![]()
![]()
![]()
C.E. Harwood et E.R. Williams
CSIRO Division of Forestry
P.O. Box 4008, QVT, Canberra 2600
Australie
RESUME
Cet article rapporte les résultats des essais internationaux de provenances d'Acacia mangium coordonnés par la FAO et le CSIRO, mis en place dans les années quatre-vingts. Les données provenant de 19 sites dans le Sud-Est et le Sud asiatique, en Australie et aux Fidji ont été étudiées. Au total 24 provenances locales de cinq régions, dont deux provenances de Papouasie-Nouvelle-Guinée (PNG), 19 de la région de Cairns au Queensland (QCR), une de l'extrême nord du Queensland (FNQ), une de Céram (CERAM) et une d'Irian Jaya (IRIAN) ont été testées. Aucun des essais n'a testé les 24 provenances, et les sous-ensembles testés variaient d'un site à l'autre. L'analyse de la variance de la hauteur et du diamètre pour chaque site d'essai montre des différences significatives entre les provenances testées.
Les données de hauteur et de diamètre ont été converties en accroissements annuels moyens pour l'analyse de la variance entre les différents sites d'essais. On constate des différences hautement significatives dans les performances entre sites d'essais, entre régions de provenance et entre provenances locales dans une même région de provenance. La croissance était généralement plus rapide sur les sites se trouvant au voisinage de l'équateur, avec des accroissements annuels moyens en hauteur de l'ordre de 3–4 m, et plus lente sur les sites plus éloignés de l'Equateur au Bangladesh, en Chine méridionale, à Taiwan et aux Fidji. Les provenances de PNG fournissent régulièrement les meilleures performances, suivies par la provenance Claudie River de FNQ. Les provenances de la région QCR (16°–18°30'S) et celles de CERAM et IRIAN avaient presque toujours une croissance plus lente que celles de PNG et FNQ.
INTRODUCTION
Acacia mangium Willd est devenu depuis une vingtaine d'années une essence de reboisement importante dans les tropiques humides. Des récoltes de semences en vue d'essais internationaux de provenances, coordonnées par le CSIRO et la FAO, ont été effectuées au début des années quatrevingts (Doran & Skelton, 1982; Turnbull et al., 1983). Les résultats publiés d'un certain nombre d'essais de provenances (Atipanumpai, 1989; Chung et al., 1990; Hadi et al., 1990) montrent des différences importantes dans la croissance de différentes provenances naturelles. Le Centre de semences forestières d'Australie a en 1990 effectué une enquête pour rassembler les données les plus récentes de tous les essais connus à partir de lots de semences FAO/CSIRO. Plusieurs des réponses reçues contenaient des données jusqu'alors non publiées, ce qui a permis une analyse de la croissance sur un large éventail de stations tropicales.
MATERIELS ET METHODES
Provenances testées
Le Tableau 1 fournit des données détaillées sur les provenances d'A. mangium incluses dans les essais, tandis que le Tableau 2 présente les principaux paramètres climatiques concernant les provenances du Queensland et de Papouasie-Nouvelle-Guinée, calculées au moyen du programme informatique BIOCLIM (Booth et al., 1988).
Les lots de semences testés étaient tous des lots mélanges provenant d'au moins cinq semenciers bien espacés (10 ou plus dans la plupart des cas), et peuvent être considérés comme représentant des provenances locales. On peut logiquement les classer en cinq régions de provenance géographiquement distinctes. Le terme de région de provenance est employé ici pour désigner une grande zone géographique dans l'aire naturelle de l'espèce, et n'implique pas que les provenances locales d'une même région de provenance aient nécessairement les mêmes caractéristiques génétiques (Turnbull & Griffin, 1984). Ces régions de provenance sont les suivantes:
Queensland, région de Cairns (QCR) - sud de l'aire d'A. mangium le long de la côte orientale du Queensland (Australie), au sud du parallèle 15°S. Dans cette région de provenance on trouve de nombreuses populations de petite à moyenne taille en lisière de forêt dense, s'étendant en latitude sur plus de trois degrés jusqu'à 18°30'S et en altitude du niveau de la mer à environ 400 m. Dix-neuf lots de semences ont été inclus dans les essais.
Extrême nord du Queensland (FNQ) - au nord du parallèle 13°S. Présence limitée en lisière des forêts-galeries le long des cours d'eau qui coulent vers les côtes est et ouest du Cap York: Claudie, Pascoe, Wenlock, Olive, Jardine. Une seule récolte provenant de six arbres de Claudie River a été testée dans les essais.
Papouasie-Nouvelle-Guinée (PNG) - A. mangium se trouve dans toute la zone de plaines du sud de la Province occidentale de Papouasie-Nouvelle-Guinée. Il y est commun, se trouvant localement en populations de taille variable sur les stations bien drainées. Les populations de la province occidentale sont contiguës à des populations étendues du sud-est de l'lrian Jaya (Gunn & McDonald, 1991). Les deux lots de semences de PNG utilisés dans les essais étaient séparés l'un de l'autre d'environ 180 km.
Céram, Indonésie (CERAM) - A. mangium se rencontre dans l'île de Céram en Indonésie. Une petite population a été échantillonnée près de Piru (Turnbull et al., 1983).
Irian Jaya, Indonésie (IRIAN) - Une seule population de faible étendue (15 ha) à Sidei dans la péninsule de Vogelkop (Turnbull et al., 1983).
Sites d'essais
Le Tableau 3 fournit des informations sur les sites d'essais. Des données étaient disponibles pour les sites de Bornéo (Sabah, Sarawak et Sud Kalimantan), de Thailande, de Chine méridionale, de Taiwan, du Bangladesh, de l'île Melville dans l'extrême nord de l'Australie, et des Fidji. On espère inclure dans un prochain numéro d'IRGF des données sur les essais d'Amérique Centrale, de Malaisie Péninsulaire, des Philippines, des îles Salomon et d'Afrique occidentale.
RESULTATS
Analyse des données par essai
Le Tableau 4 indique les lots de semences testés dans chaque essai. Outre les différentes combinaisons de lots de semences testés, il y avait des différences entre les sites d'essais en ce qui concerne les techniques sylvicoles telles que doses d'engrais appliquées et calendrier des désherbages, ce qui a sans aucun doute influé sur la croissance. Ces facteurs rendent approximatives les comparaisons de potentialités des différents sites.
Tous les essais rapportés ici utilisaient un dispositif en blocs complets aléatoires avec répétitions, toutefois la taille et la forme des parcelles et le nombre de répétitions variaient. L'analyse de la variance pour chaque site d'essais a été effectuée soit par l'organisme qui conduisait l'essai, soit dans le cours de la présente étude. On a constaté des différences significatives entre provenances en ce qui concerne la hauteur et le diamètre à hauteur d'homme sur tous les sites d'essais (la hauteur n'a pas été mesurée sur deux des sites, et le diamètre n'a pas été mesuré sur un site). Pour les sujets à plusieurs brins, on a admis comme diamètre le diamètre équivalent d'un arbre à tronc unique (c'est-à-dire le diamètre donnant la même surface terrière). Cette hypothèse devra être confirmée pour tous les sites.
Le taux de survie au moment des mesures était élevé sur la plupart des sites, comme le montre le tableau résumé ci-dessous:
| Site d'essai | Age au moment de l'évaluation de l'essai (ans) | Fourchette de taux de survie des différentes provenances | |
|---|---|---|---|
| Sabah Forest Industries | |||
| Site A | 4,0 | 70 – 86 % | |
| Site B | 4,0 | 64 – 91 % | |
| Site C | 3,0 | 70 – 78 % | |
| Site D | 3,0 | 70 – 85 % | |
| Safoda, Sabah | 4,0 | non disponible | |
| Sarawak (5 sites) | 4,0 | non disponible | |
| Sud Kalimantan site 1 | 2,5 | 100 % | |
| Sud Kalimantan site 2 | 7,0 | 60 – 72 % | |
| Thailande | 2,5 | 98 – 100 % | |
| Chine méridionale (2 sites) | 4,0 | non disponible | |
| Taiwan | 4,5 | non disponible | |
| Ile Melville, Australie | 4,5 | 75 – 98 % | |
| Fidji | 3,0 | 17 – 77 % | |
| Bangladesh | 1,5 | 32 – 80 % | |
L'âge au moment des mesures variait de 1,5 à 7 ans, mais était dans la plupart des cas compris entre 3 et 4,5 ans. L'examen de séries chronologiques de données sur la croissance dans les essais de Taiwan (2,5, 3,5 et 4,5 ans) et de Barito, Sud Kalimantan (2 et 7 ans) indique des taux de croissance en hauteur et en diamètre constants au cours des premières années suivant la plantation. Cela signifie que les comparaisons d'accroissements annuels moyens entre différents sites seraient peu influencées par les différences dans l'âge des essais au moment des mesures. On a en conséquence calculé les accroissements annuels moyens en hauteur et en diamètre pour chaque provenance sur chaque site, et ces données ont été ensuite analysées.
Analyse de variance entre sites et entre provenances
Etant donné que huit des provenances du Tableau 1 étaient absentes dans tous les essais de Bornéo, et que cinq autres provenances n'étaient présentes que dans l'un de ces essais, il fut décidé de faire deux analyses séparées, l'une pour les douze sites de Bornéo et l'autre pour les sept sites restants dans les autres pays, où les provenances étaient représentées de manière bien plus complète. Ces deux analyses de variance portaient sur la variation entre sites, entre régions de provenance, entre provenances d'une même région de provenance (c'est-à-dire les 13 provenances de la région QCR et les 2 provenances de la région PNG), et l'interaction entre site et région de provenance.
Pour les accroissements annuels moyens tant en hauteur qu'en diamètre, et pour les deux groupes d'analyses (Bornéo et autres), on a constaté des différences hautement significatives entre sites (P < 0,001), entre régions de provenances et entre provenances d'une même région de provenance. L'interaction site/région de provenance était également hautement significative (P < 0,001) pour les essais de Bornéo, mais tout juste significative (P < 0,05) pour les autres essais. Les analyses de variance seront rapportées plus en détail dans une prochaine publication.
Résumé des performances des différentes régions de provenance sur les différents sites d'essais
Les performances des cinq régions de provenance aux différents sites d'essais sont résumées dans le Tableau 5, et illustrées pour les sites autres que ceux de Bornéo dans les Figures 1 et 2. Les accroissements annuels moyens indiqués pour les régions de provenance QCR et PNG sont les moyennes des lots de semences provenant de ces régions testés sur chaque site. Etant donné que différents groupements de provenances de ces deux régions ont été testés sur les différents sites, les comparaisons entre sites ne sont pas exactes.
On constate une nette tendance à une croissance plus rapide sur les sites proches de l'équateur, avec un accroissement annuel moyen en hauteur de l'ordre de 4 m sur certains sites. La croissance sur des sites proches du tropique du Cancer en Chine méridionale, à Taiwan et au Bangladesh, ainsi qu'aux Fidji (17°50'S), est bien plus lente, avec des accroissements de seulement 1–2 m/an. En Chine méridionale, la croissance plus lente à Leilin par rapport à Haikang peut être attribuée à la fertilité moins bonne du sol (Pan Zhi-gang, comm. pers. 1990).
Une différence importante apparaît également dans les performances entre les différentes régions de provenance. PNG est presque toujours la meilleure région de provenance là où elle a été testée, quoiqu'elle n'ait été incluse que dans deux essais à Bornéo. L'unique provenance FNQ a également donné de bons résultats partout où elle a été incluse, mais avec une croissance légèrement plus lente que PNG dans la plupart des localités. Ces tendances restent valables pour les accroissements en hauteur comme en diamètre, de sorte que l'avantage de PNG et FNQ sur les trois autres régions de provenance en ce qui concerne la production en volume est important dans la plupart des cas. Les résultats d'un essai de provenance en Côte d'Ivoire, résumés par Souvannavong (1990), sont en accord avec la tendance générale à la supériorité des provenances PNG et FNQ par rapport aux trois autres régions.
Dans les essais à Taiwan et en Chine méridionale, l'une des deux provenances QCR a poussé plus vite que PNG et FNQ, mais les différences n'étaient pas significatives. CSIRO no13237 et 13239 étaient les meilleures provenances QCR à Taiwan, tandis que 13242 était la meilleure sur les deux sites de Chine méridionale. CSIRO no13242 a également donné d'aussi bons résultats que les deux provenances PNG au Bangladesh. Aux sites A et B de Sabah Forest Industries, les provenances FNQ ont été dépassées par la provenance CERAM et une ou deux provenances QCR, mais là encore les différences n'étaient pas significatives. Sur tous les autres sites d'essais où elles ont été incluses, les provenances PNG et FNQ ont dépassé les provenances CERAM et IRIAN, et toutes les provenances QCR. Il ne faut pas perdre de vue, toutefois, que la région de provenance FNQ n'est représentée dans ces essais que par un seul lot de semences venant de six semenciers.
Les performances relatives de QCR, CERAM et IRIAN sont variables, CERAM et IRIAN étant généralement meilleures sur des sites proches de l'équateur, tandis que les provenances QCR les dépassent en Chine méridionale, à Taiwan, au Bangladesh, aux Fidji et à l'île Melville (Australie).
Forme
Pour de nombreux sites d'essais on ne dispose pas encore d'évaluations de forme, de sorte que l'on n'a pas tenté à ce stade d'analyse des différences entre sites et entre provenances en ce qui concerne la forme. Les pourcentages d'arbres à tronc unique sur deux sites d'essais au Sabah, le site de l'île Melville en Australie et le site de Côte d'Ivoire (Souvannavong, 1990) sont indiqués dans le Tableau 6 qui illustre l'amplitude de la variation de la forme entre provenances et les différences importantes que l'on constate entre sites.
DISCUSSION
Il ressort des résultats des essais rapportés ici, et d'autres essais de provenances réalisés par la suite, qu'il existe des différences génétiques importantes entre régions de provenance et entre provenances locales, se traduisant par des différences constantes et significatives dans leurs performances de croissance. L'analyse d'isoenzymes indique pour A. mangium une faible diversité génétique (Moran et al., 1989). Les études d'isoenzymes n'examinent qu'un petit sous-ensemble du génome, et en conséquence ne fournissent pas une mesure absolue de la variabilité génétique d'ensemble. Toutefois, on peut noter que les deux provenances indonésiennes avaient la plus faible diversité d'isoenzymes et les moins bonnes performances dans les essais de provenances. Leurs faibles performances pourraient bien être dues à la consanguinité dans des populations issues d'une base génétique étroite, peut être fondées par une seule graine transportée par un oiseau, ou provenant d'une introduction par l'homme.
La vitesse de croissance des provenances PNG reste plus élevée que celle de la plupart des provenances QCR sur les sites d'essais de climat à hiver frais de Taiwan et de Chine méridionale. La croissance plus lente de PNG par rapport à QCR sur ces sites était prévisible si l'on considère que les provenances QCR sont originaires de climats à hivers plus frais et sont donc a priori mieux adaptées (Tableau 2). Si quelques provenances QCR l'emportaient sur PNG et FNQ en Chine et à Taiwan, les différences n'étaient toutefois pas significatives, et comme on l'a noté plus haut l'interaction entre région de provenance et site d'essai pour les sites autres que ceux de Bornéo était tout juste significative pour les accroissements annuels moyens en hauteur, et non significative pour les accroissements en diamètre.
Il est intéressant de noter que la croissance sur le site D de Sabah Forest Industries, à une altitude de 1000 m et avec une température moyenne estimée de 20°C, était bien plus rapide (accroissement moyen en hauteur 3,46 m) que sur les sites du Bangladesh, de Chine méridionale et de Taiwan où la température moyenne annuelle est de l'ordre de 22–24°C, et où l'accroissement moyen en hauteur n'était que de 1–2 m. Il apparaît que les basses températures hivernales arrêtent la croissance pendant plusieurs mois sur les sites subtropicaux (You Yintian, 1989), tandis qu'elle se poursuit toute l'année dans les conditions tempérées du site équatorial d'altitude du Sabah où il y a peu de variation saisonnière des températures. Toutefois, la faible dominance apicale sur le site d'altitude du Sabah a pour résultat des arbres de forme médiocre à tiges multiples (Tableau 6; Sim Boon Liang, comm. pers. 1990). En Thailande, des périodes de faible amplitude thermique journalière s'associent à une croissance rapide en diamètre, cependant il est difficile de distinguer cet effet de celui des variations saisonnières de pluviosité (Atipanumpai, 1989).
Depuis que les essais internationaux rapportés ici ont été réalisés, le Centre de semences forestières d'Australie a entrepris des récoltes en vue de nouvelles recherches, dont des récoltes d'arbres individuels sur un total de plusieurs centaines de semenciers dans de nombreuses provenances naturelles d'A. mangium, en particulier dans les régions de provenance PNG, FNQ et QCR (Gunn & Midgley, 1991; Gunn & McDonald, 1991). Les récoltes d'arbres individuels ont été utilisées pour établir les populations de base en vue d'un certain nombre de programmes d'amélioration génétique. Toutefois, on ne sait rien jusqu'à présent sur les gains génétiques en croissance et en forme que l'on pourrait obtenir par croisement entre différentes régions de provenance d'A. mangium.
REMERCIEMENTS
Nous sommes très reconnaissants aux organisations et aux personnes mentionnées au Tableau 3 qui nous ont fourni des données sur les essais, dont une grande partie n'avait pas été publiée. Le Dr Garth Nikles a aidé à rassembler ces données et a fourni de précieux commentaires sur une première rédaction de cet article. M. Patrick Milimo a aidé au classement des données, et M. Tom Jovanovic a calculé les paramètres climatiques pour les différentes provenances au moyen du programme informatique BIOCLIM. Le projet Semences d'arbres australiens, financé par le Bureau australien d'assistance au développement (ACIAR) et géré par le Centre de semences forestières d'Australie (ATSC), a fourni un appui pour permettre la préparation de cet article.
REFERENCES
Atipanumpai, L. (1989). Acacia mangium: Studies on the genetic variation in ecological and physiological characteristics of a fast-growing plantation species. Acta Forestia Fennica 206.
Booth, T.H., Hutchinson, M.F. & Jovanovic, T. (1988). Niche analysis and tree species introduction. Forest Ecology and Management 23, 47–59.
Chung, Jeng-Der, Hsui, Yen-Ray, Chang, Tein-Yuang & Yang, Jenq-Chuan (1990). Provenance variation of tree height, dbh and volume in A. mangium at young ages. Quarterly Journal of Chinese Forestry. 23, 77–86.
Doran, J.C. & Skelton, D.J. (1982). Récoltes de semencer d'Acacia mangium en vue d'essais internationaux de provenances. Ressources génetiques forestières informations, no 11, 48–50. FAO, Rome.
Gunn, B.V. & McDonald, M.W. (1991). Seed Collections of Eucalyptus urophylla, E. pellita and Acacia species from Indonesia, August–November 1990. Australian Tree Seed Centre.
Gunn, B.V., & Midgley, S.J. (1991). Progress in seed supply of 4 selected tropical Acacia species by the Australian Tree Seed Centre. Paper to ACIAR Workshop on Tropical Acacias, Bangkok, Feb. 11–15 1991.
Hadi, T.S., Adjers, G. & Vuokko, R. (1990). Performance of different provenances of Acacia mangium at 30 months after planting on an alang-alang (Imperata cylindrica) grassland site in South Kalimantan, Indonesia. Technical Report II/IV, June 1990, Indonesian Ministry of Forestry, Directorate General of Reforestation and Land Rehabilitation.
Miller, R.R. & Hepburn, A.J. (1989). A review of the growth of Acacia mangium at the Bengkoka affore-station and settlement project, North Sabah. Regional Symposium on Recent Development in Tree Plantations of Humid/Subhumid Tropics of Asia. Universiti Pertanian Malaysia, 5–9 June 1989.
Moran, G.F., Muona, O. & Bell, J.C. (1989). Acacia mangium: A tropical forest tree of low genetic diversity. Evolution 43, 231–235.
Souvannavong, O. (1990). Recherches sur Acacia mangium Willd., espèce de plantation d'avenir en zones tropicales humides d'Afrique centrale et occidentale. pp. 79–91, Vol. 1(2), IUFRO 19ème Congrès mondial, IUFRO, Montréal.
Turnbull, J.W. & Griffin, A.R. (1986). The concept of provenance and its relationship to infraspecific classification in forest trees. pp. 157–189 in B.T. Styles (ed.) Infraspecific classification of wild and cultivated plants. The systematics Association Special Volume no. 29. Clarendon, Oxford.
Yuo Yintian (1989). Introduction and utilization of Acacia mangium in China. Chinese Academy of Forestry Report, 277p.
Zashimuddin, M., Latif, M.A., Khan, S.A. & Davidson, J.(1985). Performance of different provenances of Acacia mangium Willd in Bangladesh. Bano Biggyan Patrika. Forest Research Institute, Chittagong, Bangladesh. 12. 57–61.
Tableau 1. Liste des provenances incluses dans les essais internationaux de provenances d'A. mangium.
| CSIRO no.1 | Rég. prov.2 | Lat °S | Long °E | Alt m | nb. semenciers | Localité |
| 12990 | QCR | 16 34 | 145 35 | 400 | 10 | Julatten |
| 12991 | QCR | 16 17 | 145 31 | 60 | 10 | Daintree |
| 12992 | QCR | 16 30 | 145 32 | 30 | 10 | Rex Range |
| 13229 | FNQ | 12 44 | 143 13 | 60 | 6 | Claudie River |
| 13230 | QCR | 17 53 | 146 06 | 5 | 10 | Mission Beach |
| 13231 | QCR | 17 42 | 145 57 | 40 | 5 | NW de Silkwood |
| 13232 | QCR | 17 41 | 148 05 | 5 | 10 | Cowley Beach |
| 13233 | QCR | 17 06 | 145 48 | 20 | 10 | N.E. Walshs Pyramid |
| 13234 | QCR | 17 02 | 145 48 | 20 | 10 | E. de Cairns |
| 13235 | QCR | 17 35 | 146 05 | 20 | 5 | Mourilyan Bay |
| 13236 | QCR | 17 48 | 146 05 | 10 | 5 | Kurrimine |
| 13237 | QCR | 17 50 | 146 01 | 20 | 10 | El Arish |
| 13238 | QCR | 17 56 | 146 02 | 70 | 10 | Mission Beach |
| 13239 | QCR | 17 55 | 145 52 | 50 | 10 | Tully |
| 13240 | QCR | 18 14 | 145 50 | 60 | 5 | Cardwell-Ellerbeck |
| 13241 | QCR | 18 21 | 146 03 | 50 | 5 | Broken Pole Ck |
| 13242 | QCR | 18 26 | 146 01 | 60 | 10 | Abergowrie S.F. |
| 13279 | QCR | 16 17 | 145 31 | 60 | 25 | Daintree |
| 13459 | PNG | 8 45 | 141 25 | 30 | 20 | Morehead |
| 13460 | PNG | 8 50 | 143 08 | 10 | 18 | Oriomo R. |
| 13534 | QCR | 16 32 | 145 25 | 60 | 70 | Cassowary Range |
| 13621 | CERAM | 3 04 | 128 12 | 150 | 99 | Piru, Ceram |
| 13622 | IRIAN | 0 46 | 133 34 | 30 | 15 | Sidei |
| 13846 | QCR | 16 31 | 145 24 | 60 | 75 | Mossman |
1 Numéro d'Identification de lot de semences CSIRO, Australian Tree Seed Centre.
Tableau 2. Paramètres climatiques estimés par BIOCLIM pour les provenances du Queensland et de Papouasie-Nouvelle-Guinée.
| CSIRO no. | lat °S | long °E | alt m | TMA* °C | MTMC* °C | MTMF* °C | PAM* mm | PMTS* mm |
| PNG | ||||||||
| 13459 | 8 45 | 141 18 | 30 | 26.2 | 32.3 | 20.8 | 1764 | 102 |
| 13460 | 8 50 | 143 08 | 10 | 26.3 | 32.3 | 20.9 | 2090 | 184 |
| FNQ | ||||||||
| 13229 | 12 44 | 143 13 | 60 | 26.0 | 33.3 | 18.8 | 1876 | 50 |
| QCR | ||||||||
| 12990 | 16 34 | 145 35 | 400 | 23.0 | 32.5 | 12.3 | 2002 | 123 |
| 12992 | 16 30 | 145 22 | 30 | 25.1 | 31.8 | 17.4 | 2128 | 96 |
| 13230 | 17 53 | 146.06 | 5 | 23.8 | 31.6 | 14.4 | 3156 | 225 |
| 13231 | 17 42 | 145 57 | 40 | 23.6 | 31.6 | 14.2 | 3601 | 281 |
| 13232 | 17 41 | 146 05 | 5 | 23.8 | 31.5 | 14.8 | 3258 | 246 |
| 13233 | 17 06 | 145 48 | 20 | 24.3 | 31.4 | 16.2 | 2161 | 80 |
| 13234 | 17 02 | 145 48 | 20 | 24.4 | 31.3 | 16.5 | 2162 | 107 |
| 13235 | 17 35 | 146 05 | 20 | 23.7 | 31.4 | 14.8 | 3243 | 249 |
| 13236 | 17 46 | 146 05 | 10 | 23.8 | 31.5 | 14.5 | 3277 | 247 |
| 13237 | 17 50 | 146 01 | 20 | 23.7 | 31.6 | 14.2 | 3411 | 268 |
| 13238 | 17 56 | 146 02 | 70 | 23.8 | 31.8 | 14.0 | 2061 | 236 |
| 13239 | 17 55 | 145 52 | 50 | 23.7 | 31.9 | 13.8 | 3678 | 338 |
| 13240 | 18 14 | 145 58 | 60 | 23.0 | 32.1 | 13.6 | 2061 | 75 |
| 13241 | 18 21 | 146 03 | 50 | 23.8 | 32.2 | 13.3 | 2061 | 96 |
| 13242 | 18 26 | 146 01 | 60 | 23.9 | 32.3 | 13.1 | 1951 | 89 |
| 13279 | 16 17 | 145 31 | 60 | 25.2 | 31.7 | 18.0 | 2781 | 185 |
| 13846 | 16 31 | 145 24 | 60 | 25.1 | 31.9 | 17.4 | 1977 | 86 |
*MAT = Température moyenne annuelle
*MTMC = Moyenne des température maxima du mois le plus chaud
*MTMF = Moyenne des température minima du mois le plus froid
*PMTS = Pluviométrie du moyenne du brimetre les plus sec
*PAM = Pluviométrie annuelle moyenne
Les donnés climatique pour CERAM et IRIAN n'ont pas pu étre estimées
Tableau 3. Sites d'essais inclus dans l'analyse.
a) L'information sur les sites d'essais est donnée ci-dessous dans l'ordre suivant: Code du site (souligné), emplacement, organisation conduisant l'essai, source d'information.
SbA, SbB, SbC, SbD, Sipitang, Sabah, Malaysia (4 sites d'essais), Sabah Forest Industries. Sim Boon Liang (comm. pers. 1990)
Sfda, Bengkoka, Sabah, Malaysia, SAFODA. Miller & Hepburn (1989)
Swk1, Swk2, Swk3, Swk4, Swk5, Sarawak, Malaysia (5 sites d'essais), Forest Department, Sibu, Sarawak. Forest Silviculturist (comm. pers. 1990)
Sk1, South Kalimantan, Indonésie, Enso/Ministère des Forêts d'Indonésie. Hadi et al. (1990)
Sk2, South Kalimantan, Indonesia, Barito Group. P. Havmoller (comm. pers. 1991)
Thai, Lad-Krating, Thailande, Kasetsart University. Atipanumpai (1989)
Chi L, Chi H, Chine méridionale (2 sites d'essais, Leilin & Haikang), Académie forestière de Chine, Pan Zhi-Gang (comm. pers. 1990).
Taiw, Chung-Pu, Taiwan, Centre de recherche forestière de Taiwan. Chung et al. (1990).
Melv, Melville Island, Northern Territory, Australie, Conservation Commission of the Northern Territory. M.W. Haines (comm. pers. 1991).
Fiji, Nukurua, Fiji, Fiji Ministry of Forests. L.R. Jiko (comm. pers. 1990).
Bdsh, Keochia, Bangladesh, Institut de recherche forestière du Bangladesh. Zashimuddin et al. (1985).
b) Données sur les sites:
| Site | Lat. | Long. | Alt. m | Temp M.A.* °C | P.A.M.* mm | |
| SbA | 5° N | 115°30 E | 100 | 26 | 3100 | |
| SbB | n.d.* | n.d. | 500 | 24 | 3100 | |
| SbC | n.d. | n.d. | 650 | 23 | 3200 | |
| Sbd | n.d. | n.d. | 1300 | 20 | 2000 | |
| Sfda | 6°55 N | 117°05 E | n.d. | 26 | 2600 | |
| Sw1 | 1°03 N | 109°55 E | 35 | 26 | n.d. | |
| Sw2 | 2°13 N | 111°30 E | 30 | 26 | n.d. | |
| Sw3 | 2°17 N | 112°00 E | 30 | 26 | n.d. | |
| Sw4 | 3°22 N | 113°37 E | 35 | 26 | n.d. | |
| Sw5 | 3°45 N | 113°49 E | 170 | 26 | n.d. | |
| SK1** | 3° S | 115° E | 20 | 26 | 2100 | |
| SK2** | 3° S | 115° E | 20 | 26 | 2100 | |
| Thai | 13°42 N | 101°06 E | 80 | 28 | 1220 | |
| ChiL | 20°38 N | 110°31 E | 50 | 22.6 | 1473 | |
| ChiH | 20°55 N | 110°06 E | n.d. | 22.9 | 1701 | |
| Taiw | 22°30 N | 120°20 E | n.d. | n.d. | n.d. | |
| Melv | 11°34 S | 130°34 E | 20 | 25 | 1750 | |
| Fiji | 17°50 S | 178°20 E | 50 | 25 | 3000 | |
| Bdsh | 22°06 N | 92°05 E | n.d. | 24 | 2590 |
Tableau 4. Lots de semences testés à chaque site d'essai.
| SbA | SbB | SbC | SbD | Sfda | Sw1 | Sw2 | Sw3 | Sw4 | Sw5 | SK1 | SK2 | Thai | ChiL | ChiH | Taiw | Melv | Fiji | Bdsh | |
| 12990 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 |
| 12991 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 12992 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 |
| 13229 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 |
| 13230 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 |
| 13231 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 |
| 13232 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 |
| 13233 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 |
| 13234 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 |
| 13235 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 13236 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 |
| 13237 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 |
| 13238 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 13239 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 |
| 13240 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 |
| 13241 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 |
| 13242 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 |
| 13279 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 |
| 13459 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 |
| 13460 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 |
| 13534 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 13621 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 13622 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 |
| 13846 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
Tableau 5. Accroissement annuel moyen en hauteur et en diamètre: performances des différentes régions de provenance et moyenne générale sur chaque site d'essai.
| Site d'essais | AAM en hauteur (m) | AAM en diamétre MAI (cm) | ||||||||||
| CERAM | IRIAN | QCR | FNQ | PNG | MOY.# | CERAM | IRIAN | QCR | FNQ | PNG | MOY.# | |
| SbA | 3.80 | 3.60 | 3.13 | 3.64 | 3.30 | 3.35 | 3.10 | 2.78 | 3.30 | 2.92 | ||
| SbB | 2.50 | 2.38 | 2.16 | 2.29 | 2.23 | |||||||
| SbC | 4.15 | 3.99 | 3.92 | 4.19 | 4.22* | 3.98 | 4.04 | 3.80 | 3.91 | 4.46 | 4.30* | 3.97 |
| SbD | 3.52 | 3.31 | 3.44 | 3.69 | 3.46 | 3.80 | 3.37 | 3.76 | 4.48 | 3.79 | ||
| Sfoda | 2.68 | 2.98 | 3.27 | 2.98 | 2.85 | 2.96 | 3.23 | 2.98 | ||||
| Swk1 | 1.92 | 1.88 | 1.42 | 2.17 | 1.63 | 1.62 | 2.36 | 1.85 | 2.83 | 2.01 | ||
| Swk2 | 2.10 | 3.08 | 2.74 | 2.68 | 1.55 | 3.07 | 2.83 | 2.62 | ||||
| Swk3 | 1.80 | 2.13 | 2.11 | 2.05 | 1.70 | 2.13 | 2.33 | 2.16 | ||||
| Swk4 | 2.71 | 3.21 | 3.32 | 3.18 | 2.53 | 3.21 | 3.47 | 3.23 | ||||
| Swk5 | 3.43 | 3.52 | 3.34 | 3.40 | 2.93 | 3.54 | 3.19 | 3.21 | ||||
| SK1 | 3.62 | 3.58 | 4.44 | 3.88 | 3.26 | 3.29 | 4.64 | 3.73 | ||||
| Sk2 | 2.96 | 2.93 | 3.46 | 3.05 | 3.35 | 3.96 | 4.23 | 3.63 | ||||
| Thai | 3.06 | 2.92 | 3.33 | 3.44 | 3.02 | 4.77 | 5.09 | 5.56 | 5.84 | 5.20 | ||
| ChiL | 0.76 | 0.78 | 0.92 | 1.14 | 1.12 | 0.93 | 0.84 | 0.83 | 1.22 | 1.31 | 1.39 | 1.20 |
| ChiH | 1.54 | 2.10 | 2.22 | 2.28 | 2.07 | 1.46 | 2.15 | 2.15 | 2.60 | 2.12 | ||
| Taiw | 2.04 | 1.83 | 2.29 | 2.70 | 2.56 | 2.30 | 1.48 | 1.54 | 2.83 | 3.44 | 3.01 | 2.75 |
| Melv | 2.76 | 2.47 | 3.19 | 3.42 | 3.80 | 3.23 | ||||||
| Fiji | 1.76 | 1.39 | 2.29 | 2.80 | 2.81 | 2.26 | 1.73 | 1.19 | 2.34 | 2.91 | 3.07 | 2.31 |
| Bdsh | 0.84 | 0.96 | 1.33 | 1.02 | ||||||||
# Moyenne de toutes les provenances locales testés sur le site
Tableau 6. Pourcentage d'arbres à tronc unique sur trois sites d'essai, par provenance.
| Site d'essais | ||||||
| Lot de semences CSIRO | Provenance | Sabah SbC1 | Sabah SbD1 | Melville Island Australie2 | Coté d'Ivoire3 | |
| région | nom | |||||
| 12991 | QCR | Daintree | 34 | 18 | - | - |
| 12992 | QCR | Rex Range | 51 | 13 | 41 | - |
| 13229 | FNQ | Claudie River | 50 | 24 | 11 | 89 |
| 13232 | QCR | Cowley Beach | 44 | 22 | - | - |
| 13233 | QCR | Walshs Pyramid | 47 | 26 | - | 84 |
| 13234 | QCR | E. de Cairns | 17 | |||
| 13238 | QCR | Mission Beach | 48 | 17 | - | |
| 13240 | QCR | Cardwell-Ellerbeck | 10 | 77 | ||
| 13241 | QCR | Broken Pole Ck | 45 | 33 | - | 76 |
| 13459 | PNG | Morehead | - | - | 22 | |
| 13460 | PNG | Oriomo R | - | - | 12 | 75 |
| 13534 | QCR | Cassowary Range | 51 | 37 | - | - |
| 13864 | QCR | Mossman | 92 | |||
| 13621 | CERAM | Piru, Ceram | 42 | 23 | 1 | 56 |
| 13622 | IRIAN | Sidei | 66 | 33 | 12 | 69 |
| - | PNG | n.d.* | 47 | - | - | - |
| - | Sabah | (d'une plantation á Lahad Datu) | 68 | 28 | - | - |
1 Sim Boon Liang, comm. pers. 1990. Pourcentage d'arbre à tronc unique
2 M.W. Haines, comm. pers. 1990. Pourcentage d'arbre à tronc unique à 3.0 m de hauteur
3 Souvannavong (1990). Pourcentage d'arbre à tronc unique
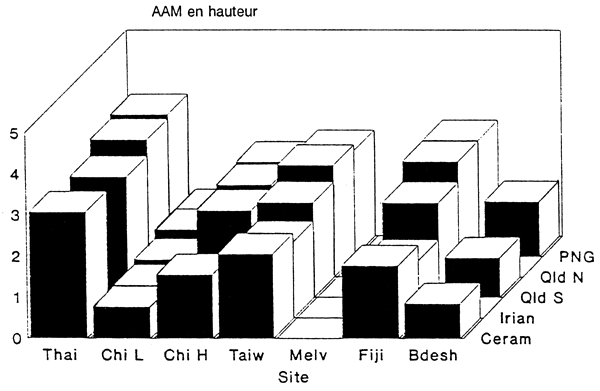
Figure 1. Accroissement annuel moyen en hauteur des différentes régions de provenance pour les sites d'essai autres que ceux de Bornéo.
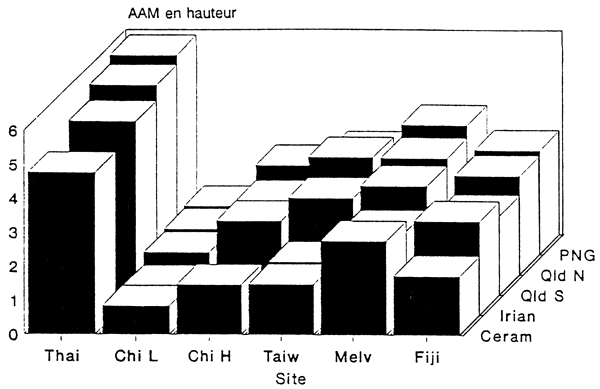
Figure 2. Accroissement annuel moyen en diamètre des différentes régions de provenance pour les sites d'essai autres que ceux de Bornéo.
1 Manuscrit reçu en juin 1991.
![]()
![]()
![]()