![]()
![]()
![]()
Sous les latitudes moyennes et élevées (supérieures à 35°) la distribution des mises bas dans l'année n'est pas uniforme et la plupart des races locales d'ovins et caprins donnent naissance aux jeunes à la fin de l'hiver et au début du printemps. Dans les races primitives de moutons tels que le mouton de Soay, originaire du nord de l'Ecosse, les agnelages ont lieu en avril. Beaucoup d'autres races de mouton en Europe du Nord sont dans la même situation. Toutefois, avec la diminution de la latitude, la distribution saisonnière des parturitions des races locales est de plus en plus variable. Chez les Ile-de-France, en France, deux pics de mises bas sont enregistrés, l'un en automne, l'autre en hiver; pour les races méditerranéennes de brebis (la Chios en Grèce, la Barbarine en Tunisie), les agnelages d'automne sont plus fréquents que ceux de printemps. Chez la brebis mérinos australienne, les mises bas peuvent avoir lieu sur plusieurs saisons. Enfin, chez les races tropicales ou subtropicales, comme les brebis peule au Niger ou les brebis Black Belly à La Barbade, les agnelages se produisent toute l'année.
Dans l'espèce caprine, la même situation est observée: les caprins des latitudes moyennes et élevées sont très saisonnés (Alpine, Saanen et Poitevine en France), alors que sous les tropiques ou les subtropiques, aucun saisonnement net et répétable d'une année sur l'autre n'est observé. C'est le cas de la chèvre créole des Antilles françaises, de la chèvre SRD (Sem Raça Defînida) au Brésil, de la chèvre naine ouest-africaine et de bien d'autres populations vivant sous de telles latitudes. Ainsi, la distribution des mises bas au cours de l'année est clairement fonction de la latitude.
Puisque la gestation dure cinq mois chez les petits ruminants, la période des accouplements fertiles des races saisonnées a lieu en fin d'été et en automne/hiver. Les variations saisonnières d'activité sexuelle concernent à la fois le mâle et la femelle.
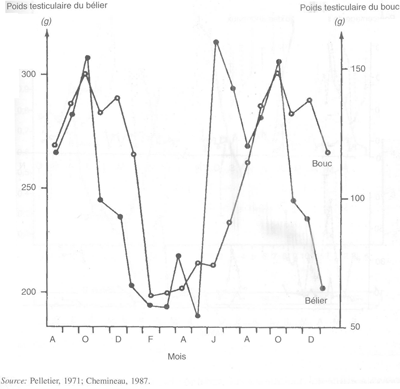
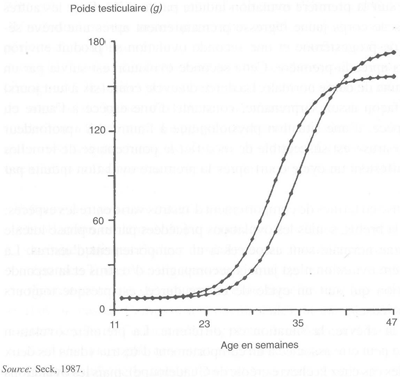
Sous les latitudes moyennes et élevées, la spermatogénèse ne s'arrête pas, mais le nombre de spermatozoïdes produits par le testicule diminue à certaines saisons de l'année. Un gramme de testicule de bélier Ile-de-France produit 12,2 x 106 spermatozoïdes en automne contre seulement 9,3 x 106 au printemps, à cause de la diminution du rendement de la spermatogénèse. Quelques classes de cellules germinales dégénèrent bien que leur vitesse de développement ne soit pas modifiée. Ces modifications saisonnières de l'activité spermatogénétique entraînent des changements importants de poids testiculaire: de 200 g en mai à plus de 300 g en août chez le bélier Ile-de-France et de moins de 100 g en mai à plus de 150 g en septembre chez le bouc alpin (figure 25). Ces variations de production spermatique peuvent également être observées dans les éjaculats, à condition cependant, que la fréquence de récolte soit suffisamment élevée. En dehors de la saison sexuelle, le nombre total de spermatozoïdes par éjaculat diminue plus rapidement avec le numéro d'ordre des éjaculats successifs, que pendant la saison sexuelle.
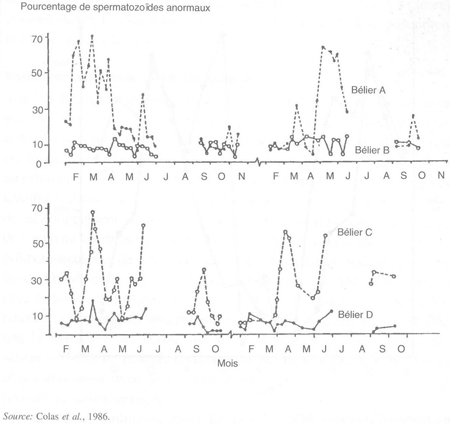
L'importante diminution observée dans l'activité spermatogénétique du bélier pendant le printemps est associée à une augmentation de la fréquence des spermatozoïdes porteurs d'anomalies morphologiques. Chez certains mâles, le pourcentage de spermatozoïdes anormaux peut atteindre plus de 70 pour cent certains mois de l'année, entraînant une stérilité temporaire des animaux (figure 26). Cette caractéristique se répète d'une année sur l'autre pour les mêmes animaux, au printemps.
Chez le bouc, aucune variation saisonnière importante du pourcentage d'anomalies spermatiques n'a été mise en évidence; toutefois, un changement saisonnier de la motilité des spermatozoïdes se produit, associé à une diminution sévère de la fertilité de ceux-ci. Cette altération a été observée quatre années consécutives chez les mêmes mâles, et chaque fois pour une Murée d'environ trois mois, mais à une période imprévisible au cours du printemps et de l'été.
FIGURE 25 Variations saisonnières du poids testiculaire du bouc alpin et du bélier Ile-de-France
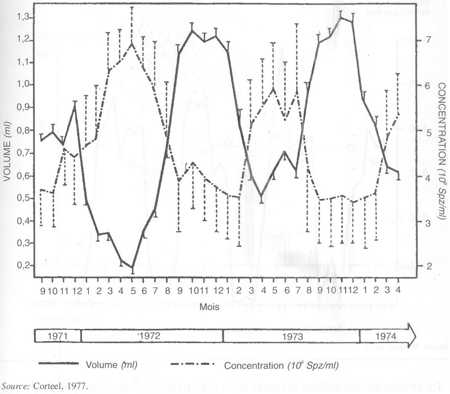
Chez les races saisonnées, le volume de l'éjaculat est élevé durant la saison sexuelle et il diminue au printemps, pour atteindre son minimum pendant l'été. La concentration spermatique de l'éjaculat en spermatozoïdes suit une évolution inverse (figure 27). Ces variations reflètent celles de la synthèse et de la sécrétion du plasma séminal par les glandes annexes qui sont stimulées quand la testostérone est haute pendant la saison sexuelle, et au repos quand elle est basse durant la contre-saison.
FIGURE 26 Variations à long terme du pourcentage de spermatozoïdes anormaux chez quatre béliers, montrant les variations saisonnières et individuelles de ce paramètre
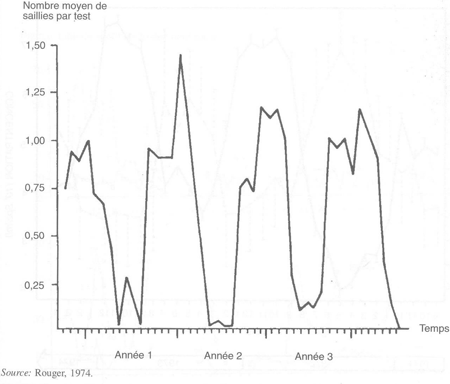
Ces variations sont aussi associées à une diminution de l'intensité du comportement sexuel pendant le printemps, chez les béliers et les boucs (figure 28). Sans entraînement régulier, les montes et les saillies cessent chez presque tous les animaux pendant quelques semaines ou mois au cours du printemps/été; chez ceux qui continuent de s'accoupler, le temps de réaction augmente. Toutefois, ces variations sont atténuées si un entraîne- ment régulier est effectué. Pour la collecte de la semence, l'entraînement régulier des mâles tout au long de l'année, à heures et jours fixes, évite l'arrêt complet du comportement sexuel et avec une telle conduite il est possible de collecter la quasi-totalité des mâles entraînés (voir Collecte et conservation de la semence, chapitre 4).
FIGURE 27 Variations saisonnières du volume de l'éjaculat et de sa concentration en spermatozoïdes chez cinq boucs alpins, m ± sem
En revanche, les béliers et boucs des races tropicales et subtropicales, s'ils sont alimentés correctement, ne manifestent pas de variations saisonnières de leurs activités spermatogénétique et comportementale (figure 29). Dans certains cas, toutefois, la situation peut être compliquée par le fait que dans les pays tropicaux et subtropicaux, les températures élevées des saisons chaudes provoquent l'apparition de spermatozoïdes anormaux et morts. C'est le cas du bélier de la race Barbarine de Tunisie (figure 30).
FIGURE 28 Variations saisonnières du nombre de saillies dans des tests de 10 minutes chez des boucs alpins non entraînés
Au contraire des mâles ovins et caprins qui produisent des spermatozoïdes tout au long de l'année, les brebis et les chèvres des races saisonnées cessent de manifester des œstrus et d'ovuler pendant plusieurs mois successifs.
FIGURE 29 Variations mensuelles du diamètre testiculaire et du nombre de saillies dans des tests de 25 minutes chez six boucs créoles
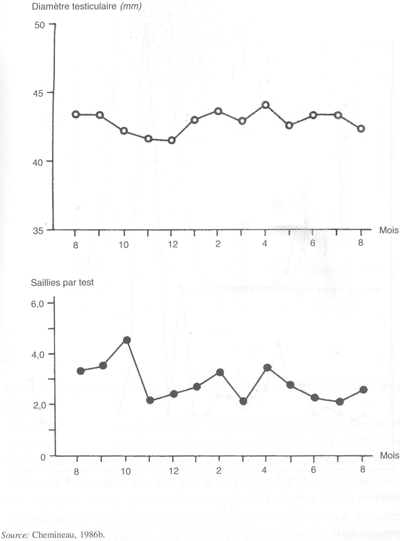
FIGURE 30 Apparition au cours de l'année de spermatozoïdes morts dans la semence de bélier de race barbarine
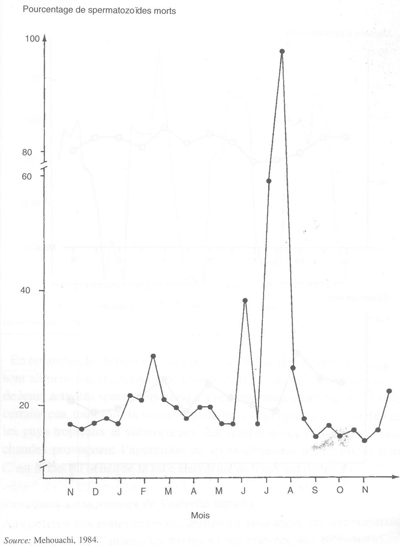
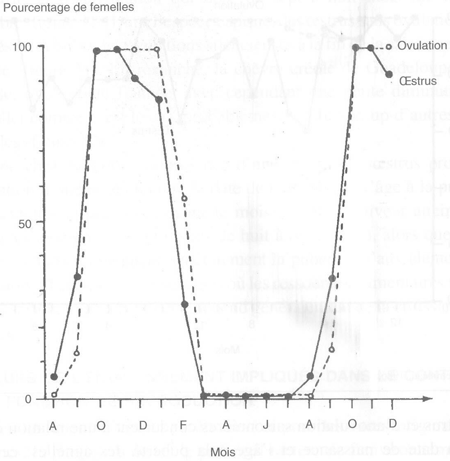
FIGURE 31 Variations saisonnières dans l'apparition mensuelle du comportement d'œstrus et de l'ovulation chez 15 chèvres alpines
Les brebis Ile-de-France cessent leur cyclicité ovulatoire et œstrale pendant six mois (de fin janvier à fin juillet), sauf un à deux cycles en mai, et les brebis Préalpes du Sud pendant quatre mois (de mars à fin juillet). Quelques races sont très saisonnées (Suffolk, Texel, Blackface, etc.) alors que d'autres (Ile-de-France, Préalpes-du-Sud, Dorset Horn, mérinos, etc.) le sont moins. Des ovulations silencieuses (ovulations non associées à un comportement d'œstrus) peuvent se produire pendant l'anœstrus et sont généralement observées au début de la saison sexuelle. Ces périodes d'anœstrus et d'anovulation saisonnières conduisent à une relation étroite entre la date de naissance et l'âge à la puberté des agnelles; celles-ci n'atteignent la puberté que pendant les mois correspondant à la saison sexuelle de l'adulte. Si l'anœstrus se situe tous les ans approximativement à la même période, des différences entre années existent dans la date moyenne du début et de la fin des activités ovulatoire et œstrale.
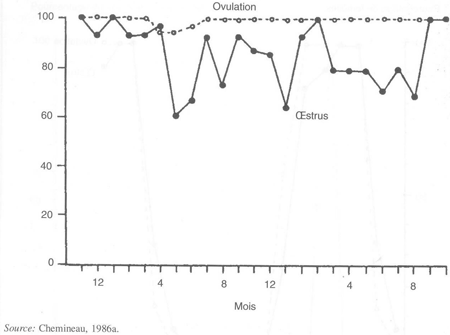
FIGURE 32 Variations saisonnières dans l'apparition mensuelle du comportement d'œstrus et de l'ovulation chez 15 chèvres créoles
Lorsque la latitude diminue, le saisonnement des races locales est de moins en moins marqué et les durées individuelles d'anœstrus raccourcissent. Dans les régions subtropicales, quelques races maintiennent leur cyclicité ovulatoire toute l'année (D'Man au Maroc, Ossimi en Egypte) et d'autres présentent un faible saisonnement de leurs activités ovulatoire ou œstrale (Barbarine en Tunisie, Rhamani en Egypte); mais aucune d'entre elles ne manifeste les importantes variations observées chez les races des latitudes plus élevées. Sous les tropiques, les brebis, si elles sont alimentées correctement, ovulent toute l'année (Black-Belly de La Barbade, Créole des Antilles françaises, Peule du Niger).
Sous les climats tempérés, les chèvres laitières ont également une saison d'anœstrus et d'anovulation qui dure de sept à huit mois (de mars à septembre; figure 31). Dans l'espèce caprine, des œstrus sans ovulation sont observés au début et des ovulations silencieuses à la fin de la saison sexuelle annuelle (figure 31). En revanche, la chèvre créole de Guadeloupe, race tropicale, ovule toute l'année, avec cependant une petite diminution en juin/juillet comme c'est le cas, probablement, de beaucoup d'autres races tropicales (figure 32).
Comme chez la brebis, la présence d'une saison d'anœstrus provoque l'apparition d'une relation entre la date de naissance et l'âge à la puberté. Les chevrettes alpines nées avant le mois de mars peuvent atteindre la puberté à l'automne suivant (âgées de huit à onze mois), alors que celles nées après mars n'atteignent généralement la puberté qu'aux alentours de 16-18 mois. Dans les zones tropicales où les ressources alimentaires varient selon la saison, l'âge à la puberté dépend généralement de la croissance des animaux.
Sous les latitudes moyennes et élevées, et pour les races originaires de ces zones, la photopériode est le principal facteur de l'environnement qui contrôle les variations saisonnières de reproduction des petits ruminants. Dans les deux sexes, l'activité gonadique et le comportement sexuel varient avec la durée du jour. Les autres facteurs de l'environnement, comme la température, le régime alimentaire ou les facteurs sociaux, agissent comme des modulateurs de l'activité sexuelle. Sous les latitudes tropicales ou subtropicales, les races locales d'ovins et de caprins semblent moins sensibles aux faibles variations photopériodiques existant dans ces zones, alors que les autres facteurs de l'environnement jouent un rôle bien plus important. Nous examinerons donc ici successivement les effets de ces différents facteurs de l'environnement sur la reproduction des ovins et des caprins.
FIGURE 33 Activité ovulatoire de brebis Ile-de-France et de chèvres alpines recevant une alternance de jours longs (16 heures de lumière/8 heures d'obscurité) et de jours courts (8 heures de lumière/16 heures d'obscurité)
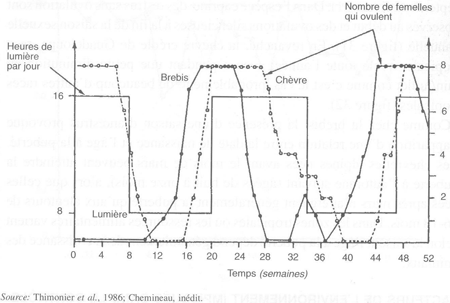
Races des latitudes moyennes et élevées. Quand les races européennes d'ovins sont transportées dans des pays de latitude moyenne de l'hémi- sphère Sud, tels que l'Australie et l'Argentine, elles réajustent leur saison de reproduction à l'automne austral. Expérimentalement, un décalage de six mois des changements de la durée du jour annuelle, induit un décalage de six mois dans l'activité sexuelle de béliers Ile-de-France, de brebis Southdown et de différents croisements de races anglaises. L'activité sexuelle reste liée à l'automne photopériodique. Le même effet est égale- ment démontré lorsqu'un régime photopériodique artificiel avec une période inférieure à 12 mois est utilisé. Ainsi, des régimes lumineux qui reproduisent en six mois les variations photopériodiques annuelles, induisent, à six mois d'intervalle, des périodes d'activité sexuelle chez le bélier et chez la brebis. Des résultats identiques sont obtenus chez la chèvre, mais la réponse semble plus tardive que celle obtenue chez les ovins (figure 33).
FIGURE 34 Mélatonine plasmatique chez des brebis Ile-de-France en jours longs et en jours courts
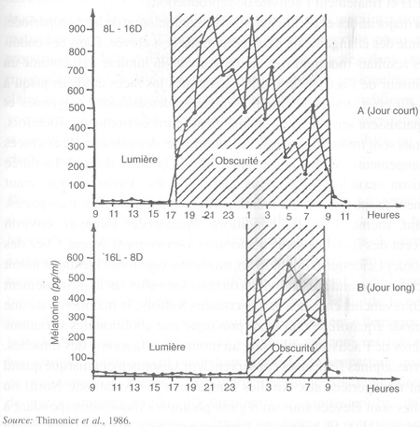
Les mécanismes physiologiques impliqués dans le contrôle photopériodique de la reproduction ne sont, pour le moment, que partiellement connus. La lumière est transmise des yeux à la glande pinéale par voie nerveuse, laquelle inclut le ganglion cervical supérieur. La glande pinéale synthétise et sécrète la mélatonine dans le plasma sanguin quand les lumières sont éteintes et s'arrête lorsque celles-ci sont allumées (figure 34). Le rythme circadien de sécrétion de mélatonine, qui dépend donc de la durée du jour, détermine l'activité des neurones hypothalamiques qui contrôlent la sécrétion de LH et finalement l'activité de reproduction.
Dans la majorité des expérimentations de manipulation de la photopériode, l'amplitude des changements photopériodiques est élevée. Dans ces conditions, les résultats montrent très clairement que la lumière agit comme un synchroniseur de l'activité sexuelle dans toutes les races étudiées jusqu'à présent. En effet, toutes les races originaires des latitudes moyennes et élevées paraissent sensibles aux changements photopériodiques. Toutefois, les résultats sont moins clairs lorsque l'on soumet des animaux de ces races à des changements de plus faible amplitude de la durée du jour. La durée de la saison sexuelle de la brebis mérinos a été décrite comme étant inversement proportionnelle à la latitude où les animaux sont transportés. Cependant, même dans des conditions équatoriales (Kenya), environ 50 pour cent des femelles sont en anœstrus à un moment donné. Chez des brebis Border Leicester et Southdown, un régime équatorial identique induit un asynchronisme entre les individus ou reste sans effet sur le saisonnement initial. En revanche, chez des brebis croisées Suffolk, le maintien sous une photopériode équatoriale constante provoque une abolition des variations saisonnières de l'activité ovulatoire, au moins chez la moitié des femelles. Des chèvres alpines françaises conservent leur saisonnement marqué quand elles sont transportées aux Antilles françaises (16° de latitude Nord) ou lorsqu'elles sont élevées sous un régime photopériodique correspondant à cette latitude (11 à 13 heures de lumière par jour), même si une légère augmentation de la durée de la saison sexuelle est observée les deux premières années. Le maintien de ces races ovines et caprines «tempérées» sous un régime photopériodique «tropical» induit l'apparition de cycles œstraux et ovulatoires anormaux. Les pourcentages de cycles courts, d'ovulations silencieuses et de comportements d'œstrus sans ovulation sont deux fois plus élevés chez les chèvres maintenues sous un régime tropical (11 à 13 heures de lumière), que chez les chèvres témoins (recevant 8 à 16 heures de lumière par jour). Ces anomalies sont surtout apparentes durant les deux premières années pendant lesquelles les femelles subissent cette photopériode.
Ainsi, même si les résultats qui viennent d'être présentés montrent qu'il existe une certaine variabilité dans la réponse, aucune race originaire des climats tempérés ne manifeste de changement important dans la durée de sa saison sexuelle lorsqu'elle est transportée sous un climat tropical. En particulier, une saison d'anœstrus est toujours observée pour une large proportion de la population. Lorsqu'elles sont placées dans un régime équatorial strict (aucune variation photopériodique), la situation est moins claire et il semble bien que certaines races, dans ces conditions, puissent voir leur saisonnement fortement atténué. Cela pose la question de savoir si les races originaires des faibles latitudes sont sensibles ou non aux changements de la photopériode.
Races des faibles latitudes. Quand des brebis javanaises à queue fine sont soumises à une alternance de trois mois de jours courts (huit heures de lumière par jour)/six mois de jours longs (16 heures de lumière par jour)/trois mois de jours courts, les femelles continuent à ovuler sans interruption, comme dans les conditions naturelles (3° de latitude Sud) bien que le comportement d'œstrus disparaisse à la fin de la période de jours longs. Des boucs nains africains, importés en Allemagne (51° de latitude Nord) ne montrent pas de variations saisonnières très nettes de leur comportement sexuel ou de leur production spermatique quantitative ou qualitative. Des brebis D'Man marocaines soumises à une photopériode de grande amplitude (celle d'Edimbourg) ne manifestent pas de changements dans leurs ovulations cycliques régulières en comparaison des témoins laissées dans des conditions naturelles. Toutefois, les résultats sont encore trop limités pour pouvoir tirer des conclusions définitives, mais ils suggèrent que les animaux des races tropicales ou subtropicales n'ont pas adapté complètement leur activité de reproduction aux nouvelles conditions photopériodiques caractérisées par une plus grande amplitude.
En conclusion, les races des latitudes moyennes et élevées sont soumises naturellement à des variations photopériodiques de grande amplitude. En leur absence, la plupart de ces races manifestent quand même des variations qui peuvent être considérées comme l'expression d'un rythme endogène annuel de reproduction, ce qui suggère une composante génétique du saisonnement de la reproduction. Les races originaires des faibles latitudes ne manifestent pas, ou presque pas, de saisonnement et semblent présenter une sensibilité réduite à la photopériode. Les variations saisonnières de reproduction sont donc toujours le résultat d'une forte interaction génotype/milieu.
Normalement, sous les latitudes moyennes et élevées, l'environnement thermique n'est pas l'entraîneur principal de l'activité sexuelle. Toutefois, en climat tropical, la température est susceptible de limiter les aptitudes de reproduction, particulièrement dans les races importées des zones tempérées.
Activité de reproduction du mâle. Le nombre maximum d'éjaculats obtenus en une heure par des béliers Dorset Horn et Border Leicester diminue quand la température ambiante augmente; en revanche, des béliers mérinos sont capables de maintenir leur activité sexuelle à des hautes températures. De surcroît, de nombreuses études indiquent clairement que des températures élevées affectent négativement la qualité de la semence avec une diminution du pourcentage de spermatozoïdes mobiles et de leur motilité, ainsi qu'un accroissement des formes anormales. Les anomalies apparaissent, généralement, au niveau de la tête (têtes piriformes, acrosomes endommagés, spermatozoïdes sans queue), mais également par la présence de gouttelettes cytoplasmiques et de flagelles recourbés. Les effets délétères des fortes températures sur la production spermatique se produisent à la suite d'une augmentation de la température testiculaire qui provoque l'apparition de dégénérescences spécifiques avec manifestation d'anomalies à des stades critiques précis du cycle spermatogénétique. Ces effets peuvent être reproduits expérimentalement en chauffant les testicules, ou en gardant les animaux dans un endroit chaud pendant quelques heures. Les spermatozoïdes modifiés apparaissent dans l'éjaculat au cours de la seconde semaine post-traitement, et une réduction du nombre total de spermatozoïdes se produit quelques jours plus tard, à 20 jours du traitement. La fécondance d'une telle semence commence à diminuer au cours de la deuxième semaine et dure jusqu'à la troisième et quatrième semaine posttraitement. Cette diminution du pouvoir fécondant est probablement le résultat de défauts de fécondation et d'accroissement de la mortalité embryonnaire.
Le retour progressif à une qualité et une fécondabilité normales dépend de la durée et de l'intensité du stress; il nécessite de 50 à 60 jours chez le bélier. Six heures d'augmentation de la température corporelle (41°C) peuvent être suffisantes pour induire une dégénérescence spermatique chez le bélier. L'exposition de mâles à des températures naturellement élevées pendant de telles périodes sur plusieurs jours consécutifs peut déclencher des désordres similaires à ceux observés après une exposition continue en chambre climatique. Il est important de remarquer que de telles élévations de température corporelle peuvent également se produire à la suite d'une maladie ou d'une infection pouvant passer inaperçue.
La température ambiante limite audelà de laquelle la semence de béliers des races européennes est affectée est située aux alentours de 29-30°C. Mais la sensibilité des mâles à la température ambiante varie selon la race. La spermatogénèse des races ovines indigènes d'Inde n'est pas affectée par les températures élevées; l'exposition de boucs créoles locaux de Guadeloupe à la lumière solaire directe ne modifie pas la qualité de leur semence. Une variabilité intrarace existe également, ce qui suggère une possible origine génétique. Quand deux groupes de béliers mérinos sont choisis pour leur température basse ou élevée, la fertilité des brebis est plus faible lorsque celles-ci sont accouplées avec les mâles à température élevée. Les raisons de ces différences individuelles ne sont pas connues, mais, dans l'espèce caprine, la division du scrotum en deux poches distinctes qui conduit à une meilleure thermorégulation du testicule, a été décrite dans plusieurs races tropicales qui sont mieux adaptées au climat chaud.
Activité de reproduction de la femelle. Dans le cas de la brebis mérinos, un accroissement de la charge thermique radiative pendant les jours 10 à 15 du cycle œstral accroît la durée du cycle, diminue la durée, voire supprime totalement l'œstrus qui suit et, finalement, diminue la fertilité. L'exposition de brebis à des températures élevées à différents stades après l'accouplement entraîne une diminution du taux de fertilité. La principale perte d'œufs se produit au cours des premiers stades de développement embryonnaire. Des œufs fécondés (deux à 32 cellules) placés dans le tractus génital de brebis maintenues à des températures ambiantes élevées (32°C) sont dégradés lorsqu'ils entrent dans l'utérus. La perte embryonnaire maximale se produit donc lorsque le stress thermique est imposé entre les jours 1 et 16 après l'insémination. Le nombre minimum de jours consécutifs nécessaires pour observer un effet du stress thermique va de deux à sept. In vitro, les fortes températures modifient l'aptitude des œufs fécondés à se développer en embryons viables et diminuent la vitesse de développe- ment embryonnaire.
Une fois implanté, l'embryon est moins sensible à un stress thermique appliqué à la mère. Toutefois, pendant la deuxième moitié de la gestation, le maintien dans un environnement thermique contraignant provoque une diminution de la croissance fœtale qui se traduit par des poids de naissance inférieurs de moitié à ceux des agneaux nés des mères témoins. Dans ce cas, une forte corrélation négative existe (r = - 0,93) entre le poids de naissance des agneaux et la température rectale des mères. L'exposition de brebis à un stress thermique uniquement diurne ne fait que limiter la réduction du poids de naissance par rapport aux brebis soumises à un stress permanent. Ces effets de l'environnement thermique sur la mortalité embryonnaire et la croissance des fœtus se produisent sans doute à cause d'une augmentation de la température utérine. Cette augmentation pourrait être due à une diminution du flux sanguin irriguant l'utérus.
Néanmoins, comme chez le mâle, d'importantes différences existent entre races quant à leur sensibilité au stress thermique. Dans un climat tropical, où la température de l'air varie entre 22 et 30°C à midi, la fertilité de brebis Pelibuey, une race locale de Cuba, atteint 74 pour cent, comparé à seulement 25 pour cent chez des brebis Suffolk importées, que les femelles soient accouplées avec un mâle Pelibuey ou Suffolk (tableau 6). Cette importante différence raciale est due à la forte sensibilité thermique des brebis Suffolk, comme l'attestent leur rythme respiratoire et leur température rectale très élevés, comparés à ceux des brebis Pelibuey (tableau 7).
TABLEAU 6 Fertilité à l'œstrus naturel de brebis Pelibuey et Suffolk accouplées avec des béliers Pelibuey ou Suffolk, en climat tropical
| Béliers Pelibuey | Béliers Suffolk | |||
|
(%) |
(nombre) |
(%) |
(nombre) |
|
|
Brebis Pelibuey |
84 |
25 |
64 |
25 |
Source: Fuentes et Chemineau, 1990.
TABLEAU 7 Rythme respiratoire et température rectale de brebis Pelibuey et Suffolk le jour de l'œstrus, 8-10 jours et 17-18 jours plus tard
|
Brebis Pelibuey |
Brebis Suffolk |
|
|
Jour 0 |
||
|
Rythme respiratoire/minute |
61 |
110 |
|
Température rectale (°C) |
39,1 |
39,9 |
|
Jour 8-10 |
||
|
Rythme respiratoire/minute |
40 |
100 |
|
Température rectale (°C) |
38,8 |
39,7 |
|
Jour 17-18 |
||
|
Rythme respiratoire/minute |
34 |
107 |
|
Température rectale (C) |
38,8 |
39,7 |
Source: Fuentes et Chemineau, 1990.
Il semble donc que les races des zones tropicales sont mieux adaptées aux fortes températures que celles des régions tempérées.
La plupart des éleveurs d'ovins et de caprins savent que différents régimes alimentaires peuvent modifier les performances de reproduction de leurs animaux. Dans les zones tropicales ou subtropicales, la sous-alimentation est probablement un des facteurs principaux de l'environnement qui limite les performances de reproduction.
Activité de reproduction du mâle. La libido des mâles peut être sévèrement affectée par la sous-alimentation. Chez le bélier, celle-ci diminue à partir de cinq à 10 semaines après le début de la sous-alimentation et cet effet persiste tant que la sous-alimentation se poursuit. Une déficience à long terme en vitamine A conduit à une diminution de l'activité sexuelle chez le bélier; toutefois, cinq à six mois sont nécessaires avant que les symptômes ne se manifestent, à cause des réserves du foie en cette vitamine.
Une sous-alimentation sévère (400 g de poids vif en moins, par semaine, pendant 30 semaines) entraîne une diminution constante du poids testiculaire, de la concentration et du nombre total de spermatozoïdes de la semence éjaculée. Une corrélation très significative existe entre le poids testiculaire et le poids vif, mais également entre le poids testiculaire et la condition corporelle. Il existe, par conséquent, des corrélations significatives entre la production spermatique journalière (DSO) et la condition corporelle. Toutefois, si la sous-alimentation affecte sévèrement la quantité de sperme produite, elle ne semble pas modifier la qualité de celui-ci.
Des béliers maintenus à un régime d'entretien, alimentés avec un supplément riche en protéines, accroissent leur poids testiculaire. Chez des béliers mérinos, un supplément alimentaire avec de la graine de lupin pendant 15 semaines accroît le poids testiculaire de 66 pour cent; le poids vif augmente également (39 pour cent), mais relativement moins. Cet effet semble passer par l'intermédiaire d'une augmentation de l'activité de la LH chez les béliers suralimentés, plutôt que par une réponse accrue des testicules à la LH.
Dans la littérature il n'existe qu'un nombre assez limité de résultats concernant le niveau alimentaire et l'activité reproductrice des jeunes mâles. Il est connu, chez les ovins et caprins, comme dans les autres espèces, que la croissance testiculaire est étroitement corrélée avec la vitesse de croissance corporelle. Par conséquent, une sous-alimentation qui réduira la vitesse de croissance corporelle, produira un retard dans l'apparition de la puberté. Au contraire, des jeunes mâles soumis à un régime alimentaire de haut niveau, atteindront la puberté plus tôt et à un poids plus élevé que ceux soumis à un régime de bas niveau.
Finalement, il est également nécessaire de mentionner que des déficits sévères en certains éléments, comme les minéraux et les oligo-éléments, sont susceptibles d'affecter les performances reproductives des mâles.
Activité de reproduction de la femelle. La réponse, sur le plan reproduction, des femelles à des modifications du niveau alimentaire, peut être divisée en deux composantes: effets à long terme résultant d'une sous-alimentation subie à une période critique pendant le jeune âge, qui se manifeste à l'âge adulte, même si une alimentation correcte est distribuée plus tard et, effets à court terme et directs que l'on peut attribuer à des modifications transitoires des nutriments disponibles.
Chez la brebis, la sous-alimentation appliquée pendant la première année de vie diminue le taux d'ovulation et le taux de naissances multiples durant la vie d'adulte. Il apparaît toutefois, qu'il est nécessaire d'appliquer un régime très sévère pour obtenir de tels effets.
Chez des brebis adultes, la sous-alimentation peut provoquer une suppression des œstrus avec une cessation des ovulations ou l'apparition d'ovulations silencieuses. Au contraire, un haut niveau alimentaire, avant et après la mise bas, réduit l'intervalle mise bas/première ovulation et permet d'obtenir plus de brebis cycliques et saillies précocement dans la saison de reproduction.
Chez les brebis à queue grasse des zones subtropicales, il est quelquefois difficile de distinguer entre les effets à long terme et à court terme puisque le gras stocké dans la queue est utilisé comme réserve en cas de sous- alimentation. Pour ce qui est de la brebis Barbarine en Tunisie, qui n'effectue qu'une seule mise bas par an, la reprise de l'activité sexuelle post-partum est retardée chez les femelles qui n'ont pas eu la possibilité de mettre du gras en réserve pendant la fin de gestation, alors que la sous-alimentation après l'agnelage accroît la fréquence des cycles ovulatoires de courte durée et d'ovulations silencieuses, par rapport aux brebis bien alimentées. Dans la même race, une sous-alimentation sévère au printemps diminue le pourcentage de femelles répondant à 1'«effet bélier», augmente le pourcentage de cycles induits de courte durée et diminue la fertilité et la prolificité des femelles qui répondent.
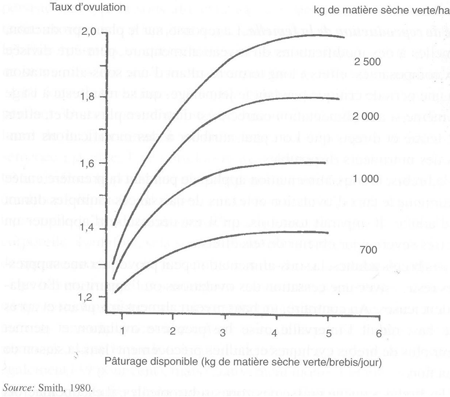
FIGURE 35 Relations entre le taux d'ovulation, les disponibilités fourragères et le contenu en matières sèches du fourrage chez des brebis maintenues au pâturage
Les effets d'une augmentation à court terme du niveau alimentaire sont bien connus. L'effet flushing est utilisé généralement quelques semaines avant la période de saillies; il produit une augmentation significative du taux d'ovulation et de la taille de la portée. Une augmentation du niveau alimentaire à deux fois le niveau d'entretien pendant seulement la durée d'un cycle œstrien, accroît le taux d'ovulation de 0,8 ovule, probablement en évitant l'atrésie des gros follicules dans les 30 dernières heures avant l'ovulation. Il semble, en fait, que le taux d'ovulation ne réponde à une modification du niveau énergétique à court terme que dans une fourchette intermédiaire spécifique de condition corporelle. En dehors de celle-ci, qui change selon le génotype, c'est la condition atteinte qui compte et il n'y a pas d'effet additionnel positif ou négatif de l'énergie ingérée.
Un cas particulier existe cependant avec 1'«effet graine de lupin», étudié essentiellement chez la brebis mérinos en Australie de l'Ouest. Une augmentation significative du taux d'ovulation (0,2 à 0,3 ovule par brebis ovulant) se produit dès six jours après le début de l'alimentation avec le lupin; cet effet est indépendant de tout accroissement mesurable du ,polds vif. Il semble dû au contenu exceptionnellement élevé en protéines des graines de lupin et à l'augmentation des substrats producteurs d'énergie.
La plupart des expériences conduites sur l'alimentation et la reproduction des mâles et des femelles sont faites en utilisant du fourrage conservé et du concentré ou avec une distribution de concentré au pâturage. Sans supplément au pâturage, les relations entre la production quantitative et qualitative de fourrage sont plus subtiles, parce qu'elles dépendent de la disponibilité fourragère et du poids d'animaux par unité de surface. Dans les conditions néo-zélandaises, lorsque la quantité de fourrage ingéré augmente, le taux d'ovulation s'accroît de façon curvilinéaire, pour plafonner à une offre située entre 3 et 4 kg de matière verte par brebis et par jour. Quand la quantité totale de fourrage augmente par hectare, le taux d'ovulation plafonne à une valeur plus élevée (figure 35). Pour le flushing, les pâturages avec dominance de trèfle (60 à 80 pour cent de trèfle blanc) sont supérieurs à ceux avec dominance de ray-grass (20 à 30 pour cent de trèfle blanc). Les taux d'ovulation les plus élevés sont obtenus après alimentation pendant six semaines, avec une disponibilité de 3,5 kg de matière verte par brebis et par jour, sur des pâturages offrant au moins 2 tonnes de matière sèche par hectare avant pâture, avec une utilisation de 40 pour cent. Ce niveau alimentaire provoque une croissance du poids vif supérieure à 1 kg par semaine.
Dans les descriptions faites du comportement sexuel mâle et femelle, les différentes interactions existant entre les sexes ont été décrites comme jouant un rôle important dans le déclenchement et le maintien du comportement sexuel dans les deux sexes. Toutefois, seuls les effets immédiats du contact avec un partenaire sur sa réponse comportementale ont été jusqu'ici considérés. Nous examinerons comment la présence permanente (ou l'absence) d'autres animaux du même sexe ou du sexe opposé sont susceptibles de modifier l'activité de reproduction du mâle à moyen ou long terme. Nous verrons également quelles sont les réponses des brebis et des chèvres à l'introduction des mâles après une période de séparation entre les sexes, soit 1'«effet mâle».
Isolement social du mâle reproducteur. Dans des conditions d'élevage intensif, la séparation des sexes s'effectue au sevrage. Dans les troupeaux laitiers ovins et caprins, celui-ci peut se produire soit 48 heures, soit 3 mois après la naissance. Dans l'espèce ovine, l'élevage des jeunes mâles en case individuelle, dans une bergerie séparée est très rare, sauf dans les petits troupeaux. Au contraire, les jeunes boucs sont souvent élevés séparément, particulièrement ceux qui vont subir les collectes de semence.
Dans l'espèce ovine, l'isolement partiel (boxes adjacents avec séparation ouverte) des jeunes mâles entre la naissance et 3-5 mois d'âge n'a pas de conséquences sur le comportement sexuel futur du bélier ou sa production spermatique, si des femelles sont présentes parmi les mâles de 3-5 mois jusqu'à la puberté. L'existence de montes mal orientées, observées sitôt après la réunion des mâles isolés depuis la naissance, n'est que transitoire (24 à 48 heures). Une telle isolation partielle n'est, en général, pas pratiquée pendant la période prépubère dans cette espèce, particulièrement grégaire.
L'isolement complet, depuis le très jeune âge jusqu'à la période prépubère (environ six mois) ferait craindre d'importantes perturbations du comportement sexuel (démarrage de l'activité copulatoire retardé ou efficacité sexuelle diminuée, voire même inhibition).
Dans l'espèce caprine, les jeunes mâles qui sont destinés à être testés sur descendance, sont classiquement élevés seuls dans des boxes individuels à partir du sevrage et sont conduits de la même façon pendant toute leur vie de reproducteur.
De telles conditions d'élevage, qui utilisent des équipements adaptés et ont des coûts d'entretien élevés, n'ont pas d'effet néfaste sur le comporte- ment sexuel du mâle. Elles ne modifient ni le moment du début de l'activité de saillie, ni l'efficacité sexuelle (définie comme le nombre de montes nécessaires pour effectuer une saillie ou éjaculer dans un vagin artificiel) et n'altèrent pas la production spermatique adulte quantitative ou qualitative. Ce mode d'élevage est préférable à l'élevage en groupe de mâles.
FIGURE 36 Début de l'activité sexuelle, au cours de tests de collecte de semence, de jeunes béliers Lacaune élevés dans trois conditions différentes pendant le jeune âge
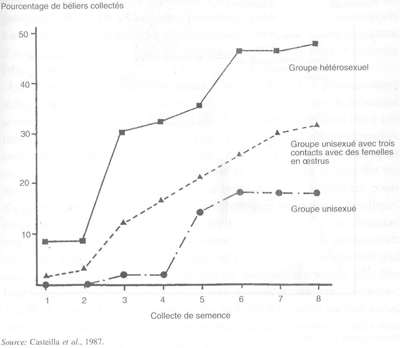
Présence de partenaires du même sexe: effets du groupe
Dans l'espèce ovine, si le sevrage n'est pas suivi de l'isolement social des jeunes mâles les uns des autres, il est habituellement suivi par la séparation des sexes et par l'élevage en groupes de même sexe. Cette séparation implique l'absence de contact avec les partenaires du sexe opposé jusqu'à la première utilisation en lutte, lors de la puberté ou plus tard. Cette privation de contact hétérosexuel pour environ trois mois, peut avoir des conséquences importantes sur l'activité sexuelle ultérieure des mâles. En effet, une proportion importante d'entre eux (quelquefois jusqu'à 70 pour cent) sont sexuellement inhibés, ce qui entraîne un retard important dans le démarrage de l'activité copulatoire ou du moment de collecte au vagin artificiel et une forte diminution de l'efficacité sexuelle. Ces perturbations peuvent limiter drastiquement l'efficacité d'un groupe de mâles dans un centre d'ia, voire même provoquer l'impossibilité totale d'utiliser des mâles intéressants sur le plan génétique. De tels problèmes réduisent par conséquent la vitesse du progrès génétique.
FIGURE 37 Début de l'activité sexuelle chez des béliers mérinos d'Arles de 18 mois élevés dans deux conditions différentes pendant le jeune âge
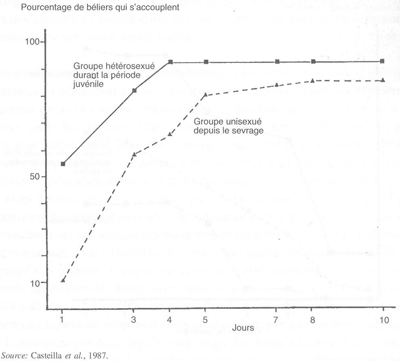
L'élevage des mâles en groupes unisexués entre l'âge de trois mois et la puberté, peut contribuer à l'apparition d'une activité homosexuelle dans les groupes de mâles. De tels animaux pourraient, par conséquent, avoir appris à associer leur comportement sexuel seulement avec des stimuli provenant d'autres mâles et à associer les montes avec l'impossibilité d'atteindre l'éjaculation.
Ultérieurement, les relations sociales interindividuelles peuvent aussi influencer fortement le comportement sexuel des mâles. L'efficacité sexuelle d'un mâle au sein d'un groupe est, en général, fortement liée à son rang social dans le groupe, en particulier si l'espace et le nombre de femelles sont limités. Dans certains cas, les mâles dominés refusent de s'accoupler en présence des mâles dominants, même si ces derniers ne peuvent les agresser (présence d'une barrière). Ces types de relations peuvent poser d'importants problèmes si les mâles dominés sont intéressants génétique- ment, ou si les mâles dominants sont stériles ou peu fertiles. En revanche, une stimulation mutuelle peut se produire dans certains cas, conduisant à une motivation sexuelle plus forte que si chaque mâle était présent tout seul avec un groupe de femelles. Ce moyen de stimulation sexuelle peut être bénéfique si la surface disponible est bien adaptée au nombre d'animaux. Toutefois, ces considérations n'ont qu'un intérêt restreint lorsque la semence est collectée au vagin artificiel, puisque les relations sociales sont alors limitées au moment de la collecte.
Dans l'espèce caprine, l'élevage de mâles en groupes de même sexe pendant la période prépubertaire est néfaste au futur comportement sexuel des boucs, particulièrement ceux qui seront collectés au vagin artificiel. Cet effet se traduit par un accroissement de la latence à l'éjaculation et la nécessité d'utiliser des stimuli artificiels, et généralement, par un retard du début de l'activité sexuelle; une inhibition complète peut même être observée chez quelques individus.
Les raisons de ces perturbations apparaissent identiques à celles présentées pour l'espèce ovine: essentiellement le développement d'une activité homosexuelle intense, conduisant à une inhibition lorsque les mâles sont placés dans des conditions hétérosexuelles. Il apparaît toutefois que, dans certains cas, les mâles les plus actifs du groupe sont aussi les plus actifs à servir le vagin artificiel; en revanche, ceux qui sont le plus soumis à des montes (c'est-à-dire ceux qui sont le moins homosexuellement actifs), sont les moins efficaces pour la collecte au vagin artificiel. Par ailleurs, les relations sociales à l'intérieur du groupe peuvent avoir des conséquences importantes. Sur ce plan, l'âge à la réunion est un facteur important qui peut modifier les réactions de peur, et qui peut avoir des conséquences sur le comportement lors de la collecte. Les boucs socialement dominants dans le groupe sont également ceux qui sont sexuellement les plus actifs.
Présence permanente de partenaires du sexe opposé
Pendant le jeune âge. Des privations sociales précoces (dans les trois premiers mois), telles que la ségrégation sexuelle, l'absence de la mère (dans le cas d'allaitement artificiel par exemple), et même l'isolement (dans le cas d'élevage en cases individuelles), n'ont pas de conséquences néfastes sur le comportement sexuel ultérieur des jeunes béliers, si elles ne sont pas poursuivies pendant la période prépubère. En effet, le rôle bénéfique d'un contact hétérosexuel pendant cette dernière période a été bien démontré. La présence postnatale de la mère renforce l'idée que l'environnement social favorable de la naissance à la puberté, est le suivant: présence de la mère et contact hétérosexuel prépubère.
Ce contact hétérosexuel a un effet particulièrement positif sur le début de l'activité copulatoire (figure 36) et sur l'efficacité des mâles, mais ne semble pas affecter la production spermatique. L'effet positif peut persister après un an chez des jeunes béliers, utilisés pour la première fois à 18 mois (figure 37).
Les modalités d'action des femelles sur le déclenchement du comporte- ment sexuel des jeunes mâles ne sont pas connues avec précision. Plusieurs expérimentations ont démontré que la présence de femelles ovariectomisées a un effet bénéfique même sur le comportement sexuel ultérieur des jeunes mâles. Cela indique que le rôle stimulant de la brebis est indépendant de son état physiologique ou de son comportement sexuel. Néanmoins, la présence de femelles sexuellement expérimentées apparaît préférable et l'induction d'une réceptivité sexuelle artificielle chez celles-ci (par traitement hormonal, par exemple) est une stimulation importante pour ces jeunes béliers et un moyen d'apprendre l'activité copulatoire.
TABLEAU 8
Effets des
conditions d'élevage et de l'âge au rassemblement 1 sur l'efficacité de la collecte de semence et sur la production spermatique chez de jeunes boucs alpins
|
Rassemblés |
Rassemblés tard |
Rassemblés |
Groupes |
Cases |
|
|
Pourcentage de collectes réussies/ 15 essais |
97 |
71 |
98 |
91 |
82 |
|
Latence à la 1re éjaculation (secondes) |
34 |
33 |
|||
|
Nombre de montes par éjaculation |
1,8 |
2,9 |
|||
|
Latence à l'éjaculation (secondes) |
42 |
62 |
|||
|
Volume de l'éjaculat (ml) |
0,8 |
0,6 |
0,9 |
0,8 |
1,0 |
|
Concentration (109 spz/ml) |
2,94 |
3,10 |
3,7 |
3,2 |
2,9 |
|
Nombre total de spz par éjaculation (109) |
2,23 |
1,79 |
3,33 |
2,56 |
2,90 |
|
Pourcentage d'éjaculats non retenus |
0,8 |
4,1 |
8,2 |
Source: Orgeur et al., 1988.
Dans l'espèce caprine, l'élevage des mâles en groupe et en ségrégation sexuelle est générateur de problèmes de comportement lors d'essais de collecte de semence au vagin artificiel.
La présence de femelles pendant la période prépubère, parmi des jeunes mâles destinés au testage sur descendance en ia, réduit ces inconvénients. De plus, une réunion précoce des jeunes mâles (avant l'âge d'un mois) est synonyme d'une moindre crainte interindividuelle et d'une tolérance plus grande qu' après une réunion tardive. Elle conduit à une meilleure efficacité à servir le vagin artificiel et à une meilleure qualité spermatique (tableau 8). Toutefois, l'élevage en boxes individuels reste préférable.
Dans cette espèce, connue pour être beaucoup moins grégaire que l'espèce ovine, la recherche du contact avec l'homme, lorsque les animaux sont élevés séparément, peut expliquer la facilité de collecte au vagin artificiel.
Dans les expérimentations décrites précédemment, la proportion de femelles était d'environ une pour quatre mâles dans l'espèce ovine et une pour quatre à 10 mâles dans l'espèce caprine.
Pendant la vie adulte, la présence permanente d'une femelle dans le groupe de mâles peut constituer une stimulation de leur activité sexuelle. Toutefois, cette stimulation tard dans la vie n'est pas indispensable si les mâles ont eu précédemment une expérience de saillie en lutte naturelle ou en récolte au vagin artificiel. Dans la plupart des cas, la présence d'une femelle pendant quelques semaines, dans un groupe de mâles, stimule les reproducteurs inhibés.
Les différentes observations présentées ci-dessus indiquent clairement que les relations interindividuelles, même avant la puberté, sont capables de modifier profondément l'expression et l'intensité du comportement sexuel du mâle. Les relations homme-animal ont également une grande importance pour l'utilisation des mâles des centres d'ia. Ces différents éléments permettent d'obtenir les meilleures conditions d'élevage des reproducteurs jeunes et adultes qui seront sélectionnés pour 1'ia.
Réponses physiologiques des femelles lors de la réintroduction des mâles après une période de séparation des sexes:l'«effet mâle». Ainsi que nous venons de le voir, la présence ou l'absence des femelles est susceptible de modifier le comportement sexuel des mâles. Toutefois, même si cette présence augmente la sécrétion de LH et de testostérone plasmatiques, il n'a pas été démontré qu'elle était susceptible de modifier l'activité spermatogénétique. En revanche, le mâle est capable, par sa seule présence parmi les femelles, de faire redémarrer leur activité ovulatoire et œstrienne. Un tel phénomène est appelé «effet mâle».
L'existence d'une distribution non uniforme des parturitions, au début de la saison sexuelle des femelles, est un phénomène qui a attiré l'attention des chercheurs depuis plusieurs dizaines d'années. Chez la brebis, il existe une relation très étroite entre la date d'introduction des mâles et le regroupement des agnelages, qui suggère que les saillies fécondantes se produisent aux alentours des jours 20-25 après l'introduction des béliers. Dans l'espèce caprine, deux pics de fécondations se produisent: le premier autour des jours 7-11 et le second autour des jours 27-35 après l'introduction des boucs.
FIGURE 38 Représentation schématique des réponses œstriennes et ovulatoires des brebis barbarines et des chèvres créoles à l'«effet mâle»
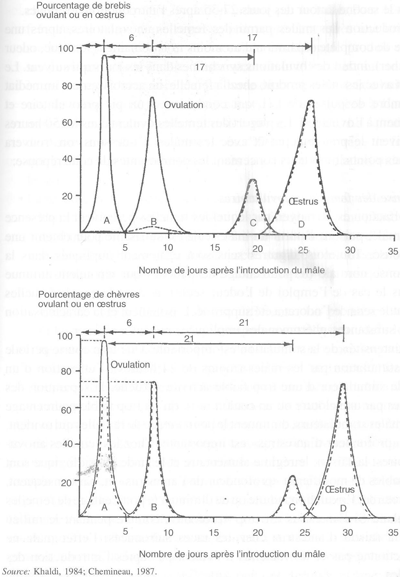
L'introduction des mâles parmi des femelles anovulatoires, après une période de complète séparation d'au moins trois semaines (son, vue, odeur et toucher), induit des ovulations synchrones dans les jours qui suivent. Le contact avec les mâles produit, chez la femelle, un accroissement immédiat du nombre de puises de LH, qui conduisent à un pic préovulatoire et finalement à l'ovulation. La plupart des femelles ovulent dans les 50 heures qui suivent le premier contact avec les mâles. Ci-dessous, on trouvera quelques points importants concernant les composantes de cette réponse:
Réponse des femelles anovulatoires
Evénements postovulatoires et comportement d'œstrus
- Chez la brebis, seules les ovulations précédées par une phase lutéale de durée normale sont associées à un comportement d'œstrus. La première ovulation n'est jamais accompagnée d'œstrus et la seconde ovulation qui suit un cycle de courte durée, est presque toujours «silencieuse». - Chez la chèvre, la situation est différente. La première ovulation induite peut être associée à un comportement d'œstrus (dans les deux tiers des cas chez la chèvre créole de Guadeloupe), mais les ovulations suivantes sont presque toujours associées à un œstrus. Dans cette espèce, le pourcentage de femelles en œstrus à la première ovulation induite diminue lorsque la «profondeur de l'anœstrus augmente».
Ces différentes observations conduisent à une meilleure compréhension des différents événements se produisant après l'introduction d'un mâle parmi un groupe de femelles anovulatoires (figure 38). Chez la brebis, la fertilité est correcte aux deux premiers pics de comportement d'œstrus qui se produisent 20 et 25 jours après l'introduction des béliers. Chez la chèvre, la fertilité est faible au premier œstrus à cause de la forte fréquence de cycles courts qui suivent la première ovulation induite. Comme la cyclicité est, en général, rétablie lorsque l'anœstrus n'est pas trop profond, les femelles luttées pendant 45 jours consécutifs ont une fertilité et une prolificité équivalant à celles des femelles déjà cycliques lors de l'introduction des boucs.
FIGURE 39 Deux exemples de l'évolution du poids testiculaire chez des béliers mérinos d'Arles au cours de leur première année d'âge
Le prétraitement des brebis et des chèvres avec un progestagène (progestérone ou FGA en une seule injection, ou sur une éponge vaginale) produit la disparition totale des cycles ovulatoires courts et augmente à près de 100 pour cent la fréquence des œstrus à la première ovulation induite. Ce traitement peut être utilisé pour induire l'œstrus chez des femelles qui doivent être inséminées artificiellement (voir Maîtrise de l'œstrus et de l'ovulation pour l'ia, chapitre 5).
L'«effet femelle». Beaucoup d'éleveurs de chèvres savent que l'apparition de l'œstrus chez quelques femelles du troupeau, peut provoquer le déclenchement d'un comportement sexuel, au moins chez quelques-unes des femelles restantes. Un tel «effet femelle» après synchronisation hormonale de quelques chèvres du troupeau a été décrit. Il n'a toutefois jamais été démontré clairement sur le plan expérimental. Chez la brebis, un effet d'entraînement des femelles en début de saison sexuelle,a été démontré.
Deux aspects différents des relations entre la santé des animaux et la production de sperme doivent être pris en considération.
Le premier concerne l'influence des maladies du reproducteur sur sa production spermatique ultérieure. Ainsi qu'il a été mentionné plus haut, une infection provoquant une augmentation de la température corporelle du mâle entraînera généralement l'apparition de spermatozoïdes anormaux dans sa semence, dans les semaines qui suivent l'infection, même si la température corporelle a diminué depuis plusieurs jours. Il est, par conséquent, vivement recommandé de suivre avec soin l'état sanitaire général de chaque reproducteur et de mesurer sa température rectale aussitôt qu'un doute survient. Quand celle-ci dépasse 39,5°C, on peut s'attendre que des spermatozoïdes anormaux apparaissent dans les semaines suivantes. Des infections peuvent également passer inaperçues par les personnes chargées des soins aux animaux; un examen régulier de la qualité de la semence (deux fois par mois, par exemple) est donc recommandé pour l'ensemble des reproducteurs, afin d'identifier ceux qui pourraient manifester une proportion élevée de spermatozoïdes anormaux dans leur semence.
Le second aspect à considérer concerne la possibilité qu'ont les reproducteurs mâles de transmettre des maladies infectieuses, par leur semence, aux femelles inséminées dans les troupeaux de production. Les agents infectieux de beaucoup de. maladies sont présents dans la semence au moment du maximum de l'infection, tels que ceux de la Blue-Tongue, du CAEV, de l'IBR-IPV, de la paratuberculose, de la listériose, de la brucellose, de la salmonellose, de la pasteurellose, de la toxoplasmose, de la leptospyrose, de la chlamydiose, de la mycoplasmose et certainement de beaucoup d'autres inconnues à ce jour. Trichomonas a également été identifié à plusieurs reprises dans la semence de bouc. Cela ne veut cependant pas dire que des animaux ayant été infectés, mais ne se trouvant plus au pic de l'infection, continuent d'excréter des germes dans leur semence. Le seul moyen sûr de ne pas transmettre d'agent infectieux par la semence reste toutefois, celui de s'assurer que les mâles producteurs sont indemnes de toute maladie contagieuse. Il est, par conséquent, très important de maintenir le troupeau de reproducteurs strictement hors du contact des animaux susceptibles d'être infectés. De plus, des contrôles sérologiques de routine doivent être effectués régulièrement pour les maladies existant dans l'aire de production (voir Bâtiments, logement et conduite des animaux, chapitre 3).
La puberté peut être définie comme l'âge et le poids auxquels les animaux sont capables de se reproduire: dans le cas des mâles lorsqu'ils sont capables de féconder une femelle après saillie (puberté mâle) et dans le cas des femelles, lorsqu'elles sont fécondées lors de l'œstrus et capables de conduire une gestation jusqu'à son terme (puberté femelle). Dans les deux sexes, la puberté est, en général, précédée d'une période de quelques semaines, appelée période prépubère, pendant laquelle une stimulation externe peut provoquer l'apparition de la puberté.
Puberté chez le mâle. Dans les deux espèces considérées, le poids testi-culaire à la naissance varie de 2 à 30 g et est constitué de tubes séminifères contenant les cellules de soutien (qui se développeront plus tard en cellules de Sertoli) et de gonocytes (d'où partiront les divisions spermatogoniales). Après la période impubère (qui dure de quelques semaines à quelques mois selon les races, la saison de naissance et le régime alimentaire), caractérisée par une augmentation lente du poids testiculaire, les premiers cycles spermatogénétiques débutent pendant la phase de croissance rapide du testicule. Les premiers spermatozoïdes sont libérés dans la lumière des tubes pendant la phase de croissance rapide et la puberté (première éjaculation) est atteinte durant la fin de cette phase. La taille testiculaire peut être utilisée, intrarace, comme un prédicteur précis du début de la puberté. Après quoi, le testicule commence une troisième phase d'évolution, la deuxième période de croissance lente de son poids. Le poids testiculaire maximum est atteint pendant la deuxième année de vie et dépend du régime alimentaire et de la saison chez les races photopériodiques. Deux exemples d'évolution du poids testiculaire au cours de la première année de vie sont représentés à la figure 39.
Le développement anatomique des organes d'évacuation dépend directement de la sécrétion de testostérone par le testicule. Chez le mâle immature, le gland du pénis et l'appendice filiforme sont complètement adhérents au prépuce. Avec la croissance corporelle, un détachement progressif des adhésions est observé et le pénis devient libre, l'appendice filiforme en premier, puis le gland. Lorsque les testicules sont d'une taille développée, le pénis est libre dans le prépuce et peut sortir de celui-ci.
Les premières manifestations de comportement sexuel, incluant des montes orientées préférentiellement vers les femelles, apparaissent dès l'âge de quelques jours chez les jeunes mâles. Toutefois, cette activité de «jeu sexuel» n'a aucun rapport avec la puberté qui se manifeste vers l'âge de quatre à six semaines, ni avec le futur comportement sexuel du reproducteur adulte. Les premiers signes de la puberté (achèvement de la spermatogénèse, présence de spermatozoïdes dans l'épididyme) apparaissent généralement à environ 30 à 40 pour cent du poids adulte, mais la puberté «comportementale» (séquences organisées conduisant à des accouplements, au lieu du «jeu sexuel» de l'enfance) n'est atteinte que vers 40 à 50 pour cent du poids adulte.
Une fois que la puberté «comportementale» s'est manifestée, la semence peut être collectée à l'aide d'un vagin artificiel. Les premiers éjaculats collectés sont, toutefois, loins d'être normaux et ne sont en général pas utilisables pour l'ia. La concentration spermatique est faible, beaucoup de spermatozoïdes sont morts et/ou anormaux, et la motilité des quelques cellules vivantes qui restent est faible. Il est alors nécessaire d'attendre quelques semaines supplémentaires (trois à 10) jusqu'à ce qu'un accroissement de la quantité et de la qualité des spermatozoïdes éjaculés soit observé.
Il existe une variation importante entre races, quant à l'âge et le poids vif auxquels la puberté est atteinte.
Puberté chez la femelle
Espèce ovine. Chez l'agnelle, une première ovulation silencieuse suivie par un corps jaune de courte durée se produit généralement à la puberté. Le premier comportement d'œstrus n'est observé que lorsqu'il y a eu, au préalable, une phase lutéale de durée normale. Par conséquent, les événements observés chez l'agnelle se produisent, habituellement, dans l'ordre suivant: première ovulation silencieuse suivie d'un corps jaune de courte durée; deuxième ovulation silencieuse suivie d'un corps jaune de durée normale; et troisième ovulation associée à un comportement d'œstrus et suivie d'un corps jaune de durée normale.
Pendant la première saison sexuelle de la femelle, la durée moyenne des œstrus est plus courte que celle des brebis adultes, ce qui peut sans doute être attribué à la fois à la plus faible sensibilité du système nerveux central à l'œstradiol 17B\?\ et à la plus faible sécrétion de celui-ci par l'ovaire. Cette diminution de la durée du comportement d'œstrus est probablement responsable, partiellement en lutte naturelle de la plus faible fertilité des agnelles par rapport à celle des adultes.
Espèce caprine. Dans cette espèce, une situation différente est souvent observée: un ou plusieurs œstrus peuvent apparaître sans ovulation associée, puis suivent œstrus et ovulation. En effet, puisque la chèvre, au contraire de la brebis, n'a pas besoin de recevoir une imprégnation par la progestérone avant l'œstradiol 17B\?\ pour le déclenchement du comportement d'œstrus, la sécrétion précoce d'œstradiol 17B\?\ provenant des premiers follicules en développement suffit pour provoquer l'apparition de l'œstrus. Il est possible toutefois, les relations entre ovaire et système nerveux central n'étant pas complètement établies, que l'ovulation ne se produise pas après la première vague de croissance folliculaire.
Dans les deux espèces, le début de la puberté dépend à la fois de l'âge et du poids vif. Les femelles sont pubères à un âge défini, seulement si elles ont atteint un développement corporel suffisant. De la même façon, les femelles deviennent pubères à un poids vif défini et à condition qu'elles aient un certain âge. Le poids vif auquel la puberté est atteinte est, en général, 40 à 60 pour cent du poids adulte, l'âge se situant alors entre cinq et 18 mois. Chez les races saisonnées, les animaux ne deviennent pubères que pendant la saison sexuelle et, par conséquent, l'âge et le poids à la puberté dépendent étroitement de la date de naissance dans l'année. Dans ces races, les femelles nées en hiver/début du printemps atteindront la puberté à l'automne/hiver suivants, uniquement si elles ont un développement corporel suffisant (c'est-à-dire si elles ont été alimentées correctement), sinon, elles devront attendre jusqu'à la saison sexuelle suivante et n'atteindront la puberté qu'à 18 mois. Le faible développement du tractus génital chez les animaux nullipares peut expliquer les résultats de fertilité peu satisfaisants lorsque les femelles sont mises en lutte trop tôt.
Pour ces raisons il n'est pas recommandé de mettre à la reproduction les jeunes femelles avant qu'elles n'aient atteint 50 à 60 pour cent du poids vif des femelles adultes.
Dans les deux espèces, la mise bas est suivie d'une période de repos sexuel pour deux raisons d'origine interne. La première est le temps nécessaire à l'involution utérine. La seconde est l'inactivité de l'ovaire, essentiellement d'origine centrale puisque celui-ci n'est pas suffisamment stimulé par les hormones gonadotropes.
Des stimulations externes peuvent également retarder la reprise de l'activité sexuelle post-partum, comme la présence des jeunes, la lactation, le niveau alimentaire prépartum et post-partum et le moment de l'année où a lieu la mise bas dans les races saisonnées. Dans ce dernier cas, comme pour la puberté, il existe une relation étroite entre la date de parturition et l'intervalle qui sépare celle-ci de la première ovulation ou du premier œstrus. Lorsque la mise bas a lieu quelques semaines avant ou pendant la première moitié de la saison sexuelle, le premier œstrus et/ou la première ovulation se produisent rapidement (30 à 60 jours plus tard); en revanche, les femelles qui agnèlent ou chevrottent pendant la deuxième moitié de la saison sexuelle ou pendant la saison d'anœstrus, attendent la saison sexuelle suivante pour reprendre leur activité sexuelle post-partum.
L'interaction entre durée de l'anœstrus post-partum et fertilité à l'ia est importante si les femelles sont synchronisées par traitement hormonal. La fertilité des femelles allaitantes ou en lactation, peu de temps après la parturition, est, en effet, toujours plus faible que celle des femelles sèches (voir Paramètres susceptibles de modifier les résultats d'ia, chapitre 6).
L'intérêt pour l'amélioration génétique des performances de reproduction est la plupart du temps centré sur la taille de portée, la puberté, l'intervalle post-partum, la durée de la saison sexuelle chez la femelle et les composantes de la production spermatique chez le mâle. Ces caractères, partiellement sous contrôle génétique, présentent une variabilité entre races et également intrarace. Il y a donc au moins deux méthodes potentiellement disponibles pour les améliorer: utiliser les ressources raciales pour le croisement et le développement d'une nouvelle lignée ou sélectionner en race pure. Une mention spéciale doit être faite pour la prolifïcité qui peut être contrôlée par un gène majeur (comme le gène «F» Booroola). L'introduction de ce gène dans une race par des croisements répétés avec des animaux porteurs est un autre moyen d'améliorer la prolificité.
Taille de portée. C'est un des caractères de reproduction qui varie d'une race à l'autre. Dans l'espèce ovine, les brebis mérinos, Charmoise (France), Suffolk (Royaume-Uni), Barbarine (Tunisie), Serres (Grèce) sont des exemples de races caractérisées par une prolificité relativement basse (entre 1,0 et 1,5 agneau par mise bas), quelles que soient les conditions d'élevage. Au contraire, chez la brebis Romanov (Europe), la brebis Chios (Grèce), la brebis D'Man (Maroc), la brebis Finnoise (Europe du Nord), connues comme des races prolifiques, la taille moyenne de portée est en général comprise entre 2,5 et 3,5 agneaux par mise bas. Quelques femelles donnent naissance à des quadruplés ou des quintuplés, quelquefois plus.
Entre ces deux groupes extrêmes de brebis, les prolifiques et les non prolifiques, sont situés tous les autres génotypes qui donnent naissance en moyenne à 1,5-2,5 agneaux par mise bas.
Dans l'espèce caprine il semble exister un plus faible nombre de races prolifiques et la taille de portée des races prolifiques apparaît plus faible que celle des brebis. La plupart des races de chèvres ont une prolificité comprise entre 1,5 et 2,5 chevreaux par mise bas. Les races non saisonnées ont tendance à avoir une taille de portée supérieure aux races saisonnées. Les composantes de la prolificité (taux d'ovulation et mortalité embryonnaire) n'ont pas été étudiées de façon aussi approfondie que dans l'espèce ovine.
La répétabilité de la taille de portée est faible; ce caractère est modifié par l'environnement et, comme d'autres caractères qui lui sont proches, il a une faible héritabilité. D'une manière générale, les estimations de la répétabilité de la taille de portée chez la brebis sont voisines de 0,10. Toutefois, la variabilité phénotypique et le coefficient de variation sont relativement élevés, ce qui permet un taux annuel de réponse à la sélection comparable à d'autres caractères à forte héritabilité, comme le poids au sevrage. De plus, l'analyse des expériences de sélection en ferme démontre clairement que la sélection sur la prolificité est un objectif qui en vaut la peine.
A noter toutefois que la synchronisation hormonale de l'œstrus ne fait pas que modifier l'expression de la taille de portée. Les prolificités induite et naturelle sont deux caractères génétiques différents: l'héritabilité est un peu plus forte pour la prolificité induite et la corrélation génétique entre ces caractères est seulement de 0,40. Paradoxalement, ces dernières considérations empêchent les éleveurs qui veulent sélectionner directement sur la taille de la portée, d'utiliser la synchronisation hormonale de l'œstrus.
Les variations de taille de portée entre les races dépendent du taux d'ovulation, du taux de fécondation et de la mortalité embryonnaire. De nombreuses études montrent que le taux d'ovulation est la composante principale de la taille de la portée. En effet, celui-ci représente la limite maximale de la prolificité; par ailleurs, les races prolifiques ont des taux d'ovulation élevés (plus de sept ovulations peuvent être observées). De plus, sa répétabilité et son héritabilité sont supérieures aux valeurs correspondantes pour la taille de la portée. Toutefois, aucun rapport n'a été publié, à notre connaissance, où une sélection sur le taux d'ovulation a entraîné une augmentation de la taille de la portée. La diminution de survie embryonnaire pour chaque corps jaune supplémentaire conduit au fait que la taille de portée maximale est située pour des taux d'ovulation intermédiaires. De surcroît, dans certaines situations, une variation génétique de survie embryonnaire contribue de façon importante à la prolificité; ainsi, les brebis Romanov ont une meilleure survie embryonnaire que les brebis finnoises.
Découvert récemment en Australie, le gène mérinos Booroola (appelé aussi gène F, comme fécondité), est un gène à effet majeur qui agit sur le taux d'ovulation de la femelle. Les femelles porteuses homozygotes, qui ont deux copies du gène (FF), ont un taux d'ovulation moyen de 4,7 et une taille de portée de 2,8; les hétérozygotes porteuses, qui ont seulement une copie du gène (F+), ont un taux d'ovulation de 2,9 et une taille de portée de 2,4 et finalement, les homozygotes non porteuses (dans la race mérinos) qui n'ont pas le gène (++), ont un taux d'ovulation et une taille de portée d'environ 1,3. Une telle découverte a de profondes implications sur la stratégie utilisée pour augmenter la fécondité. La présence d'un gène à effet majeur, identique, dans d'autres races de brebis est également suspectée. Chez le mâle, le gène F n'est apparemment pas exprimé et le seul moyen, pour le moment, d'identifier les béliers porteurs est d'organiser un testage sur descendance.
Puberté et post-partum. Ainsi que nous l'avons vu précédemment, le début de la puberté ou l'âge au premier œstrus et le début de la saison sexuelle, chez les animaux saisonnés, sont étroitement liés. L'activité sexuelle des agnelles peut donc être un bon critère de sélection pour le saisonnement de l'adulte.
Entre races, l'âge à la puberté présente d'importantes variations: ainsi, en France, 88 pour cent des primipares Romanov mettent bas avant l'âge de 24 mois, par rapport à seulement 40 pour cent des Berrichonnes.
L'activité sexuelle post-partum hors-saison présente également un déterminisme génétique chez les races saisonnées et il existe une forte variabilité inter et intrarace dans la durée de l'anœstrus post-partum, ce qui pourrait représenter un caractère à sélectionner.
Ainsi que cela a été dit, il existe une forte variabilité entre races dans la durée de la saison sexuelle. Chez les races saisonnées, il y a une forte relation (corrélation phénotypique de 0,36 à 0,80) entre la date du premier œstrus et la durée de la saison sexuelle ou le nombre d'œstrus par femelle. De plus, la date de premier œstrus est répétable et héritable. La sélection sur ce caractère se fait indirectement par les modes de conduite ou les rythmes de reproduction accélérés.
L'introduction dans les pays tempérés, de races non saisonnées venant des milieux tropicaux ou subtropicaux et conduits en race pure ou en croisement avec les races locales, pourrait être d'un intérêt certain pour accroître la fertilité à contre-saison. Néanmoins, les résultats sont toujours limités dans ce secteur et la fertilité hors-saison n' est pas perçue comme un caractère limitant. Dans ces circonstances, la sélection intrarace reste la plus intéressante.
Comportement sexuel. Le comportement sexuel est un caractère important et complexe à considérer, bien qu'il n'existe que peu d'études sur les bases génétiques de celui-ci. Des différences raciales dans le nombre de saillies par mâle (plus faible en Mérinos qu'en Finnois) ainsi que dans le temps de latence avant la collecte (les Texel réagissent plus vite que les Dorset) ont été rapportées. D'importantes variations intrarace existent également pour ce caractère qui présente, en général, une bonne répétabilité. Des héritabilités voisines de 0,30 ont été rapportées pour la capacité de saillie des béliers en lutte naturelle.
Qualité de la semence. Des différences raciales ont été mises en évidence pour la plupart des caractéristiques spermatiques (volume et concentration en spermatozoïdes de l'éjaculat, anomalies spermatiques, pourcentage de cellules vivantes) et pour la production spermatique quotidienne. Par exemple, les béliers Ile-de-France ont une production spermatique plus élevée que les béliers Romanov. Ce résultat n'est pas surprenant puisque les spermatozoïdes sont issus des spermatides rondes et que les béliers Ile-de-France ont, à 13 mois, plus de spermatides rondes que les béliers Romanov.
Il existe aussi des variations importantes entre mâles dans le pourcentage de spermatozoïdes anormaux. Au printemps, dans les races saisonnées d'ovins, quelques mâles produisent un pourcentage élevé de spermatozoïdes anormaux (proche de 100 pour cent), alors que d'autres restent à de faibles valeurs (moins de 15 pour cent). Toutefois, ces différences n'existent plus chez les mêmes mâles quand ils sont collectés pendant la saison sexuelle, à l'automne. Cette caractéristique de «photosensibilité» est répétable pour les mêmes mâles d'une année sur l'autre, et la sélection d'animaux produisant une semence de bonne qualité au printemps est déjà effectuée sur cette base dans plusieurs centres d'ia. L'héritabilité de ce caractère semble assez élevée (0,42) alors qu'elle est faible pour le pourcentage de spermatozoïdes vivants.
Des variations individuelles dans le volume et la concentration spermatique de l'éjaculat sont également rapportées; l'héritabilité du volume est élevée (0,43) mais celle de la concentration spermatique est faible (0,07).
Taille testiculaire. Beaucoup d'études ont porté sur les variations génétiques entre races concernant le développement testiculaire. Les races prolifiques d'ovins (Romanov, Finnoise, etc.) ont tendance à avoir un développement testiculaire plus précoce et plus rapide que les races non prolifiques (Ile-de-France, Dorset), mais les poids testiculaires adultes sont généralement plus élevés chez ces derniers. En dépit des effets marqués de la saison et du poids vif sur le poids testiculaire avant la puberté, les héritabilités sont assez élevées (0,15 à 0,50).
Le sens des corrélations entre les différents paramètres de reproduction ne sont pas stricts d'une race à l'autre. Le saisonnement n'est pas toujours associé à la taille de portée de la même façon: la brebis Peule (Niger) n'est pas saisonnée et de prolifïcité moyenne, la D'Man (Maroc) n'est pas saisonnée et à forte prolificité, l'Ile-de-France est saisonnée et de faible prolificité et, finalement, la Romanov est saisonnée et très prolifique.
Les caractéristiques de reproduction de ces différentes races sont généralement décrites dans leurs conditions d'élevage traditionnelles ou dans des conditions proches de la «normale» pour la race considérée. Toutefois, l'expression de ces caractères, même s'ils sont d'origine génétique, est susceptible de changer si les animaux sont transférés dans des conditions d'environnement très différentes. La forte taille de portée des races prolifiques, par exemple, peut être réduite considérablement si les femelles sont sous-alimentées ou soumises à un stress thermique (voir Effets de l'environnement thermique sur l'activité de reproduction, chapitre 2). Au contraire du taux d'ovulation et de la taille de portée, le nombre de cycles sexuels des races saisonnées n'est pas profondément affecté quand les animaux sont maintenus sous un régime alimentaire restrictif. Quand des animaux saisonnés sont transférés dans de basses latitudes, le saisonnement persiste ce qui indique que la commande de ce caractère est essentiellement d'origine génétique, bien que certaines perturbations des cycles soient observées.
La prolificité ne peut être mesurée que chez la femelle; la sélection sur ce caractère serait beaucoup plus efficace si un caractère corrélé, mesuré chez le mâle, pouvait être utilisé. Bien que plusieurs relations entre caractères mâle et femelle aient été analysées dans ce but, il n'existe pas de résultat net indiquant qu'une sélection sur un caractère mâle puisse produire un accroissement de la taille de portée. Il est rapporté toutefois, que la sélection sur la taille testiculaire, ajustée au poids vif, conduit à la sélection d'une puberté plus précoce chez la femelle. Le taux d'ovulation est également augmenté à poids vif constant, mais pas en valeur absolue.
Une remarque particulière doit être faite, dans l'espèce caprine, concernant le caractère motte (sans cornes) et ses relations avec l'intersexualité. L'absence de cornes est due à un gène autosomal dominant «P». Par conséquent, trois génotypes et deux phénotypes sont rencontrés: les homozygotes PP et les hétérozygotes Pp qui sont mottes et les homozygotes récessifs pp qui sont cornus. A l'état homozygote, le gène P entraîne la masculinisation de toutes les femelles PP qui peuvent être mal formées à divers degrés, ce qui conduit à un pseudo-hermaphroditisme. Puisque toutes les femelles homozygotes mottes PP sont stériles, toutes les femelles qui se reproduisent sont soit mottes hétérozygotes Pp ou cornues pp. Tous les mâles cornus pp sont fertiles et les mâles mottes hétérozygotes Pp sont également fertiles. Au contraire, plus de la moitié des mâles mottes PP sont partiellement ou complètement stériles. Finalement, les mâles destinés à la reproduction doivent donc être choisis de préférence cornus et tous les mâles mottes issus de deux parents mottes doivent être écartés.
Deuxième partie
Insémination artificielle des brebis
et des chèvres
![]()
![]()
![]()