Control of verticillium wilt and fusarium wilt diseases by soil solarization in Southern Spain
R. M. Jimenez-Diaz,1,3, J. Bejarano², M. A. Blanco³, J. Gomez², R. Gonzalez², and J. M. Melero²
1 Instituto de Agronomía y Protección Vegetal, CSIC, Apdo. 3048,14080 Córdoba, Spain
2 Departamento de Protección Vegetal, DGIEA, Apdo. 240, 14080 Córdoba, Spain
3 Departamento de Agronomía, ETSIA, Universidad de Córdoba, Apdo. 3048, 14080 Córdoba, Spain
Introduction
Verticillium wilt induced by Verticillium dahliae Kleb. and Fusarium wilt induced by Fusarium oxysporum-Schlecht f. sp. niveum (E. F. Smith) Snyd & Hans., are the most important diseases of cotton (Gossypium hirsutum L.) and watermelon [Citrullus lanatus (Thumb.) Matsum et Nakai], respectively, in southern Spain (2,5). Recently, devastating epidemics of Verticillium wilt have occurred in the lower Guadalquivir Valley (Marismas) of that region as a result of high inoculum densities (ID) in the soil and/or the widespread occurrence of a defoliating pathotype of the pathogen (2).
Similarly, Fusarium wilt of watermelon is threatening the greenhouse crop industry in Almeria, where intense watermelon cultivation in sand-mulched soils has lead to increased ID values of the pathogen and wilt incidence. In 1985, disease survey in greenhouse crops in Almeria indicated that 54 percent of the surveyed watermelons were affected and that the highly virulent race 2 of the pathogen is widespread in that area (5).
Control of those Fusarium and Verticillium wilt diseases is mainly by the use of resistant or tolerant cultivars. However, other control measures are needed when resistance or tolerance are unavailable or unsuitable. Soil solarization has proven effective against V. dahliae and F. oxysporum f. sp. niveum in several countries (6, 7, 8, 9, 10), and it seems more convenient than soil fumigation with methyl bromide because of toxicity, recolonization problems and economic cost.
This paper presents an overview of our studies on the use of soil solarization to control Verticillium wilt of cotton and Fusarium wilt of watermelon in southern Spain, as well as on the beneficial effect of that practice on rose planting-stock production in the absence of major pathogens.
Verticillium Wilt of Cotton
Soil solarization experiments were conducted in fields naturally infested with the defoliating pathotype of V. dahliae at the Marismas area of southern Spain from 1986-88. Two (A and B) and three fields (C-E) with a range of ED values were selected for experiments in 1986 and 1987, respectively. Inoculum density in the experimental plots was assessed by the Butterfield and DeVay method (4) in bulked samples of the upper 20-cm soil layer, at the time of tarping the soil with polyethlene sheets (PE), just after plastic removal, and at the seedling stage of the next cotton crop the following spring.
In 1986, 12 plots at each of fields A and B were irrigated in early July, of which six were solarized and six served as unsolarized controls in a randomized complete block design. Plots were solarized from mid-July through the end of August. In field A, plots were 6 x 10 m and PE sheets used were 37.5 ?m thick. Plots in field B were 3.5 x 10 m and PE sheets used were 25.0 ?m thick. Solarized and unsolarized plots were sown to cotton cultivar Coker 310, susceptible to V. dahliae and widely grown in the area, in early spring 1987. Disease incidence (percentage of plants with foliar symptoms) was assessed at the end of July and early September, and seed-cotton yield was determined in field B. for which plots were harvested by handpicking.
Soil solarization in the summer of 1986 brought V. dahliae population to negligible levels in fields A and B (Table 1). Furthermore, ID values in solarized plots remained close to 0 propagules/g dry soil (p/g) shortly after planting time in 1987, whereas they had increased in the unsolarized control (Tables 1, 2). Disease incidence in unsolarized controls was fairly high in both fields, but it was very low in solarized plots when assessed in July 28 (Table 2) and it reached 22 percent in field A on September 9. The beneficial effects of soil solarization were further indicated in field B. since seed cotton yield in solarized plots was increased by 131 percent as compared to unsolarized ones, where severe disease developed even with low ID values (Table 2).
In 1987, plots (6.5 x 15 m or 6.5 x 30 m) at each of fields C-E were either solarized or not in replicated (twice) experiments with a randomized complete block design. Plots were irrigated in mid-July and tarped with PE sheets 37.5 um thick three days later. The PE sheets were kept in position through the end of September to cope with unusual mild temperatures, cloudy periods and rainfall.
Soil solarization eradicated V. dahliae from soils with a range of ID values to undetectable ID levels (Table 1). ID values in solarized soils remained at low levels both in the 0-20 cm and 2040 cm soil layers shortly after planting time in l988 (Table 2). Furthermore, plant stand in solarized plots increased by 21-108 percent as compared to unsolarized ones, where infections by Rhizoctonia solani caused damping-off. Disease incidence in solarized plots of fields C and D was significantly reduced with respect to unsolarized controls, for which high incidence was correlated with ID values (Table 2). Similarly, seed cotton yield increased significantly after solarization, and to a larger extent in the most severely affected field C (110 percent) than in the less affected field D (33 percent) (Table 2).
Fusarium Wilt of Watermelon
Two preliminary solarization experiments were carried out in commercial plastic houses with a history of severe Fusarium wilt of watermelon at Almeria, southern Spain, in 1985 and 1986.
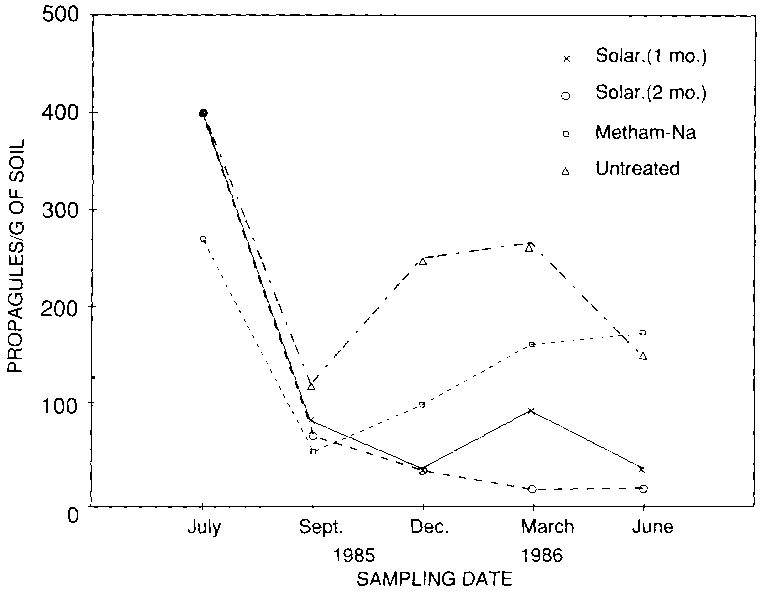
In 1985, plots were fumigated with Metham-Na (Vapam at a rate of 1 200 I/ha), or solarized for one (mid-July - mid-August) or two (mid-July - mid-September) months. An untreated plot served as control. The experiment was replicated four times in a latin square design. Soil at a depth of 10-15 cm was sampled just before and after treatments, and periodically thereafter, and assayed for their density of viable propagules of F. oxysporum (FO) by dilution plating onto a semi-selective medium (3, 5). Plots were planted to watermelon, cultivar Resistant, in mid-December and disease incidence was assessed periodically.
Density of viable (FO) propagules in control and treated soils had declined, respectively, to about 19 percent and 30 percent of their initial populations by September 1985, and it further decreased in solarized soils to stabilize at about 8 percent of the initial population after December (Fig. 1). On the contrary, FO populations in control and fumigated soils fluctuated during the period of study. Final disease incidence in plots solarized for one and two months averaged 28 percent and 25 percent, respectively, as compared to 41 percent for fumigated plots and 47 percent for the untreated controls.
A non-replicated trial was conducted in two plastic houses in the summer of 1986. Soil was solarized for 45 days in one plastic house and fumigated with DD+methyl-isothiocyanate (Ditrapex at the rate of 500 1/ha) in the other. Densities of viable FO propagules in the 10-15 cm soil layer were reduced by 64 percent and 40 percent after solarization and fumigation, respectively. Pathogenicity tests carried out with 100 FO isolates recovered from soil prior to and after treatment indicated that some 85 percent of the FO population surviving each treatment was pathogenic to watermelon cultivar Sugar Baby. Prior to treatment, the frequency of pathogenic FO isolates was 58 percent and 84 percent in soil to be solarized or fumigated, respectively.
An additional experiment was carried out in a plastic house at Almeria in 1988, with the soil artificially infested with F. oxysporum f. sp. niveum. Inoculum consisted of conidia of the pathogen mixed with talcum powder. The infested talcum was spread onto soil strips, then covered with a 10-cm sand layer where seed are placed at sowing. Following soil infestation, watermelon cultivar Sugar Baby was planted onto the infested strips to increase and homogenize the pathogen population in soil.
The experiment consisted of soil solarization for one (June 28-July 28) or two (June 28-August 28) months and fumigation with Metham-Na (Vapam at a rate of 1 200 1/ha in early July). Untreated infested or uninfested plots served as controls. There were four replicated 24-m² plots with a randomized complete block design. Soil was drip irrigated then tarped with PE sheets 4 m wide and 37.5 m m thick. Soil temperature at 10- and 20-cm depth was recorded continuously in solarized and unsolarzied plots, and averaged for each hour. Density of viable FO propagules in treated and untreated soil was assessed as before, two months before solarization and at three-month intervals from late September 1988 through early July 1989.
Plots were sown to watermelon cultivar Sugar Baby in the beginning of April 1989, following a winter crop of green beans (Phaseolus vulgaris L.). Incidence of wilted plants was recorded at weekly intervals from 17 to 38 days after planting. The influence of treatments on crop production was assessed by the biomass of non-symptomatic plants sampled when thinning the plots 43 days after planting, as well as by the number and yield of fruits harvested per plot.
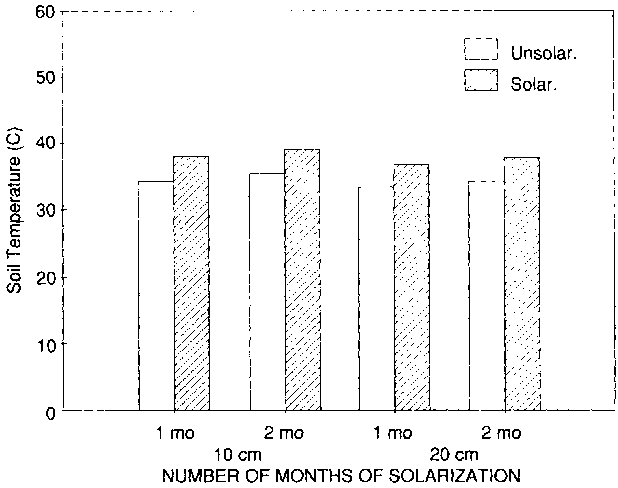
Mean temperatures at 10 and 20 cm soil depths increased 4°C and 3.3°C, respectively, during the first month of solarization and 3.7°C and 3.5°C, respectively, over the two-month solarization period (Fig. 2). Hourly temperatures varied within a range of 26.3° - 47.8°C and 28.9° - 43.3°C at 10and 20-cm soil depth, respectively, during the two months of solarization; and within the range of 25.5° - 41.6°C and 26.8° - 38.8°C during the same period in the unsolarized soil. The rather low temperature increments in solarized soil, as compared to unsolarized soil, might be due to solarization within a plastic house closed environment. Although the temperature increase was low, the population of FO in soil was significantly reduced after solarization for two months, and to a lesser extent by fumigation and one month's solarization (Table 3).
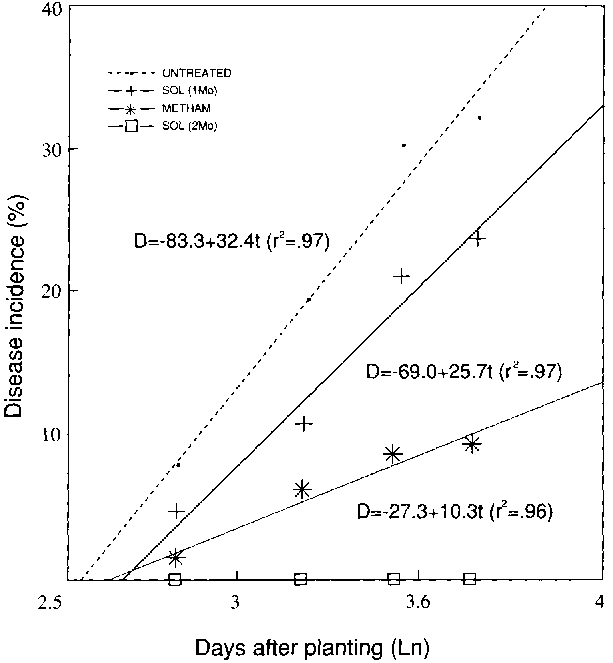
Disease progress curves in treated and untreated plots were linearized by log transformation of time after planting and they are shown in Figure 3. Rate of disease progress in plots solarized for two months was nil. In comparison, solarization for one month and fumigation with Metham-Na reduced the rating of disease progress to 79 percent and 32 percent, respectively, of that in untreated plots.
Soil solarization for two months showed the highest increases in plant dry weight, and watermelon fruit number and yield, followed by Metham-Na and one month's solarization (Table 4). While plant dry weight, and number of watermelon fruit in plots solarized for two months were significantly higher than those in the uninfested control, differences among uninfested control, solarization for one month and fumigation were not significant.
Rose Planting-Stock Production
A soil solarization experiment was conducted during 1987- 1989, on a field which had been continuously cropped to rose [Rosa indica var. major] for the last eight years near Sevilla, southern Spain. The soil in that field is a silty loam with 0.8 percent of organic matter. Soil analyses before treatments indicated that no major pathogens were present at detectable levels.
The experiment consisted of four treatments including soil solarization, fumigation with Metham-Na at the rate of 750 kg a,i./ha (Turkam 50 percent w/v at 1 500 1/ha) or methyl bromide at the rate of 100 g/m², and an untreated control. Treatments were replicated four times in a randomized complete block design, with 60 experimental plots (9.3 x 6.4 m).
Soil was solarized for eight weeks from mid-July through mid-September, 1987, by means of transparent PE 7/m wide and 50 m m thick placed in position after moistening the soil by sprinkler irrigation. Metham-Na was applied on October 1, 1987, with the soil sealed by flooding the plots after the treatment, and methyl bromide was applied two weeks later. On December 1,1987, about 500 cuttings of Indica rootstock were planted per plot, in 10 rows 85 cm apart. Rootstocks were grafted with rose cultivar Koba on April 26. Effects of soil treatments were assessed by the percentage of dead cuttings and the vigour of plant growth estimated on a 1-10 scale four months after planting, as well as by the level of weed infestation. Plants were harvested on February 2, 1989 and sorted out into four commercial qualify standards A-D.
Soil solarization for two months in the absence of major known pathogens increased the percentage of rooted cuttings and plant vigour of Indica rootstock, reduced the weed infestation, and increased the gross value production of grafted plants by 37 percent of that in untreated control. Those effects were highest in plots fumigated with methyl bromide, which increased the gross value production of grafted plants by 35 percent of that in untreated control (Table 5).
Conclusion
Our results have shown that soil solarization under environmental conditions prevailing in southern Spain is widely applicable as a control measure for Verticillium wilt of cotton and Fusarium wilt of watermelon, which severely affect these crops in the area.
Control of Verticillium wilt in the Marismas area of the Guadalquivir Valley by soil solarization is of particular significance for cotton production in southern Spain, since it demonstrated the efficacy of that measure in soil with a rather high ID of the defoliating pathotype of V. dahliae. Also, it illustrates that clay soils prevalent in that area arc suitable for solarization even in years for which environmental conditions in summer time do not seem favourable.
Experiments to control Fusarium wilt of watermelon under plastic houses have shown that solarization for two months provides a better control of the disease than fumigation with Metham-Na, and it resulted in higher increases of plant growth and yield. Solarization during one month, in July, was not highly effective in either eradicating the pathogen from soil or decreasing disease incidence. Therefore, when solarization under plastic house conditions is not feasible for more than 45 days, it should be integrated with fumigation at low dose (1) and should meet optimal environmental conditions (probably from mid-July on).
References
1. Ben-Yephet, Y., J. M. Melero-Vara, and J. E. DeVay. 1988. Interaction of soil solarization and metham-sodium in the destruction Verticillium dahliae and Fusarium oxysporum f. sp. vasinfectum. Crop Protection 7:327-331.
2. Blanco-Lopez, M. A., J. Alcazar-Bejarano, J. M. Melero-Vara, and R. M. Jimenez-Diaz. 1989. Current status of verticillium wilt of cotton in southern Spain: pathogen variation and population in soil. Pages 123-132 In: NATO ASI Series, Vol H28. Vascular Wilt Diseases of Plants Ed. E. C. Tjamos and C. Beckman. Springer-Verlog, N.Y.
3. Bouhot, D. and F. Rouxel. 1971. Technique selective et quantitative d' analyse des Fusarium oxysporum et Fusarium solani de sol Mode d' emploi. Ann. Phytopathol. 3(2):251-254.
4. Butterfield, E. J. and J. E. DeVay. 1977. Reassessment of soil assays for Verticillium dahliae. Phytopathology 7:1073-1078.
5. Gonzalez-Torres, R., R. M. Jimenez-Diaz, and J. Vazuez-Gomez. 1988. Incidencia y distribucion de las Fusariosis vasculares del melon y la sandia en Andalucia. Investigacion Agraria. Produccion y Proteccion Vegetal 3:377-392.
6. Greenberger, H., J. Katan, and Y. Keren. 1986. Control of Fusarium wilt in watermelon, 1984-5. B & C Tests 1:25. APS Press, Minneapolis, MN.
7. Katan, J. 1981. Solar heating solarization. of soil for control of soilborne pests. Annual Review of Phytopathology 19:211-236.
8. Martyn, R. D. and T. K. Hartz. 1986. Use of soil solarization. to control Fusarium wilt of watermelon. Plant Disease 70:762-766.
9. Meler, J. M., R. Gonzalez, J. Gomez, J. Bejarano, and M. J. Basallote. 1989. La solarizacion de suelos mediante acolchado en Andalucia. Plasticulture 82:73-82.
10. Pullman, G. S., J. E. DeVay, R. H. Garber, and A. R. Weinhold. 1981. Soil solarization. Effects on Verticillium wilt cotton and soilborne populations of Verticillium dahliae, Pythium spp., Rhizoctonia solani and Thielaviopsis basicola. Phytopathology 71:954-959.
Table 1. Effect of soil solarization. on the population of Verticillium
dahliae in soil
| Inoculum density (p/g) | |||||
| Year | Field | Treatmenta | Before treatment | After treatment | |
| 1986 | |||||
| A | Unsolarized | 23.3 | 16.1 | ||
| Solarized | 27.3 | 0 | |||
| B | Unsolarized | 2.4 | 2.8 | ||
| Solarized | 1.8 | 0.2 | |||
| 1987 | |||||
| C | Unsolarized | 20.8 | 68.3 | ||
| Solarized | 20.0 | 0 | |||
| D | Unsolarized | 3.0 | 34.3 | ||
| Solarized | 3.5 | 0 | |||
| E | Unsolarized | 7.3 | 42.0 | ||
| Solarized | 7.5 | 0 | |||
a Plots were solarized from July 9 - August 31 in 1986, and from
July 16-September 30, in 1987.
Table 2. Effect of soil solarization on Verticillium wilt
and seed cotton yield
| Year | Field | Treatment | Inoculum
density at planting (p/g) |
Disease incidence (%) |
Seed cotton yield (kg/ha) |
|
| Upper layer |
Lower layer |
|||||
| 1987 | ||||||
| A | Unsolarized | 51.3 | - | 90.5 | ||
| Solarized | - | - | 5.0 | |||
| B | Unsolarized | 6.0 | - | 60.1 | 2 093 | |
| Solarized | 0.1 | - | 4.5 | 4 833 | ||
| 1988 | ||||||
| C | Unsolarized | 28.8 | 5.8 | 87.7 | 1 562 | |
| Solarized | 1.5 | 0.3 | 13.5 | 3 279 | ||
| D | Unsolarized | 15.3 | 5.0 | 55.0 | 3 425 | |
| Solarized | 0.3 | 0 | 1.8 | 4 575 | ||
Table 3. Population of Fusarium oxysporum in
treated soil as a percentage of the initial population before
treatment
Sampling date |
||||
| Treatment | 28/9/88 | 30112/88 | 6/4/89 | 3/7/89 |
| Infested control | 21.1 | 51.1 | 23.3 | 24.4 |
| Solarization (1 mth) | 5.8 | 15.9 | 32.7 | 7.1 |
| Solarization (2 mth) | 0.9 | 3.7 | 15.6 | 18.4 |
| Metham-Na | 2.8 | 3.2 | 2.4 | 7.2 |
Table 4. Effects of soil treatment on growth and yield of
watermelon in sand-mulched plastic house
| Treatment | Plant Dry weight (g) |
Number of fruits per plot |
Yield (kg/plot) |
| Infested control | 78.2 x | 10.0 x | 6.5 x |
| Uninfested control | 111.9 x y | 16.0 x | 16.5 x y |
| Solarization (1 mth) | 116.2 x y | 13.0 x | 15.1 x y |
| Solarization (2 mth) | 213.9 z | 27.5 y | 32.0 z |
| Metham-Na | 178.2 | 17.5 x y | 21.9 y z |
a Values followed by the same letter within columns are not
significantly different (P=0.05) according to Student's t
test.
Table 5. Effects of soil treatments on growth of Rosa
indica rootstocka
| Treatment | Incidence of dead cattingsa |
Plant growth vigourb |
Incidence of weed infestation (Number/1.25 m²) (%) |
Gross value production ($ US) |
| Untreated | ||||
| control | 5.6cx | 1.8cx | 112.5 x | 1 377 x |
| Solarization | 4.0 xy | 6.0 y | 32.5 y | 1 881 y |
| Methyl | ||||
| bromide | 3.1 y | 8.1 z | 3.5 z | 1 863 y |
| Metham-Na | 3.1 y | 3.4 x | 24.3 y | 1 783 z |
a Determined four months after planting.
b Based on 1-10 scale (1= least vigour, 10= greatest vigour).
c Mean of four replications. Values followed by the same letter
within columns are not significantly different (P= 0.5) according
to Student's t test.
Figure 2. Soil temperature in sand-mulched greenhouse in solarized and unsolarized soil.
Figure 3. Fusarium wilt of watermelon in greenhouse treated soils - 1989.
Control of fusarium solani broad bean by solar heating of the soil in Northern Iraq
A.R.T. Sarhan, Department of Biology, College of Science, University of Salahaddin, Erbil, Iraq
Abstract
Covering moistened field soil with transparent polyethylene sheets during the hot season caused an increase in soil temperature which resulted in the control of Fusarium solani, the causal agent of root rot disease in broad bean. The populations of the pathogen were reduced in the mulched soils. Soil covered for six weeks during July and August was sown with broad bean seeds and this resulted in a significant decrease in the percentage of diseased plants and rotten roots. Consequently, plant growth was increased. This method of control is less costly than chemical ones, non-hazardous and easy to apply in agriculture.
Introduction
Broad bean (Faba vulgaris Moench) is an important crop in Iraq. According to our previous work (12), it was found that among the soilborne pathogens of this crop, Fusarium solani (Maritus), causes a serious seed and root rot disease. At present control of these diseases in Iraq depends on chemical methods (10), or biological methods (11). Soil solarization, a process that utilizes the sun to heat soil under plastic sheets, was reported to be successful in controlling many soilborne pathogens (2, 3, 5, 9, 13). No previous work has been done regarding the effect of soil solarization on this pathogen in Iraq. The objectives of this research were to determine whether soil solarization would control root rot disease in broad bean caused by F. solani and the populations of pathogen in the soil, and also to see whether soil solarization would influence the growth of plants.
Materials and Methods
Field experiments were carried out in naturally infested soil in northern Iraq (Erbil area), where broad bean had been grown frequently. F. solani was aseptically isolated on potato - dextrose - agar (PDA) and identified by the Commonwealth Mycological Institute (C MI) . Individual plots 2 x 4 m were arranged in a randomized complete block design consisting of four treatments each with five replications. Solarization was carried out on soils moistened by irrigation, two days before mulching, to increase thermal sensitivity of F. solani and improve heat transfer. Thereafter, plots were irrigated every two to three weeks until the plastic was removed. Plots were mulched with 0.05 mm transparent polyethylene plastic sheets for two, four and six weeks during July and August. Soil temperatures were measured and recorded for solarized and control treatments at 5, 10 and 20 cm depths. Soil samples were taken from solarized and control soils, after removal of the polyethylene sheets, to assay the populations of F. solani using the dilution method as described by Al-Hassan et al. (2). Plots which were solarized for six weeks were used in further experiments.
Broad bean seeds (cv. Ekwadelgii) were sown the following autumn on 15 October. Results were expressed as percentage of diseased plants and as an average of disease incidence several times during the season. Fusarium root rot was estimated using a 0 - 4 scale, where 0 = healthy roots and 4 = roots completely destroyed. At each evaluation, 25 plants were uprooted from each plot and their roots were washed under tap water and examined for disease symptoms. To measure the effect of soil solarization on growth of plants, the shoot lengths and dry weight of plant were determined at different times after emergence during the growth season.
Results and Discussion
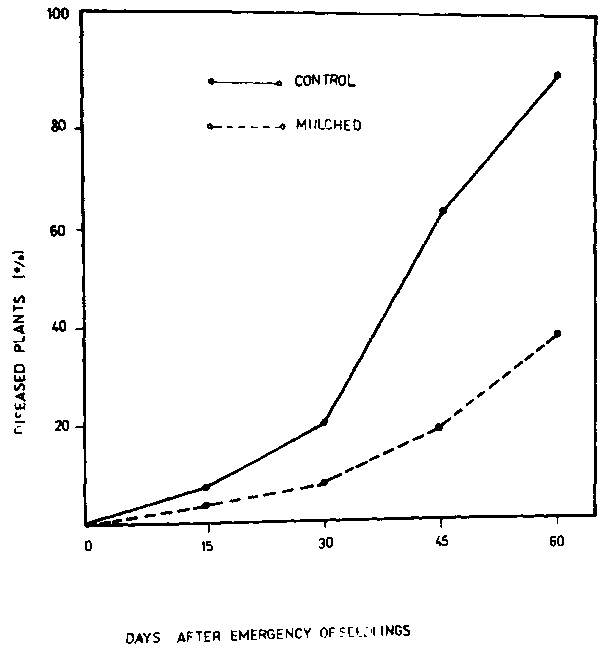
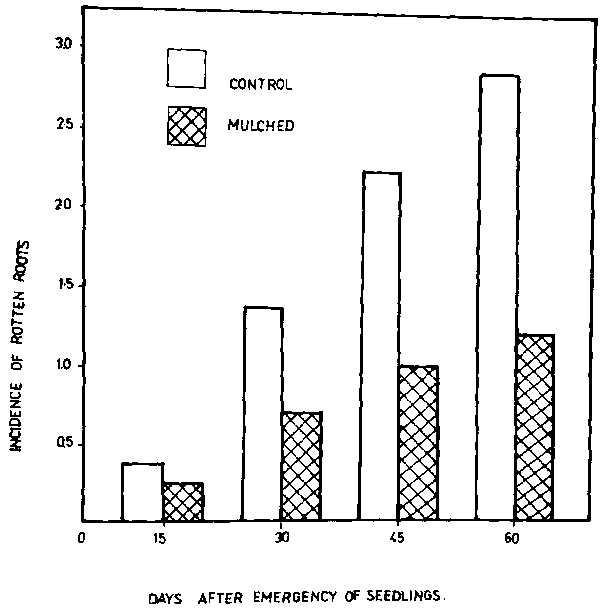
Results in Table 1 show the effect of soil solarization on maximum soil temperatures. The mulching of moist soils raised the soil temperature and the highest temperature was in the upper layer at 5 cm depth. Soil temperatures at the other depths were also increased in mulched soil compared to the unmulched soil. The differences in the maximum temperatures between mulched and unmulched soils after six weeks were 17°, 13° and 12°C at the 5, 10 and 20 cm depths, respectively. Soil solarization after four and six weeks reduced significantly the soil population of the pathogen at all depths compared to the unmulched treatment (Table 2). Results showed that F. solani population densities were considerably reduced after four and six weeks of soil mulching. These results are in accord with those obtained by Nelson and Wilhelm (8), who reported that temperatures achieved at the upper layers by soil mulching are in the range of those lethal to plant pathogens. Figure I shows the percentage of diseased plants in the solarized and unmulched soils during the season. The percentage of diseased plants after 60 days from emergence dropped from 89 percent in the unmulched treatment to 38 percent in the solarized ones. Disease incidence of Fusarium root rot of broad bean plants in solarized soil was significantly reduced after 30, 45 and 60 days compared to unmulched treatments (Fig. 2).
Results presented here agree with those obtained by Garber and Houston (4) and Pullman et al. (9), who demonstrated that soil heating at sublethal temperatures impaired the ability of some soilborne plant pathogens to penetrate the plant and cause plant disease. This probably indicates that solar healing causes an alteration in the soil microflora which involves a biological control in addition to the thermal effect on the soilborne pathogens. However, Moorman (6) showed that wilt disease incidence in eggplants caused by Verticillium did not differ significantly between solarized (black polyethylene) and unmulched soils.
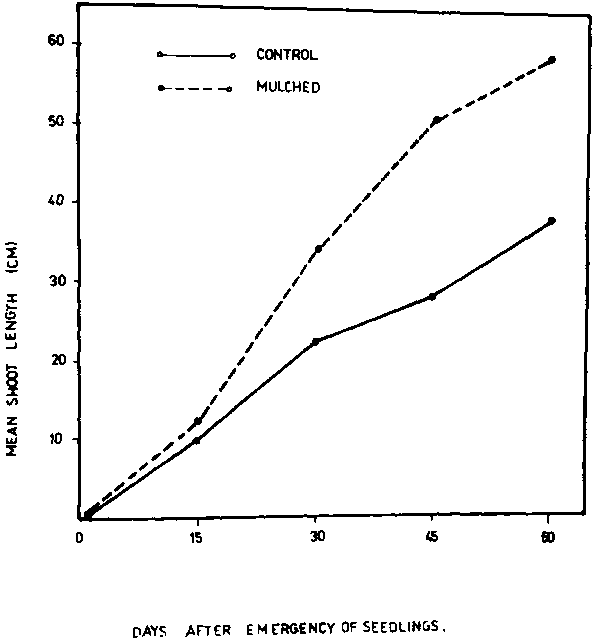
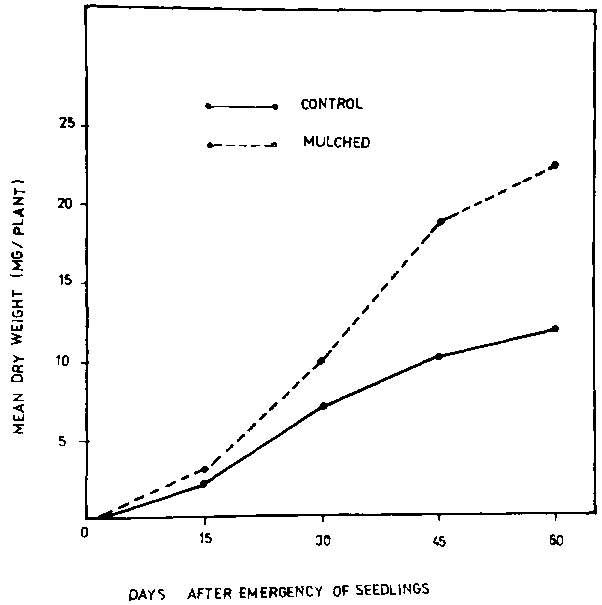
The effect of soil solarization on shoot lengths of broad bean plants is shown in Figure 3. Soil solarization improved plant growth. The increase in the growth of plants was more pronounced after 45 and 60 days from emergence. Plant dry weights (Fig. 4) indicated that plants grown in solarized soil had significantly greater growth than those in the unmulched soil. These findings confirm the results of Moosa et al. (7) on the effect of soil mulching on growth of watermelon, who reported that soil mulching speeded up the release of nutrient available in the soil for plant use. Finally, it should be recognized that soil solarization can be carried out because it is less costly, non-hazardous and easy to apply in agriculture.
References
1. Abu-Gharbieh, W. 1989. Use of solar energy for control of soil-borne pests and pathogens. Arab J. Pl. Protection 7(1): 108 (Abstract).
2. Al-Hassan, K.K., H.A. Mahmoud, M.T. Abd-Alhamid, and L.l. Mohamed. 1985. Effect of soil energy and some chemical compounds on soil fungi in plastic houses. 1. Agric. Water Reso. 4(3):83-93.
3. Al-Raddad, A.M. 1979. Soil disinfestation by plastic tarping. M.Sc. Thesis, Univ. Jordan, Amman. pp. 95.
4. Garber, R.H. and B.R. Houston. 1966. Penetration and development of Verticillium albo-atrum in the cotton plant. Phytopathology 56:1121-1126.
5. Jones, J. P., A.J. Overman, and C.M. Geraldson. 1972. The effect of mulching on the efficacy of DD - MENCS for control of Fusarium wilt of tomato. Plant Dis. Reptr. 56(11): 953-956.
6. Moorman, G.W. 1982. The influence of black plastic mulching on infection rates of Verticillium wilt and yield of eggplant. Phytopathology 72:14121414.
7. Moosa, M., R.M. Al-Safah, and I.M. Jassim. 1985. The effect of plastic mulch colours on growth of watermelon plants and some soil properties. J. Solar Energy Res. 5(1):39-53.
8. Nelson, P.E. and S. Wilhelm. 1958. Thermal death range of Verticillium albo-atrum. Phytopathology 48:613-616.
9. Pullman, G.S., J.E. DeVay, R.H. Garber and A.R. Weinhold. 1981. Soil solarization: Effect on Verticillium wilt of cotton and soilborne populations of Verticillium dahliae, Pythium spp., Rhizoctonia solani, and Thielaviopsis basicola. Phytopathology 71:954-959.
10. Sarhan, A.R.T. and F.M. Toma. 1989. Study on potential of some systemic fungicides, soil fumigants and solar energy for control of broad bean rootrot. J. Coll Edu., Salahaddin Univ. 1(3):20-34.
11. Sarhan, A.R.T. and M.E. Taha. 1989. Biological control of root rot disease of broad bean. 3rd Nat. Conf. of Pests and Dis. of Veg. and Fruits in Egypt and Arab Countries, Ismailia. p. 85.
12. Sharif, F.M., A.R.T. Sarhan, A.M. Hegazi, A.H. Fatah and M. Salam. 1987. Fungi associated with broad bean seed and their control. J. Univ. Kuwait (Sci.) 14:331-336.
13. Zaid, A.W., W. Ismail and F. Bishaya. 1989. The use of soil solarization to control soil-borne pathogens in great Libyan Jamahiriya. Arab J. Pl. Protection 7(1):85 (Abstract).
Table 1. Effect of soil solarization. on maximum soil
temperatures at various soil depths during experiments
| Date of soil mulching¹ | Soil
temperatures (C) at different depths (cm) |
||
| 5 | 10 | 20 | |
| July 1 to 15 | |||
| Control | 38 | 31 | 24 |
| Mulched | 52 | 43 | 38 |
| July 1 to July 30 | |||
| Control | 40 | 32 | 27 |
| Mulched | 58 | 45 | 38 |
| July 1 to August 15 | |||
| Control | 42 | 35 | 32 |
| Mulched | 59 | 48 | 44 |
1 Soil was irrigated two days before mulching
Table 2. Effect of soil solarization on the survival of
Fusarium solani at different depths
| Treatment* | Fusarium
propagules/g of soil at different depths (cm) |
||
| 5 | 10 | 20 | |
| Control | 216 a | 264 a | 338 a |
| Solarized 2 weeks | 190 a | 232 a | 289 b |
| Solarized 4 weeks | 49 b | 84 b | 93 c |
| Solarized 6 weeks | 19 c | 38 c | 55 d * |
1 Transparent polyethylene sheets were placed on the soil July 1.
* Values within columns followed by the same letter are not significantly different (Duncan's multiple range test, P = 0.05).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}