![]()
![]()
![]()
H.H. Shugart and T.M. Smith
Herman H. Shugart is W.W. Corcoran Professor and Chairman of the Department of Environmental Sciences, University of Virginia.
Thomas M. Smith is an assistant professor with the Department of Environmental Sciences. University of Virginia, USA.
Moving toward a systems analysis of the global boreal forest.
A boreal forest stand in southern Finland
The boreal forests represent one of the most important softwood timber resources on earth. These forests also represent the largest single pool of living organic carbon on the terrestrial surface. There is evidence that the productivity of the forest during the growing season is of sufficient magnitude to produce the annual oscillation in the global atmospheric carbon dioxide concentration. The forests are the habitat of important fur-bearing and game animals, rare and endangered species and other species of conservation interest. In their more isolated domains, boreal forests represent some of the last truly wilderness areas on Earth.
This multidimensional importance of the boreal forest has stimulated much discussion on the possible response of these forests to predicted environmental changes, especially those caused by "acid rain" and climate change associated with increased concentrations of atmospheric carbon dioxide.
Along with the expected year-to-year variations in weather and other normal environmental fluctuations, large areas of the boreal forests are in or near industrial regions and have been exposed to significant changes in air quality (SOx, NOx, ozone etc.) over the past several decades. Even in areas that are distant from industrial centres, the long-distance airborne transport of pollutants have resulted in lowered pH levels in precipitation.
On a global scale, the carbon dioxide concentration of the atmosphere is now about 25 percent greater than what are thought to be the preindustrial revolution levels. The climatic changes predicted by computer models of the Earth's "weather machine" (general circulation models or GCMs) in response to a doubling of atmospheric carbon dioxide appear to be biologically significant, at least at the higher northern latitudes (Shugart et al., 1986).
Uncertainty is one of the key words that arises in any discussions evaluating the possibility and impact of global climate change (Bolin et al., 1986). This includes the uncertainty of the magnitude of the increase in the Earth's carbon dioxide ambient as well as the temporal pattern of the projected increase, and the uncertainty of the predictions regarding regional patterns of climatic change produced by the GCMs (particularly with regard to the climatic variables that involve water - precipitation, cloudiness, soil moisture etc.).
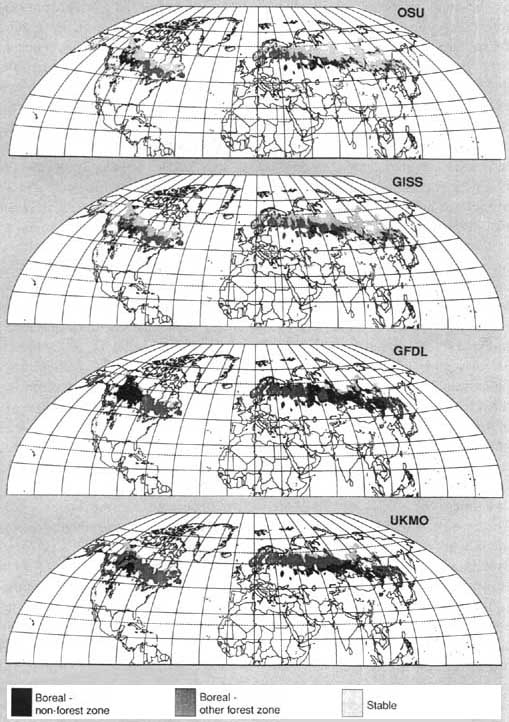
The Intergovernmental Panel on Climate Change concluded in its recent reports (1989a. 1989b) that climate change could result in poleward shifts in climate zones. which control forest species distribution, and potentially large losses from forest decline. The boreal forests are therefore one of the ecosystems with the greatest potential for being most affected (Fig. 1). The patterns of these changes will depend not only on the responses of individual species, but also on the constraints of land-use practices. Such forest responses could have major impacts on forest productivity, conservation management and availability of forest products.
Experiments and field studies on such long-term and large-scale processes are almost inevitably incomplete. Simulation models can be used to evaluate hypotheses generated by field studies and ecological experiments. and also in situations where the more traditional approach is less applicable; for example, for studies that span several research generations, such as the study of processes involved in forest succession. This potential accounts for the large number of computer models that have been developed to predict dynamic change in the boreal forest (for reviews, see Fries, 1974; Shugart, 1984; Shugart and Prentice, 1992).
What is the nature of a boreal forest model that can predict forest responses to changing environmental forces? No one model or approach can meet these requirements, and none will in the future, at least in the absence of new information on ecological effects of atmospheric contaminants, relationships of carbon storage and release to climate, and so on. However, one class of model that can be tested to some degree for climate change effects, and which can include the fundamental mechanisms in the carbon dioxide ambient response of plants, is that of forest dynamics based on the growth of individual trees.
Individual-based models of forest dynamics
The consideration of individual trees in models to project global ecosystem dynamics may at first seem paradoxical. However, it is the interaction between individual trees and their local environment that determines processes such as gap-phase regeneration which, in turn, crucially affect the composition, structure and texture of forests over large areas. These processes can be effectively simulated by individual-based models. Furthermore, data on the size distribution of individual trees are relatively easy to obtain, provide an important basis for testing dynamic models and have historically been used as the starting point for understanding large-scale forest processes.
The relative abundance of tree species in forests is a consequence of differences in regeneration success, the growth rates of individual trees in different environments and mortality rates. These, in turn, are influenced by a range of factors, some of which are controlled (at least in part) by the biology and ecology of the species involved. For example, the regeneration success of trees is actually a complex consequence of tree fecundity, germination patterns, seedling physiology and dispersal and a legion of other factors. Realistic forest simulation models can be used to investigate the relative sensitivity of birth, growth and death parameters in a variety of forests.
Individual trees attain sufficient size to locally alter their own micro-environment and that of subordinate trees. The species, shapes and sizes of trees in a forest can have a direct influence on the local forest environment. The environment, too, has a profound influence on the performance of different species, shapes and sizes of trees. Thus, there can be a feedback from the canopy tree to the local microenvironment and subsequently to the seedling and sapling regeneration that may become the next canopy.
The micro-environmental alteration caused by a canopy tree is perhaps most easily observed in the case of the forest light environment. The nature of the leaf - area profile and canopy geometry are dominant factors in the amount and nature of the light at the forest floor. Trees can also alter the local environment with respect to the nature of soil moisture, soil nutrient availability and other factors.
The potential importance of individual tree interaction in understanding forest environments implies that these considerations be taken into account when interfacing environmental issues with forest ecosystem dynamics. The expression, "unable to see the forest for the trees", used idiomatically to address an over attention to detail in the face of the obvious, comes to mind when one is attempting to predict larger-scale dynamics from small quantum behaviour. As will be discussed later, this fear is allayed to some degree by the impressive computational power of modern digital computers.
Computer models that simulate the dynamics of a forest by following the fates of each individual tree were first developed in the mid-1960s. Following the development of the earliest such model, by Newnham in 1964, foresters at several institutions proceeded to formulate further models of forest change that used a digital computer to dynamically modify a map of the sizes and positions of each tree in a forest. These models have become more and more popular as the computer power available has increased.
As Huston, DeAngelis and Post (1988) point out, an advantage of such models is that two assumptions, often used when modelling populations, are not necessary. They are:
the unique features of individuals are unimportant to the degree that individuals are identical; and the population is "perfectly mixed" so there are no local spatial interactions of any great magnitude.
These assumptions seem particularly inappropriate for trees.
Furthermore, there is a considerable body of information on the performance of individual trees (growth rates, establishment requirements, height/ diameter relations) that can be used directly in estimating the parameters of such models. Most individual tree models of forests are based on simple rules of interaction among individuals (e.g. shading, competition for limited resources etc.) in conjunction with simple rules of birth, death and growth of individuals.
Individual-based forest gap models
To illustrate the performance of individual tree models and their application in the prediction of changes in boreal forest dynamics in response to changing environmental conditions, we will discuss forest gap models.
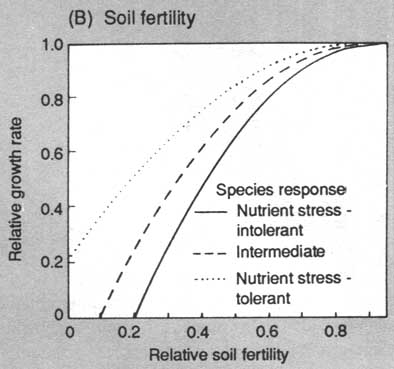
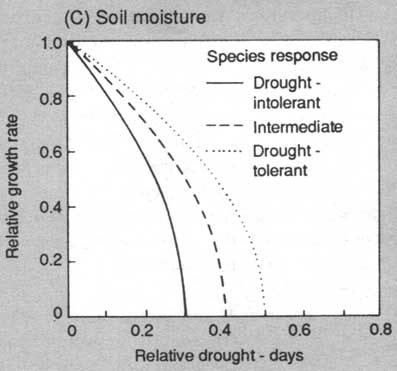
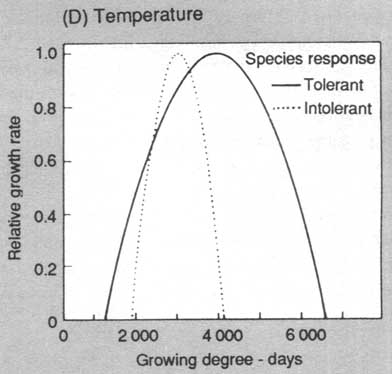
FIGURE 2. Response functions used to modify annual diameter growth of trees for the environmental constraints of (A) light, (B) soil fertility, (C) soil moisture (D) temperature. Most species can be assigned to response categories for light, nutrients and moisture; temperature responses are based on geographic ranges of species
Gap models are ecological versions of earlier forestry models based on individual tree dynamics. Individual - based forest gap models simulate the annual establishment, diameter growth and mortality of trees on a defined plot. The general approach has been applied to a wide variety of forested systems around the world, including boreal forests in North America and Eurasia. Although the models differ in their inclusion of specific processes pertinent to the dynamics of the particular sites being simulated (in the case of boreal forest: permafrost formation, effects of light angle, competition between mosses and tree seedlings etc.), they share a common set of characteristics.
Model structure. In gap models, each plant is recognized as a unique entity with respect to the processes of establishment, growth and mortality. This allows computation of species- and size-specific changes in each tree. The precise location of each individual tree is not considered in most gap models. The assumption is that each individual on a plot of a given size is influenced by (and influences) the growth of all other individuals on that plot. The spatial scale of the models is an area corresponding to the zone of influence of a single maximum-size individual. The vertical structure of the canopy is modelled explicitly. The sizes of individuals (diameter, height and leaf area) are used to construct a vertical leaf - area profile. Light extinction is computed and the light environment for each individual is calculated.
Environmental constraints and resource competition. All gap models simulate individual tree response to the light availability at different height intervals on the plot. Other resources are incorporated to varying degrees in different versions of the model. In extant boreal forest models, these other constraints include soil moisture, fertility and temperature as well as disturbances such as fire, harvest and windthrow. It is through these constraints also that the effects of new stresses, such as air pollution, or potentially beneficial changes, such as carbon dioxide fertilization, can be added to the models. These may be incorporated in a variety of ways, including alterations in growth rates, regeneration success or mortality rates of individual trees. These expected responses must be generated by running more detailed models or from field observations. Frequently, the projected responses to environmental change are developed as possible scenarios and the models are used to determine how a forest might change as a consequence of alterations in tree-level biology.
Environmental responses are modelled via a "constrained potential" paradigm in which a tree has a maximum potential behaviour under optimal conditions (maximum diameter increment, survival or establishment rate). This optimum is then reduced according to the environmental context of the plot (shading, drought) to give the behaviour under specific ambient conditions (See Fig. 2).
Competition in the model is indirect and depends on the relative performance of different trees under the environmental conditions of the model plot. These conditions may be influenced by the trees themselves (e.g. a tree's leaf area influences the light available beneath it) or may be modelled as extrinsic and not influenced by the trees (e.g. temperature). Competition may operate in two modes.
Competition for light is asymmetric: a tree at a given height absorbs light and reduces the resource available to trees at lower positions in the canopy. In most gap models, competition for below-ground resources (water and nutrients) is symmetric: each tree experiences the same resource level common to the plot. Thus, competition depends strictly on the context of the modelled gap: a tree that has the best performance relative only to other trees on the plot is the most successful. Competitive success depends on the environmental conditions of the plot, the species present and the relative sizes of the trees; each of these varies through time in the model.
Modelling tree biology
Growth. The growth of an individual is calculated using a function that predicts an expected diameter increment under optimal conditions, given a tree's current diameter. This optimum increment is then modified by the environmental response functions (described earlier), and the realized increment - an annual ring - is added to the tree.
Mortality, The death of an individual is modelled as a stochastic or chance process. Most gap models have two components of mortality: age-related and stress-induced. The age-related component applies equally to all individuals of a species and depends on the maximum expected longevity for the species with a typical annual mortality rate of about I to 2 percent. Stress is defined with respect to a minimal diameter increment (typically 10 percent of the optimum); individuals failing to meet this minimal condition are subject to an elevated mortality rate.
Establishment. Tree establishment and regeneration rates are also largely stochastic, with maximum potential establishment rates constrained by the same environmental factors that modify tree growth. Each simulation year, a pool of potential recruits is filtered through the environmental context of the plot and new individuals are established.
Fir (Abies spp.)

Whole-plant response and ecophysiological processes
The individual -based gap models outlined here simulate the annual growth of trees using a species-specific optimal growth curve. This diameter change can be re-expressed as annual diameter growth and then modified as a function of environmental conditions by using a series of functions relating percentage maximum growth to resource availability (light, water, nutrients and temperature see Fig. 2). The environmental conditions are represented as an integrated index on an annual basis.
This approach is in contrast to the more physiological process models which may function on an hourly or daily time-scale and relate environmental conditions at these scales to the processes of photosynthesis and transpiration at either the leaf- or whole-canopy level. The two approaches have been reconciled by using physiological models to generate the annual responses of whole trees. This integration, summarized as a response function similar to those currently used in the model, is then used in a gap model to extrapolate the whole tree response to larger-scale conditions at the community level over longer periods. One outcome of these later experiments has been to gauge increased forest water yields in response to a doubled carbon dioxide level in the atmosphere and the alteration in the water use efficiencies of individual trees.
Poplar (Populus spp.)

Thus, gap models should not be expected to predict the long-term dynamics of a small forest plot, although the average of many simulations may predict the behaviour of a larger area. This aspect of gap models is quite realistic; there are good reasons to expect that replicate small plots of land, even under ideal simulated environmental conditions, would not show the same successional pattern, whereas the average behaviour of an ensemble of plots is likely to be more consistent.
Testing a boreal forest simulator
The testing of simulation models able to project long-term forest responses under changing conditions has generally involved inspecting a model's ability to reproduce historical data, data from stands of a known age and patterns of forest associated with environmental gradients. One important test that has been used in several cases is the direct comparison of forest yield tables referring to different intervals of time with equivalent output models. Gap models have been used for the prediction of species composition changes over time-spans as long as 18000 years, along various environmental gradients, in response to different disturbance regimes and in response to stress and disease. In such tests, several model plots are averaged to estimate stand-level characteristics.
Because these models produce data in the same form as forest inventory data (the diameter and species of each tree on a plot of a known size), the model output can be compared with samples from actual forest stands to obtain a direct comparison of average tree diameters, basal areas and stocking densities for stands of a known age. In most tests, the model predictions lie within the natural range of variation seen in actual forest stands. One can also use the models to reconstruct larger forest landscape patterns (including variations in the soil, slope, aspect and age of stands). These predictions can then be compared with forest patterns seen in actual locations.
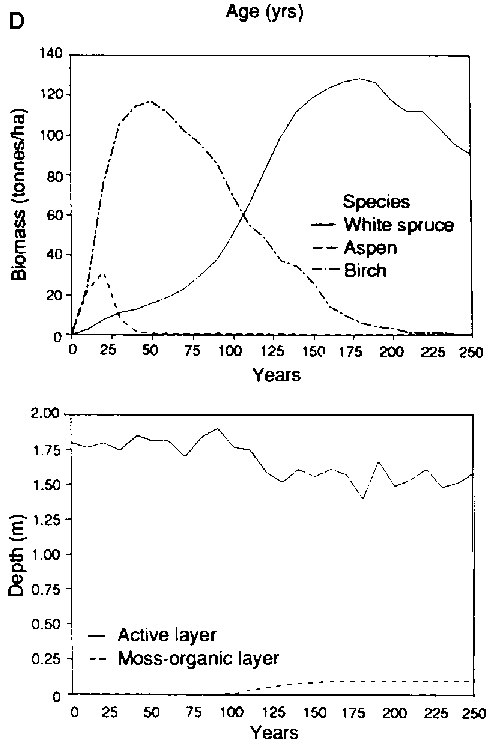
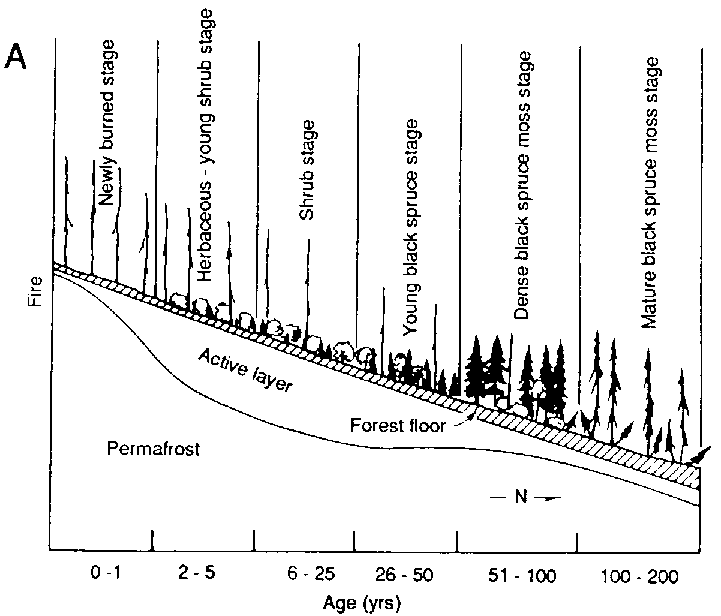
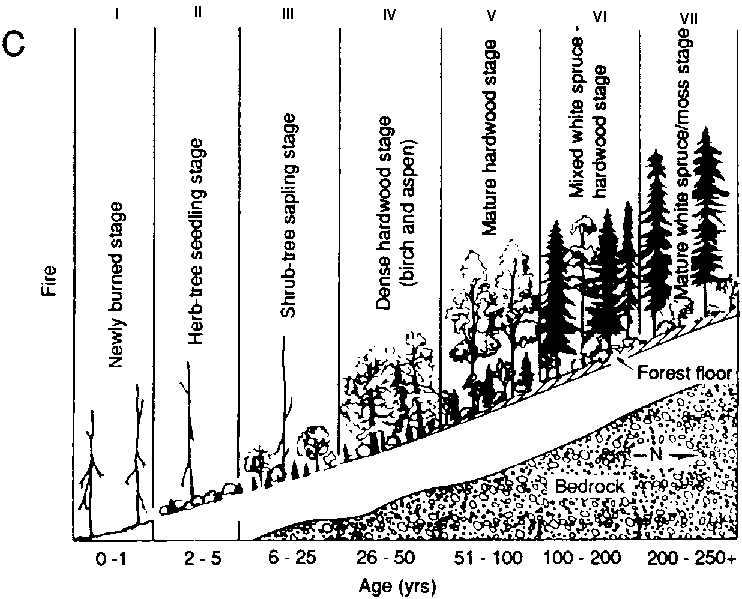
As an example, Fig. 3 illustrates one of the testing protocols for Bonan's model of forest structure and succession for North American boreal forests. Bonan (1989) and Bonan and Korzukin ( 1989) simulated forest succession and permafrost dynamics of naturally regenerated forests on both north-facing and south-facing slopes under environmental conditions appropriate to Fairbanks, Alaska. Fairbanks is in the "zone of discontinuous permafrost". This region features permafrost formations and successions with black spruce (Picea mariana) on north-facing slopes, and a more complex birch (Betula spp.), aspen (Populus spp.) and white spruce (Picea glauca) succession on permafrost-free south-facing slopes (Bonan and Shugart, 1989). The model was tested for its ability to reproduce the landscape pattern, the composition and structure of specific stands of different ages on different slopes as well as the presence, absence and depth of permafrost under diverse conditions (Fig. 3). Bonan also inspected the ability of the model to reproduce stand structure in a widely dispersed set of locations throughout North America (Bonan, Shugan and Urban, 1990; see also Pastor and Post, 1988).
There is a clear need to continue the development of computer models capable of simulating the responses of forests to changing environmental stress on both local and global scales. Cooperative efforts are emerging at the international level to construct computer models already developed. These models can be used to project forest change and synthesize new information and insights as they are further developed. Such models represent an important step forward in international forest ecology research and in our ability to predict forest responses to global change.
FIGURE 3. Testing a boreal forest simulator for Its ability to reproduce the structure, landscape pattern and succession of forests in the vicinity of Fairbanks, Alaska (A) Successional pattern of forests on north-facing slopes following a forest fire. The figure Is arranged as a chronosequence with the age of the forest increasing from left to right (a) Simulated response of forests (biomass by species; depth of the active, unfrozen soil layer; depth of the moss and organic matter foyer) over successional time (C) Successional pattern of forests on south-facing slopes following e forest fire (D) Simulated response of forests over successional time (biomass by species; depth of the active, unfrozen soil layer; depth of the moss and organic matter layer)
FIGURE 3 - B
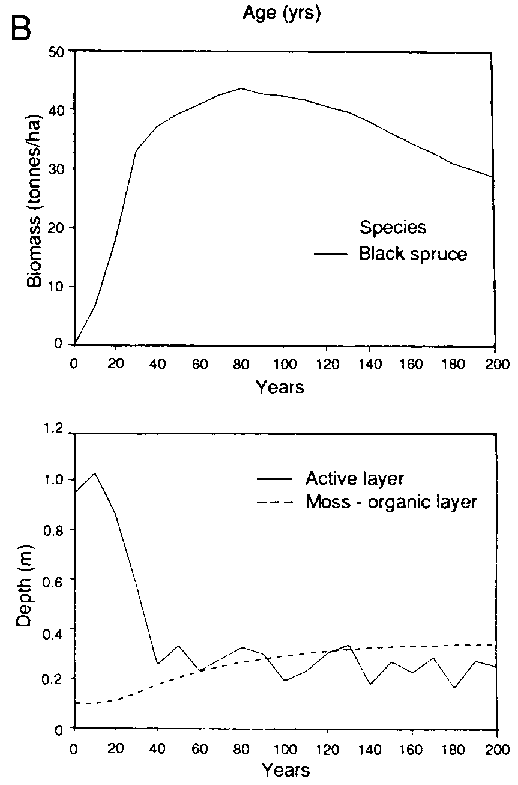
FIGURE 3 - D
For example, since 1984 there has been an international effort to develop a "unified boreal forest model" capable of predicting the dynamic change of North America's and Eurasia's northern forests in response to environmental stress and climate change (Shugart, Leemans and Bonan, 1992). The model represents the continuing collaboration of more than 40 scientists from all the nations with boreal forest resources and from many countries with significant alpine forests (e.g. Switzerland and Bulgaria). Much of the model was developed by coordinating the doctoral dissertations of Dr G.B. Bonan (University of Virginia) and Dr R. Leemans (Uppsala University). We have presented results from the North American version of the resultant "unified boreal zone model" in the section above.
The development of a unified simulator for a major forest biome at the global level has several important implications. Originally, the model (Shugart, Leemans and Bonan, 1992) was designed to evaluate how one of the most important terrestrial ecosystems might respond as a whole to global climate change. The boreal forest - particularly its wood and soils - is a major terrestrial carbon reservoir.
In addition, the model represents a uniform synthesis of a large body of knowledge (the book describing the formulation of the model includes a bibliography of more than 1400 citations). The model is valuable for developing research priorities and sharing knowledge regarding boreal forests It is available without charge and is intended for widespread application in a range of forestry problems. The designers of the unified boreal simulator intend the present model to be iteratively improved as comparative studies are carried out on different parts of the boreal forests.
Ågren, G.I., McMurtrie, R.E., Parton, W.J., Pastor, J. & Shugart, H.H. 1991. State - of the - art models of production- decomposition in conifer and grassland ecosystems. Ecol. Appl., 1:118-138.
Bolin, B., Doos, B.R., Jager, J. & Warrick, R.A., eds. 1986. The greenhouse effect, climatic change and ecosystems, p. 207-270. Chichester, John Wiley.
Bonan, G.B. 1989. Environmental factors and ecological processes controlling vegetation patterns in boreal forests. Landscape Ecol., 3: 111-130.
Bonan, G.B. & Korzukin, M.D. 1989. Simulation of moss and tree dynamics in the boreal forest of interior Alaska. Vegetation, 84: 31-44.
Bonan, G.B. & Shugart, H.H. 1989. Environmental factors and ecological processes in boreal forests. Annu. Rev. Ecol. Syst., 20: 1-28.
Bonan, G.B., Shugart, H.H. & Urban, D.L. 1990. The sensitivity of some high-latitude boreal forests to climatic parameters. Climatic Change, 16: 9-29.
Fries, J., ed. 1974. Growth models for tree and stand simulation. Res. notes 30. Stockholm, Royal College of Forestry.
Holdridge, L.R. 1967. Life zone ecology, San lose Costa Rica, Tropical Science Center.
Huston, M., DeAngelis, D.& Post, W.M. 1988. New computer models unify ecological theory. Bioscience, 38(10): 682-691.
IPCC. 1989a. Land-use management: adaptive responses to climate change. Working Paper for the Resource Use and Management Subgroup of Working Group 3.
IPCC. 1989b. Biological diversity: adaptive responses to climate change. Working Paper for the Resource Use and Management Subgroup of Working Group 3.
Pastor, J. & Post, W.M. 1988. Response of northern forests to CO2 - induced climate change. Nature, 334(6177): 55-58.
Shugart, H.H. 1984. A theory of forest dynamics: the ecological implications of forest succession models. New York. Springer-Verlag. 278 pp.
Shugart, H.H., Antonovsky, M. Ya, Jarvis, P.G. & Sandford, A.P. 1986. CO2, climatic change and forest ecosystems. In B. Bolin, B.R. Doos J. Jager &: R.A. Warrick, eds. The greenhouse effect, climatic change and ecosystems. p. 475-521. Chichester John Wiley.
Shugart, H.H., Leemans, R. & Bonan, G.B., eds. 1992. A systems analysis of the global boreal forest. Cambridge, Cambridge Univ. Press. 542 pp.
Shugart, H.H. & Prentice, I.C. 1992. Individual tree-based models of forest dynamics and their application in global change research. In H.H. Shugart, R. Leemans & G.B. Bonan, eds. A systems analysis of the global boreal forest, p. 313-333. Cambridge, Cambridge Univ. Press.
Smith, T.M., Leemans, R. & Shugart, H.H. 1991. Sensitivity of terrestrial carbon storage to CO2 - induced climate change: comparison of five scenarios based on general circulation models. Climatic Change, 17. (in press)
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}