![]()
![]()
![]()
G.M. MacDonald
Glen M. MacDonald est professeur adjoint au Département de géographie de l'Université McMaster d'Hamilton (Canada).
Les prélèvements de pollen fossile provenant de lacs et tourbières permettent d'identifier le profil à long terme de la modification de la fréquence des essences et l'abondance relative de ces dernières dans les forêts boréales.
Le bon aménagement des forêts boréales aux fins de la conservation et de l'utilisation durable des ressources demande une surveillance approfondie des forêts qui permette de détecter et de maîtriser les effets des modifications naturelles et artificielles du milieu. C'est peut-être le changement climatique de la planète dû aux activités humaines qui menace le plus gravement et de la manière la plus étendue la forêt boréale (Emanuel, Shugart et Stevenson, 1985; Kauppi et Posch, 1985, 1988). La réaction de la végétation aux changements de vastes étendues naturelles qui pourraient être déclenchés par le changement climatique pourrait comporter également une modification de la structure des groupements végétaux, un déplacement de l'aire d'extension géographique des espèces végétales et des modifications de la structure génétique des populations.
Cependant, les données dont on dispose sur l'état actuel des forêts boréales sont limitées à la fois dans le temps et dans l'espace. L'utilisation d'images obtenues par satellite, une connaissance approfondie de la réalité de terrain et des études de peuplements peuvent permettre d'élargir la couverture spatiale de ces données. Mais, dans le meilleur des cas, l'étendue temporelle reste de l'ordre de 50 à 100 ans, c'est-à-dire moins que la durée de vie potentielle de la plupart des essences de la forêt boréale. L'absence de données à long terme soulève deux questions. Premièrement, a-t-on besoin de données à long terme? Deuxièmement, existe-t-il une source d'indicateurs susceptibles de remplacer rétrospectivement ces données? La réponse à la première question est évidente. La végétation que l'on trouve aujourd'hui dans la zone boréale se développe depuis la fin de la dernière période glaciaire, il y a environ 10000 ans. Des données de surveillance sur une période de 50 ans représentent un échantillon de 1/200e de cette durée. Etant donné que la vie moyenne des essences boréales se situe entre 80 et 400 ans et que les incidences de modifications naturelles du milieu ne se font sentir qu'au bout d'une génération au moins, il ne fait aucun doute que ces données d'observation ne permettront pas à elles seules de détecter d'éventuelles modifications progressives mais significatives de la forêt.
En s'efforçant de répondre à la deuxième question, le présent article décrit d'abord la manière dont les prélèvements de pollen fossile provenant de lacs et tourbières permettent d'identifier le profil à long terme de la modification de la fréquence des essences et l'abondance relative des essences dans les forêts boréales. Il tire quelques importantes conclusions de ces données concernant l'évolution de la structure des groupements d'essences des forêts boréales, le rythme et le profil des changements de la répartition géographique des essences boréales et la distribution géographique de la diversité génétique.
Les essences arborées boréales dominantes (Picea, Pinus, Larix, Abies, Betula et Populus) sont pollinisées par le vent et produisent de grandes quantités de pollen facilement transporté. L'essentiel de ce pollen ne remplit jamais sa fonction de reproduction et va en partie se déposer dans les lacs et les tourbières où, à l'abri de l'air, la partie extérieure des grains de pollen peut se conserver indéfiniment. Les paléo-écologistes ont des protocoles bien établis d'analyse du pollen et des spores provenant des lacs et tourbières. Divers types de dispositifs de carottage sont utilisés pour prélever des sections verticales des sédiments des lacs et tourbières. Les échantillons de sédiments sont traités à l'aide d'une série de réactifs chimiques qui digèrent la plus grande partie de la matière minérale et organique tout en laissant intacts le pollen et les spores. Les grains sont ensuite identifiés et comptés au microscope.
FIGURE 1
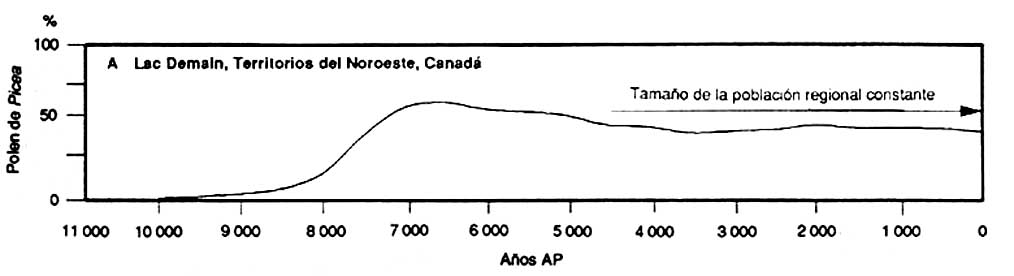
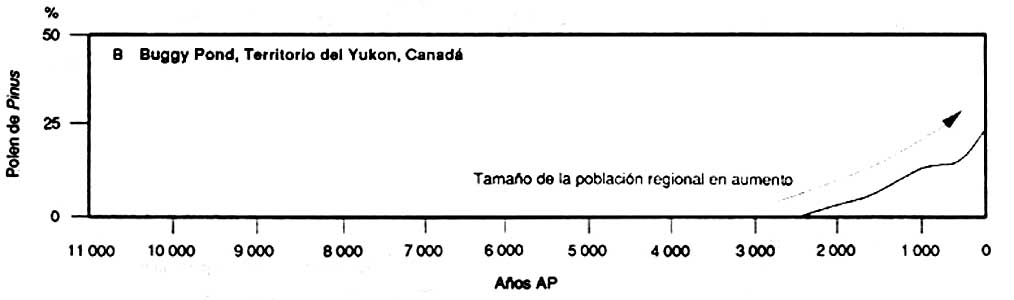
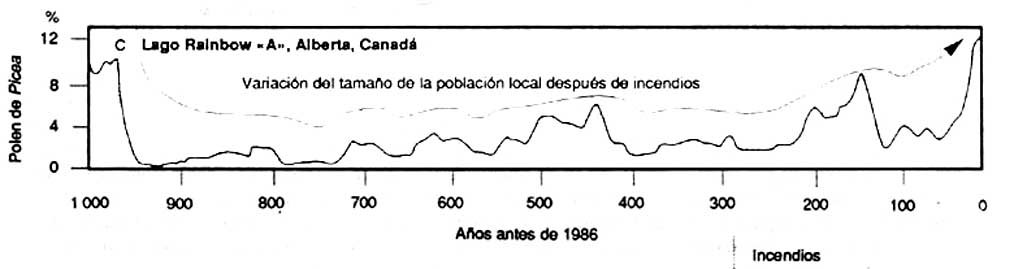
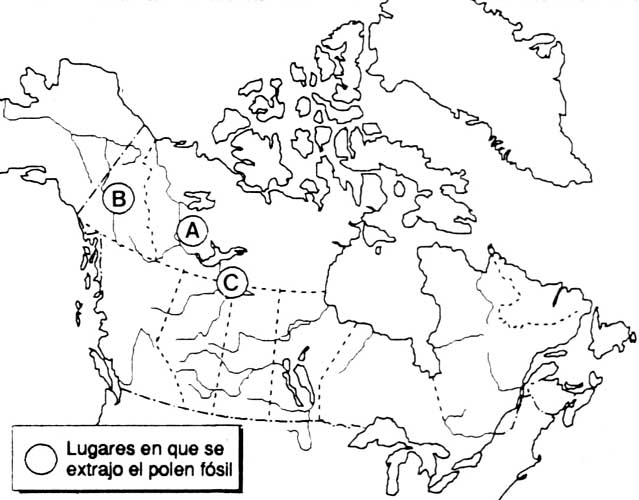
Les informations que fournissent les études de pollen fossile peuvent permettre de mieux comprendre l'évolution a long terme de la composition de la forêt boréale, contrairement aux autres sources d'information. Les pollens fossiles provenant de petits lacs de l'ouest des Territoires du Nord-Ouest (A) montrent que les effectifs de la population régionale de Picea spp. ont été relativement constants pendant les 4000 à 5000 dernières années; les données provenant des pollens fossiles du centre du Yukon (B) montrent que les populations de Pinus spp. ont rapidement progressé pendant les 2000 dernières années; les pollens fossiles d'un très petit lac du nord de l'Alberta (C) indiquent que l'abondance de Picea spp. dans la végétation locale a varié après des incendies
A Lac Demain, Territoires du Nord-Ouest, Canada
B Buggy Pond, Territoire du Yukon, Canada
C Rainbow Lac «A». Alberta. Canada
Sites des prélèvements de pollen fossile
Le pollen de la plupart des essences boréales peut permettre de remonter au genre, mais dans certains cas, par exemple pour Picea glauca et P. mariana, le pollen est suffisamment particulier pour permettre d'identifier l'espèce (Hansen et Engstrom, 1985). Le contrôle chronologique des prélèvements de pollen est généralement assuré par datation au radiocarbone.
Les proportions du mélange de végétation sont difficiles à déterminer à partir de prélèvements de pollen fossile. Des genres tels que Picea et Pinus, qui produisent de grandes quantités de pollen, sont en général sur-représentés dans les prélèvements de pollen par rapport à des genres tels que Abies et Larix, qui en produisent de plus petites quantités. L'étendue dont provient l'échantillon représenté dans les prélèvements de pollen fossile tirés de sédiments de lacs est inversement liée à la vitesse d'implantation du grain de pollen et directement proportionnelle au rayon du lac ou de la tourbière où l'échantillon a été prélevé (Prentice, 1988).
Le pollen de Pinus spp. est généralement léger et facilement transporté par les vents de haute altitude. On a calculé que la zone dont proviennent 70 pour cent du pollen de Pinus spp. déposé dans un petit lac (300 m de rayon) peut s'étendre dans un rayon de 59 km du site (Prentice, 1988). En revanche, la zone dont proviennent 70 pour cent du pollen plus lourd produit par Picea spp. peut s'étendre dans un rayon de 2 km seulement autour du même lac (Prentice, 1988). Etant donné que le pollen déposé dans les lacs donne un tableau complet de l'état de la végétation dans une zone allant de quelques dizaines à des milliers de km2, les qualités d'échantillonnage spatial des prélèvements de pollen ont été rapprochées des données de télédétection (Webb, Laseki et Bernabo, 1978). En ce qui concerne les images obtenues par satellite, chaque pixel fournit un échantillon complet de la réflectance spectrale de la végétation et d'autres types de couverture dans une zone de 100 m2 à plus d'un million de m2. Cependant, les prélèvements de pollen peuvent permettre de suivre en permanence l'état de la végétation dans la zone dont provient l'échantillon sur des milliers d'années, tandis que les données obtenues par satellite ne remontent qu'aux années 60.
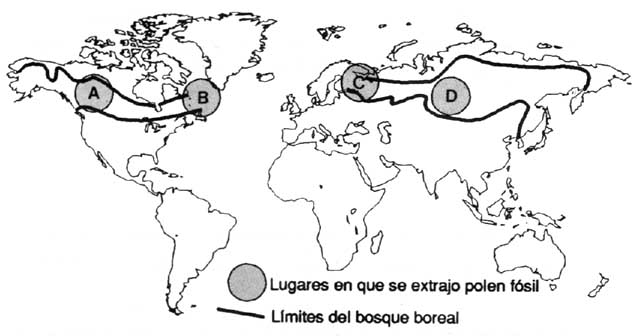
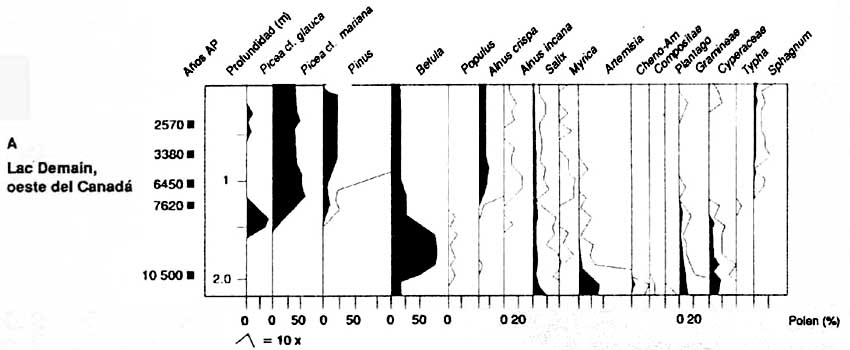
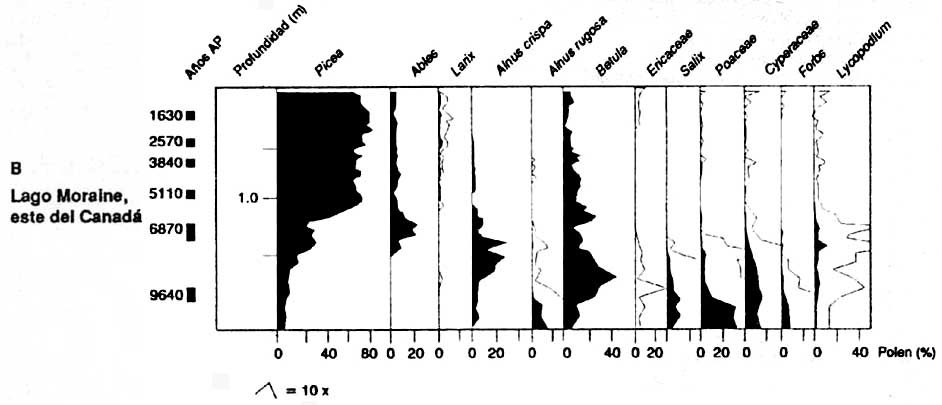
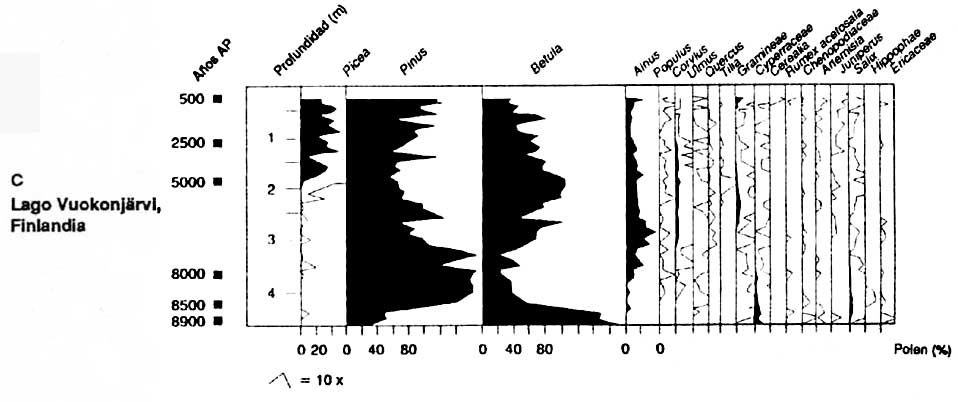
Des prélèvements de pollen fossile provenant de l'ouest et de l'est du Canada, de la Finlande et de la Sibérie (figure 2) donnent une perspective de l'évolution des groupements végétaux boréaux après la dernière période glaciaire, il y a environ 10000 ans. Ces prélèvements de pollen montrent que la forêt boréale évolue de manière très différente dans chacune de ces régions.
Le premier genre d'arbres à apparaître dans les prélèvements de pollen postglaciaire dans la région boréale de l'ouest du Canada est Populus (figure 2A MacDonald, 1987). Picea glauca et P. mariana sont devenus dominants dans la végétation entre 9000 et 8000 ans BP (année BP = 14C années avant 1950). Betula papyrifera est également devenu une composante importante de la forêt à ce moment-là. Le dernier genre d'arbres à s'implanter dans la région a été Pinus (P. contorta spp. latifolia et P. banksiana). Les pins sont devenus d'importantes composantes de la végétation entre 7000 et 4000 ans BP. Dans le nord-est du Canada, la première végétation de la région boréale méridionale a été caractérisée par des graminées et buissons (figure 2B - Engstrom et Hansen, 1985). Le genre Picea n'a dominé qu'à partir de 8000 à 7000 ans BP, P. mariana remplaçant P. glauca comme essence dominante vers 6000 ans BP. Abies balsamea est également devenu une composante importante de la végétation à ce moment-là.
Dans l'est de la Finlande (figure 2C Huttunen et Stober, 1980), Betula spp. dominait la végétation en 9000 ans BP. Pinus sylvestris est devenu l'essence arborée prédominante dans les prélèvements de pollen en 8500 BP. Picea abies n'est devenu une importante composante de la végétation qu'en 5000 ans BP.
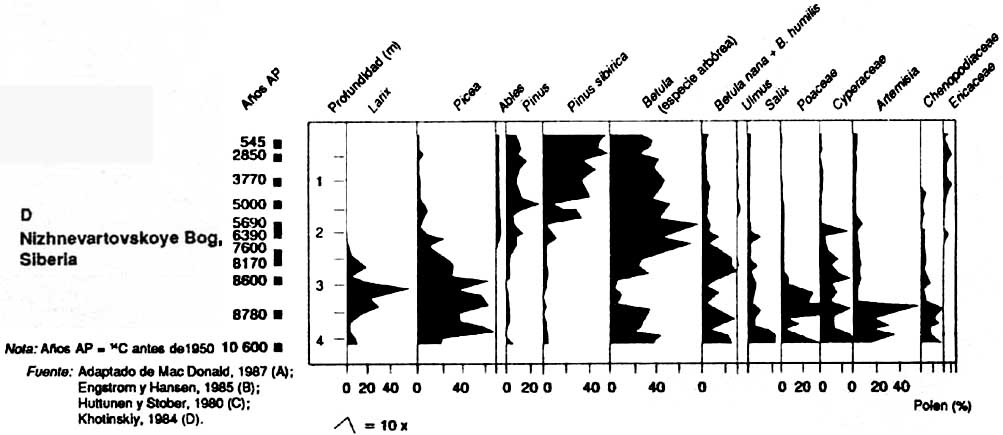
La première végétation postglaciaire dans l'ouest de la Sibérie (figure 2D Khotinskiy, 1984) comprenait les genres Larix et Picea ainsi que les essences arborées de Betula. Entre 8000 et 5000 ans BP, les genres Abies et Pinus ont beaucoup progressé aux dépens de Larix et de Picea.
A Lac Demain, Territoire du Nord-Ouest, Canada
Tourbière de Nizhnevartovskoye, Sibérie
FIGURE 3
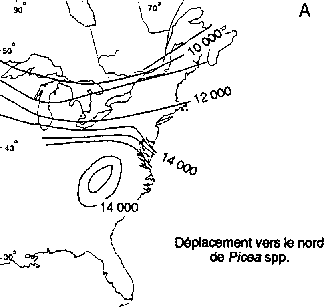
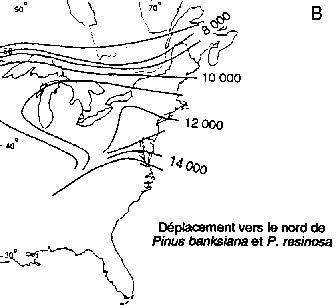
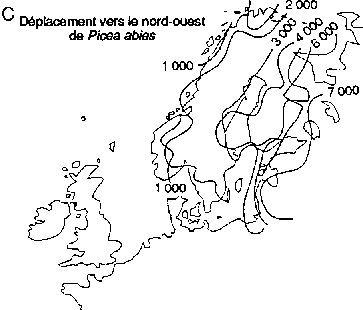
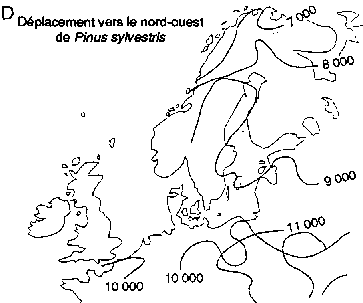
Déplacement vers le nord des genres Picea et Pinus dans la région boréale actuelle de l'est de l'Amérique du Nord (A, B); déplacement vers le nord-ouest de Picea abies et de Pinus sylvestris en Fenno-Scandie (C. D)
Déplacement vers le nord de Picea spp.
Déplacement vers le nord de Pinus banksiana et P. resinosa
Déplacement vers le nord-ouest de Picea abies
Déplacement vers le nord-ouest de Pinus sylvestris
Ces données montrent clairement que, dans toutes ces régions, la forêt boréale a évolué pour prendre sa forme actuelle pendant les 5000 dernières années. Il est à noter que les quatre sites qui ont fait l'objet de prélèvements se trouvent au centre de la forêt boréale et sont relativement inactifs au point de vue de l'évolution de la végétation postglaciaire par rapport à des sites de la périphérie méridionale et septentrionale. En Amérique du Nord, en Europe et en Asie, les bordures de la forêt boréale ont subi une modification marquée de la végétation due aux variations climatiques pendant les 10000 dernières années (Hyvarinen, 1976; Khotinskiy, 1984; Ritchie, 1987). Les facteurs pouvant avoir conditionné l'évolution de la forêt pendant la période postglaciaire restent à débattre. Cependant, les travaux récents laissent penser qu'à des échelles temporelles de milliers d'années et à des échelles spatiales sous-continentales, le changement de climat a probablement été le principal facteur qui a déterminé les rythmes et modes d'évolution de la forêt boréale (Prentice, Bartlein et Webb, 1991).Il est donc aussi probable que la réaction de la végétation boréale au changement climatique prévu sera très variable selon les régions au point de vue des modes et des rythmes d'évolution des groupements végétaux.
Des réseaux géographiques des sites de pollen fossile ont été utilisés pour établir des reconstructions cartographiques des modifications passées des aires d'extension des essences boréales en Europe de l'Ouest et en Amérique du Nord (figure 3). L'aspect le plus significatif de ces modifications a été la remontée vers le nord de certaines essences après le dernier réchauffement postglaciaire. Ces données sont importantes pour l'aménagement de la forêt boréale, et cela pour deux raisons.
Premièrement, les données que fournit le pollen fossile montrent les rythmes auxquels diverses essences, ayant certains degrés de variation intraspécifique, peuvent modifier leur aire d'extension à la suite de modifications du milieu naturel. Les estimations de la vitesse maximale des déplacements vers le nord de Picea spp. après le dernier réchauffement postglaciaire vont de 376 m/an dans l'est de l'Amérique du Nord (Delcourt et Delcourt, 1987) à 500 m/an dans le nord-ouest de l'Europe (Huntley et Birks, 1983). La vitesse maximale de migration des pins du Nord (Pinus banksiana, P. strobus et P. resinosa) dans l'est de l'Amérique du Nord a été estimée à 613 m/an (Delcourt et Delcourt. 1987). Dans le nord-ouest de l'Europe, la vitesse maximale de migration de P. sylvestris a été estimée à 1500 m/an (Huntley et Birks, 1983). Davis (1989) a fait remarquer que bien que ces vitesses de migration semblent rapides, le changement climatique imputable à l'effet de serre demandera peut-être une modification des aires d'extension à des rythmes de 3000 m/an.
Mélèze (Larix spp.)

Deuxièmement, les données que fournit le pollen mettent en évidence de grandes différences interspécifiques de la vitesse de propagation vers le nord des essences boréales. Dans l'est et dans l'ouest de l'Amérique du Nord (figure 3A), Picea mariana et P. glauca ont réussi à occuper une bonne partie de leur aire d'extension actuelle assez rapidement après le réchauffement postglaciaire (Davis, 1981; Delcourt et Delcourt, 1987; MacDonald, 1987). En revanche, l'arrivée de Pinus spp. à ses limites septentrionales de l'est de l'Amérique du Nord a été beaucoup plus tardive (figure 3B). Pinus contorta spp. latifolia migre probablement encore vers le nord dans le nord-ouest du Canada (MacDonald et Cwynar, 1985). En Fenno-Scandie (Danemark, Finlande, Norvège, Suède), Pinus sylvestris a gagné ses limites actuelles relativement tôt (figure 3D) après la fin de la période glaciaire. En revanche, Picea abies n'a atteint ses limites actuelles du nord-ouest en Scandinavie que dans les 500 dernières années (figure 3C). On connaît encore mal les facteurs qui ont régi ces vitesses de migration intra et interspécifiques. L'une des explications que l'on avance pour ces phénomènes à l'échelle du continent est que chaque essence a réagi de manière très individualiste aux changements climatiques en raison des différences génétiques et des degrés d'adaptabilité (Huntley et Webb, 1989). Cela laisse penser que le changement climatique futur pourrait entraîner des ajustements individualistes analogues des limites de l'aire d'extension des essences arborées boréales.
Il y a longtemps que les généticiens forestiers étudient les données relatives au pollen fossile, afin de mieux comprendre les relations qui existent entre l'histoire des essences arborées boréales au pléistocène et la répartition actuelle des variations génétiques de ces essences.
Les profils de diversité génétique des genres nord-américains tels que Picea, Pinus et Abies et l'évolution divergente d'essences voisines telles que Pinus contorta et P. banksiana ont été attribués à la perturbation des aires d'extension provoquée par les glaciations du quaternaire (Critchfield, 1984) qui a séparé les populations autrefois contiguës entre l'est et l'ouest, le sud et le nord. Cependant, des recherches récentes menées sur les pollens fossiles dans l'ouest du Canada ont permis de conclure que le faible degré de variabilité génétique qui caractérise les populations de Pinus contorta spp. latifolia du Yukon ne peut être attribué à l'isolement des pins du Yukon pendant le dernier maximum de glaciation. En effet, les données que fournissent les pollens fossiles montrent que Pinus contorta n'est apparu au Yukon qu'il y a environ 4000 ans (MacDonald et Cwynar, 1985).
Il est possible que la faible variabilité génétique des populations du Yukon tienne à la façon dont les pins ont migré vers le nord après la fin de la dernière glaciation. La migration des pins vers le nord dans l'ouest du Canada a probablement été accomplie par l'implantation de petits groupements d'«éclaireurs». A leur tour, ces petites populations isolées ont probablement fourni les semences nécessaires à la croissance de la population à l'extrême nord de l'aire d'extension de ces essences et à l'implantation de nouveaux groupements d'éclaireurs. Les pertes aléatoires de variabilité génétique lices à chacun de ces événements sont peut-être à l'origine du gradient de variabilité génétique décroissante vers le nord observé pour Pinus contorta spp. latifolia (Cwynar et MacDonald, 1987). Ce mode de migration a peut-être aussi entraîné une sélection pour les semences légères, faciles à transporter, qui caractérisent les populations de pins du Yukon (MacDonald et Cwynar, 1985).
Les recherches décrites plus haut ont deux incidences importantes pour la compréhension de la modification de la structure génétique des forêts boréales. D'abord, la différence génétique considérable entre les populations du Yukon et les populations plus méridionales s'est constituée sur une période de moins de 4000 ans. Cela laisse penser que les différences significatives et étendues de structure génétique peuvent apparaître assez rapidement sans que les aires de végétation ne soient perturbées par des glaciations. Ensuite, si la perte de diversité génétique ou la sélection des semences pour leur légèreté a été due au mode de migration vers le nord, des pertes analogues et/ou des changements morphologiques ont toute probabilité d'être observés si les aires d'extension changent rapidement à la suite du réchauffement futur de la planète.
Les données tirées de l'analyse des pollens fossiles permettent d'identifier les variations des associations végétales et de l'abondance relative des essences dans la végétation forestière boréale sur plusieurs millénaires. Les propriétés de détection spatiale de ces données peuvent être, en gros, comparées à celles de données de télédétection fournies par les satellites. Les prélèvements de pollen fossile sont une importante source de données de base qui permettent d'interpréter l'évolution des forêts boréales.
Les groupements végétaux des forêts boréales ont acquis leur configuration actuelle dans les 5000 dernières années ou moins. Il existe d'importantes différences régionales dans les modes d'évaluation des groupements boréaux. L'évolution future des forêts boréales peut varier selon les régions et donner des structures de groupements sans équivalent actuel.
Les rythmes et modes d'évolution des aires d'extension des essences boréales depuis la fin de la dernière période glaciaire montrent qu'il existe une très grande variabilité interspécifique. Cette variabilité tient à des réactions très individualistes des essences au milieu. Les réactions des essences arborées à l'évolution future de l'environnement seront probablement très variables d'une essence à l'autre.
Depuis la dernière glaciation, il s'est produit pour certaines essences arborées des modifications observables de la répartition à grande échelle de la diversité génétique. Peut-être l'évolution future de l'environnement favorisera-t-elle des pertes considérables de diversité génétique des espèces végétales boréales.
Critchfield, W.B. 1984. Impact of the Pleistocene on the genetic structure of North American conifers. Proc. North American Forest Biology Workshop, 8: 70-118.
Cwynar, L.C. & MacDonald, G.M. 1987. Geographical variation of lodgepole pine in relation to population history. Am. Nat., 129: 463-469.
Davis, M.B. 1981. Quaternary history and stability of forest communities. In D.C. West, H.H. Shugart & D.B. Botkin, eds. Forest succession: concepts and application. New York, Springer-Verlag.
Davis, M.G. 1989. Lags in vegetation response to greenhouse warming. Climatic Change, 15: 7582.
Delcourt, P.A. & Delcourt, H.R. 1987. Long-term forest dynamics in the temperate zone. New York, Springer-Verlag.
Emanuel, W.R., Shugart, H.H. & Stevenson, M.P. 1985. Climate change and the broad-scale distribution of terrestrial ecosystem complexes. Climatic Change, 7: 29-43.
Engstrom, D.R. & Hansen, B.C.S. 1985. Postglacial vegetation change and soil development in southeastern Labrador as inferred from pollen and chemical stratigraphy. Can. J. Bot., 63: 543-561.
Hansen, B.C.S. & Engstrom, D.R. 1985. A comparison of numerical and qualitative methods of separating pollen of white and black spruce. Can. J. Bot., 63: 2159-2163.
Huntley, B. & Birks, H.J.B. 1983. An atlas of past and present pollen maps for Europe: 0-13000 years ago. Cambridge, Cambridge Univ. Press.
Huntley, B. & Webb III, T. 1989. Migration: species, response to climatic variations caused by changes in the Earth's orbit. J. Biogeog., 16: 5-19.
Huttunen, P. & Stober, J. 1980. Dating of palaeomagnetic records from Finnish lake sediment cores using pollen analysis. Boreas, 9: 193-202.
Hyvarinen, H. 1976. Flandrian pollen deposition rates and tree line history in northern Fennoscandia. Boreas, 5: 163-175.
Kauppi, P. & Posch, M. 1985. Sensitivity of boreal forests to possible climatic warming. Climatic Change, 7: 45-54.
Kauppi, P. & Posch, M. 1988. A case study of the effect of CO2 - induced climatic warming on forest growth and the forest sector: A. Productivity reactions of northern boreal forests. In M.L. Parry, T.R. Carter & N.T. Konjin, eds. The impacts of climatic variations on agriculture, p. 183- 195. Dordrecht, Kluwer Academic Publishers.
Khotinskiy, N.A. 1984. Holocene vegetation history. In A.A. Velichko, H.E. Wright, Jr. & C.W. Barnosky, eds. Late Quaternary environments of the Soviet Union, p. 179-200. Minneapolis, Univ. Minnesota Press.
MacDonald, G.M. 1987. Postglacial vegetation history of the Mackenzie River basin. Quaternary Res., 28: 245-262.
MacDonald, G.M. & Cwynar, L.C. 1985. A fossil pollen-based reconstruction of the late Quaternary history of lodgepole pine (Pinus contorta ssp. latifolia) in the western interior of Canada. Can. J. Forest. Res., 15: 1039-1044
Prentice, I.C. 1988. Records of vegetation in time and space: the principles of pollen analysis. In B. Huntley & T. Webb III, eds. Vegetation history, p. 17-42. Dordrecht, Kluwer.
Prentice, I.C., Bartlein, P.J. & Webb III, T. 1991. Vegetation and climate in eastern North America since the last glacial maximum. Ecology, 72: 2038-2056.
Ritchie, J.C. 1987. Postglacial vegetation of Canada. Cambridge, Cambridge Univ. Press.
Webb III, T., Laseki, R.A. & Bernabo, J.C. 1978. Sensing vegetational patterns with pollen data: choosing the data. Ecology, 59: 1151-1163.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}