![]()
![]()
![]()
H.H. Shugart et T.M. Smith
Herman H. Shugart est professeur (chaire W.W. Corcoran) et directeur du Département des sciences de l'environnement à l'Université de Virginie.
Thomas M. Smith est professeur assistant au Département des sciences de l'environnement de l'Université de Virginie (Etats-Unis).
Vers une analyse fonctionnelle de l'ensemble de la forêt boréale
Peuplement forestier boréal dans le sud de la Finlande
Les forêts boréales constituent l'une des plus grandes sources de bois de conifères de la terre. Elles représentent aussi la plus vaste réserve de carbone organique vivant à la surface du globe. Il est établi que la productivité de la forêt pendant sa période végétative est d'une amplitude suffisante pour expliquer la variation annuelle de la concentration de dioxyde de carbone dans l'atmosphère. Les forêts sont l'habitat de nombreux animaux à fourrure, de gibier, d'espèces rares ou menacées, ainsi que d'autres espèces dont la préservation est souhaitable. Les régions les plus reculées des forêts boréales sont parmi les rares dernières véritables étendues sauvages du globe.
Cette importance multidimensionnelle de la forêt boréale a suscité beaucoup de discussions quant à l'évolution possible de ces forêts face aux modifications escomptées de l'environnement, plus particulièrement sous l'effet des «pluies acides» et du changement climatique associé aux concentrations croissantes de CO2 dans l'atmosphère. Outre qu'elles subissent les fluctuations météorologiques prévisibles d'une année à l'autre ainsi que les variations normales de l'environnement, d'importantes parties de la forêt boréale sont situées dans des régions industrielles ou à proximité de celles-ci et ont été exposées à des changements significatifs de la qualité de l'air (SOx, NOx, ozone, etc.) durant les dernières décennies. Les longues distances parcourues dans l'air par les matières polluantes ont entraîné une diminution du pH des précipitations, même dans les régions éloignées des centres industriels.
A l'échelle mondiale, la concentration de CO2 dans l'atmosphère est actuellement supérieure de quelque 25 pour cent au niveau qu'elle aurait atteint à l'époque précédant la révolution industrielle. Les changements climatiques prévus par les modèles informatiques de la «machine météorologique» de la terre (modèles de la circulation générale ou MCG), sous l'effet du doublement du CO2 dans l'atmosphère, semblent être biologiquement significatifs, pour le moins dans les latitudes élevées de l'hémisphère nord (Shugart et al., 1986).
«Incertitude» est un des mots-clés qui revient dans chaque débat sur l'éventualité et l'impact d'un changement climatique à l'échelle planétaire (Bolin et al., 1986). Cette incertitude porte, entre autres, sur l'amplitude de l'accroissement du CO2 ambiant et la répartition temporelle de l'accroissement projeté, et sur les prévisions quant à la structure régionale du changement climatique faites à partir de modèles de la circulation générale (notamment en ce qui concerne les variables climatiques se rapportant à l'eau précipitations, nébulosité, humidité du sol, etc.).
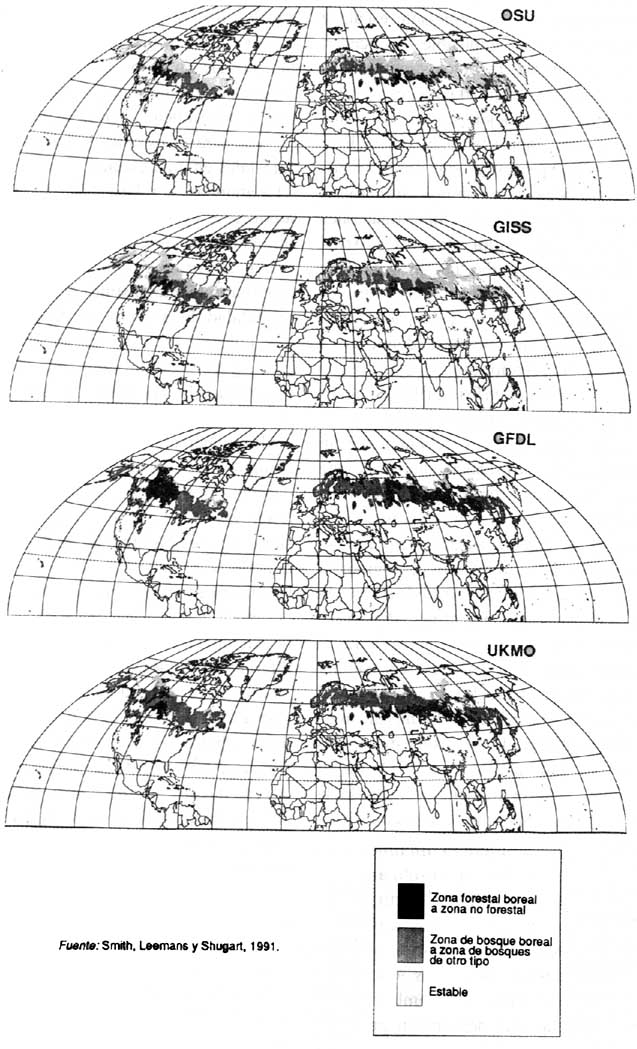
Le Groupe d'experts intergouvernemental pour l'étude du changement climatique a souligné, dans des rapports récents, (IPCC, 1989a, 1989b) que le changement de climat pourrait entraîner le déplacement vers les pôles de zones climatiques qui influent sur la répartition des espèces forestières et, partant, un déclin considérable de la forêt. Ainsi, les forêts boréales sont un des écosystèmes qui risquent d'être le plus touchés (figure 1). La forme que prendront ces changements dépendra non seulement de la réaction de chaque espèce individuelle, mais aussi des contraintes liées aux modes d'utilisation des terres. La réaction des forêts pourrait avoir des incidences considérables sur la productivité forestière, l'aménagement pour la conservation et l'approvisionnement en produits forestiers.
Les vérifications expérimentales et les études faites sur le terrain sont presque inévitablement incomplètes lorsqu'il s'agit de processus à long terme d'une telle ampleur. Des modèles de simulation peuvent être utilisés pour évaluer les hypothèses engendrées par des recherches sur le terrain et des expérimentations écologiques, et aussi dans des situations où une approche plus traditionnelle est moins applicable, par exemple les études qui couvrent des recherches sur plusieurs générations, telle l'étude des processus liés au renouvellement de la forêt. Ce potentiel explique le grand nombre de modèles informatiques qui ont été conçus pour prévoir le changement dynamique de la forêt boréale (Fries, 1974; Shugart, 1984; Shugart et Prentice, 1992).
Comment doit se présenter une modélisation de la forêt boréale susceptible de prévoir les réactions de la forêt aux forces modifiant l'environnement? En réalité, il n'existe pas de modèle ou d'approche qui puisse répondre à ces exigences, et il n'y en aura pas à l'avenir, du moins tant que l'on n'en saura pas plus sur les effets écologiques des polluants atmosphériques, sur les effets climatiques de la rétention du carbone et de sa libération, et ainsi de suite. Toutefois, une des modélisations que l'on peut vérifier dans une certaine mesure pour les effets du changement climatique, et qui peut intégrer les mécanismes fondamentaux de la réponse des plantes au CO2 ambiant, est celle de la dynamique des forêts fondée sur la croissance des arbres individuels.
Modélisation de la dynamique des forêts basée sur l'individu
La prise en compte des arbres individuels dans un modèle visant à projeter la dynamique globale d'un écosystème peut, de prime abord, sembler paradoxale. Toutefois, c'est l'interaction des arbres individuels avec leur environnement local qui détermine des processus comme la phase de régénération de la trouée qui, à son tour, affecte d'une manière fondamentale la composition, la structure et la texture des forêts sur une vaste étendue. Ces processus peuvent être effectivement simulés par des modèles basés sur les individus. En outre, les données sur la répartition de la taille des arbres individuels sont relativement faciles à obtenir; elles fournissent une source importante d'information pour tester les modèles dynamiques et ont été utilisées historiquement comme point de départ pour comprendre l'évolution de la forêt sur une grande échelle.
L'abondance relative des espèces d'arbres dans les forêts est la conséquence des différences au niveau du succès de la régénération, au niveau des taux de croissance des arbres individuels dans des environnements différents, et au niveau des taux de mortalité. Ceux-ci, à leur tour, sont influencés par un ensemble de facteurs, dont certains dépendent (du moins en partie) de la biologie et de l'écologie des espèces considérées. Par exemple, le succès de la régénération des arbres est en fait le résultat complexe de la fécondité de l'arbre, des modes de germination, de la physiologie des jeunes plants, de la dispersion et de quantité d'autres facteurs. Des modèles réalistes de simulation de forêts peuvent être utilisés pour étudier la sensibilité relative des paramètres de naissance, de croissance et de mortalité d'une variété de forêts.
Des arbres individuels atteignent une taille suffisante pour affecter localement leur propre micro-environnement et celui des arbres subordonnés. Les espèces, les formes et les tailles des arbres de la forêt peuvent avoir une influence directe sur l'environnement forestier local. L'environnement, à son tour, exerce une influence profonde sur la performance des différentes espèces, la forme et la taille des arbres. Ainsi, l'arbre à étage dominant peut avoir une influence sur le micro-environnement local et, par conséquent, sur le développement du jeune plant qui pourra former le couvert forestier successif.
La modification du micro-environnement sous l'effet d'un arbre à étage dominant peut sans doute être observée le plus facilement dans le cas de l'environnement lumineux de la forêt. La nature du profil de la surface des feuilles et la configuration de la cime sont les facteurs dominants qui agissent sur la quantité et la nature de la lumière parvenant jusqu'au sol forestier. Les arbres peuvent aussi modifier l'environnement local en agissant sur l'humidité du sol et sa capacité nutritive, et sur d'autres facteurs.
Pour comprendre les environnements forestiers, l'importance potentielle des interactions des arbres individuels doit être prise en compte lorsque l'on cherche à établir l'interface entre les facteurs environnementaux et la dynamique de l'écosystème forestier. L'expression populaire «l'arbre qui cache la forêt», utilisée pour souligner l'importance accordée au détail au détriment de l'évidence, vient à l'esprit lorsque l'on tente de prévoir une dynamique à grande échelle à partir d'un petit quantum de comportement. Mais, comme il sera mentionné par la suite, cette crainte est, dans une certaine mesure, modérée par la puissance impressionnante des ordinateurs numériques modernes.
Les premiers modèles informatiques qui simulent la dynamique d'une forêt en fonction de l'évolution de chaque arbre individuel ont vu le jour au milieu des années 60. Dans la foulée de la première modélisation du genre, créée par Newnham en 1964, les forestiers des différentes institutions ont mis au point de nouveaux modèles de changement forestier utilisant un ordinateur numérique pour modifier dynamiquement la carte des tailles et des emplacements de chaque arbre dans une forêt. L'utilisation de ces modèles s'est développée au fur et à mesure qu'augmentait la capacité des ordinateurs.
Comme le font remarquer Huston, De Angelis et Post (1988), l'avantage de tels modèles est que souvent deux hypothèses de modélisation des populations ne sont pas nécessaires. Ces hypothèses sont les suivantes:
· les particularités uniques des individus sont tellement peu différenciées que les individus semblent identiques;· la population est «parfaitement mélangée» de sorte qu'il n'y a pas d'interactions spatiales locales d'importance significative.
Ces hypothèses semblent particulièrement inappropriées dans le cas des arbres.
En outre, il existe une masse d'informations sur les performances des arbres individuels (taux de croissance, conditions d'implantation, relations hauteur/diamètre) qui peuvent être utilisées directement pour évaluer les paramètres de tels modèles. La plupart des modélisations de forêts basées sur l'arbre individuel sont fondées sur de simples règles d'interactions entre les individus (par exemple, ombre, concurrence pour les ressources limitées, etc.) associées des règles élémentaires sur la naissance, la mort et la croissance des individus.
Modélisation de la forêt basée sur l'individu
Pour illustrer la performance des modèles basés sur des arbres individuels et leurs applications dans la prévision du changement de la dynamique des forêts boréales, sous l'effet d'une modification de l'environnement, nous examinerons les modélisations de type «gap model».
Les «gap models» sont des versions écologiques des premiers modèles forestiers basés sur la dynamique des arbres individuels. Les modélisations de la forêt basées sur les individus simulent les données annuelles concernant l'implantation, la croissance du diamètre et la mortalité des arbres sur une parcelle définie. Cette approche générale a été appliquée à une large variété de systèmes forestiers dans le monde, y compris les forêts boréales d'Amérique du Nord et d'Eurasie. Bien que les modèles diffèrent en ce qui concerne l'introduction de processus importants dans les dynamiques de sites particuliers simulés (dans le cas de la forêt boréale: la formation de permafrost, les effets de l'incidence de la lumière, la concurrence entre les mousses et les jeunes plants, etc.), ils ont en commun un certain nombre de caractéristiques.
Structure des modèles. Dans les «gap models», chaque plante est reconnue comme une entité unique pour ce qui est de l'implantation, de la croissance et de la mortalité. Cela permet l'estimation des espèces et des changements de taille spécifiques des arbres. L'emplacement précis de chaque arbre individuel n'est pas considéré dans la plupart des «gap models». On suppose que chaque individu sur une parcelle d'une taille donnée est influencé par la croissance de tous les autres individus de la parcelle (et réciproquement). L'échelle spatiale sur laquelle les modèles fonctionnent est une surface correspondant à la zone d'influence d'un seul individu de taille maximale. La structure verticale du couvert est simulée d'une manière explicite. Les tailles des individus (diamètre, hauteur et superficie des feuilles) sont utilisées pour construire le profil vertical de la surface du feuillage. La perte de lumière est également calculée par l'ordinateur, et l'environnement lumineux est calculé pour chaque individu.
Contraintes de l'environnement et concurrence pour les ressources. Tous les «gap models» simulent la réponse d'un arbre individuel à la quantité de lumière disponible à plusieurs intervalles de hauteur sur la parcelle. D'autres ressources sont intégrées à des degrés variables dans différentes versions du modèle. Dans les modélisations existantes de la forêt boréale, ces autres contraintes comprennent l'humidité du sol, la fertilité, la température, aussi bien que les perturbations comme les feux, les coupes et les arbres abattus par le vent. C'est aussi à travers ces contraintes que les effets de nouvelles agressions comme les dommages provoqués par la pollution de l'air, ou de changements potentiellement bénéfiques comme l'enrichissement en CO2, peuvent être rajoutés aux modélisations. Ces facteurs peuvent être introduits de différentes manières, notamment en modifiant les taux de croissance, la réussite de la régénération ou le taux de mortalité des divers arbres. Les réactions escomptées sont calculées à partir de modèles plus détaillés ou d'observations sur le terrain. Fréquemment, les réactions éventuelles à un changement de l'environnement sont explicitées sous forme de scénarios possibles, et les divers modèles sont utilisés pour déterminer comment une forêt pourrait évoluer en fonction des modifications au niveau biologique de l'arbre.
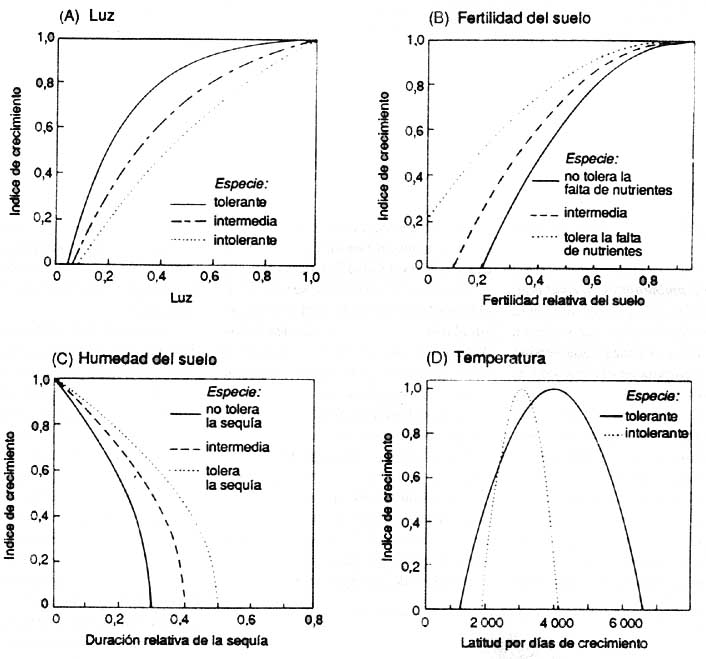
Les réactions à l'environnement sont simulées au moyen d'un paradigme de «potentiel limité» selon lequel un arbre a un comportement à potentiel maximal dans des conditions optimales (à savoir, augmentation maximale du diamètre, taux de survie ou d'implantation). Cet optimum est ensuite réduit en fonction du contexte environnemental de la parcelle (par exemple, ombre, sécheresse) pour arriver à un comportement en rapport avec le milieu ambiant (figure 2).
La concurrence dans le modèle est indirecte et dépend de la performance relative des différents arbres selon les conditions environnementales sur la parcelle type. Ces conditions peuvent être influencées par les arbres eux-mêmes (ainsi, la surface du feuillage d'un arbre influe sur la lumière disponible au-dessous), ou bien elles peuvent être considérées comme des variables exogènes (par exemple, la température). La compétition peut s'opérer de deux manières. La compétition pour la lumière est asymétrique: l'arbre à une hauteur donnée absorbe la lumière et réduit la ressource disponible pour les arbres situés aux niveaux inférieurs du couvert forestier. Dans la plupart des «gap models», la concurrence pour les ressources du sous-sol (eau, éléments nutritifs) est symétrique: chaque arbre dispose du même niveau commun de ressources que toute la parcelle. Ainsi, la concurrence dépend strictement du contexte de la trouée modélisée: seul l'arbre qui a la meilleure performance par rapport aux autres arbres de la parcelle est le plus prospère. Le succès, dans ce cas, dépend des conditions écologiques de la parcelle, des espèces présentes et de la taille relative des arbres; chacun de ces facteurs évolue avec le temps dans le modèle.
Modélisation de la biologie de l'arbre
Croissance. La croissance d'un individu est estimée à partir d'une fonction de · croissance qui prévoit, à partir du diamètre réel de l'arbre, l'augmentation escomptée dans des conditions optimales. Cet accroissement optimal est ensuite modifié par les fonctions de réaction à l'environnement (décrites ci-dessus), et l'accroissement effectif - correspondant au cerne annuel - est ajouté à l'arbre.
Mortalité. La mortalité des individus est définie de façon stochastique ou aléatoire. La plupart des modèles ont deux composantes de mortalité: l'une relative à l'âge et l'autre relative aux agressions. La composante relative à l'âge est appliquée de manière égale à tous les individus d'une espèce et dépend du maximum de longévité espérée pour cette espèce, avec un taux de mortalité annuel caractéristique pouvant varier de 1 à 2 pour cent. Les agressions sont définies en fonction du diamètre minimal de croissance (généralement 10 pour cent de l'optimum). Les individus qui n'atteignent pas ce minimum sont sujets à un taux élevé de mortalité.
Implantation. L'implantation des arbres et leur régénération sont aussi très largement stochastiques, avec des taux d'implantation potentiels maximaux limités par les mêmes facteurs environnementaux qui modifient la croissance de l'arbre. A chaque année simulée correspond un ensemble de recrues potentielles qui sont «filtrées» à travers le contexte écologique de la parcelle, et de nouveaux individus sont installés.
Sapin (Abies spp.)

Réaction globale de la plante et processus écophysiologiques
Les «gap models» basés sur les individus décrits ci-dessus simulent la croissance annuelle des arbres en utilisant les courbes optimales de croissance spécifique à chaque essence. Ce changement de diamètre peut être réexprimé sous forme de diamètre de croissance annuelle et ensuite modifié en fonction des conditions de l'environnement, au moyen d'une série de fonctions établissant une corrélation entre le pourcentage de croissance maximale et la disponibilité des ressources (lumière, eau, éléments nutritifs et température; voir figure 2). Les conditions environnementales sont représentées sous forme d'indice intégré sur une base annuelle.
Cette approche se distingue des modèles davantage basés sur le processus physiologique qui peuvent fonctionner sur une base horaire ou journalière et qui mettent en relation les conditions environnementales correspondantes avec les processus de photosynthèse et de transpiration au niveau du feuillage ou de l'ensemble du couvert. Les deux approches ont été conciliées par l'utilisation de modèles physiologiques pour donner les réactions annuelles de l'arbre dans sa totalité. Cette intégration, résumée comme une fonction de réaction similaire à celles utilisées couramment dans le modèle, est ensuite appliquée à un «gap model» pour extrapoler la réaction globale de l'arbre aux conditions plus larges au niveau de la communauté sur des périodes plus longues. Ces dernières expérimentations ont permis, entre autres, de mesurer l'accroissement de la production d'eau des forêts en réponse au doublement du niveau de CO2 dans l'atmosphère et la modification des capacités d'utilisation de l'eau par les arbres individuels.
Ainsi, on n'attend pas des «gap models» qu'ils prévoient à long terme la dynamique d'une petite parcelle de forêt, bien que la moyenne de nombreuses simulations puisse permettre de prévoir le comportement d'une plus grande surface. Cet aspect des modèles est tout à fait réaliste: il y a de bonnes raisons de penser que, même dans des conditions idéales de simulation de l'environnement, la réplication de petites parcelles de terre ne donnerait pas un schéma identique de succession, alors que le comportement moyen d'un ensemble de parcelles est probablement plus équilibré.
Test de simulateur de forêt boréale
Le test des modèles de simulation conçus pour projeter l'évolution à long terme des forêts sous l'effet d'un changement de conditions a généralement consisté à étudier la capacité des modèles à reproduire des données historiques, des données provenant de peuplements d'âge déterminé et des comportements forestiers associés aux variations de l'environnement. Un des tests importants qui a été utilisé dans plusieurs cas consiste en une comparaison directe des tableaux du rendement forestier établis à différents intervalles de temps avec les résultats équivalents du modèle. Les «gap models» ont été utilisés pour prévoir les changements de composition des espèces sur des périodes de temps allant jusqu'à 18000 ans, en fonction des variations de l'environnement, par rapport à différents régimes de perturbation et par rapport aux contraintes et aux maladies. Dans de tels tests, on fait la moyenne de plusieurs parcelles types pour estimer les caractéristiques au niveau des peuplements.
Peuplier (Populus spp.)

Etant donné que ces modèles fournissent des données qui se présentent sous la même forme que celles des inventaires forestiers (diamètre et espèce de chaque arbre sur une parcelle de taille déterminée), les résultats qu'ils produisent peuvent être comparés à des échantillons tirés de peuplements forestiers réels, afin d'obtenir une comparaison directe du diamètre moyen des arbres, des aires de base et des densités de peuplement pour des massifs forestiers d'âge connu. Dans la plupart des tests, les prévisions de la simulation restent dans les limites naturelles des variations relevées dans les peuplements réels. On peut aussi utiliser la modélisation pour reconstruire un schéma de paysage forestier plus grand (y compris la variation des sols, des pentes, des aspects ou de l'âge des peuplements). Ces prévisions peuvent être ensuite comparées à des structures forestières existantes.
Ainsi, la figure 3 représente un des protocoles de test pour le modèle de Bonan de structure et d'évolution forestière pour les forêts boréales de l'Amérique du Nord. Bonan (1989) et Bonan et Korzukin (1989) ont simulé l'évolution de la forêt et la dynamique du permafrost de forêts régénérées naturellement sur les pentes tant nord que sud dans des conditions environnementales correspondant à celles de Fairbanks (Alaska). Fairbanks est dans la «zone de permafrost discontinu». Cette région est caractérisée par une formation de permafrost et des successions d'épicéa noir (Picea mariana) sur les pentes de la face nord, et une succession plus complexe de bouleaux (Betula spp.), peupliers (Populus spp.) et épicéa blanc (Picea glauca) sur les pentes de la face sud sans permafrost (Bonan et Shugart, 1989). Le modèle a été testé pour mesurer sa capacité de reproduire le type de paysage forestier, la composition et la structure de peuplements spécifiques d'âges variés sur des pentes différentes, ainsi que la présence, l'absence et la profondeur du permafrost sous diverses conditions (figure 3). Bonan a aussi étudié la capacité du modèle à reproduire la structure d'un peuplement dans divers emplacements largement dispersés en Amérique du Nord (Bonan, Shugart et Urban, 1990; voir aussi Pastor et Post, 1988).
FIGURE 3
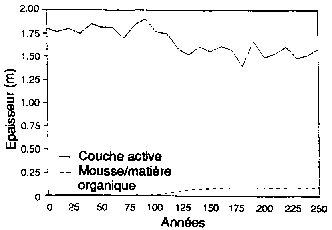
Expérimentation d'un simulateur de forêt boréale pour sa capacité a reproduire la structure, les types de paysage et la succession des forêts, aux alentours de Fairbanks (Alaska). (A) Succession de forêts sur les pentes de la face nord après un feu de forêt Le schéma est présenté comme une série de séquences chronologiques, l'âge de la forêt croissant de gauche a droite. (B) Réponse simulée des forêts (biomasse par espèces, épaisseur de la couche de sol active non gelée; épaisseur de la couche de mousse et de matière organique) à des moments successifs. (C) Types successifs de forêts sur les pentes de la face sud après un feu de forêt. (D) Réponses simulées de forêts à des moments successifs (biomasse par espèces; épaisseur de la couche de sol active non gelée; épaisseur de la couche de mousse et de matière organique) à des moments successifs.
Figura A - Age (années)
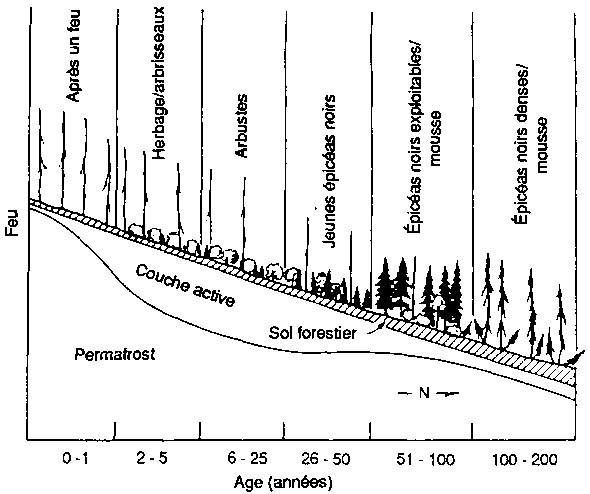
Figura B1 - Biomasse (tonnes/ha)
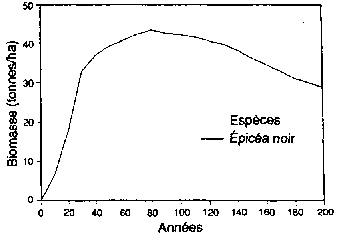
Figura B2 - Épaisseur (m)
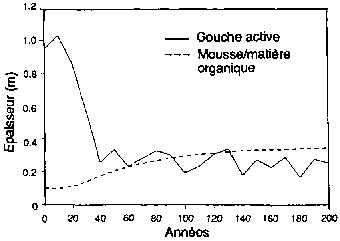
Figura C - Age (années)
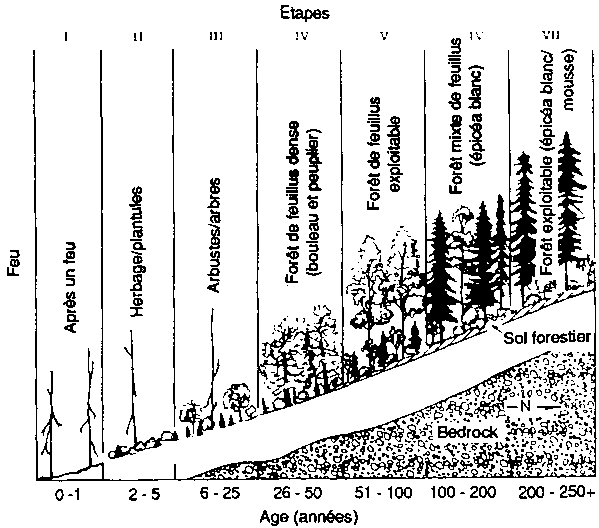
Figura D1 - Biomasse (tonnes/ha)
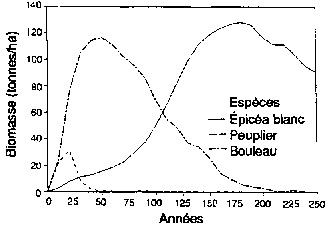
Figura D2 - Épaisseur (m)
Il existe un net besoin de continuer à développer les modèles informatiques capables de simuler la réaction des forêts aux changements des contraintes de l'environnement à l'échelle tant locale que mondiale. Des efforts sont déployés au niveau international pour construire en commun des modèles informatiques évolués. Ces modèles peuvent être utilisés pour prévoir les changements que subiront les forêts et faire la synthèse des nouvelles informations perspectives à mesure qu'elles apparaissent. Ils représentent un progrès important dans la recherche internationale en matière d'écologie forestière et dans la capacité de l'homme à prévoir comment les forêts s'adapteront au changement des conditions sur la terre.
Ainsi, depuis 1984, un effort international vise à développer un «modèle unifié de la forêt boréale», capable de prévoir le changement de dynamique face aux contraintes de l'environnement (y compris le changement climatique) pour les forêts boréales de l'Amérique du Nord et de l'Eurasie (Shugart Leemans et Bonan, 1992). La modélisation illustre la collaboration permanente de plus de 40 scientifiques appartenant à toutes les nations dotées de forêts boréales et à plusieurs pays ayant de grandes forêts alpines (par exemple, Suisse et Bulgarie). Une grande partie du modèle a été élaborée à partir des thèses de doctorat de G.B. Bonan (Université de Virginie) et de R. Leemans (Université d'Uppsala). Nous avons présenté des résultats de la version nord américaine du «modèle unifié de la zone boréale» dans la section précédente.
La mise au point d'un simulateur unifié pour biome forestier de première importance à l'échelle planétaire a plusieurs implications importantes. A l'origine, le modèle (Shugart, Leemans et Bonan, 1992) était conçu pour évaluer comment un des plus importants écosystèmes terrestres pouvait réagir en bloc au changement climatique mondial. La forêt boréale constitue un des plus grands réservoirs de carbone de la terre, particulièrement le carbone stocké dans le bois et les sols de la forêt. De plus, la modélisation représente une synthèse uniforme à travers une masse considérable de connaissances (l'ouvrage décrivant la formulation du modèle a une bibliographie de plus de 1400 titres). Le modèle est précieux pour ce qui est du développement des priorités de la recherche et du partage des connaissances sur les forêts boréales. Il est disponible gratuitement et a été conçu pour un usage très large s'appliquant à toute une gamme de problèmes forestiers. Les auteurs du simulateur boréal unifié souhaitent que le modèle actuel soit amélioré parallèlement au développement des études comparatives sur les différentes parties des forêts boréales.
Ågren G.I., McMurtrie, R.E., Parton, W.J., Pastor, J. & Shugart, H.H. 1991. State-of-the-art models of production-decomposition in conifer and grassland ecosystems. Ecol. Appl., 1:118-138
Bolin, B., Dons, B.R., Jager, J. & Warrick, R.A. eds. 1986. The greenhouse effect. climatic change and ecosystems, p. 207-270. Chichester. John Wiley.
Bonan, G.B. 1989. Environmental factors and ecological processes controlling vegetation patterns in boreal forests. Landscape Ecol., 3: 111-130.
Bonan, G.B. & Korzukin, M.D. 1989. Simulation of moss and tree dynamics in the boreal forest of interior Alaska. Vegetation, 84: 31-44.
Bonan, G.B. & Shugart, H.H. 1989. Environmental factors and ecological processes in boreal forests. Annu. Rev. Ecol. Syst., 20: 1-28.
Bonan, G.B., Shugart, H.H. & Urban, D.L. 1990. The sensitivity of some high-latitude boreal forests to climatic parameters. Climatic Change, 16: 9-29.
Fries, J., ed. 1974. Growth models for tree and stand simulation. Res. notes 30. Stockholm. Royal College of Forestry.
Holdridge, L.R. 1967. Life zone ecology. San Jose, Costa Rica Tropical Science Center.
Huston, M., DeAngelis, D.& Post, W.M. 1988. New computer models unify ecological theory. Bioscience. 38(10): 682-691.
IPCC. 1989a. Land -use management: adaptive responses to climate change. Working Paper for the Resource Use and Management Subgroup of Working Group 3.
IPCC. 1989b. Biological diversity: adaptive responses to climate change. Working Paper for the Resource Use and Management Subgroup of Working Group 3.
Pastor, J. & Post, W.M. 1988. Response of northern forests to CO2-induced climate change. Nature, 334(6177): 55-58.
Shugart, H.H. 1984. A theory of forest dynamics: the ecological implications of forest succession models. New York, Springer-Verlag. 278 pp.
Shugart, H.H., Antonovsky, M.Ya., Jarvis, P.G. & Sandford, A.P. 1986. CO2, climatic change and forest ecosystems. In B. Bolin B.R. Doos, J. Jager & R.A. Warrick, eds. The greenhouse effect, climatic change and ecosystems. p. 475-521. Chichester, John Wiley.
Shugart, H.H., Leemans, R. & Bonan, G.B., eds. 1992. A systems analysis of the global boreal forest. Cambridge, Cambridge Univ. Press. 542 pp.
Shugart, H.H. & Prentice, I.C. 1992. Individual tree-based models of forest dynamics and their application in global change research. In H.H. Shugart R. Leemans & G.B. Bonan, eds. A systems analysis of the global boreal forest, p. 313-333. Cambridge, Cambridge Univ. Press.
Smith, T.M., Leemans, R. & Shugart, H.H. 1991. Sensitivity of terrestrial carbon storage to CO2-induced climate change: comparison of five scenarios based on general circulation models. Climatic Change, 17. (Sous presse)
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}