![]()
![]()
![]()
par
Gösta Eriksson, Gene Namkoong et James Roberds1
RESUME
On s'accorde généralement à penser que pour donner les meilleurs résultats, les programmes de conservation doivent tabler sur la variation génétique. Néanmoins, il y a d'importantes divergences d'opinions concernant les méthodes à utiliser pour la conservation génétique dans ces programmes. La source de cette controverse réside dans des hypothèses formulées sur les ressources génétiques et leur dynamique. On pense à tort que l'évolution des populations naturelles conduit à une adaptation maximale. Au contraire, la sélection, la dérive génétique et la migration des gènes sont des forces d'évolution qui interagissent de manière complexe, de sorte qu'une adaptation maximale ne peut jamais être obtenue. Les changements constants se produisant dans l'environnement obligent aussi la sélection naturelle à agir dans de multiples directions. En outre, la plupart des caractères ayant une importance sont contrôlés par un certain nombre de loci génétiques qui ont des allèles avec des effets interchangeables. C'est pourquoi les ressources génétiques d'aujourd'hui devraient être considérées comme le point de départ des activités de conservation génétique et non pas comme un objectif des programmes de conservation.
La conservation génétique devrait viser avant tout à créer des conditions qui renforceront l'évolution future des ressources génétiques. Les méthodes utilisées devraient assurer que les objectifs du programme soient réalisés, prenant en compte à la fois la dynamique et la structure génétique. Le Système d'amélioration en populations multiples, qui associe conservation et amélioration génétique constitue en ce sens la meilleure méthode de gestion des programmes de conservation.
INTRODUCTION
Il y a un large consensus sur la nécessité de conserver les espèces et les généticiens forestiers et les gens informés s'accordent à penser que la variation génétique est un élément nécessaire dans tout programme de conservation. La variation est nécessaire non seulement pour assurer l'adaptabilité actuelle des espèces mais aussi pour l'évolution continue de l'espèce. Toutefois, les désaccords sont importants quant aux méthodes de conservation des gènes. On s'interroge toujours, par exemple, sur la taille idéale des échantillons, les méthodes in situ par opposition aux méthodes ex situ et la régénération naturelle par opposition à la régénération artificielle. Dans le présent article, nous examinons la variation génétique dans une optique d'évolution et nous décrivons une stratégie de programme qui répond aux objectifs de conservation des gènes forestiers. Pour mettre au point un programme efficace, nous devons respecter les principes scientifiques de la génétique tout comme les principes moraux. Nous nous pencherons d'abord sur les principes avant de proposer quelques directives pour un programme de conservation.
RESSOURCES GENETIQUES
Un motif important de désaccord concernant les méthodes de conservation porte sur les hypothèses formulées sur la nature des ressources génétiques et leur dynamique. Une hypothèse porte sur le nombre de gènes qui influent sur un caractère. Si chaque caractère était contrôlé par un locus unique, comme on pourrait le croire, chaque allèle à chaque locus serait important et toute perte diminuerait la capacité d'une population de réagir aux pressions environnementales. De là la préférence pour de grandes collections et de grandes réserves pour conserver des allèles, et plus rare est un allèle, plus importante doit être la collection. Toutefois, la plupart des caractères importants sont contrôlés par de nombreux loci et chaque locus a des effets qui contrôlent l'expression d'un grand nombre de caractères. Ainsi, l'effet d'un allèle à un locus peut souvent être remplacé par n'importe lequel de plusieurs allèles à ce locus ou à d'autres loci qui agissent de la même manière ou selon plusieurs autres mécanismes métaboliques. On donnera un exemple simplifié pour illustrer ce type de contrôle génétique. Nous supposons que tous les allèles ayant un indice commun contribuent dans la même mesure à l'adaptabilité, indépendamment du locus. Des allèles avec différents indices, toutefois, contribuent de différentes manières à l'adaptabilité. En ce cas, le génotype A1A1B1B2C1C2 aura la même adaptabilité que A2A2B1B1C1C1, et il en sera de même pour tous les autres génotypes qui ont quatre allèles à indice 1. Pour parvenir à un produit final donné, il y a de nombreuses options et l'évolution ne dépend pas de gènes uniques.
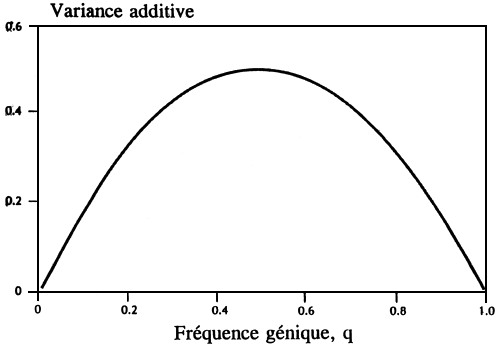
Les moyennes des caractères peuvent être déplacées en remplaçant les allèles qui ont des effets additifs et les plus grands déplacements sont dus à des allèles qui ne sont ni très communs ni très rares. La Figure 1 montre que des allèles à des fréquences inférieures à 0,01 ou supérieures à 0,99 ont peu d'effet sur la variance génétique additive et cela explique qu'il n'y a guère d'effet immédiat sur l'adaptabilité. Il n'est donc pas nécessaire de sauvegarder tous les allèles à tous les loci et, en fait, l'évolution au cours des prochaines générations ne dépendra pas d'allèles rares et leur sauvegarde n'est pas le problème immédiat. Certains se préoccupent à juste titre au sujet du rôle des allèles rares dans l'évolution à long terme, mais nous pouvons gérer la ressource en tenant compte de ce fait sans devoir créer des collections ou des réserves plus grandes pour conserver des allèles uniques.
Figure 1. Relation entre la variance additive et la fréquence génique à un seul locus avec deux allèles, l'action des gènes étant totalement additive.
Un autre motif de désaccord réside dans l'hypothèse que l'évolution dans la régénération naturelle maximalise toujours l'adaptabilité et que la meilleure stratégie de conservation consiste à intervenir le moins possible. Cependant, les résultats de la sélection reflètent les effets de conditions antérieures et, en outre, la sélection n'est pas la seule force d'évolution qui donne sa forme à la ressource génétique. Etant donné que les principaux effets de la sélection se font sentir au niveau de l'organisme individuel, les effets de la sélection sur les caractères spécifiques dépendent de la manière dont le caractère est lié à d'autres caractères qui constituent l'adaptabilité totale. Ainsi, les caractères sont parfois corrélés négativement, ce qui signifie que le progrès obtenu dans un caractère aura une incidence défavorable sur un autre caractère. La plasticité phénotypique permet également aux individus de survivre même s'ils ne sont pas génétiquement adaptés idéalement à un environnement particulier. De plus, un certain nombre de gènes et de caractères sans importance pour l'adaptabilité sont changés simplement sous l'effet d'une corrélation fortuite. Par ailleurs, des gènes individuels agissent habituellement sur plusieurs caractères de manière pléiotropique, si bien qu'il est rare que l'on puisse dire qu'un caractère est à un niveau optimal. Le changement constant de l'environnement, avec ou sans événements importants comme le changement climatique mondial, oblige la sélection naturelle à agir dans de multiples directions. Par conséquent, on n'obtiendra jamais une adaptabilité maximale ne serait-ce que pour cette raison.
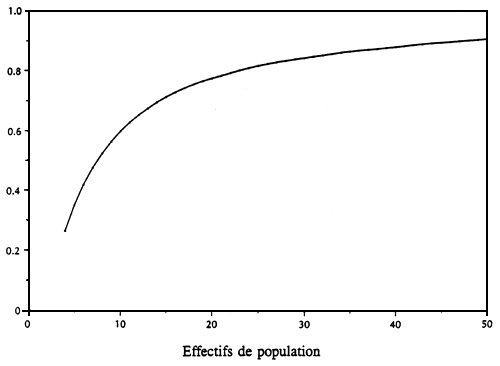
Parmi les autres forces d'évolution qui affectent la structure des ressources génétiques, citons la dérive génétique et sa contrepartie, la migration. Le petit effectif d'une population permet aux processus aléatoires d'influer fortement sur les distributions génétiques et de réduire la variation génétique au sein de ces populations. La Figure 2 montre qu'à peine plus de 50 % de la variance additive reste après 10 générations pour un effectif de population (Ne) = 8 et que plus de 25 % de la variance est perdue à Ne = 16. La dérive génétique est une grande force d'évolution qui peut annuler l'effet de la sélection naturelle dans les petites populations.
Figure 2. Fraction restante de la variance additive après dix générations pour différents effectifs de population. Aucune autre force d'évolution n'est supposée influer sur le résultat.
Fraction restante de la variance additive après dix générations
Pour comprendre les effets de la migration, il est utile d'examiner le concept de cercles environnementaux de sélection (Selective Environmental Neighbourhoods - SEN) introduit par Brandon (1990). Dans un SEN donné, les arbres ne changent pas de rang par rapport à l'adaptabilité. Là où il y a un flux de gènes entre plusieurs SEN, l'effet de la sélection naturelle est amoindri par rapport à ce qu'il aurait pu être a priori. Cela produit une forte variation dans l'adaptabilité au sein de chaque population à travers les SEN, mais entraîne aussi une adaptation réduite pour tout site donné. Ainsi les croisements entre différents SEN sont probablement à l'origine de la variation importante dans les caractères adaptatifs observés dans des peuplements de Pinus sylvestris et Picea abies (Eriksson 1982). La dérive génétique peut être marquée dans certains peuplements forestiers tropicaux, mais pour de nombreuses essences des forêts boréales et tempérées pollinisées par le vent, la migration est importante et va contrecarrer les effets de la dérive, du moins périodiquement.
En résumé, les forces d'évolution de la sélection, de la dérive et de la migration agissent de manière complexe et ne nous induisent pas à penser que la nature engendre la perfection. L'état actuel du système génétique peut être considéré comme un point de départ pour l'avenir, mais cela ne signifie pas qu'il est “sacré”, et son entretien sans changement ne doit pas être l'objectif de conservation. Néanmoins, il est clair que quels que soient les résultats de l'évolution, des espèces ont survécu jusqu'ici et représentent donc une solution viable à leurs problèmes passés. Mais l'on continue de se demander si elles sont optimales pour les conditions futures, notamment devant les changements survenant dans l'environnement (y compris un éventuel changement climatique mondial).
Pour les essences forestières, le désaccord sur les méthodes de conservation semble reposer, du moins en partie, sur des équivoques quant à la nature de la ressource génétique. Si la ressource pouvait être comprise comme un système dynamique, alors la structure de toute espèce serait mieux considérée comme l'une des nombreuses structures possibles et la structure présente, par sa nature même, comme un état passager. En tentant de geler l'état actuel, on irait à l'encontre de la dynamique naturelle du système et on ne répondrait pas aux objectifs de conservation génétique des arbres forestiers.
ASPECTS MORAUX
Etant donné qu'il y a de fortes contraintes en ce qui concerne le temps, le personnel et les fonds pour la conservation, nous devrons toujours faire des choix sur les génotypes, les populations ou les espèces dans lesquelles investir notre temps, nos ressources financières et nos talents. Une affirmation sur le plan moral est souvent énoncée sous la forme de ce que l'on appelle un “faux raisonnement naturaliste” selon lequel tout ce qui vient de la nature est bon par essence. Cela débouche sur deux principes; tout ce qui existe devrait être conservé et mieux vaut ne pas agir qu'agir n'importe comment. Ces principes nous obligent alors à ne pas faire de choix ou à conserver activement seulement par caprice. Aucun de ces résultats ne fournit de directives ou de priorités raisonnées en matière de conservation pouvant justifier un investissement public. En outre, l'affirmation donne une importance non méritée à ce qui en fait est un état biologiquement passager sans justifier ce choix. L'erreur consiste dans le fait de prétendre qu'un état particulier du monde est bon sans justifier cette affirmation autrement qu'en proclamant simplement sa vérité.
Pour justifier la conservation sur le plan moral, il faut expliquer en termes convaincants pourquoi, dans un monde de contraintes, des investissements devraient être faits en faveur des programmes de conservation. Il est très important de préciser pour le bien de qui nous conservons. La plupart reconnaîtront qu'il faut tenir compte du bien-être des êtres humains et que c'est surtout pour le bien au moins des générations actuelles et futures que la plus grande partie des activités de conservation sont entreprises. Cela vient de l'idée que les êtres humains ont des valeurs intrinsèques qui se différencient de la valeur qu'ont les outils qui leur servent. Le problème des conservationnistes est de savoir si des entités biologiques autres que les êtres humains ont une valeur intrinsèque et, si oui, comment incorporons-nous le bien intrinsèque de celles-ci dans les objectifs de conservation? Cette question a fait couler beaucoup d'encre, mais dans le cadre du présent article, on peut noter que même si nous n'attribuons pas à d'autres êtres des valeurs intrinsèques, il reste encore assez d'arguments en faveur de la conservation de presque toutes les espèces (Norton 1987). Un programme de conservation fondé uniquement sur la valeur intrinsèque des générations d'aujourd'hui et de demain n'excluerait pas la conservation d'autres espèces présentant un intérêt direct ou indirect pour l'homme mais pourrait en fait comprendre un large éventail d'espèces en raison de leur utilité pour les êtres humains, de leur utilité pour les espèces directement utiles à l'homme, etc. Fonder la conservation des espèces sur leur utilité directe ou indirecte pour l'homme évite aussi le problème d'essayer de justifier comment des groupes d'individus peuvent revendiquer la moindre valeur intrinsèque. Il semble qu'il n'y ait pas de propriétés de groupes comme des espèces considérées distinctes de leurs membres pris séparément qui donnent à ce groupe une importance particulière. Ainsi, la rareté est une propriété de groupe, mais elle n'implique pas en soi une valeur intrinsèque autant qu'elle implique que la gestion peut réussir à sauver des gènes en sauvant les quelques génotypes restants. Un autre argument encore plus valable en faveur de la conservation de grande échelle des espèces peut prendre comme point de départ leur valeur pour les humains et, dans ce cas, il est beaucoup plus facile de mettre en place des priorités. Il ne faut pas oublier que nous adoptons ici une vue large de ce qui constitue une valeur pour les générations actuelles et futures.
OBJECTIFS
Dans les sections précédentes et auparavant (Eriksson et al. 1993), nous avons affirmé que la nature est dynamique et cela implique que la conservation génétique devrait aussi être dynamique. Le potentiel pour une évolution dynamique est particulièrement important à une période où les changements dans l'environnement sont de taille. Il s'ensuit qu'un objectif important est la création de bonnes conditions pour l'évolution future.
Un objectif subordonné, mais important est de saisir des gènes à des fréquences > 0,01 et de tirer des adaptations existantes pouvant se présenter dans différentes populations. Ce dernier objectif pourrait être considéré comme contraire à ce qui a été énoncé plus haut. Il s'agit en fait d'augmenter la probabilité de saisir la variance additive la plus grande possible dans les populations de ressources génétiques. Cet objectif cadre parfaitement avec nos arguments précédents. Un troisième objectif consiste à incorporer une composante d'amélioration associée. C'est une fonction particulièrement importante pour les programmes de conservation comportant des espèces très utiles du point de vue commercial.
Les arbres ne sont pas les seuls organismes qui constituent un écosystème forestier. De nombreuses autres espèces dépendent de certaines essences forestières pour leur survie et leur bien-être. On les appelle espèces associées par opposition aux essences forestières (espèces cibles) qui sont l'axe autour duquel s'articule un programme de conservation forestière. Pour rester crédibles, il est important d'inclure la conservation des espèces associées parmi nos objectifs.
Certains conservationnistes plaident en faveur de la conservation de la structure actuelle des gènes, cela principalement parce que la structure actuelle sert de référence pour des comparaisons futures. D'autres prétendent que les populations qui sont menacées devraient avoir la priorité. Ces objectifs ont l'un et l'autre des avantages mais revêtent moins d'importance que notre premier objectif.
Comme il a été dit dans la section précédente, il ne suffit pas qu'un élément génétique soit rare pour justifier sa conservation pour des raisons morales.
CONNAISSANCES GENETIQUES
L'analyse de la structure des populations génétiques fournit l'information de base nécessaire pour mettre au point de bonnes méthodes d'échantillonnage pour les programmes de conservation des gènes. La structure peut varier en fonction du caractère pour lequel elle est déterminée. Pour ce qui est de la structure de la population, les caractères peuvent être regroupés en deux grandes classes: marqueurs moléculaires et caractères mesurables. On sait maintenant que la première classe de caractères affiche une variation limitée entre populations dans de nombreuses espèces, alors que les caractères mesurables affichent une variation considérable aussi bien entre populations qu'à l'intérieur d'une même population dans un grand nombre d'espèces. Cela est une conséquence de l'évolution antérieure des deux types de caractère. Les marqueurs moléculaires sont neutres ou presque et reflètent l'évolution à long terme alors que bon nombre des caractères mesurables sont associés à l'adaptation et en conséquence reflètent des faits récents relatifs à la sélection. Avant de mettre en oeuvre un programme d'échantillonnage pour la conservation, il est important d'étudier les structures des populations pour les deux types de caractères. Il faudrait éviter l'échantillonnage fondé exclusivement sur les données relatives aux marqueurs moléculaires.
Pour la majorité des essences forestières, nous n'aurons jamais les connaissances suffisantes sur la structure génétique tant pour les marqueurs moléculaires que pour les caractères mesurables. La rareté de l'information ne saurait justifier l'inaction. Les connaissances disponibles ne permettant pas de jeter les bases d'un programme de conservation, il faut partir d'hypothèses fondées sur la connaissance des structures possibles des populations. Ces hypothèses peuvent s'appuyer sur les caractéristiques écologiques de l'espèce ainsi que sur son habitat naturel.
La recherche concernant la structure d'une population devrait être axée sur des espèces ayant différentes combinaisons de caractères écologiques. Ces espèces peuvent servir de modèles pour les programmes de conservation d'autres espèces. Quand on disposera de données sur un nombre suffisant d'espèces, on pourra élaborer des directives applicables à des espèces dont la structure des populations est inconnue.
METHODES
La création de bonnes conditions pour l'évolution future a été identifiée plus haut comme l'objectif principal pour la conservation génétique. La méthode de conservation choisie pour une espèce devrait ainsi garantir son potentiel d'évolution. Le moyen le plus facile d'y parvenir est de mettre au point une collection de métapopulations à l'intérieur d'une même espèce. La conservation ne devrait pas être limitée à la préservation de la variance additive existante mais devrait encourager l'accroissement de ce type de variance chaque fois que possible. Le Système d'amélioration en populations multiples (Multiple Population Breeding System - MPBS) mis au point par Namkoong (1976, 1984) fournit une méthode pour réaliser cet objectif. Suivant ce système, une population combinée d'amélioration et de ressources génétiques est divisée en une vingtaine de sous-populations comptant chacune à peu près 50 entités génétiques. Les avantages de diviser la population en sous-populations sont résumés ci-après:
Dans ce contexte, la conservation génétique devient plus qu'une simple préservation, elle se transforme en un processus dynamique qui peut inclure l'amélioration des arbres à long terme, et en tirer parti. Dans ce processus, la variance entre populations est accrue alors que la variance à l'intérieur d'une même population est maintenue. Pour un effectif de population de 50, le taux de perte de variance additive est de 1 % par génération. Avec une gestion judicieuse, une partie de cette perte peut être neutralisée par la sélection. La fréquence de mutation dans des loci contrôlant des caractères mesurables a été estimée à 10-2 – 10-3 par génération dans certaines espèces (Lande et Burrowclough 1987). Ainsi, les mutations dans les caractères adaptatifs devraient compenser une partie de la perte de variance additive due à la dérive génétique.
Sous sa forme la plus élaborée, le MPBS exige la création de sous-populations dans un large éventail de conditions environnementales. Les plantations hors de l'aire naturelle actuelle de l'espèce peuvent aussi être utiles dans la préparation d'avenirs incertains. Cette forme de MPBS sera utilisée de préférence pour des essences forestières d'intérêt commercial en raison des coûts élevés que cela comporte. Toutefois, les espèces qui ont une grande importance pour l'existence de nombreuses autres espèces sont aussi candidates à cette forme de MPBS. Les avantages du MPBS peuvent aussi être obtenus avec la conservation génétique in situ d'espèces commercialement moins importantes à condition que l'échantillonnage des populations soit effectué de manière à maximiser l'adaptabilité existante. Selon les conditions économiques, la gestion active peut être prescrite de manière à ce que les individus désirés soient sélectionnés pour servir de semenciers pour la prochaine génération.
La recherche a montré que de nombreuses plantes menacées dépendent particulièrement des caractéristiques de l'habitat. Pour atteindre l'objectif de la conservation génétique pour des espèces associées, la diversité de l'habitat devrait être un objectif. L'expansion de certaines sous-populations dans la variante in situ du MPBS pour inclure quelques centaines d'hectares, conjuguée à un plan de gestion visant à créer une diversité maximale de l'habitat à l'intérieur de ces vastes sous-populations devraient assurer la conservation génétique pour une majorité d'espèces associées (voir Varela et Eriksson 1995). Cette pratique toutefois risque de ne pas être efficace pour certains animaux de grande taille.
Pour conclure, le MPBS est la méthode la plus efficace de conservation génétique des arbres forestiers mais elle pâtit du coût élevé de la plantation. Des variantes in situ du MPBS à employer avec des espèces jugées actuellement moins importantes peuvent aussi être conçues pour aider à réaliser l'objectif principal de la conservation génétique des arbres forestiers: créer de bonnes conditions pour l'évolution future.
REFERENCES
Brandon (1990). Adaptation and Environment. Princeton Univ. Press, Princeton, New Jersey. 214pp.
Eriksson, G. (1982). Ecological genetics of conifers in Sweden. Silva Fenn. 16: 149–156.
Eriksson, G.; Namkoong, G. & Roberds, J. (1993). Dynamic gene conservation for uncertain futures. For. Ecol. Manag. 62: 15–37.
Lande, R. & Burrowclough, G.F. (1987). Effective population size, genetic variation, and their use in population management. In Viable Populations for Conservation (Ed. M.E. Soulé), New York, Cambridge Univ. Press: 87–123.
Namkoong, G. (1976). A multiple index selection strategy. Silvae Genet. 25:199–201.
Namkoong, G. (1984). Strategies for gene conservation in forest tree breeding. In Plant Gene Resources: A Conservation Imperative. (Eds. C.W. Yeatman, D. Kafton, and G. Wilkes). AAAS Selected Symposium 87. Westview Press, Boulder CO.: 79–89.
Norton, B.C. (1987). Formal and substantive priority systems. In Why preserve Natural Variety, (Ed. M. Cohen). Princeton Univ. Press, Princeton, New Jersey: 243–257.
Varela, M.C. and Eriksson, G. (1995). Multipurpose gene conservation in Quercus suber - A Portuguese example. Silvae Genet. Sous presse.
Resources Génétiques Forestières No 23. FAO, Rome (1996)
Manuscrit reçu en mai 1995
![]()
![]()
![]()