulf of Guinea by the FIS fleet. Stretta and Slepoukha (1986) define a temperature bracket of 20° C to 31°C for sets containing only yellowfin (all size classes mixed) made on schools not associated with logs. It is noteworthy that this interval of temperature between 20° C and 31° C corresponds approximately to the minimum and maximum values found in the eastern tropical Atlantic.
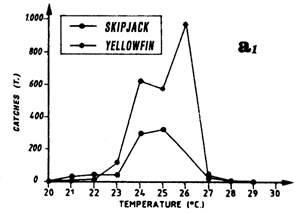
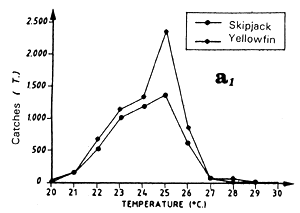
Only the sample of FIS and Spanish tuna boat catches from the period 1980 to 1983 will be analyzed, this being judged the most significant because of the amount caught by these two fleets and their complete geographic coverage of the zone. These two fleets together contribute, during the period studied, to 74% of industrial surface catches in the zone. Figure 7.3 presents the distribution of tonnages and the cumulative frequency of yellowfin caught (all sizes included) by temperature class by these purse seiner fleets. More than 60% of catches come from water with temperatures over 26.0° C.
Catch distribution of yellowfin as a function of temperature differ for the three classes classically differentiated (figure 7.4): fish less than 5 kg, those between 5 and 30 kg and those over 30 kg. For the first category, fishing is done essentially between 22° C and 30° C. For the second category (yellowfin from 5 to 30 kg), the catch curve presents a plateau of 24° C to 28° C. On the other hand, for yellowfin more than 30 kg, the largest part of the catches took place in waters where the surface temperature is over 25° C with 34% of the catches for the class 28° C.
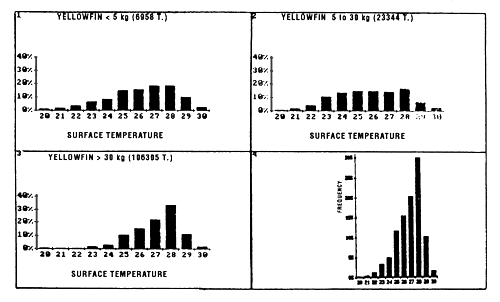
Figure 7.4 Catches of yellowfin as a function of surface temperature for FIS and Spanish purse seiners for the period 1980 to 1983 (subsample of sets for which the surface temperature and species composition are known). (1) yellowfin less than 5 kg; (2) yellowfin from 5 to 30 kg; (3) yellowfin greater than 30 kg; (4) all sizes.
Adult yellowfin are also frequently caught by longline below the thermocline where the temperatures are between 15°C and 20° C in the Gulf of Guinea.
In the Gulf of Guinea, Bages and Fonteneau (1979) find that skipjack is fished from 22° C to 29° C, is in the same thermal range as yellowfin. In an analysis on 1152 sets, containing only skipjack, Stretta and Slepoukha (1986), define a thermal range between 20° C and 31° C.
The sample used to describe the relation between surface temperatures and skipjack catches is the same as for yellowfin.
Skipjack distribution by size class as a function of surface temperature differs according to fish less than 2 kg, or between 2 and 6 kg and those over 6 kg (figure 7.5.). For fish less than 2 kg and those from 2 to 6 kg, the curves are close and both present a maximum of catches for the class 28° C. On the other hand, the catch distribution for skipjack over 6 kg presents a peak centered on the 25° C class which alone represents nearly 50% of the catches (the scarcity of catches of this category, 560 tonnes only, greatly limits however the significance of this result).
In the eastern tropical Atlantic, Pianet (1981) puts the thermal preference for bigeye between 20° C and 29° C and a thermal preference between 20° C and 22° C for bigeye fished by surface gear along the coast between Cape Lopez and Angola.
As for yellowfin, we have analyzed distribution of bigeye catches by purse seiners as a function of temperature for individuals less than 5 kg, for those between 5 and 30 kg, and for those more than 30 kg. For bigeye less than 5 kg (figure 7.6), one notes the existence of two modes centered on 23° C and 28° C, making up 60% of the catches between 26° C and 30° C; the very reduced size of the sample (445 tonnes), due essentially to very poor identification of small bigeye in log books, makes this result not very significant. For bigeye from 5 to 30 kg, the catch curve by temperature class presents approximately a plateau between 23° C and 29° C. On the other hand, for bigeye more than 30 kg, 62% of the tonnages fished, are at temperatures under 25° C.
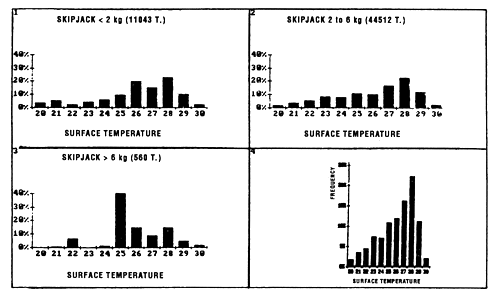
Figure 7.5 Catches of skipjack as a function of surface temperature for FIS and Spanish purse seiners for the period 1980 to 1983 (subsample of sets for which the surface temperature and species composition are known). (1) skipjack less than 2 kg; (2) skipjack from 2 to 6 kg; (3) skipjack greater than 6 kg; (4) all sizes.
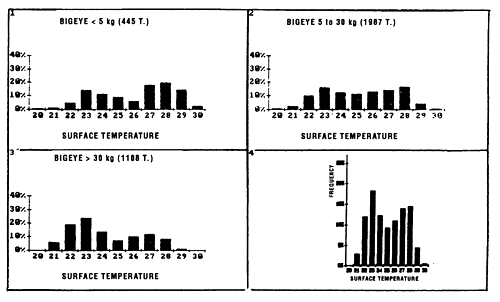
Figure 7.6 Catches of bigeye as a function of surface temperature for FIS and Spanish purse seiners for the period 1980 to 1983 (subsample of sets for which the surface temperature and species composition are known). (1) yellowfin less than 5 kg; (2) yellowfin from 5 to 30 kg; (3) yellowfin greater than 30 kg; (4) all sizes.
Adult bigeye are in fact mainly caught with longlines, particularly with deep longlines, in waters below the thermocline where temperatures are between 10° C and 15° C.
The dissolved oxygen content of the water mass plays an important role in the physiology of tuna as we have seen in paragraph 6.1.6. Sharp (1978) sets the minimum needs in oxygen to 1.5 ml/l for yellowfin and 0.5 ml/l for bigeye. Barkley et al., (1978) propose the value of 5 ppm (around 3.5 ml/l) of dissolved oxygen as the value to use to develop a model of skipjack distribution. These authors were thus able to draw skipjack distribution maps for the Pacific. In the Atlantic Ocean, Ingham et al., (1977) and Evans et al., (1979) have traced potential skipjack distribution maps from the maximum depth of the 18° C isotherm or of the 3.5 ml/l of dissolved oxygen isopleth. The latter authors find that skipjack is capable of living between 40° N and 34° S. These authors think that in the coastal upwelling zones, in the latitudes above 15°, skipjack habitat is conditioned by the depth of the 18° C isotherm, while in the equatorial band, between 15° N and 15° S, this habitat is instead conditioned by the concentration of dissolved oxygen. However, in the south-east region of the Atlantic off Angola, skipjack are generally caught in the third quarter between 30 and 50 meters in zones where the oxycline is very shallow (figure 3.13.).
In the Gulf of Guinea, outside of the Bay of Biafra (north-east of the Gulf of Guinea), the surface salinity varies from 32 to 36‰. It is not possible to characterize the tuna distribution zones by surface salinities, although the low salinity probably explains the low tuna catches in the zone north-east of the Gulf of Guinea.
Sea water salinity does not seem to have a direct effect on tuna (Blackburn, 1965; Sund et al., 1981). In the framework of studies done on skipjack and yellowfin in captivity, Dizon (1977), did not observe variations in swimming speed or in the behavior of these tuna when the salinity fell from 34 to 29 ‰. Staying in the range of values that one habitually finds in the natural environment, one can acknowledge that tuna were not sensitive to natural variations in salinity. However, Donguy et al. (1977) note very clear relations between the size of skipjack catches and salinity in the western Pacific. In examples presented by these authors, the surface salinity is linked to particular oceanographic conditions such as equatorial upwelling due to easterly winds and the phenomenon of “doming” toward 10° S by the west wind. In these two cases, Donguy et al. (1977), discover the presence of a superficial enrichment. The surface salinity is, in this case, only one of the principal indicators of these mechanisms of enrichment and not the parameter acting directly on the behavior of skipjack.
In the Gulf of Guinea, Caverivière et al. (1976) demonstrate for yellowfin a relation between salinity and abundance of larvae or spawning: they have never observed larvae in waters where salinity was over 33.5‰. On the other hand, for bigeye and skipjack, larvae have been found in waters at 31‰.
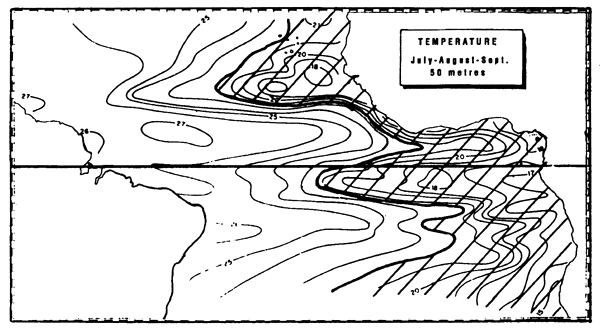
Figure 7.7 Zone of the superficial thermocline in the intertropical Atlantic, defined by the 22° isotherm at a depth above 50 m (in the third quarter, after Merle, 1978).
In this paragraph, we will not deal with the vertical distribution of tuna approached in chapter 6. The term thermocline used by fishermen is related more to the depth of the homogeneous surface layer or “depth of the thermocline”, than to the vertical temperature gradient found below this homogeneous layer.
Green (1967) analyzed the rate of positive sets as a function of the depth of the thermocline. From his study, based on 1855 purse seine sets in the eastern tropical Pacific, he concluded that the rate of positive sets increases when the upper mixed layer is shallow. For this author, the best rates of positive sets are found when the thermocline is less than 18.4 m and the temperature gradient in the thermocline is over 0.55° C/m. This author thinks that these higher rates of positive sets are explained by the low content of dissolved oxygen (< 1 ml/l) observed just under the thermocline. Blackburn and Williams (1975) find that in the eastern tropical Pacific, the maximum in skipjack abundance is in zones where the homogeneous layer is less than 40 meters. Sharp (1978) studied the variations of the vulnerability of yellowfin fished by purse seine as a function of time of the day, depth of the homogeneous layer, and depth of the 23° C and 15° C isotherms during a period of 6 months. The choice of these isotherms was been made according to physiological criteria. For tuna fishing not associated with schools of dolphins, this author finds in the eastern tropical Pacific a strong correlation (r = 0.94) between catch per unit of effort and the depth of the 23° C isotherm; he concludes that the maximum vulnerability of yellowfin in this region appears in zones where the 23° C isotherm is at a depth greater than 45 meters and where the 15° C isotherm is above 73 meters. Note that in the western tropical Pacific, purse seiners fish in waters where the thermocline is at a depth over 150 m. Evans et al., (1980) show as well that in the eastern tropical Pacific, the number of positive sets containing skipjack is maximum when the homogeneous layer is at 15 meters.
No detailed analysis of this problem has been done in the study sector, however, one notes that, taken as a whole, the fishing zone of tropical tuna by surface gears in the zone coincides with the one where the thermocline is close to the surface. This is clearly shown in figure 7.7.
Acoustic telemetry experiments show that a school of yellowfin swims mainly between the surface and the base of the thermocline while an isolated individual has a tendency to head toward the thermocline (Carey and Olson, 1981). In the Atlantic, by using an ultrasonic tag, Levenez (1981) shows that a skipjack can make numerous large vertical movements and cross the thermocline; he even observes a skipjack passing rapidly from a temperature of 28° C at the surface to a temperature under 11° C at 400 meters. At this depth, the oxygen content was probably under 1.5 ml/l. Skipjack dive therefore into cold waters for brief but repeated excursions in order to permit direct dissipation of heat produced by muscles into the environment (paragraph 6.1.5.).
Before describing and analyzing the different productive systems, we will study the productivity of oligotrophic waters preceding the establishment of the productive system. Oligotrophic water is water depleted of nitrates and in which there is no new production; only production by regeneration exists.
It is historically acknowledged that the tropical Atlantic is mildly productive, with the exception of zones where the environmental conditions bring an abundant quantity of nutrients to the surface (Herbland et al., 1983). In the eastern tropical Atlantic during the warm season, Herbland and Voituriez (1977) have defined a Typical Tropical Structure (TTS), that is, a two-layer system where the surface layer is lacking in nitrate. We will borrow from Herbland et al. (1983) the description of the typical tropical structure: “Under a warm homogeneous layer, containing no nitrate, little chlorophyll and saturated in oxygen, the nitracline always coincides with the chlorophyll maximum and the oxycline. Its level is over or under the maximum temperature gradient depending on the depth and especially the intensity of the latter”. These authors specify that “the depth of the nitracline will then be an indicator of the intensity of the flux of nitrate in the euphotic zone and that the depth of the nitracline then becomes the conductor that permits the linking of the most productive domes to the deepest convergences”. This typical tropical structure ceases to exist when nitrate appears on the surface. In the tropical zone, it is generally at the base of the homogeneous layer that primary production is strongest, where the appearance of nutrient salts in the euphotic layer stimulates new production. However, the functioning of an oligotrophic ecosystem is still largely an enigma (Herbland, pers. com.).
The intensity of the new production derived from deep water nitrates defines the richness of a zone in phytoplankton, since production from regeneration consumes nitrogenous organic compounds, urea and amino acids, that have already cost the ecosystem energy to produce.
Brandhorst (1958) points out that one of the most important factors influencing the aggregation of pelagic fish in tropical waters is the presence of food dependent on phytoplankton production; in the eastern tropical Pacific, this author finds an inverse relation between the depth of the thermocline and the quantity of zooplankton, and this quantity of zooplankton appeared to be linked, in some regions, to tuna abundance.
In the eastern tropical Atlantic, Le Borgne et al. (1983) also find that in typical tropical structures, the biomass of zooplankton is all the more elevated since the upper mixed layer is thin, ie. since the thermocline is closer to the surface. By comparison, in ohgotrophic waters where this layer is thick, the biomass of zooplankton is low.
If one considers the macroplankton-micronecton (animals from around 0.5 to 10 cm), Roger (1982) finds in oligotrophic waters, a low biomass of these animals with 311 mg/m2 in dry weight against 1165 mg/m2 in the richest zone in the tropical Atlantic (from this author, 1 g of dry weight per m2 of macroplankton-micronecton represents 5 tonnes of fresh biomass per km2 in annual average). Taxonomically, crustaceans represent 57% of the total micronecton biomass and fish 29%. In the oligotrophic zones, 47% of the biomass is concentrated in the first 100 meters during the night against 7.1% during the day.
Table 7.1 Table comparing the mean values of water column primary production expressed in mgC/m2/h encountered in different productive systems of the Gulf of Guinea.
| Maurit. upwel. | C. Lopez Front | Guin. & Ang. Domes | Equator | Convergence | ||
| Therm. R. | upwel. | |||||
| Water column Prim. Prod. | ||||||
| Mean values mgC / m2 / h | 137 – 200 (1) | 73 (2) | 80 (1) | 70 (3) | 133 (4) | 38 (3) |
(1) VOITURIEZ and HERBLAND (1982)
(2) Value calculated by VOITURIEZ and HERBLAND (1977) according to DUFOUR and STRETTA 1973
(3) VOITURIEZ and HERBLAND (1977)
(4) VOITURIEZ and HERBLAND (1978)
In the tropical Atlantic, there are different processes for enrichment of water masses. In table 7.1., we have combined the maximum primary production rates observed in the eastern tropical Atlantic within different productive systems.
These enrichment zones have a common point: the rising of the thermocline in the euphotic layer. Zones of nutrient upwelling in the euphotic layer are the center of the fertilization of the water mass (Margalef, 1978). In artificial fertilization experiments of Mediterranean oligotrophic waters with nitrates and phosphates, Fiala et al.(1976) find that the maximum development of phytoplankton takes place after 6 to 10 days.
In the eastern tropical Atlantic, the intensive development of phytoplankton during the enrichment in nitrates of the euphotic layer will be limited by intensive grazing by herbivores. The positive linear relation between chlorophyll and dry weight of zooplankton found by Le Borgne et al. (1983), expresses the coupling between zooplankton and phytoplankton biomass. This relation is as applicable in oligotrophic water as in equatorial upwelling. Le Borgne et al. (1983) note that with increase in primary production, there is not only an increase in phytoplankton biomass but also an increase in size of cells. Moreover, Jones (1982) notes also that the largest individuals at each trophic level have a tendency to grow with increase in primary production and that this tendency appears at high levels of the food chain. Also, fish of commercial size can feed at progressively lower levels of the food chain. With an increase in primary production, the size of the largest individuals at each trophic level grows, and the food chain tends to be relatively short (Jones, 1982). Also this author points out the fact that in a surface upwelling zone, the horizontal transport of the water mass is an important factor as it must last for a sufficiently long time in order to permit the development of the food chain.
Le Borgne et al. (1983) also mention the “possibility (for micronecton) during an increase in phytoplanktonic biomass to augment the algal component of their ration to the detriment to other particles: it could go straight to a herbivorous diet from an omnivorous diet”. This variation in diet can be also be accompanied by a short circuit in the food chain. Jones (1982) suggests this for the food chain ending in tuna in the Gulf of Thailand. It is thus that the food chain beginning at phytoplankton and ending at tuna can take two “paths” (figure 7.8). The first one, the longest, goes in two stages to end up at tuna: first herbivores in the widest sense, then through fish larvae and juveniles and/or invertebrate predators. The second path goes in only one stage to end up at tuna: sardinellas, anchovies, mackerel (small coastal pelagics) and micronecton.
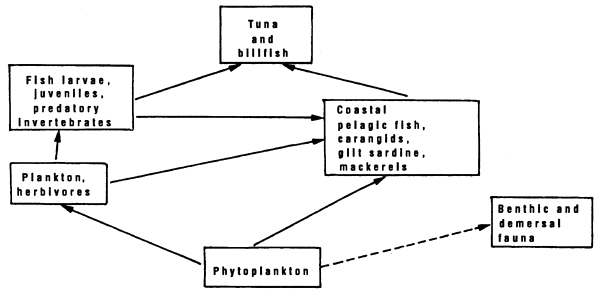
Figure 7.8 Diagram of the food chain leading to tunas in the gulf of Thailand; after Jones, 1981.
In upwelling zones, the largest herbivores are fish. This form of food chain short circuit is described for tuna off Baja California (Griffiths, 1965 and Blackburn, 1969a). In the frontal zone, yellowfin and skipjack feed mainly on one species: the red crab Pleuroncodes planipes; this pelagic crab is a herbivore.
These patterns are valid for coastal upwelling zones and frontal zones associated with them in the Gulf of Guinea. In the absence of frontal structures stemming from a cold surface upwelling, the maturation processes of the water mass are slower. However, Herbland et al. (1983) specify “it is not necessary that the nutrients reach the surface for the zone to be productive and the mixed layer the centre of great activity”. There is then the question of knowing whether this productivity is sufficient to support tertiary production capable, in turn, to support tuna concentrations. Herbland et al. (1983) estimates “that the short term variations of hydrological conditions have probably more importance than one thinks.”
Independent of the complexity of productivity mechanisms that come into play, it is striking to note the strong correlation that exists between zones of large planktonic production (figure 7.9) and those of large tuna catches (figure 4.11).
In the Gulf of Guinea, Dragovich (1970) describes the trophic chain leading to tuna by analyzing the stomach contents of fish ingested by yellowfin and skipjack. The results of this study confirm the dependence of these prey organisms below macrozooplankton. From Roger and Grandperrin (1976), in the western tropical Pacific, the migrant fauna that lives at depth during the day and only comes to the surface layers in the night, seems to not participate in the diet of tuna, which feed essentially at the expense of organisms that remain in the upper 200 meters during the day.
Paragraph 6.1.4. analyses the diet and feeding behavior of tuna. We recall in summary that:
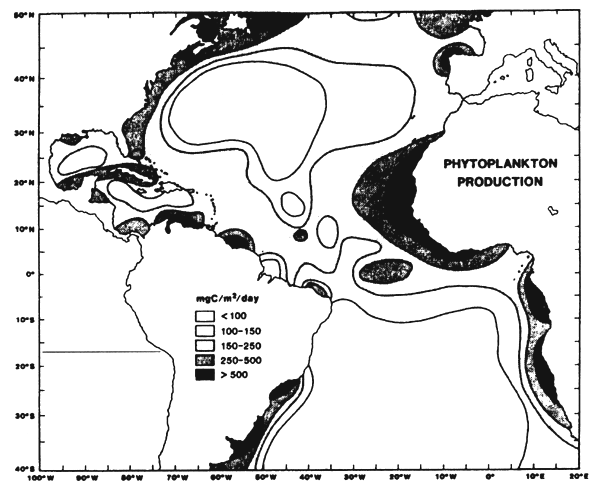
Figure 7.9 Mean phytoplankton production in the Atlantic (after Koblentz-Mishke et al., 1970; FAO, 1972).
Sund et al. (1981) estimate that tuna larvae are found in zones where environmental conditions are compatible with spawning and the survival of larvae and where the drift (or transport) of larvae carries them in zones where physical parameters and food conditions are optimum. We have seen in paragraph 6.1.1.6. that in the Gulf of Guinea, Caverivière et al. (1976), demonstrate that yellowfin larvae are especially abundant in warm waters with a temperature over 28° C. The presence of warm waters is also an indispensable element for sexual maturation and spawning of skipjack.
Sund et al. (1981) ask the classic and important question of knowing if migration or the movement of tuna toward spawning zones is “active”, “passive” or the result of chance. In the Gulf of Guinea, although information on yellowfin and skipjack reproduction are fragmentary, one can reasonably think that genetic migrations of these two species are at the same time active and passive; this is the conclusion adopted by Williams (1972) for skipjack in the Pacific.
The five environment parameters that we mention: surface temperature, content of dissolved oxygen, depth of the thermocline, its associated temperature gradient, and available food, must be considered together as it is the environment parameter that will be close to the critical value for a species, at a size and at a given physiological level, that will become the limiting factor in the distribution of tunas.
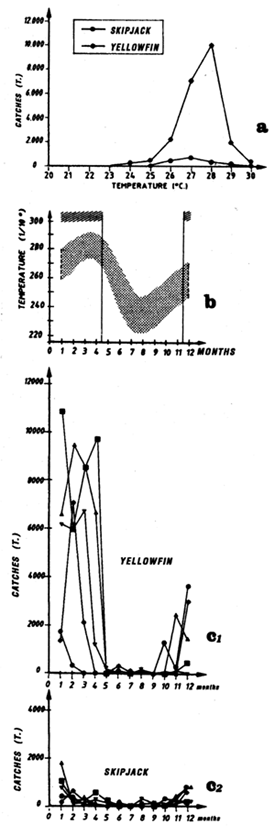
For each spatio-temporal stratum, we will present five types of graphs:
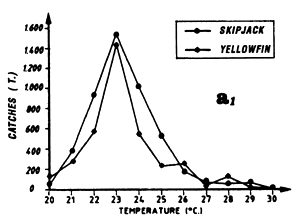
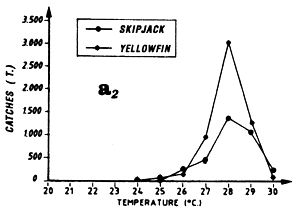
The first (a) presents catch in tonnes as a function of surface temperature for yellowfin, skipjack and bigeye for FIS and Spanish purse seiners from 1980 to 1983 (from catch samples for which surface temperature at time and place of the catch is known).
The second (b) presents average monthly surface temperatures (in 1/10 of a degree) from 1964 to 1984 in each one of the sectors studied. These data are from merchant vessel files (NODC, USA). On this graph, the vertical lines enclose the fishing period or periods that have been retained for analysis.
The third type of graph (c) represent catch by month in the fishing zone examined by FIS and Spanish tuna boats from 1979 to 1983.
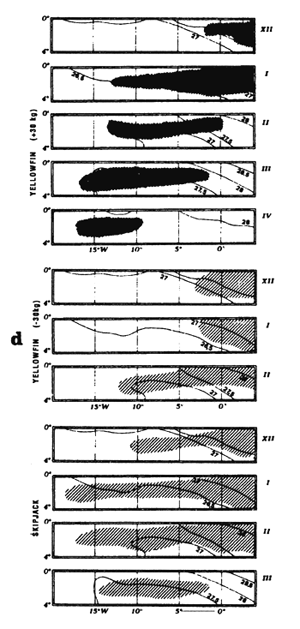
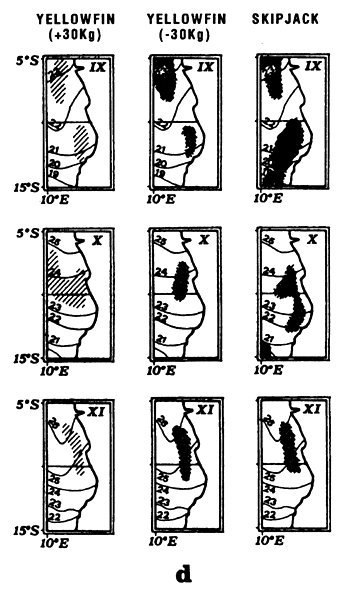
The fourth type of graph (d) consists of monthly fishing maps with precise localization of purse seine sets for large yellowfin, small yellowfin and skipjack by FIS and Spanish purse seiners from 1980 to 1983. On the maps representing yellowfin catches, we have taken the positions of the monthly isotherms taken from the atlas of Hastenrath and Lamb (1977). This type of representation gives an image of average surface temperatures found in the fishing zone. One must not loose sight however that these temperatures are not concomitant to fishing data and only constitute an average statistical reality of surface temperatures in the sector.
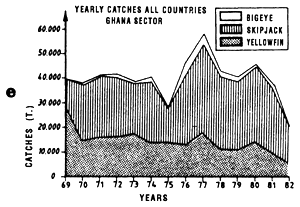
The fifth graph (e) presents an estimation of total catches by all surface gears for the three species of tuna from 1969 to 1982 in each of the geographic sectors studied.
The hydrology of this zone has been mostly described during boreal winter at the moment where coastal upwelling, one of the most important ocean wide, develops.
The Mauritania zone is historically one of the first fishing zones explored by European tuna fleets. The fishing season extends from June to October; it starts at the end of the cold season and continues during the entire warm season (figure 7.10.b.). One finds the phenomenon of hydrological transition in the distribution of tuna catches as a function of temperature, with catches of yellowfin, skipjack and bigeye in waters between 22° C and 28° C (figure 7.10.a.). Maps of catch localization (figure 7.10.d.) show that tuna are caught to the limit of the continental shelf to the south-west of the Arguin shoal, in immediate proximity to a zone which is very rich in coastal pelagic fish. The fishing zones are very restricted and it is only from the month of October that tuna disperse toward the south.
The hydrology of the Senegal-Mauritania maritime province is described by Berrit (1962), Rossignol and Aboussouan (1963), Champagnat and Domain (1978) and Rebert (1979). This region is situated in the zone of oscillation of the intertropical front to the north where a system of tradewinds is established. The influence of these winds is felt up to 10° N in winter and up to 21° N in summer. A resulting cold season appears from December to May and a warm season starting in June. In the cold season, the winds intensify the seasonal cooling on the coast and this particularly strong upwelling brings cold waters to the surface, on average less than 22° C, with temperatures locally on the low side of 18° C; these cold waters are rich in nutrients. In the warm season, the trade winds weaken, the upwellings cease, and the cold waters are covered with warm waters, more than 27° C, associated with the north equatorial counter current.
Surface temperature maps show the presence of a frontal structure from March to June off the Bijagos Islands between 8° N and 13° N. This zone symmetrical to the one of Cape Lopez in the southern hemisphere. Although the Bijagos front has not been studied as well the one of Cape Lopez and is probably less well characterized, one can reasonably think that the hydrological and enrichment phenomena are similar but however less important.
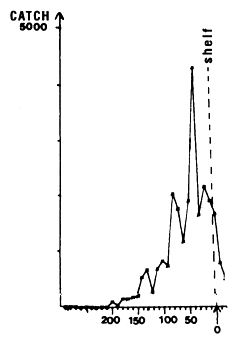
Figure 7.12 Longitudinal distribution of skipjack catches by 10 mile intervals in the zone 14 to 15° N during the month of August, 1980 to 1983. The edge of the continental shelf is situated approximately at the point marked 0 along the x axis.
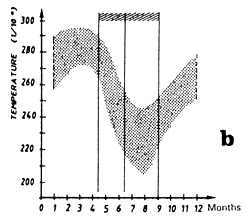
On can distinguish two fishing seasons in this zone: the first in April and May, the second from July to September. These two fishing periods are very clearly differentiated at the catch level as a function of the surface temperature:
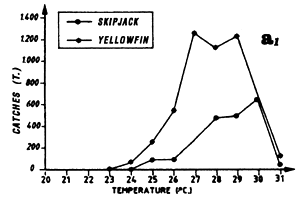
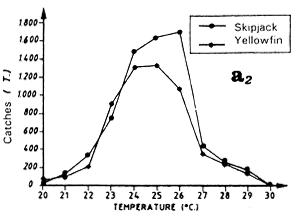
During the fishing season that extends from April to May, the period that corresponds to the end of the cold season and the beginning of the warm season, one finds the transition in the tuna catch distribution as a function of temperature, with catches of yellowfin and skipjack in waters over or equal to 25° C and catches in temperatures under 25° C (figure 7.11.a.) The maximum of catches is observed at 23° C. From Cayré (1985), skipjack that sustain catches observed off Senegal and the Cape Verde Islands in April-May, come from a zone situated off Liberia. These skipjack stay in the Senegal zone all summer.
The second fishing period, from July to September, is the most important for catches. The yellowfin and skipjack fishing occurs in warm waters where the temperature is between 26° C and 30° C (figure 7.11.a.). Note that almost all of the catches are made in a very narrow fringe, just above the 200 meter isobath and at the immediate proximity of the continental shelf. We have shown in figure 7.12 the total skipjack catches for the months of August from 1980 to 1983, by FIS and Spanish purse seiners, between 13° N and 15° N as a function of longitude, that is to say, approximately the distance to the coast. The catch peak is between 17° 45' and 17° 55' west. In order to explain this extreme tuna concentration, one must envisage the hypothesis that these fish feed actively on important coastal pelagic stocks that stay the summer in the sector between two upwelling seasons. The estival biomass of coastal pelagic fish in the sector is estimated by echo-integration to be on the average several hundreds of thousands of tonnes (Levenez pers. com.). This hypothesis must be confirmed by stomach contents observations.
Figure 7.10 Catches and temperature in the “Mauritania” sector:
Distribution of yellowfin and skipjack according to surface temperature, FIS baitboats, 1980 to 1983, fishing operations for which surface temperature is known, from May to November.
Mean monthly surface temperature (in tenths of degrees) taken by merchant vessels in the sector from 1964 to 1984 (the period corresponding to the fishing season is indicated by dashes).
Catches of bigeye (c1), yellowfin (c2) and skipjack (c3) per month by FIS baitboats from 1969 to 1984.
Locations of monthly catches of yellowfin, skipjack and bigeye in the sector from May to November.
Annual catches of yellowfin, skipjack and bigeye estimated for all surface tuna fleets, 1969 to 1982 (in metric tonnes).
MAURITANIA
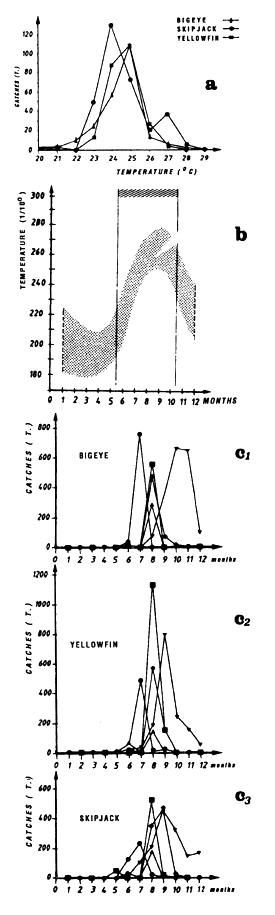 | 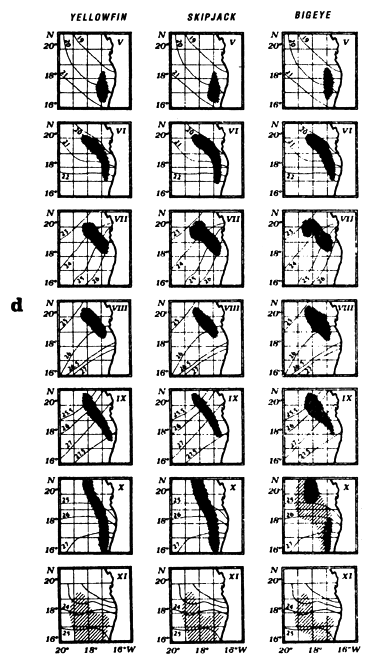 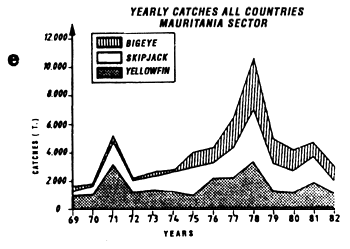 |
Figure 7.11 Catches and temperature in the “Senegal” sector:
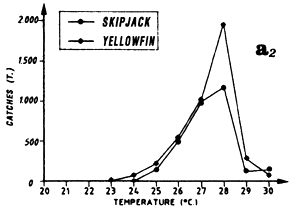
Distribution of yellowfin and skipjack according to surface temperature, FIS and Spanish purse seiners, 1980 to 1983, hauls for which surface temperature is known, (a1) from April to May and (a2) from July to September.
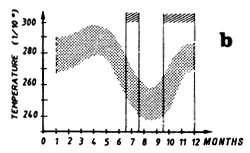
Mean monthly surface temperature (in tenths of degrees) taken by merchant vessels in the sector from 1964 to 1984 (the period corresponding to the fishing season is indicated by dashes).
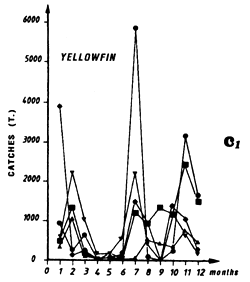
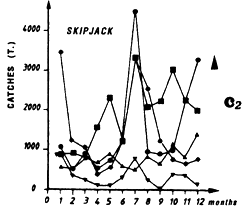
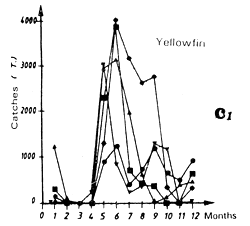
Catches of yellowfin (c2) and skipjack (c1) per month by FIS and Spanish purse seiners from 1969 to 1984.
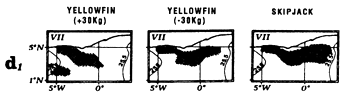
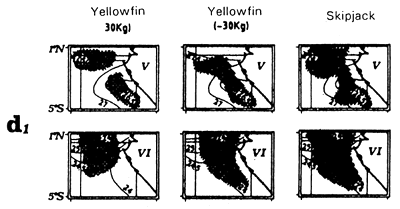
Locations of monthly catches of large yellowfin (+30kg) yellowfin, small yellowfin (-30kg) and skipjack in the sector from April to May (d1) and from July to September (d2).
Annual catches of yellowfin, skipjack and bigeye estimated for all surface tuna fleets, 1969 to 1982 (in metric tonnes).
SEGENAL
 |  |
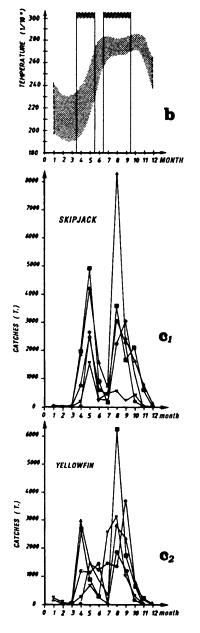 | 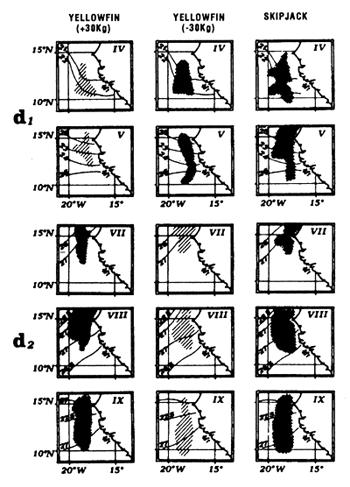 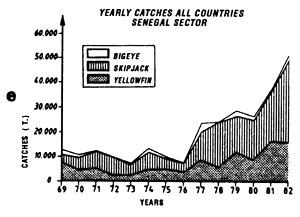 |
Figure 7.13 Catches and temperature in the “Guinea” sector:
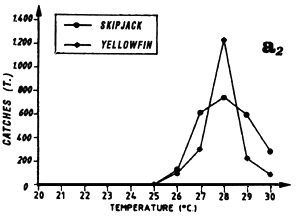
Distribution of yellowfin and skipjack according to surface temperature, FIS and Spanish purse seiners, 1980 to 1983, hauls for which surface temperature is known, (a1) from March to May and (a2) from September to November.
Mean monthly surface temperature (in tenths of degrees) taken by merchant vessels in the sector from 1964 to 1984 (the period corresponding to the fishing season is indicated by dashes).
Catches of yellowfin (c1) and skipjack (c2) per month by FIS and Spanish purse seiners from 1969 to 1984.
Locations of monthly catches of large yellowfin (+30kg) yellowfin, small yellowfin (-30kg) and skipjack in the sector from April to May (d1) and from July to September (d2).
Annual catches of yellowfin, skipjack and bigeye estimated for all surface tuna fleets, 1969 to 1982 (in metric tonnes).
GUINEA
 |  |
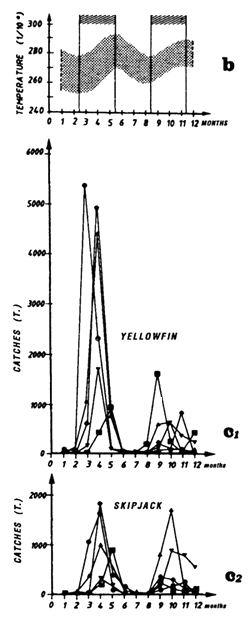 | 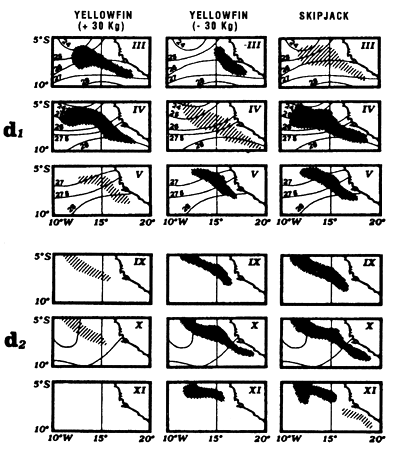 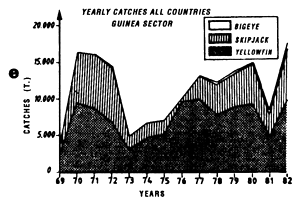 |
Table 7.2 Table comparing the mean values of zooplankton production expressed in mg encountered in different productive systems of the Gulf of Guinea.
| Angola Dome | Oligot. waters | N. E. C. | Equator | Convergence | ||
| Therm. R. | upwel. | |||||
| ZOOPL. PROD. (micro.+meso.) dry weight mg | 525 | 139 – 360 | 683 | 394*-1 318 | 592-2 360 | 570-994 |
This region is a boundary zone between the northern regions subject to the climatic regime of the northern hemisphere and the southern regions under the climatic regime of the austral hemisphere. The annual amplitude of surface temperature variations is low, on the order of 2° to 3° C (figure 7.13.b.). An upward movement of the thermocline in boreal summer associated with the cyclonic circulation of the eastern extremity of the north equatorial counter current, is expressed in the thermocline in the form of a dome. This hydrological structure has been studied by several authors: Rossignol and Meyrius (1964), Mazeika (1967), Voituriez and Dandonneau (1974) as well as Voituriez and Herbland (1982). Voituriez and Dandonneau (1974) show that “this particular structure of the thermocline, which does not reach the surface, brings about an important increase in primary production, which one would think would be favorable to the development of the trophic chain leading to the presence of food for tuna”.
Two fishing seasons have been distinguished in this region: the first from March to May and the second of less importance, from September to November.
Large yellowfin and skipjack catches from March to May can be linked to the fact that this region is situated at the edge of the Senegal-Mauritania upwelling and the front with which it is associated (Berrit, 1962). It is probable that this frontal structure and advection of waters stemming from coastal upwelling contribute to the development of food favorable to the presence of tuna at the beginning of the year. The catch distribution as a function of temperature is a reflection of the hydrological complexity of this zone at this time of year. Yellowfin and skipjack catches as a function of temperature (figure 7.13.a.) are made in a wide temperature range (25° C to 30°C). Moreover, these large catches can also be the result of concentrations linked to yellowfin and skipjack reproduction. Albaret (1977), demonstrates a spawning area for yellowfin (less important however than the one located in the inner of the Gulf of Guinea) off Guinea during the first quarter of the year.
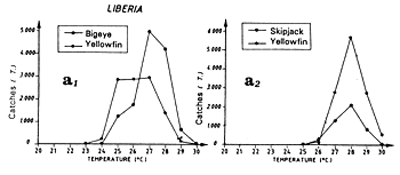
The second fishing season starts in this region at the end of boreal summer, at a time where the dome structure of the thermocline is present but abated due to subsidence of the north equatorial counter current. Catch distribution as a function of surface temperature reflects the spatial homogeneity of temperatures at this time. More than 80% of catches are taken in a thermal range from 27°C to 29° C (figure 7.13.a.). As for the Senegal and Mauritania zones, one must envisage the hypothesis that tuna come to feed at the edge of the continental shelf at the expense of the large coastal pelagic fish stock present off the Bijagos Islands. One must also consider the complementary hypothesis that tuna feed in the sector on the pelagic production stemming from the enrichment of the Guinea Dome. Another hypothesis, which should not be dismissed, is that of fishing associated with fish aggregation gears; Stretta and Slepoukha (1986) point out that in the coastal zones off Liberia, the proportion of fishing on logs in relation to total fishing in the zone is over 40% (paragraph 6.1.7.).
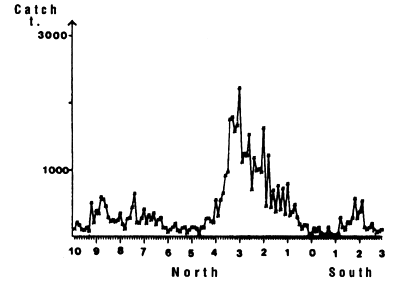
Figure 7.15 Latitudinal distribution of skipjack catches by 10 mile intervals in the zone 25°W to 10°W from 10°N to 3°S from November to March, 1980 to 1983, FIS and Spanish purse seiners.
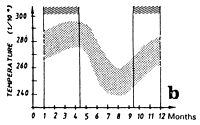
The Liberia zone is a complex zone for which few hydrological studies have been made. It is a zone where the amplitude of the annual thermal signal is large (figure 7.14.b). The cooling of waters in this zone during boreal summer is linked to the coastal cooling to the east of this zone in the Ivorian maritime province and to cooling coupled to the equatorial upwelling (Grouzov, 1971). This zone is a convergence zone between the north equatorial counter current and the north part of the south equatorial current. Table 7.2, presenting different values of micro and mesozooplankton production, shows that within this convergence zone this productivity is not negligible.
Two fishing seasons in this zone have been distinguished. The first from January to April and the second from October to December. In this zone, skipjack have a tendency to congregate between 2 and 4°N (figure 7.14.d. and 7.15). As for the Guinea region, yellowfin and skipjack catches during the first season are made over a wide thermal range (figure 7.14.a1). On the other hand, the thermal range in which the second fishing season takes place is much more restrained from 27 to 29°C (figure 7.14.a2.). Tuna abundance in the sector and their strong concentration between the latitudes 2°N and 4°N remain poorly explained, although this distribution is probably in relation to environmental factors.
The equatorial region of the Atlantic is characterized by the presence of two mechanisms for enrichment of the water mass. The first occurs in January-February and the largest values of chlorophyll are between 1°30'S and 3°S. This high productivity is associated with the presence of the thermal ridge observed between 2 and 3°S. A thermal ridge is a rising of the thermocline in the euphotic layer, the thermocline does not however reach the surface. This ridge is a permanent characteristic of the thermal structure in the equatorial zone of the Atlantic Ocean. It has been observed at 4°W in November 1971 (Dufour and Stretta, 1973a), at 10°W in November 1973 (Stretta, 1975) and up to 20°W (Voituriez and Herbland, 1977). The last authors remark that in this type of structure, maximum chlorophyll is observed at the nitracline which does not necessarily coincide with the pycnocline. If the nitracline is not too deep, there is a maximum of primary production that can reach 70 mgC/m2 /h (table 7.1). The presence of this thermal ridge near 2 to 3°S, will create a permanent source of enrichment and ecosystem variations will occur following a state of equilibrium. Dandonneau (1975) notes that in this type of ecosystem, the contributions of nutrient salts are sufficiently slow that the two populations of phytoplankton and zooplankton have the time to adapt. The effect of this thermal ridge is particularly clear in examining figure 7.16 which represents yellowfin catches in the sector as a function of latitude, by intervals of 10 nautical miles. The maximum of catches is between 1°30'S and 3°S.
Figure 7.14 Catches and temperature in the “Liberia” sector:
Distribution of yellowfin and skipjack according to surface temperature, FIS and Spanish seiners; 1980 to 1983, hauls for which surface temperature is known, (a1) from October to December and (a2) from January to April.
Mean monthly surface temperature (in tenths of degrees) taken by merchant vessels in the sector from 1964 to 1984 (the period corresponding to the fishing season is indicated by dashes).
Catches of yellowfin (c1) and skipjack (c2) per month by FIS and Spanish seiners from 1969 to 1984.
Locations of monthly catches of large yellowfin (+30kg) yellowfin, small yellowfin (-30kg) and skipjack in the sector from October to December (d1) and from January to April (d2).
Annual catches of yellowfin, skipjack and bigeye estimated for all surface tuna fleets, 1969 to 1982 (in metric tonnes).
 |  |
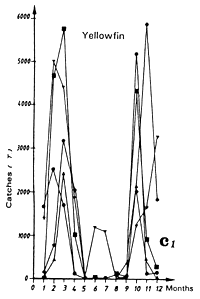 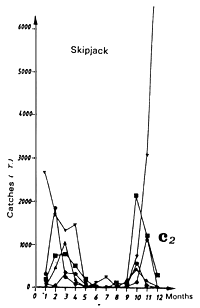 | 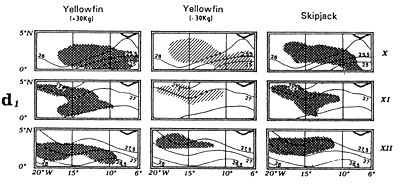 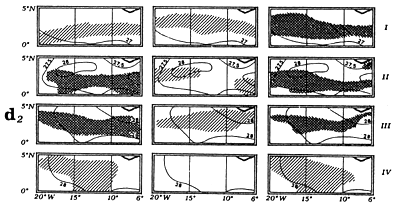 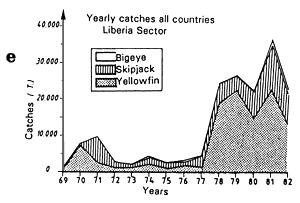 |
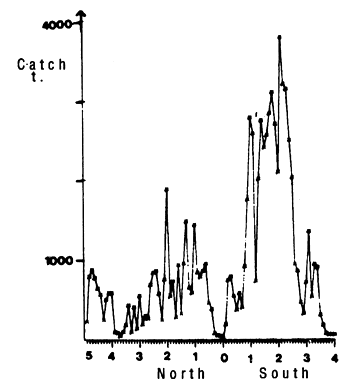
Figure 7.16 Latitudinal distribution of yellowfin catches by lo mile intervals in the zone 10°W to the equator, between 5°N and 4°S, from November to March, 1980 to 1983, FIS and Spanish purse seiners.
Although the thermal ridge seems to be a permanent feature of the thermal structure in the Atlantic during the warm season, the second enrichment mechanism, on the other hand, corresponds to upwelling that develops in the cold season in the equatorial region from July to September (or equatorial divergence). According to Voituriez and Herbland (1977), this upwelling is characterized by cooling and enrichment in nitrates between 0°30'N and 1°30'S, centered on 0°30'S. The bulk of production is in the surface. The cold waters resulting from this divergence encounter warm waters present to the north and form a frontal zone (that does not reach the extent of the one off Cape Lopez). In waters stemming from upwelling, productivity is elevated (table 7.1) with large variations linked to the strong turbulence that prevails at the equator. According to Voituriez and Hisard (1975), this turbulence is contrary to phytoplankton development.
Le Borgne (1977) has shown that the production of zooplankton is higher in the cold season (July-September) than in the warm season (December-March), the catch rate of production between the two seasons being very close to that of the biomass. This author estimates the average annual productivity of the equatorial zone at 21.5 tonnes of fresh matter per square mile.
The Equatorial zone could be qualified as being a region where only yellowfin is fished (figure 7.17.e). Fishing starts in this zone after the upwelling period in high productivity waters of the equatorial divergence. The weekly sea surface temperature maps furnished by the NOAA (GOSSTCOMP maps) demonstrate, in this region, the presence of important cells of cold water upwelling that create very localized enrichments of the water mass.
The predominance of yellowfin of more than 30 kg (figure 7.17.d) is linked to the fact that this zone seems to be one of the principal areas of yellowfin reproduction (paragraph 6.1.1.). According to Albaret (1977), the maturation of female yellowfin ovaries starts at the beginning of December and lasts until March and spawning occurs during the first quarter. Most of the spawning takes place in the eastern tropical Atlantic in two zones: the principal, which is in the “inner” Gulf of Guinea from 5° N to 5° S and extends from the African coast to the Greenwich Meridian (figure 6.9). A second spawning zone, is found off Liberia. The gonads of tuna caught in the fishing zone to the west of the Greenwich Meridian and to the south of the equator have never been sampled, but one can hypothesize that these individuals are also in the phase of reproduction during the first quarter. Migrating yellowfin, with out doubt associated to their reproductive cycle, thus find favorable spawning conditions.
The almost complete absence of yellowfin fishing in the zone from May to November remains difficult to explain as the waters of this sector are productive all year and the water temperature is compatible with their presence.
Yellowfin catches are principally observed between 26 and 29° C (figure 7.17.a.) and there is also a distinct displacement toward the west of the fishing locations between December and April (figure 7.17.d.). This displacement is made in all of the thermal ridge zone between the equator and 3° S; this movement can not be explained by search for food, the entire zone being under the influence of the equatorial enrichment process.
The Ghana Zone extends from the Côte d'Ivoire to Ghana and includes the continental shelves of these two countries. The hydrology of the Ivorian continental shelf is described by several authors: Varlet (1958), Morlière (1970), Morlière and Rebert (1972), Lemasson and Rebert (1973) and Binet (1976). From these articles, it is concluded that warm (28° C), desalinated (S < 35‰) tropical waters cover the central south Atlantic during most of the year.
Enrichment of this coastal zone comes from coastal upwellings that occur sporadically in January and February determining a “small cold season” during the “great warm season”. A more intense upwelling, for which the release mechanisms are analyzed by Picaut (1983), appears from July to September. This upwelling brings cold (22° C), salinated water (S> 35‰), rich in nutrients, to the surface layer. Binet (1976) shows that the primary production responds to environment enrichment brought about by upwelling and that the major part of the secondary production is linked to temperature variations. The nutrient contributions from river origin seem quite important in this zone.
The Ghana zone such as is represented in figure 7.2. includes the coastal fringe of the Ivory-Ghanaen continental shelf in the Cape Three Points region and the maritime province of the high seas up to 1° N between 5° E and 6o W. In this region during boreal summer near 3° N, a transition zone forms between the cold waters stemming from the coastal upwelling and the cold waters stemming from the equatorial divergence. This transition zone where a mixing takes place between warm tropical waters, defined above, and cold waters originating from the north and south, can be observed using airborne radiometers. This transitional water mass, of a temperature between 24.5 and 26.5° C has been observed between 0 and 10° W (the maritime region to the west of 10°W was outside of the observation zone) and between 2.30° N and 3.30° N.
Without frontal zone formation, such as the one observed off Cape Lopez, there are doubtlessly exchanges, according to the concept developed by Frontier (1978), between cold masses derived from coastal and equatorial upwellings and this mass of warm waters. This would explain that during boreal summer, it is in this transitional water mass (represented on the graph between 24° C and 27° C) that most of the total catch takes place (figure 7.18.a).
During the period 1980 to 1983 for FIS and Spanish purse seiners, two fishing seasons can be distinguished: the first in July, and the second, being more important, from October to December. However, Tema pole and line boats that have permanently exploited this zone for 20 years, have catch rates of skipjack, yellowfin and juvenile bigeye that do not indicate a distinct seasonality (figure 4.37).
The first purse seine fishing period in July is localized off Cape Three Points at the edge of the continental shelf which is particularly wide at this place (figure 7.18.d1). During this period, fishing is in waters with temperatures between 23 and 27° C. This means that fishing was in relation with cells of cold upwellings along the continental shelf. Traditionally, for example from 1967 to 1979, this fishing period extended in general until the months of August and September. The current shortening of the fishing season remains unexplained. Specifically the possible effect of a modification of environment change to explain this shift is hypothetical.
Figure 7.17 Catches and temperature in the “Equator” sector:
Distribution of yellowfin and skipjack according to surface temperature, FIS and Spanish purse seiners, 1980 to 1983, hauls for which surface temperature is known, from December to March.
Mean monthly surface temperature (in tenths of degrees) taken by merchant vessels in the sector from 1964 to 1984 (the period corresponding to the fishing season is indicated by dashes).
Catches of yellowfin (c1) and skipjack (c2) per month by FIS and Spanish purse seiners from 1969 to 1984.
Locations of monthly catches of large yellowfin (+30kg) yellowfin, small yellowfin (-30kg) and skipjack in the sector from April to May (d1) and from July to September (d2).
Annual catches of yellowfin, skipjack and bigeye estimated for all surface tuna fleets, 1969 to 1982 (in metric tonnes).
| EQUATOR | |
 |  |
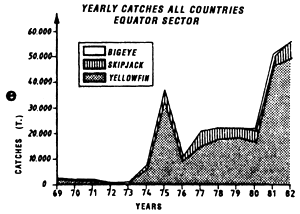 | |
Figure 7.18 Catches and temperature in the “Ghana” sector:
Distribution of yellowfin and skipjack according to surface temperature, FIS and Spanish purse seiners, 1980 to 1983, hauls for which surface temperature is known, (a1) in July and (a2) from October to December.
Mean monthly surface temperature (in tenths of degrees) taken by merchant vessels in the sector from 1964 to 1984 (the period corresponding to the fishing season is indicated by dashes).
Catches of yellowfin (c1) and skipjack (c2) per month by FIS and Spanish purse seiners from 1969 to 1984.
Locations of monthly catches of large yellowfin (+30kg) yellowfin, small yellowfin (-30kg) and skipjack in the sector in July (d1) and from October to December (d2).
Annual catches of yellowfin, skipjack and bigeye estimated for all surface tuna fleets, 1969 to 1982 (in metric tonnes).
| GHANA | |
 |  |
 |  |
 |  |
 |  |
During the second fishing season that occurs in this sector from October to December, most of the catches of the two species by purse seiners is in a thermal range from 26° C to 29° C (figure 7.18.a2). Fishing occurs much more in the high seas than during the month of July and at times when there is an increase in temperature of surface waters (figure 7.18.b.).
Another fishing period, short and not important, takes place during the first quarter of the year; it can be linked to enrichment of the zone due to coastal upwelling during the small cold season (Binet, 1976). At this time, an area to the south of the Ghana zone is included in one of the principal yellowfin spawning areas (Albaret, 1977).
The oceanic region of Cape Lopez, the limits of which are indicated in figure 7.2, is characterized by the presence of a frontal structure in boreal summer, between warm, desalinated waters to the north, and cold an salty waters to the south. The frontal zone of Cape Lopez consists, in fact, of an association of three types of fronts (Stretta, 1977):
The hydrology of this zone is described by numerous authors (Berrit, 1962; Dufour and Stretta, 1973; Morlière et al., 1974; Hisard et al., 1975; Stretta, 1977 and Wauthy, 1977) and has been presented in paragraph 3.3.3.
In this region, near Cape Lopez, an important whaling industry developed from 1910 to the 1950s (Collignon and Roux, 1950). During the whaling season that ran from mid June to the end of October, 16 to 20 whales from 12 to 18 m and 45 tonnes (Megaptere jubarte or Humpback) were fished off Cape Lopez and processed on land. In 1949, 1356 whales were caught (Acolas, 1950). The presence in the zone of high cetacean biomass obviously corresponds to the presence of large quantities of available food for predators at the top of the food chain.
Dufour and Stretta (1973) show that:
However as Frontier (1978) points out, this frontal zone is biologically richer than the two adjacent zones as “at the level of contact between two ecosystems of different maturities, an matterenergy is exported from the younger to the more mature. Warm oligotrophic water, exploiting cold rich water, becomes populated with mobile higher trophic levels elements”.
In their synthesis on Pacific tuna, Sund et al. (1981), commenting on the results of Dufour and Stretta (1973), point out the fact that zooplankton can accumulate in a passive and mechanical way at an interface and that micronecton, capable of active displacement, can been attracted to the frontal zone by zooplankton. This conclusion is not true for all taxa of micronecton and all do not migrate at the same time. These authors point out therefore that micronecton nets are, in general, very poor sampling gears for animals constituting the tuna diet.
This important biomass of edible animals for tuna, leads to a maximum concentration of tuna schools of the warm side of the frontal system (Stretta, 1977).
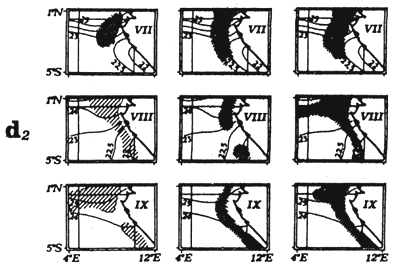
Two fishing periods in the Cape Lopez zone have been distinguished, the first in May and June and the second in July, August and September. As these three types of fronts previously described are overlapping, it is difficult to associate a fishing “stratum” specifically to one of these frontal structures. It is possible that fishing in May and June (figure 7.19.dl.) is coupled more with desalinized fronts than during the second fishing period from July to September (figure 7.19.d2.). During this second fishing period, hydrological conditions are often characterized by an accentuation of the thermal type frontal system and to the formation of a “bubble” of warm water (figure 7.20). It is within and at the periphery of this bubble of warm water that tuna are concentrated (Stretta, 1977, Stretta et al., 1977). These very specific thermal structures can develop from 1° N to 1° S between Sao Thomé Island and Cape Lopez. To the south of Cape Lopez, small very localized fronts can also develop along the coast bordering the continental shelf and be the sight of important concentrations of tuna. One must not loose sight that the coastal zone to the south of Cape Lopez is a spawning zone of Sardinella aurita in the cold season (Gheno and Fontana, 1981).
In the Cape Lopez region, the fishing maps indicate that tuna are not concentrated close to the continental shelf, as observed in other zones. Tuna exploit therefore more the productivity of the oceanic frontal system than that linked to the continental shelf. The biomass of coastal pelagic fish present in the Cape Lopez zone is relatively low compared to that observed more to the south in the upwelling zone of Angola. This appears clearly on the map of average catches of these species (figure 7.21) (One can, to a first approximation, consider that the biomasses of these species are in each zone proportional to the catches).
Here we will consider the fishing zone from 5° S to 15° S and from the coast to 10° E. This zone is characterized by the presence of a thermal dome and by strong coastal upwelling during austral winter. The Angola dome has been provisionally demonstrated by Mazeika (1967) and studied by Gallardo et al. (1974). This dome of Angola is much less distinct than the one of Guinea and even its existence is currently questionable (paragraph 3.3.4.). Nevertheless it remains true that an upward movement of the thermocline appears in February-March, to the north of the northern limit of the tradewinds (Voituriez and Herbland, 1988). During this period of the year, it is possible that the presence of tuna in this zone is linked to the productivity resulting from the thermal dome as suggested by Beardsley (1969). Moreover, Herbland and Stretta (1973) draw distribution maps of micronecton in the Angola dome and attempt to link the quantity of micronecton to the presence of tuna. In austral winter, the development of coastal upwelling leads to the formation of sharp frontal structures as demonstrated by temperatures measured by aerial radiometry in September 1973 (Vercesi, 1973).
One principal fishing period has been distinguished in this zone from September to November. Also there are sporadic catches in the first quarter during the years 1973 to 1975.
At the end of austral winter after the coastal upwelling activity, purse seiners and pole and line boats regularly fished off Angola until 1974 (figure 7.22.e). Ingham et al. (1977) present maps of skipjack schools locations in October-November 1968. All tuna school observations are very close to the coast, as is well shown in figure 4.10 showing the localities of catches during the period 1969 to 1974. From 1980 to 1983, skipjack fishing is also situated in the coastal zone in immediate proximity to the continental shelf (figure 7.22.d.). In September 1973, yellowfin and especially skipjack fishing occurs in very marked frontal zones between the 20 and 22.8° C isotherms, as temperatures taken by aerial radiometry in September 1973 show (Vercesi, 1973).
Figure 7.19 Catches and temperature in the “Cape Lopez” sector:
Distribution of yellowfin and skipjack according to surface temperature, FIS and Spanish purse seiners, 1980 to 1983, hauls for which surface temperature is known, (a1) from May to June and (a2) from July to September.
Mean monthly surface temperature (in tenths of degrees) taken by merchant vessels in the sector from 1964 to 1984 (the period corresponding to the fishing season is indicated by dashes).
Catches of yellowfin (c1) and skipjack (c2) per month by FIS and Spanish purse seiners from 1969 to 1984.
Locations of monthly catches of large yellowfin (+30kg) yellowfin small yellowfin (-30kg) and skipjack in the sector from April to May (d1) and from July to September (d2).
Annual catches of yellowfin, skipjack and bigeye estimated for all surface tuna fleets, 1969 to 1982 (in metric tonnes).
CAPE LOPEZ
 |  |
 |  |
 |  |
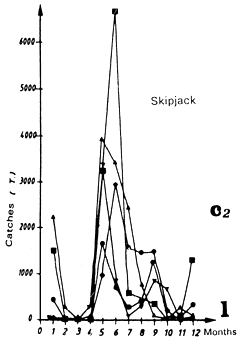 | 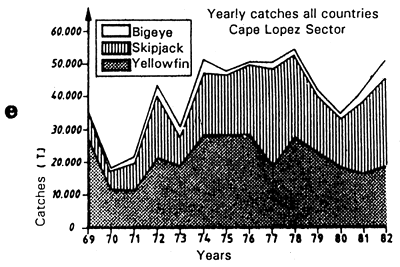 |
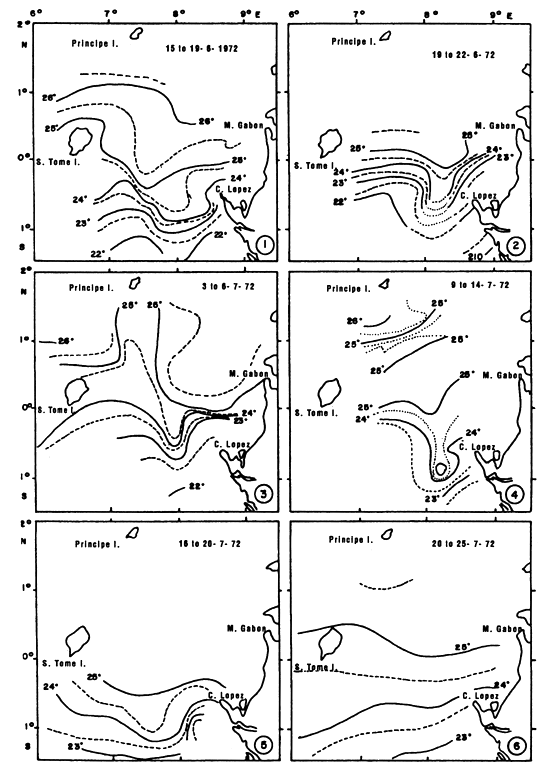
Figure 7.20 Surface isotherms observed by 5 day period in the Cape Lopez zone in June and July 1972 (after Stretta, 1977).
Figure 7.21 Mean catches of gilt sardine, carangids and scombrids during the 1979 to 1982 period (based on CECAF and ICSEAF statistical bulletins).
The description of the principal spatio-temporal fishing strata in the Gulf of Guinea show that significant tuna catches occur in the thermal range from 22° C to 29° C. Catch distribution as a function of surface temperature is very variable according to the regions considered. Aside from the surface temperature on the day of fishing, the environment parameters will act: (1) at the level of the tuna biomass, (2) at the level of fishing strategy and (3) at the level of fishing tactics.
(1) At the level of the tuna biomass.
Does environmental variability influence the dynamics of exploited stocks? There have been few studies of this subject.
Conceptually, the environment variability could intervene directly on the catchability of the stock, therefore on its rate of fishing mortality and indirectly on the biomass of the stock. Currently, the only study of effect of environmental anomalies on tuna exploitation is the one by Cayré and Roy (1985). They remark that in fishing zones off the Côte d'Ivoire (including the one we have named Ghana Zone) and off Senegal, the catch rates (expressed in c.p.u.e.), seem to respond negatively to all large thermal anomalies, whether positive or negative. These authors mention the fact “that there is no simple correlation between catch rates (c.p.u.e.) of yellowfin in the eastern tropical Atlantic and the inter-annual variability of surface temperature considered as an indicator of biological productivity in the upwelling zones”.
Variations in environmental parameters in a “nursery” could also influence the quality of food so that it would not be appropriate to the development of larvae and thus have effects on growth and mortality of larval and juvenile stages. This could result in an an exceptionally poor age class and bring about a fall in catches in a habitually very productive zone. This seems to be what happened in the Cape Lopez zone in 1975 (Stretta, 1977).
(2) At the level of fishing strategy.
Hydrological conditions favoring enrichment of a zone and therefore the presence or absence of fish will have an effect on the fishing strategy of the fleets. The food chain that comes from high primary production started by the arrival of nutrients in the euphotic layer ending up as prey animals for tuna is, at all links, subject to the synergistic action of physico-chemical and biological parameters. These parameters will directly influence the resources. Poor enrichment conditions in a zone could also influence “migratory routes” of tuna and the availability of resources.
The first hypothesis that we accept in order to explain the presence of tuna in a zone is that there is a link between its presence and the productivity of the water mass, that is to say the richer the water mass, the higher the probability of finding tuna.
The skipjack migration map (chapter 5, figure 5.21) shows that skipjack and young yellowfin displace from one fishing zone to another. From the Cape Lopez zone in the second quarter, skipjack migrate toward the Ghana zone in the third quarter and toward the Liberia and Guinea zones during the fourth quarter and the first quarter of the following year. These skipjack and young yellowfin are caught in zones that have, beforehand, undergone a process of enrichment and maturation of the water mass (Stretta and Slepoukha, 1983b).
These displacements are conditioned by an incessant quest for food to support their high energetic requirements. The presence and quantity of food available in a region will have an influence on the location and duration of fishing. Certain fishing strata are at the edge of the continental shelf above which coastal pelagic fishing is particularly active. This is the case along the continental shelf of Mauritania, Senegal, Ghana and Angola (map 7.22). This dependence of tuna vis-a-vis coastal pelagics must however be modulated. Thus, following the collapse of the sardinella stock of the Ivory-Ghana region in 1973, yellowfin and skipjack catches did not immediately diminish in the region and it was not until 1975 that tuna catches were particularly low. Regarding the duration of a fishing period, it is probable that the enrichment processes of the water mass was not a unique occurrence in time. The temperature maps of the Cape Lopez fishing zone in 1972 (figure 7.20) and those of 1974 and 1975 (Stretta, 1977) show that cold waters originating from the coastal upwelling along the coast of Gabon create several frontal systems from June to July. This process also occurs during the Ghana fishing period off the Côte d'Ivoire in boreal summer (Stretta et al., 1973). From January to March, very localized fishing is linked to the development of the “small cold season” that accompanies the appearance of cold waters in this sector (Morlière, 1970 and Binet, 1976). This fishing stratum was not analyzed above because of its very irregular character.
Can the large catches in the Guinea zone during the first fishing stratum, from March to May, be considered to be associated with the production of the Guinea Dome that seems to disappear toward the end of September (Merle, 1979)? The presence of the thermal dome of Guinea alone does not seem to explain catches of skipjack and yellowfin from November to March. If Mazeika's (1978) temperature maps at 20 m from January to April, as well as those at 20 and 50 m from January to March (Merle (1978), do not show a domed thermal structure in this zone, the disappearance of the dome seems to be put to question following recent oceanographic cruises (Le Bouteiller, pers. comm.). Moreover, the formation of a frontal zone between waters originating from the Senegal upwelling and the warm waters could support production in the region of the Guinea dome. Also, fishing in this sector could be associated with water mass enrichment by river sources or by convergence cells. Another hypothesis, not to be dismissed, is that of fishing with fish natural aggregation devices; Stretta and Slepoukha (1986) point out that in the coastal zones of Liberia, the proportion of fishing on debris is over 40%.
In the Gulf of Guinea, tuna are fished in waters where no productive system is indicated before-hand or has not been detected on the surface). Would the absence (or the low intensity) of structures favoring enrichment of water masses have an influence on the presence of fish?
In the Equatorial zone, the presence of the thermal ridge could maintain high productivity all year as recently proposed by Herbland et al. (1985). In the warm season, especially in the equatorial zone, a form of enrichment occurs, as mentioned above, which is difficult to quantify and linked to the action of long waves trapped at the equator, favoring an upwelling of infrathermoclinal waters (rich in nitrate) in the euphotic layer. Sometimes, these upwelling cells reach the surface and can be detected. Herbland et al. (1983) consider that in the eastern tropical Atlantic, short term variations of hydrological conditions (vertical and horizontal circulation and associated turbulent mixing) have probably more importance than one would think; it seems that the entire equatorial zone (between 1o N and 7o S) is subjected to “vertical pulsations” very limited in space and time, poorly explained, and, in any case, not foreseeable. A meteorological satellite equipped with an infrared radiometer is the preferred gear for locating this type of variation of surface hydrological conditions that is the basis for local enrichment of the water mass.
To the first hypothesis based on tuna displacement conditioned principally by their quest for food, the hypothesis of concentrations linked to reproduction must not be omitted. Yellowfin spawning occurs mainly during the first quarter of the year and occurs in the eastern tropical Atlantic in two zones, the principal zone being in the “heart” of the Gulf of Guinea from 5o N to 5o S and from the African coast to the Greenwich meridian; a second spawning zone is off Liberia (Albaret, 1977). For skipjack, the problem is different as this species seems to have a more opportunistic reproductive strategy. Cayré and Farrugio (1986) suggest that the zones where the temperature is over 24° C, and in which the biomasses of phytoplankton and zooplankton are high, seem particularly favorable to skipjack reproduction in the Atlantic.
Following the works of Kitchell et al. (1978), energetic needs of Pacific skipjack and juvenile yellowfin have been estimated. From these authors, a skipjack less than 1 kg consumed between 7.3% and 19% of its weight per day. For juvenile yellowfin, the values obtained are close to those calculated for skipjack. The more recent and complete works of Olson and Boggs (1986) in the Eastern Pacific estimate the food ingested daily by yellowfin to between 3.9% and 6.7% of the weight of individuals, depending on estimation methods. The biomass of the three principal species (yellowfin, skipjack and bigeye) is estimated at around 1.5 million tonnes (chapter 8) and would need annually from 21 to 37 million tonnes of food. These energetic needs are enormous. Moreover, we know that tuna have a very opportunist feeding behavior. It is not surprising therefore that zones rich in tuna are more often those where food is abundant.
To find zones of tuna concentration, it would suffice therefore to look for high density food regions; the search for these regions leads us to locate zones of high productivity in the ocean and to follow them in space and time. We have seen above that in tropical waters, the system that plays a primary role in enrichment of the epipelagic layer is the upward movement of the thermocline, whether to the surface (in the case of upwelling or fronts), or just to the euphotic layer (as in the case of domes or the equatorial ridge).
This leads us to examine the concept of the hydrobiological history of water masses. This notion will permit us to approach the problem of tuna food localization.
The superficial cooling process, indicating the enrichment of water, can often be specifically detected by satellite. One of the regrettable limits in the use of satellite remains the current impossibility of detecting hydrological structures in the form of a ridge or thermal dome, which do not translate into superficial cooling. However it will probably be possible in the future to detect this type of structure by analysis of water color from satellite equipped with a radiometer analyzing the visible spectrum back scattered by the sea.
In favorable cases where it is possible to observe the formation of a productive zone by satellite (primary and secondary productivity), it can be seen that some delay is required to observe the concentration of tuna in the sector. In the simplest case, it will be possible to follow changes in space and time of the thermal “perturbations” by satellite in order to forecast periods and zones where the probability of finding prey animals and tuna will be high. Analysis of detailed fishing statistics, daily catches with positions in degrees and minutes, will enable the observation of the timetable of the appearance of tuna in the zone, provided the zone is explored by the fishery.
Figure 7.22 Catches and temperature in the “Angola” sector:
Distribution of yellowfin and skipjack according to surface temperature, FIS and Spanish purse seiners, 1980 to 1983, hauls for which surface temperature is known: from September to Novemb
Mean monthly surface temperature (in tenths of degrees) taken by merchant vessels in the sector from 1964 to 1984 (the period corresponding to the fishing season is indicated by dashes).
Catches of yellowfin (c2) and skipjack (c1) per month by FIS and Spanish purse seiners from 1969 to 1984.
Locations of monthly catches of large yellowfin (+30kg) yellowfin, small yellowfin (-30kg) and skipjack in the sector from September to November.
Annual catches of yellowfin, skipjack and bigeye estimated for all surface tuna fleets, 1969 to 1982 (in metric tonnes).
| ANGOLA | |
 |  |
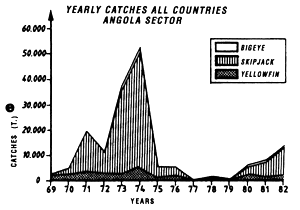 | |
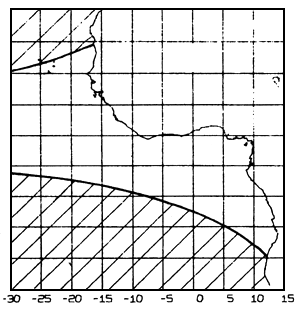
Figure 7.23 Zone of strong July winds (speed above 6 m/s), i.e. unsuitable for purse seine operations (based on Hastenrath and Lamb, 1977).
On the basis of the principal of following a water mass in space and time, the predictive model “Previpeche” developed by Stretta and Slepoukha (1983, 1983a) makes predictions of favorable or unfavorable zones for the presence of tuna in the eastern tropical Atlantic.
The analysis of Mendelsohn and Roy (1986) confirms the statistical value of this concept: these authors, analyzing the relation between catches yellowfin and skipjack by the FIS tuna fleet and environmental data (surface temperature and wind speed) from 1969 to 1979 by fifteen day period, show that the environmental parameters in certain zones could be indicators of the oceanographic mechanisms that are the origin of favorable fishing conditions.
Several serious problems remain concerning the exact relation between the richness of a zone in food and tuna abundance:
It is unknown if the observation, made by Roger and Grandperrin in the Pacific, that tuna do not seem to feed on migrant fauna can be transposed in the Gulf of Guinea; hydrological conditions are very different in both oceans, especially the depth of the homogeneous layer. Hosever, Roger (pers. comm.) thinks that the pattern of the Pacific would be applicable to the Atlantic Ocean.
The importance of coastal pelagic fish in the diet of tuna remains difficult to estimate. This would be important to know as the ecology and dynamics of these species is intrinsically different to that of planktonic organisms, in particular their potential capacity to survive longer than plankton in zones poor in food (at least for adults).
The ability of tuna to survive long periods with little food in its natural environment, in particular during the migration or reproductive phases, remains largely unknown.
(3) At the level of fishing tactics
Limits and variations of the environmental parameters will have a short term action on catch to which the fisherman will be particularly susceptible. Surface temperature, depth and gradient of the thermocline, content of dissolved oxygen, wind force, current, will act on the behavior and availability of the fish.
A thin homogeneous layer accompanied by oxygen under 3.5 ml/l as is found off Angola (Ingram et al., 1977), will favor the phenomenon of fish concentration.
A strong wind, for example over force 4 on the Beaufort scale, does not permit purse seiners to work efficiently and only pole and line boats can fish: thus, not many purse seiners can operate in Mauritania during summer, a zone where the trade winds blow. The zones of strong wind can cover large surfaces in the study zone as shown in figure 7.23. Table 7.3 notes the scarcity of purse seine sets carried out in strong winds. This table gives, for FIS purse seiners, percentages of sets carried out in the Gulf of Guinea in winds under or over force 4 on the Beaufort scale. Only 2.8% of sets took place in a wind over or equal to force 4, even though these winds are frequent in the zone.
Although the effect of the current seems to have little influence (Stretta and Slepoukha, 1986), near the equator (from 10 to 20 miles on either side), in spite of the presence of tuna, a purse seine master will hesitate to set a purse seine because of the presence of the Lomonosov under current. On the surface, the south equatorial current moves to the west-north-west, and around 20 – 40 meters, the Lomonossov current moves to the east, that which brings about shearing of the net and a strong probability of damage and a null set.
It happens at times that the fish are concentrated in a very restrained space. It is the phenomenon of concentration described by Fonteneau (1985) where several thousands of tonnes of tuna are concentrated in the surface dozens of miles from the coast. This could be the result of a concentration of food linked to the appearance of specific hydrological conditions (current convergence, formation of a front particularly active, etc.). It is also possible that the fish are aggregating in a spawning area.
Fish can be found associated with an natural FAD (fish aggregation device); we can not give an environmental explanation for this type of association.
A purse seiner can be faced with a particular fish behavior and will have difficulties encircling them: this is the case when the fish are dispersed, swim too fast or are found too deep. Can these behaviors be attributed to environment parameters?
There are situations that also influence on fishing tactics, although environment parameters are not implicated in the tuna catch; we will limit ourselves to listing some. A fishing gear can have null catches in an area with abundant fish, as is the case for purse seiners finding themselves on a school of large yellowfin or bigeye. A pole and line boat can find itself in a tuna rich zone during baiting; it can also find itself in a region where schools of tuna do not respond to bait and refuse to bite the hooks as a result, for example, of a high abundance of natural food.
Free access to exclusive economic zones constitutes also a factor poorly appreciated by scientists. It is very probable that most of the purse seine fleets operating in the region have recently restrained their fishing activity in certain coastal sectors, not because of the absence of tuna, but because of the absence of fishing agreement with neighboring countries.
Table 7.3 Table comparing percentages of sets by 10 tonne class made with winds from force 1 to 4 and above force 5.
| Null sets | 1 to 30 T | 11 to 20 T | 21 to 30 T | 31 to 40 T | 41 to 50 T | 51 T | Nbr sets | |
|---|---|---|---|---|---|---|---|---|
| Force 1 to 4 | 20.8% | 27.8% | 18.5% | 10.9% | 6.5% | 4.9% | 10.7% | 5 839 |
| Force 5 + | 30.2% | 23.4% | 19.7% | 6.8% | 3.7% | 6.2% | 9.9% | 168 |
The present analysis shows complexity of relations between tuna and environment. If it seems certain at present that zones rich in tuna are in general zones ecologically the most productive, the linkages between tuna and the environment prove to be very complex to analyze and understand in detail, in particular when predictions are to be made at an small spatio-temporal scale. These must be based as much on biological knowledge pertaining to species (biology, behavior, state of stocks), as on knowledge pertaining to environment. Numerous questions remain on mechanisms in play, and on the respective importance of various factors. Only new detailed research oriented toward the study of the relations between tuna and their environment will answer these multiple questions.
![]()
![]()
![]()